
Extracts from the Leaves of Cissus verticillata Ameliorate High-Fat Diet-Induced Memory Deficits in Mice

 , , and
, , and 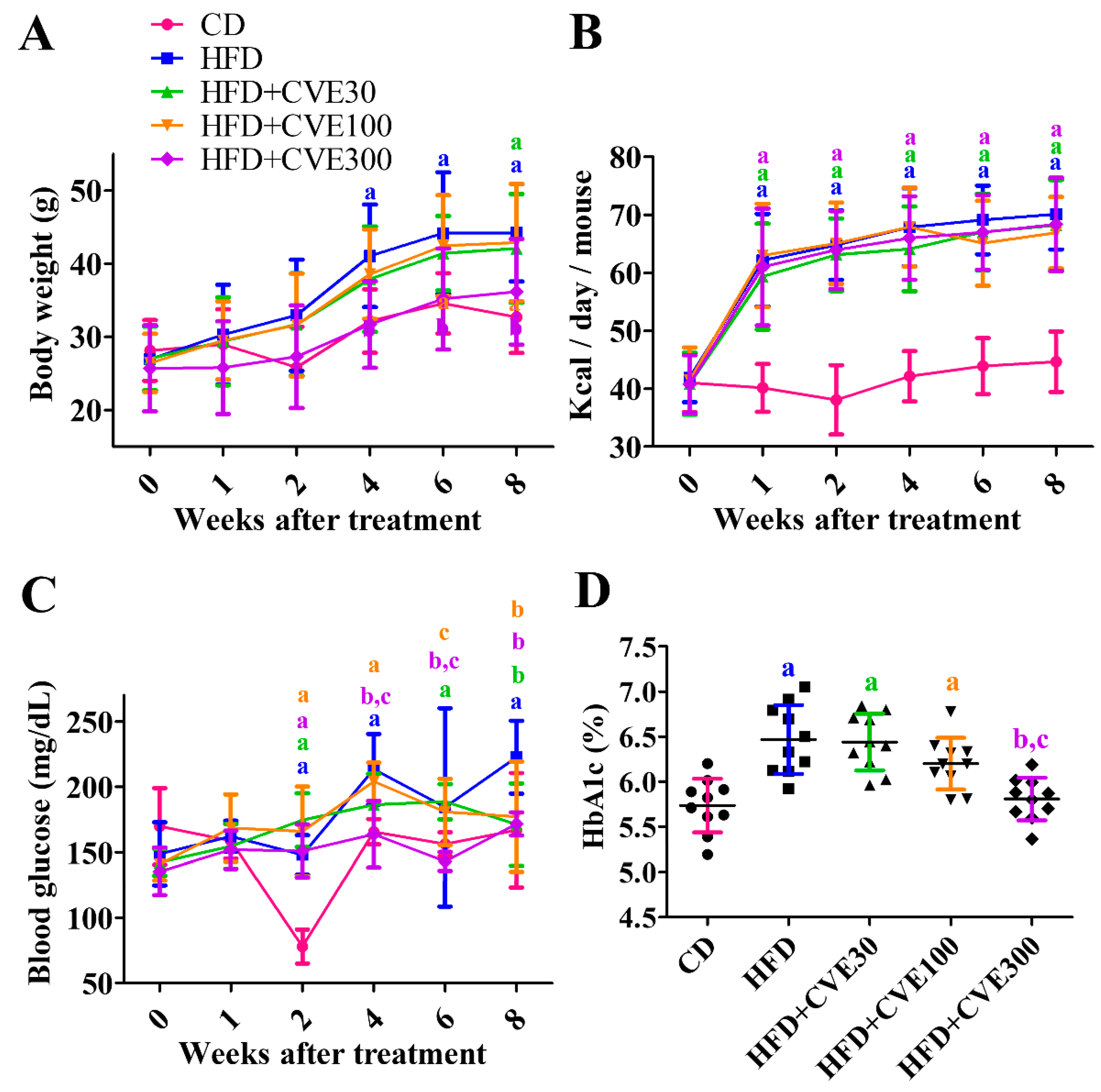{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of CVE on Body Weight, Blood Glucose, and HbA1c Levels
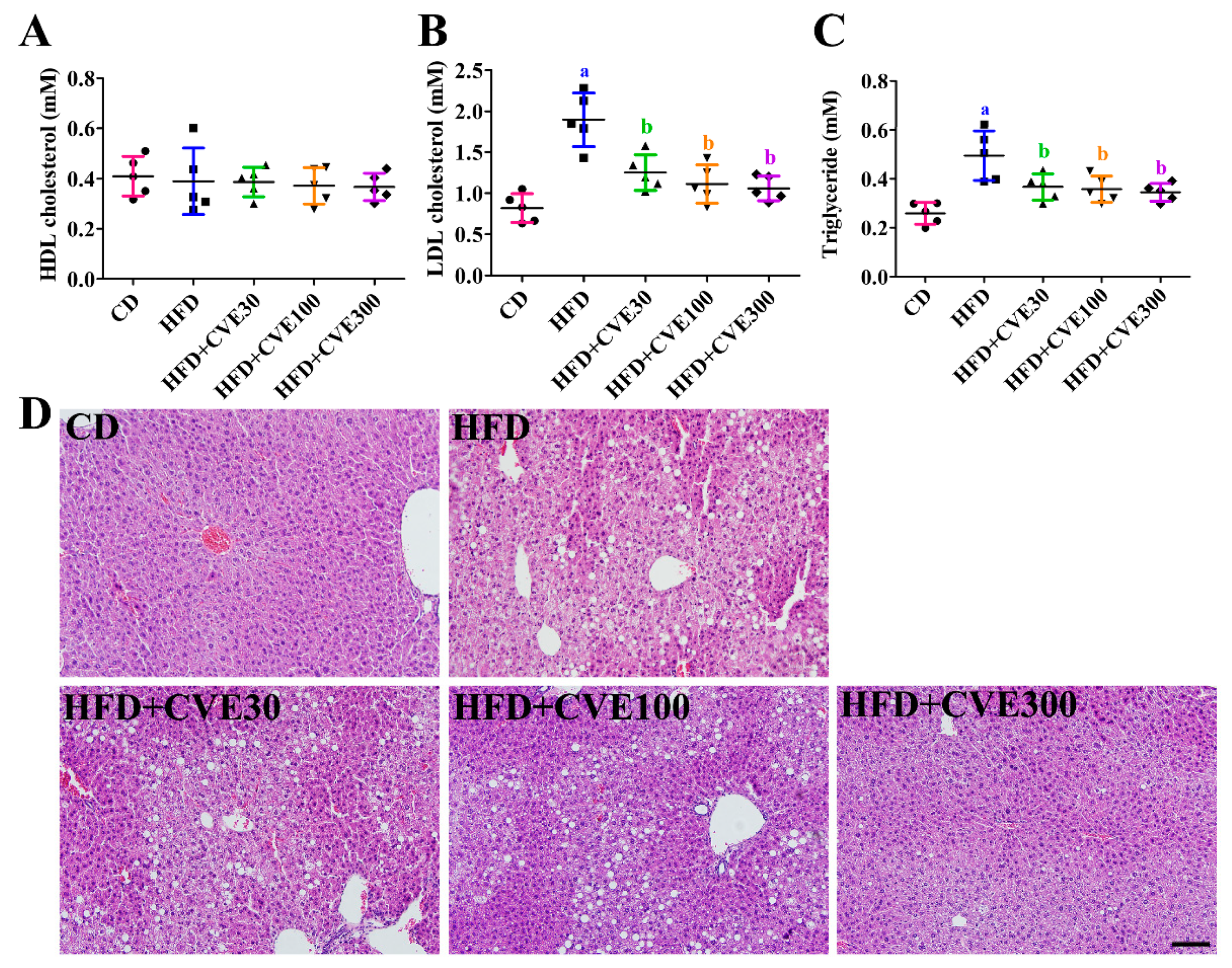
2.2. Effect of CVE on Biochemical Parameters in the Serum
2.3. Effect of CVE on Liver Morphology
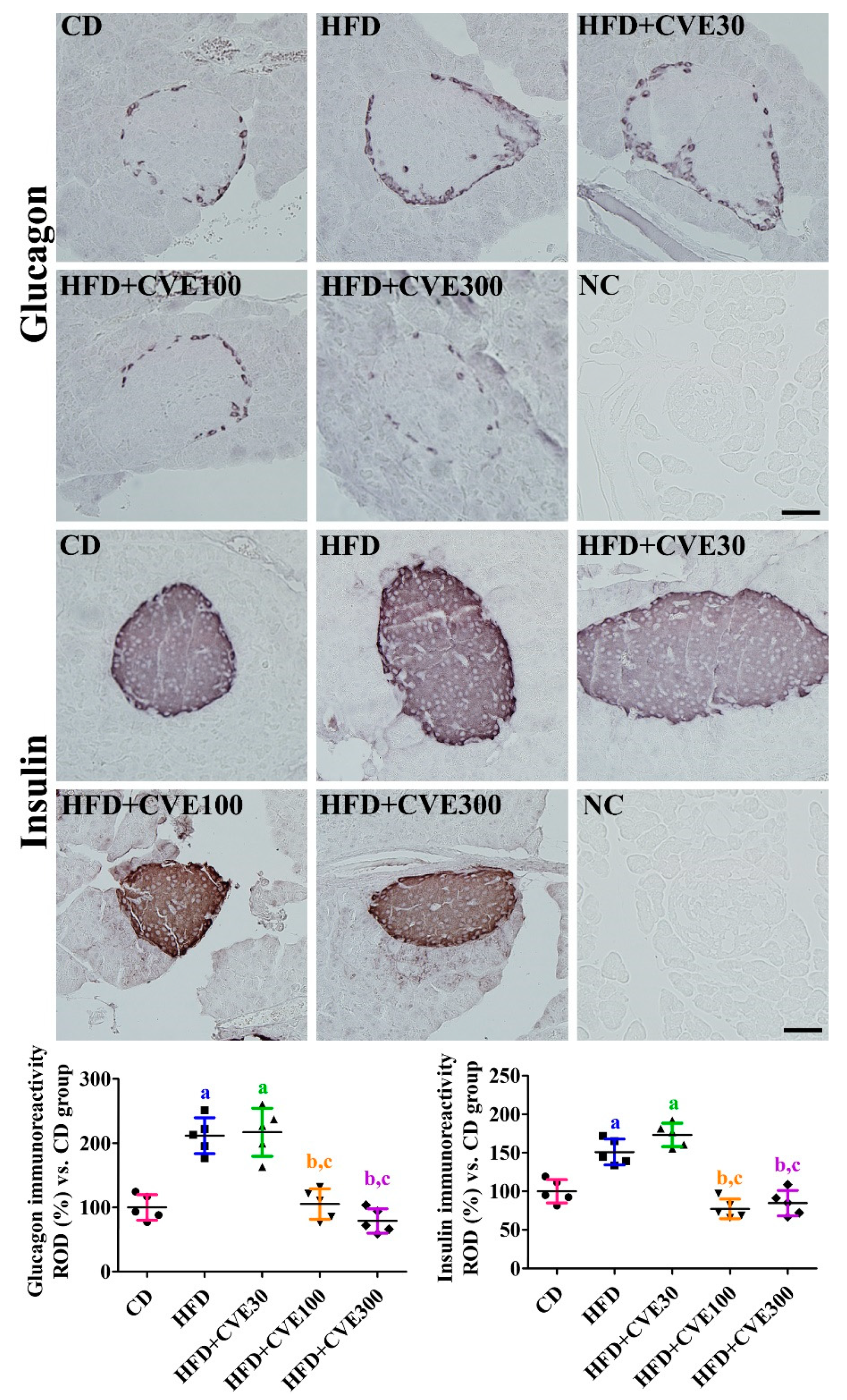
2.4. Effect of CVE on Insulin and Glucagon Secreting Cells in the Pancreas
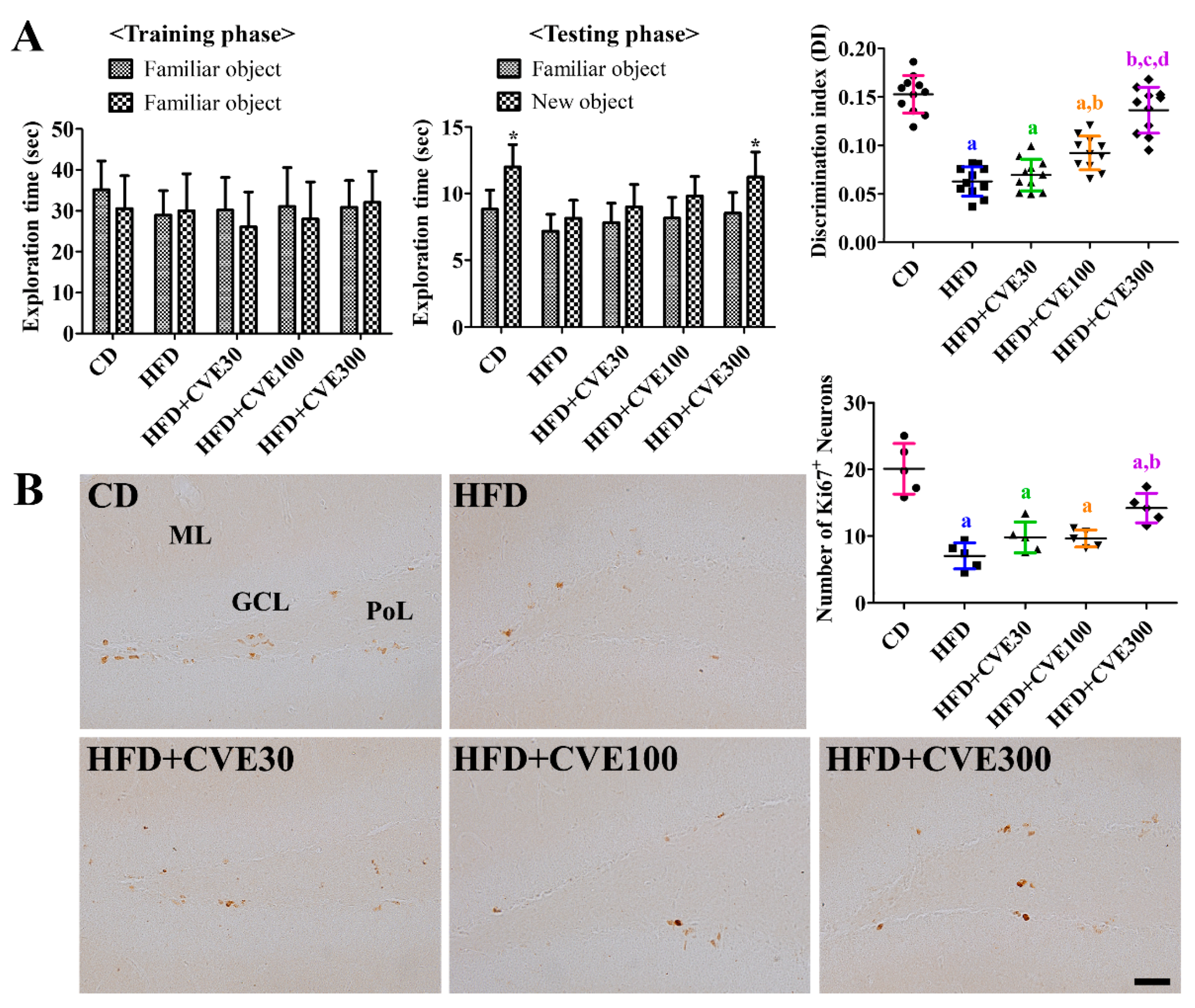
2.5. Effect of CVE on Novel Object Recognition Memory
2.6. Effect of CVE on Proliferating Cells and Differentiated Neuroblasts in the Dentate Gyrus
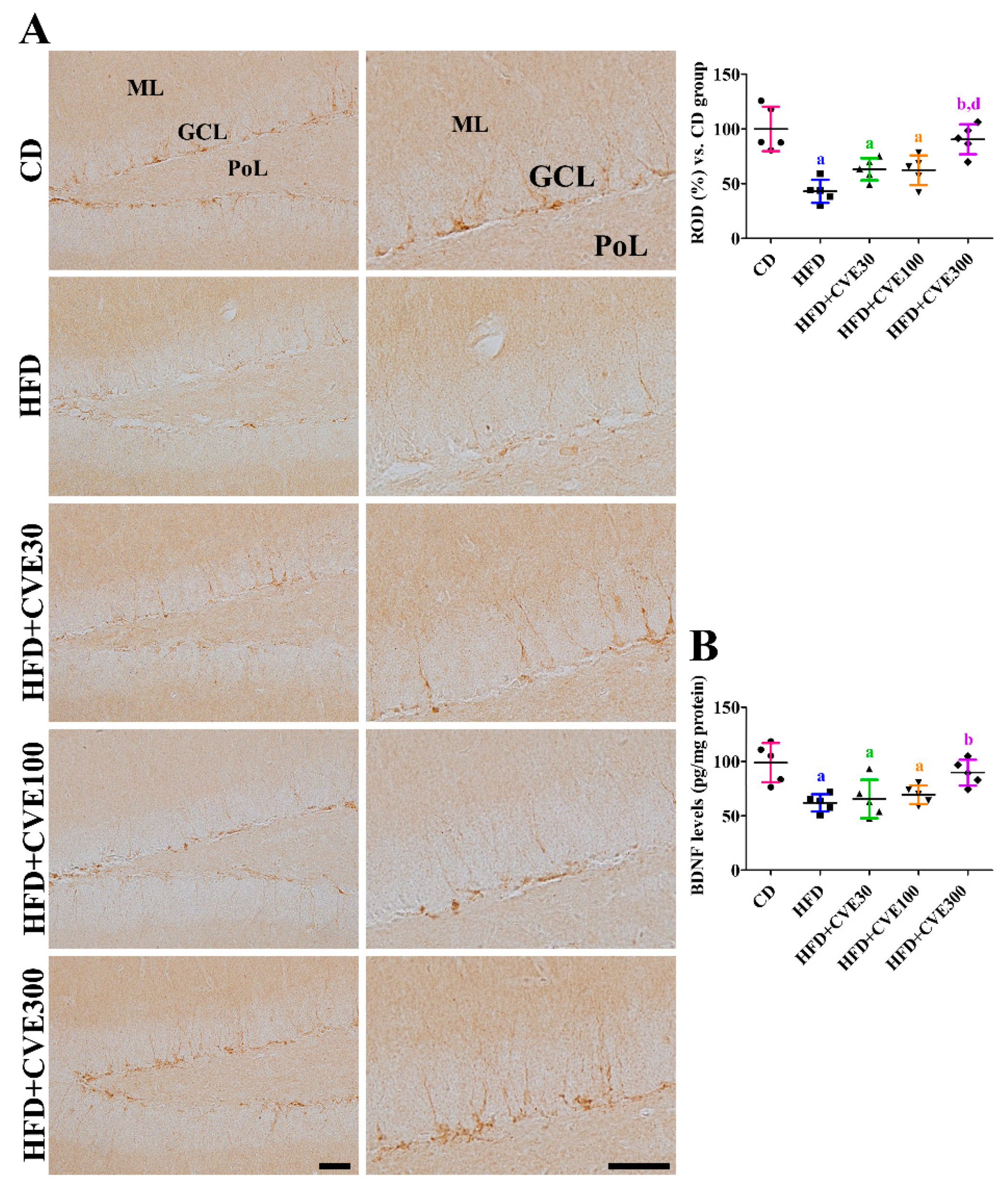
2.7. Effect of CVE on Brain-Derived Neurotrophic Factor (BDNF) Levels in the Hippocampus
3. Discussion
4. Materials and Methods
4.1. Preparation of CVE
4.2. Experimental Animals
4.3. Experimental Groups
4.4. Novel Object Recognition Memory
4.5. Biochemical Experiments and Morphological Study of Peripheral Tissues
4.6. Morphological Study in the Hippocampus
4.7. Data Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tchkonia, T.; Morbeck, D.E.; Von Zglinicki, T.; Van Deursen, J.; Lustgarten, J.; Scrable, H.; Khosla, S.; Jensen, M.D.; Kirkland, J.L. Fat tissue, aging, and cellular senescence. Aging Cell 2010, 9, 667–684. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, D.; Strakovsky, R.; Zhang, Y.; Pan, Y.X. Hepatic cellular senescence pathway genes are induced through histone modifications in a diet-induced obese rat model. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G558–G564. [Google Scholar] [CrossRef]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic adipose tissue inflammation linking obesity to insulin resistance and type 2 diabetes. Front. Physiol. 2020, 10, 1607. [Google Scholar] [CrossRef] [PubMed]
- Ye, J. Mechanisms of insulin resistance in obesity. Front. Med. 2013, 7, 14–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabia, S.; Kivimaki, M.; Shipley, M.J.; Marmot, M.G.; Singh-Manoux, A. Body mass index over the adult life course and cognition in late midlife: The whitehall II cohort study. Am. J. Clin. Nutr. 2009, 89, 601–607. [Google Scholar] [CrossRef] [Green Version]
- Arnold, S.E.; Lucki, I.; Brookshire, B.R.; Carlson, G.C.; Browne, C.A.; Kazi, H.; Bang, S.; Choi, B.R.; Chen, Y.; McMullen, M.F.; et al. High fat diet produces brain insulin resistance, synaptodendritic abnormalities and altered behavior in mice. Neurobiol. Dis. 2014, 67, 79–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lizarbe, B.; Soares, A.F.; Larsson, S.; Duarte, J.M.N. Neurochemical modifications in the hippocampus, cortex and hypothalamus of mice exposed to long-term high-fat diet. Front. Neurosci. 2019, 12, 985. [Google Scholar] [CrossRef] [PubMed]
- Kamal, A.; Ramakers, G.M.; Gispen, W.H.; Biessels, G.J. Hyperinsulinemia in rats causes impairment of spatial memory and learning with defects in hippocampal synaptic plasticity by involvement of postsynaptic mechanisms. Exp. Brain Res. 2013, 226, 45–51. [Google Scholar] [CrossRef]
- Duarte, J.M.N.; Skoug, C.; Silva, H.B.; Carvalho, R.A.; Gruetter, R.; Cunha, R.A. Impact of caffeine consumption on type 2 diabetes-induced spatial memory impairment and neurochemical alterations in the hippocampus. Front. Neurosci. 2019, 12, 1015. [Google Scholar] [CrossRef] [PubMed]
- Kerti, L.; Witte, A.V.; Winkler, A.; Grittner, U.; Rujescu, D.; Flöel, A. Higher glucose levels associated with lower memory and reduced hippocampal microstructure. Neurology 2013, 81, 1746–1752. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Chang, G.Q.; Dourmashkin, J.T.; Leibowitz, S.F. Distinct phenotypes of obesity-prone AKR/J, DBA2J and C57BL/6J mice compared to control strains. Int. J. Obes. 2006, 30, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.K.; Kim, I.Y.; Kim, D.W.; Yoo, K.Y.; Kim, Y.N.; Yi, S.S.; Won, M.H.; Lee, I.S.; Yoon, Y.S.; Seong, J.K. Strain-specific differences in cell proliferation and differentiation in the dentate gyrus of C57BL/6N and C3H/HeN mice fed a high fat diet. Brain Res. 2008, 1241, 1–6. [Google Scholar] [CrossRef]
- Shao, X.; Wang, M.; Wei, X.; Deng, S.; Fu, N.; Peng, Q.; Jiang, Y.; Ye, L.; Xie, J.; Lin, Y. Peroxisome proliferator-activated receptor-γ: Master regulator of adipogenesis and obesity. Curr. Stem Cell Res. Ther. 2016, 11, 282–289. [Google Scholar] [CrossRef]
- Gao, F.; Zang, L.; Wu, D.Y.; Li, Y.J.; Zhang, Q.; Wang, H.B.; Tian, G.L.; Mu, Y.M. Pioglitazone improves the ability of learning and memory via activating ERK1/2 signaling pathway in the hippocampus of T2DM rats. Neurosci. Lett. 2017, 651, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Pipatpiboon, N.; Pratchayasakul, W.; Chattipakorn, N.; Chattipakorn, S.C. PPARγ agonist improves neuronal insulin receptor function in hippocampus and brain mitochondria function in rats with insulin resistance induced by long term high-fat diets. Endocrinology 2012, 153, 329–338. [Google Scholar] [CrossRef]
- Yoo, D.Y.; Kim, W.; Kim, D.W.; Nam, S.M.; Jung, H.Y.; Kim, J.W.; Lee, C.H.; Choi, J.H.; Won, M.H.; Yoon, Y.S.; et al. Cell proliferation and neuroblast differentiation in the dentate gyrus of high-fat diet-fed mice are increased after rosiglitazone treatment. J. Vet. Sci. 2014, 15, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Vickers, A.; Zollman, C.; Lee, R. Herbal medicine. West J. Med. 2001, 175, 125–128. [Google Scholar] [CrossRef]
- Charen, E.; Harbord, N. Toxicity of herbs, vitamins, and supplements. Adv. Chronic Kidney Dis. 2020, 27, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, J.A. Systematics of Vitaceae in South America. Can. J. Bot. 2007, 85, 712–721. [Google Scholar] [CrossRef]
- Beserra, F.P.; Santos Rde, C.; Périco, L.L.; Rodrigues, V.P.; Kiguti, L.R.; Saldanha, L.L.; Pupo, A.S.; da Rocha, L.R.; Dokkedal, A.L.; Vilegas, W.; et al. Cissus sicyoides: Pharmacological mechanisms involved in the anti-inflammatory and antidiarrheal activities. Int. J. Mol. Sci. 2016, 17, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salem, P.P.O.; Vieira, N.B.; Garcia, D.A.; Nicácio, K.J.; Dias, D.F.; de Paula, A.C.C.; Assis, D.M.; Caldas, I.S.; Novaes, R.D.; Marinho, M.V.; et al. Anti-urolithiatic and anti-inflammatory activities through a different mechanism of actions of Cissus gongylodes corroborated its ethnopharmacological historic. J. Ethnopharmacol. 2020, 253, 112655. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Xu, C.T.; Lin, D.D.; Qin, J.K.; Ye, G.J.; Deng, Q.H. Anti-inflammatory polyphenol constituents derived from Cissus pteroclada Hayata. Bioorg. Med. Chem. Lett. 2016, 26, 3425–3428. [Google Scholar] [CrossRef]
- Dutta, T.; Paul, A.; Majumder, M.; Sultan, R.A.; Emran, T.B. Pharmacological evidence for the use of Cissus assamica as a medicinal plant in the management of pain and pyrexia. Biochem. Biophys. Rep. 2019, 21, 100715. [Google Scholar] [CrossRef] [PubMed]
- Mahamad, A.T.; Miaffo, D.; Poualeu Kamani, S.L.; Mahamat, O.; Kamanyi, A.; Wansi Ngnokam, S.L. Glucose, lipid and oxidative stress lowering activity of the aqueous extract from leafy stems of Cissus polyantha Gilg & Brandt in dexamethasone-induced hyperglycemia in rats. J. Diabetes Metab. Disord. 2020, 19, 1527–1535. [Google Scholar] [PubMed]
- Pepato, M.T.; Baviera, A.M.; Vendramini, R.C.; Perez Mda, P.; Kettelhut Ido, C.; Brunetti, I.L. Cissus sicyoides (princess vine) in the long-term treatment of streptozotocin-diabetic rats. Biotechnol. Appl. Biochem. 2003, 37, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Viana, G.S.; Medeiros, A.C.; Lacerda, A.M.; Leal, L.K.; Vale, T.G.; Matos, F.J. Hypoglycemic and anti-lipemic effects of the aqueous extract from Cissus sicyoides. BMC Pharmacol. 2004, 4, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oben, J.; Kuate, D.; Agbor, G.; Momo, C.; Talla, X. The use of a Cissus quadrangularis formulation in the management of weight loss and metabolic syndrome. Lipids Health Dis. 2006, 5, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatree, S.; Sitticharoon, C.; Maikaew, P.; Pongwattanapakin, K.; Keadkraichaiwat, I.; Churintaraphan, M.; Sripong, C.; Sririwichitchai, R.; Tapechum, S. Cissus Quadrangularis enhances UCP1 mRNA, indicative of white adipocyte browning and decreases central obesity in humans in a randomized trial. Sci. Rep. 2021, 11, 2008. [Google Scholar] [CrossRef]
- Panthong, A.; Supraditaporn, W.; Kanjanapothi, D.; Taesotikul, T.; Reutrakul, V. Analgesic, anti-inflammatory and venotonic effects of Cissus quadrangularis Linn. J. Ethnopharmacol. 2007, 110, 264–270. [Google Scholar] [CrossRef]
- de Almeida, E.R.; Rafael, K.R.; Couto, G.B.; Ishigami, A.B. Anxiolytic and anticonvulsant effects on mice of flavonoids, linalool, and alpha-tocopherol presents in the extract of leaves of Cissus sicyoides L. (Vitaceae). J. Biomed. Biotechnol. 2009, 2009, 274740. [Google Scholar] [CrossRef] [Green Version]
- Moto, F.C.O.; Arsa’a, A.; Ngoupaye, G.T.; Taiwe, G.S.; Njapdounke, J.S.K.; Kandeda, A.K.; Nkantchoua, G.C.N.; Omam Omam, J.P.; Pale, S.; Kouemou, N.E.; et al. Anxiolytic and antiepileptic properties of the aqueous extract of Cissus quadrangularis (Vitaceae) in mice pilocarpine model of epilepsy. Front. Pharmacol. 2018, 9, 751. [Google Scholar] [CrossRef]
- Rao Sirasanagandla, S.R.; Ranganath Pai Karkala, S.; Potu, B.K.; Bhat, K.M. Beneficial Effect of Cissus quadrangularis Linn. on osteopenia associated with streptozotocin-induced type 1 diabetes mellitus in male Wistar rats. Adv. Pharmacol. Sci. 2014, 2014, 483051. [Google Scholar]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef]
- Willebrords, J.; Pereira, I.V.; Maes, M.; Crespo Yanguas, S.; Colle, I.; Van Den Bossche, B.; Da Silva, T.C.; de Oliveira, C.P.; Andraus, W.; Alves, V.A.; et al. Strategies, models and biomarkers in experimental non-alcoholic fatty liver disease research. Prog. Lipid Res. 2015, 59, 106–125. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.S.; Uddin, M.G.; Shoibe, M.; Mahmud, A.A.; Banik, S. Evaluation of anxiolytic and hypoglycemic potential of Cissus adnata Roxb. in animal model. J. Complement. Integr. Med. 2019, 17. [Google Scholar] [CrossRef]
- American Diabetes Association. 2. Classification and diagnosis of diabetes. Diabetes Care 2017, 40 (Suppl. 1), S11–S24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roden, M. Diabetes mellitus: Definition, classification and diagnosis. Wien. Klin. Wochenschr. 2016, 28 (Suppl. 2), S37–S40. [Google Scholar] [CrossRef] [PubMed]
- Kappe, C.; Zhang, Q.; Nyström, T.; Sjöholm, A. Effects of high-fat diet and the anti-diabetic drug metformin on circulating GLP-1 and the relative number of intestinal L-cells. Diabetol. Metab. Syndr. 2014, 6, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, C.L.; Lin, H.L.; Ke, L.Y.; Yen, H.W.; Shen, K.P. Pre-germinated brown rice extract ameliorates high-fat diet-induced metabolic syndrome. J. Food Biochem. 2019, 43, e12769. [Google Scholar] [CrossRef]
- Zeng, Q.; Zhao, L.; Meng, C.; Zhao, X.; Liu, Y.; Shi, R.; Han, X.; Wang, T.; Li, J. Prophylactic and therapeutic effects of different doses of vitamin C on high-fat-diet-induced non-alcoholic fatty liver disease in mice. Biomed. Pharmacother. 2020, 131, 110792. [Google Scholar] [CrossRef] [PubMed]
- Chidambaram, J.; Carani Venkatraman, A. Cissus quadrangularis stem alleviates insulin resistance, oxidative injury and fatty liver disease in rats fed high fat plus fructose diet. Food Chem. Toxicol. 2010, 48, 2021–2029. [Google Scholar] [CrossRef]
- Lekshmi, R.K.; Rajesh, R.; Mini, S. Ethyl acetate fraction of Cissus quadrangularis stem ameliorates hyperglycaemia-mediated oxidative stress and suppresses inflammatory response in nicotinamide/streptozotocin induced type 2 diabetic rats. Phytomedicine 2015, 22, 952–960. [Google Scholar] [CrossRef] [PubMed]
- Nagai, A.; Mizushige, T.; Matsumura, S.; Inoue, K.; Ohinata, K. Orally administered milk-derived tripeptide improved cognitive decline in mice fed a high-fat diet. FASEB J. 2019, 33, 14095–14102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada-Goto, N.; Katsuura, G.; Ochi, Y.; Ebihara, K.; Kusakabe, T.; Hosoda, K.; Nakao, K. Impairment of fear-conditioning responses and changes of brain neurotrophic factors in diet-induced obese mice. J. Neuroendocrinol. 2012, 24, 1120–1125. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.; Angelucci, A.; Costantin, L.; Braschi, C.; Mazzantini, M.; Babbini, F.; Fabbri, M.E.; Tessarollo, L.; Maffei, L.; Berardi, N.; et al. Brain-derived neurotrophic factor (BDNF) is required for the enhancement of hippocampal neurogenesis following environmental enrichment. Eur. J. Neurosci. 2006, 24, 1850–1856. [Google Scholar] [CrossRef]
- Franco-Robles, E.; López, M.G. Agavins increase neurotrophic factors and decrease oxidative stress in the brains of high-fat diet-induced obese mice. Molecules 2016, 21, 998. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fu, X.; Lan, N.; Li, S.; Zhang, J.; Wang, S.; Li, C.; Shang, Y.; Huang, T.; Zhang, L. Luteolin protects against high fat diet-induced cognitive deficits in obesity mice. Behav. Brain Res. 2014, 267, 178–188. [Google Scholar] [CrossRef]
- Abidin, İ.; Aydin-Abidin, S.; Bodur, A.; İnce, İ.; Alver, A. Brain-derived neurotropic factor (BDNF) heterozygous mice are more susceptible to synaptic protein loss in cerebral cortex during high fat diet. Arch. Physiol. Biochem. 2018, 124, 442–447. [Google Scholar] [CrossRef]
- Chidambara Murthy, K.N.; Vanitha, A.; Mahadeva Swamy, M.; Ravishankar, G.A. Antioxidant and antimicrobial activity of Cissus quadrangularis L. J. Med. Food 2003, 6, 99–105. [Google Scholar] [CrossRef]
- Yu, S.; Kim, S.R.; Jiang, K.; Ogrodnik, M.; Zhu, X.Y.; Ferguson, C.M.; Tchkonia, T.; Lerman, A.; Kirkland, J.L.; Lerman, L.O. Quercetin reverses cardiac systolic dysfunction in mice fed with a high-fat diet: Role of angiogenesis. Oxid. Med. Cell. Longev. 2021, 2021, 8875729. [Google Scholar] [CrossRef]
- Zhang, F.; Feng, J.; Zhang, J.; Kang, X.; Qian, D. Quercetin modulates AMPK/SIRT1/NF-κB signaling to inhibit inflammatory/oxidative stress responses in diabetic high fat diet-induced atherosclerosis in the rat carotid artery. Exp. Ther. Med. 2020, 20, 280. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Kwon, H.J.; Jung, H.Y.; Lim, S.S.; Kang, B.G.; Jo, Y.B.; Yu, D.S.; Choi, S.Y.; Hwang, I.K.; Kim, D.W. Cissus verticillata extract decreases neuronal damage induced by oxidative stress in HT22 cells and ischemia in gerbils by reducing the inflammation and phosphorylation of MAPKs. Plants 2021, 10, 1217. [Google Scholar] [CrossRef] [PubMed]
- Yoo, D.Y.; Jung, H.Y.; Kim, W.; Hahn, K.R.; Kwon, H.J.; Nam, S.M.; Chung, J.Y.; Yoon, Y.S.; Kim, D.W.; Hwang, I.K. Entacapone promotes hippocampal neurogenesis in mice. Neural Regen. Res. 2021, 16, 1005–1110. [Google Scholar] [PubMed]
- Yoo, D.Y.; Yim, H.S.; Jung, H.Y.; Nam, S.M.; Kim, J.W.; Choi, J.H.; Seong, J.K.; Yoon, Y.S.; Kim, D.W.; Hwang, I.K. Chronic type 2 diabetes reduces the integrity of the blood-brain barrier by reducing tight junction proteins in the hippocampus. J. Vet. Med. Sci. 2016, 78, 957–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewitt, S.M.; Baskin, D.G.; Frevert, C.W.; Stahl, W.L.; Rosa-Molinar, E. Controls for immunohistochemistry: The Histochemical Society’s standards of practice for validation of immunohistochemical assays. J. Histochem. Cytochem. 2014, 62, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Kim, W.; Hahn, K.R.; Jung, H.Y.; Kwon, H.J.; Nam, S.M.; Kim, J.W.; Park, J.H.; Yoo, D.Y.; Kim, D.W.; Won, M.H.; et al. Melatonin ameliorates cuprizone-induced reduction of hippocampal neurogenesis, brain-derived neurotrophic factor, and phosphorylation of cyclic AMP response element-binding protein in the mouse dentate gyrus. Brain Behav. 2019, 9, e01388. [Google Scholar] [CrossRef]
- Jung, H.Y.; Kim, W.; Kwon, H.J.; Yoo, D.Y.; Nam, S.M.; Hahn, K.R.; Yi, S.S.; Choi, J.H.; Kim, D.W.; Yoon, Y.S.; et al. Physical stress induced reduction of proliferating cells and differentiated neuroblasts is ameliorated by fermented Laminaria japonica extract treatment. Mar. Drugs 2020, 18, 587. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, W.; Kwon, H.J.; Jung, H.Y.; Lim, S.-S.; Kang, B.-G.; Jo, Y.-B.; Yu, D.-S.; Choi, S.Y.; Hwang, I.K.; Kim, D.W. Extracts from the Leaves of Cissus verticillata Ameliorate High-Fat Diet-Induced Memory Deficits in Mice. Plants 2021, 10, 1814. https://doi.org/10.3390/plants10091814
Kim W, Kwon HJ, Jung HY, Lim S-S, Kang B-G, Jo Y-B, Yu D-S, Choi SY, Hwang IK, Kim DW. Extracts from the Leaves of Cissus verticillata Ameliorate High-Fat Diet-Induced Memory Deficits in Mice. Plants. 2021; 10(9):1814. https://doi.org/10.3390/plants10091814
Chicago/Turabian StyleKim, Woosuk, Hyun Jung Kwon, Hyo Young Jung, Soon-Sung Lim, Beom-Goo Kang, Yong-Bok Jo, Dong-Sool Yu, Soo Young Choi, In Koo Hwang, and Dae Won Kim. 2021. "Extracts from the Leaves of Cissus verticillata Ameliorate High-Fat Diet-Induced Memory Deficits in Mice" Plants 10, no. 9: 1814. https://doi.org/10.3390/plants10091814
APA StyleKim, W., Kwon, H. J., Jung, H. Y., Lim, S. -S., Kang, B. -G., Jo, Y. -B., Yu, D. -S., Choi, S. Y., Hwang, I. K., & Kim, D. W. (2021). Extracts from the Leaves of Cissus verticillata Ameliorate High-Fat Diet-Induced Memory Deficits in Mice. Plants, 10(9), 1814. https://doi.org/10.3390/plants10091814