
When Water Availability Is Low, Two Mediterranean Salvia Species Rely on Root Hydraulics


Abstract
:1. Introduction
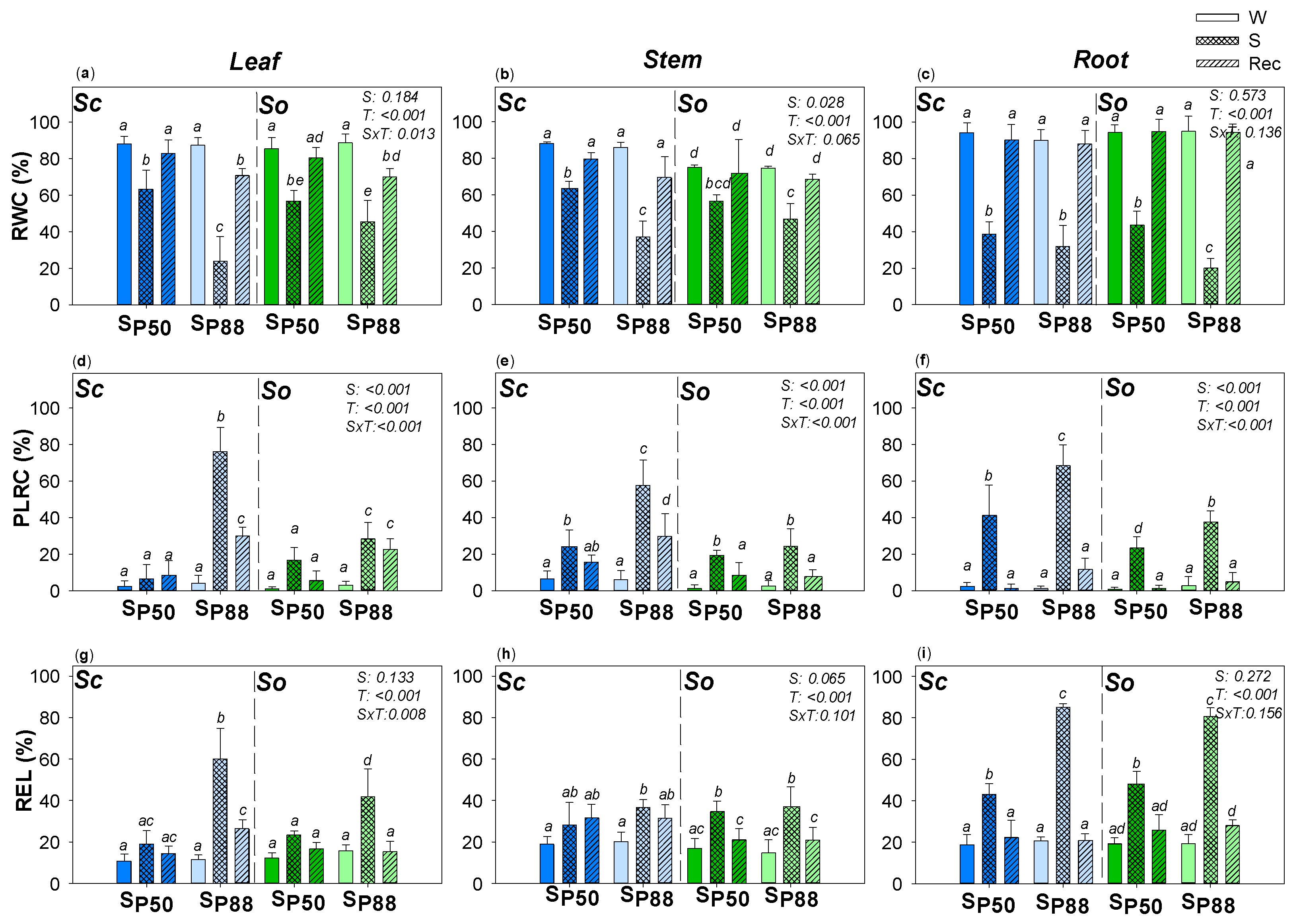
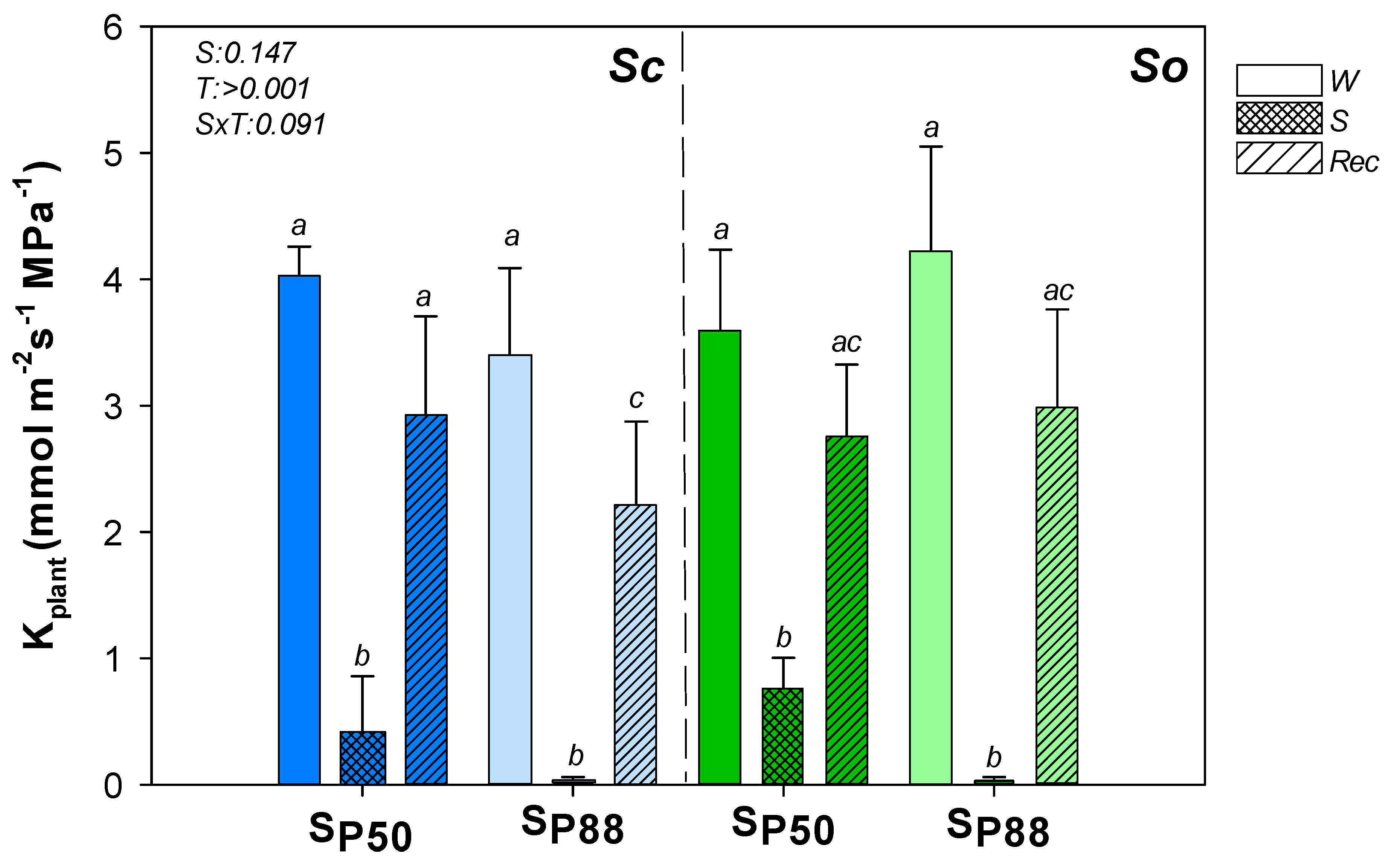
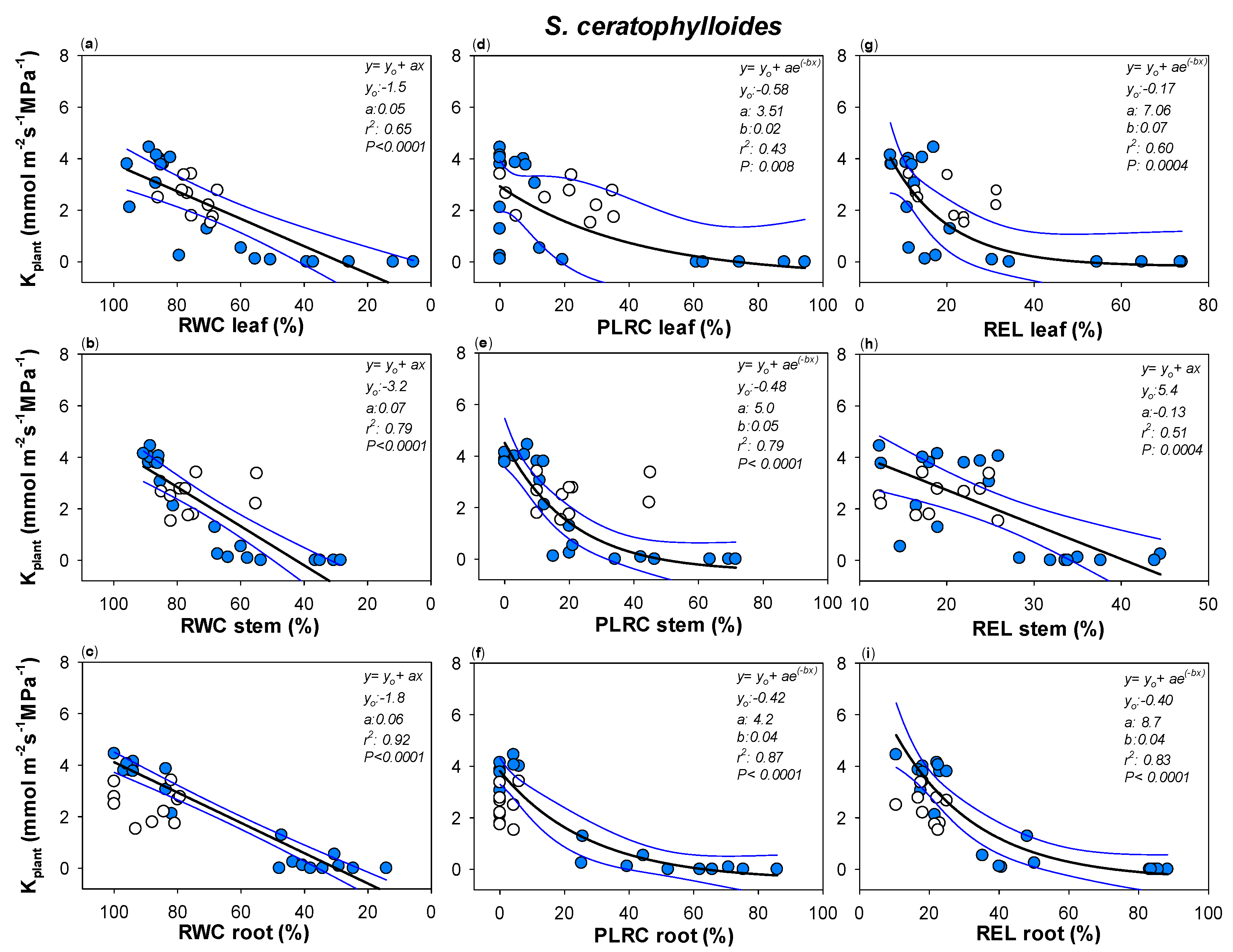
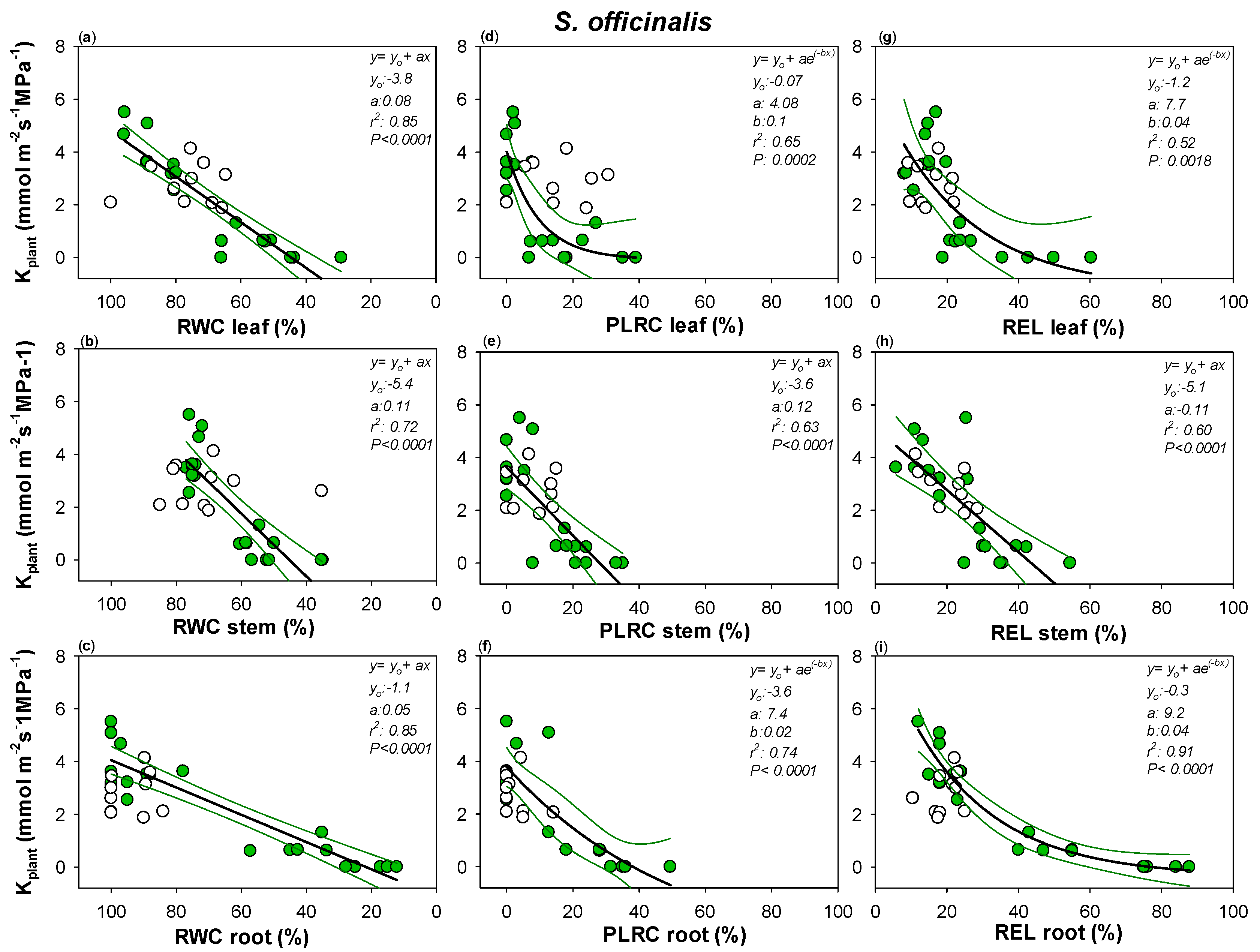
2. Results
3. Discussion
3.1. Two Different Drought Resistance Strategies but a Similar Root Hydraulics Recovery Ability
3.2. Water Content and Loss in Rehydration Capability Actually Drive Plant Hydraulics
4. Materials and Methods
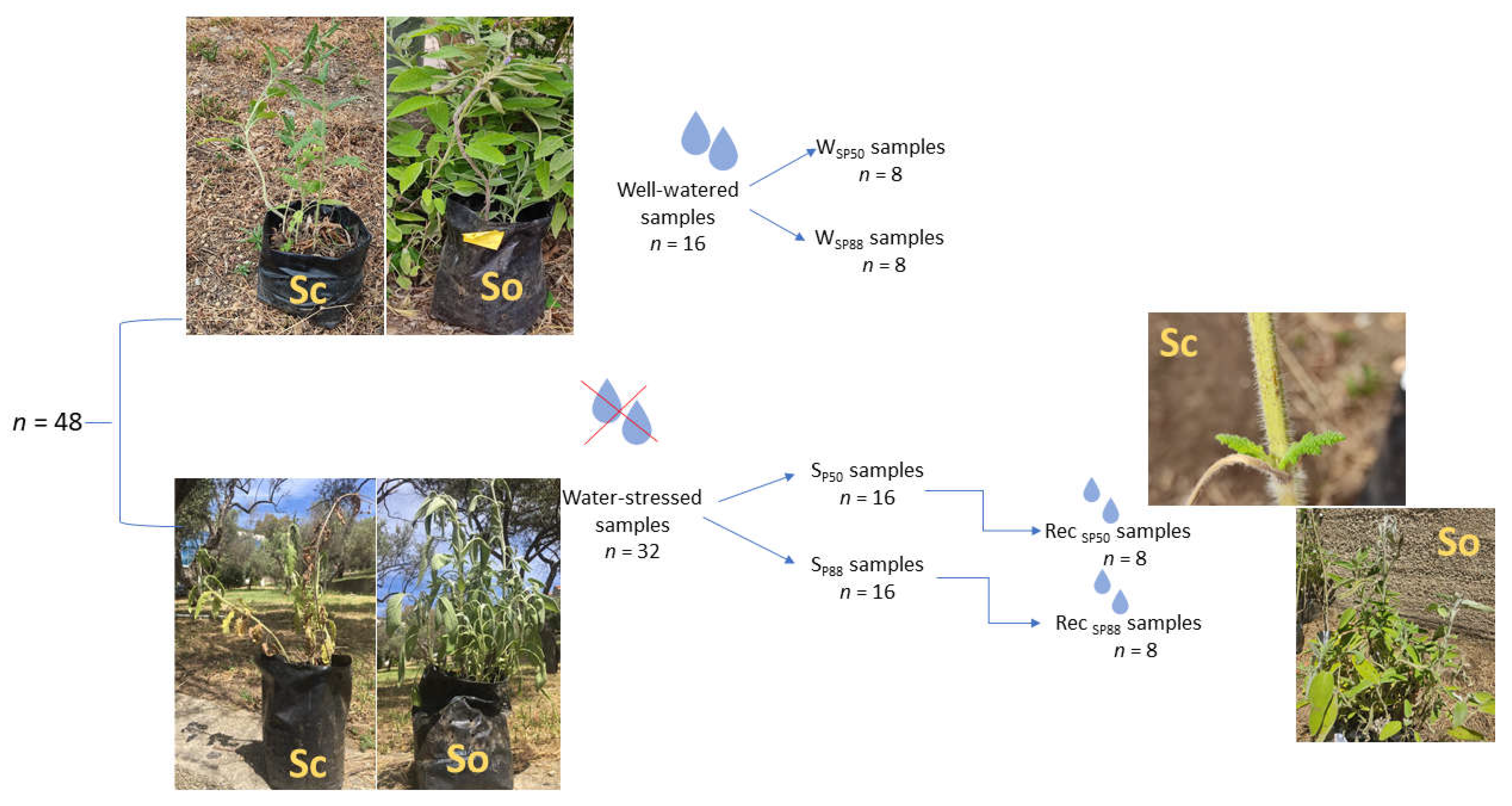
4.1. Plant Material and Growth Conditions
4.2. Gas Exchange and Water Status
4.3. Estimating the Relative Water Content, Rehydration Capacity and Cell Membrane Integrity of Leaf, Stem and Root Samples Experiencing Drought-Recovery Treatment
4.4. Structural Traits and Biomass Allocation
4.5. Plant Hydraulic Conductance Measurements by EFM
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Carnicer, J.; Coll, M.; Ninyerola, M.; Pons, X.; Sánchez, G.; Peñuelas, J. Widespread crown condition decline, food web disruption, and amplified tree mortality with increased climate change-type drought. Proc. Natl. Acad. Sci. USA 2011, 108, 1474–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menezes-Silva, P.E.; Loram-Lourenço, L.; Alves, R.D.F.B.; Sousa, L.F.; Almeida, S.E.D.S.; Farnese, F.S. Different ways to die in a changing world: Consequences of climate change for tree species performance and survival through an ecophysiological perspective. Ecol. Evol. 2019, 9, 11979–11999. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, F. Climate change hot-spots. Geophys. Res. Lett. 2006, 33, L08707. [Google Scholar] [CrossRef]
- Mariotti, A.; Pan, Y.; Zeng, N.; Alessandri, A. Long-term climate change in the Mediterranean region in the midst of decadal variability. Clim. Dyn. 2015, 44, 1337–1456. [Google Scholar] [CrossRef]
- Drobinski, P.; Da Silva, N.; Bastin, S.; Mailler, S.; Muller, C.; Ahrens, B.; Christensen, O.B.; Lionello, P. How warmer and drier will the Mediterranean region be at the end of the twenty-first century? Reg. Environ. Chang. 2020, 20, 78. [Google Scholar] [CrossRef]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S.; et al. Climate change and interconnected risks to sustainable development in the Mediterranean. Nat. Clim. Chang. 2018, 8, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Lionello, P.; Scarascia, L. The relation between climate change in the Mediterranean region and global warming. Reg. Environ. Chang. 2018, 12, 1481–1493. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Filella, I.; Estiarte, M.; Llusìa, J.; Ogaya, R.; Carnicer, J.; Bartrons, M.; Rivas-Ubach, A.; Grau, O.; et al. Assessment of the impacts of climate change on Mediterranean terrestrial ecosystems based on data from field experiments and long-term monitored field gradients in Catalonia. Environ. Exp. Bot. 2018, 152, 49–59. [Google Scholar] [CrossRef]
- Juhlke, T.R.; Van Geldern, R.; Barth, J.A.C.; Bendix, J.; Bräuning, A.; Garel, E.; Häusser, M.; Huneau, F.; Knerr, I.; Santoni, S.; et al. Temporal offset between precipitation and water uptake of Mediterranean pine trees varies with elevation and season. Sci. Total Environ. 2021, 755, 142539. [Google Scholar] [CrossRef]
- Underwood, E.C.; Viers, J.H.; Klausmeyer, K.R.; Cox, R.L.; Shaw, M.R. Threats and biodiversity in the Mediterranean biome. Divers. Distrib. 2009, 15, 188–197. [Google Scholar] [CrossRef]
- Ubran, M.C. Accelerating extinction risk from climate change. Science 2015, 348, 571–573. [Google Scholar]
- Molina-Venegas, R.; Ramos-Gutiérrez, I.; Moreno-Saiz, J.C. Phylogenetic patterns of extinction risk in the endemic flora of a mediterranean hotspot as a guiding tool for preemptive conservation actions. Front. Ecol. Evol. 2020, 8, 571587. [Google Scholar] [CrossRef]
- Rundel, P.W.; Arroyo, M.T.K.; Cowling, R.M.; Keeley, J.E.; Lamont, B.B.; Vargas, P. Mediterraneanbiomes: Evolution of their vegetation, floras, and climate. Ann. Rev. Ecol. Evol. Syst. 2016, 47, 383–407. [Google Scholar] [CrossRef]
- Molina-Venegas, R.; Aparicio, A.; Lavergne, S.; Arroyo, J. Climatic and topographical correlates of plant palaeo- and neoendemism in a Mediterranean biodiversity hotspot. Ann. Bot. 2017, 119, 229–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilagrosa, A.; Bellot, J.; Gil-Pelegrin, E. Cavitation, stomatal conductance, and leaf die-back in seedlings of two co-occuring Mediterranean shrubs during an intense drought. J. Exp. Bot. 2003, 390, 2015–2024. [Google Scholar] [CrossRef] [PubMed]
- Vilagrosa, A.; Hernández, E.I.; Luis, V.C.; Cochard, H.; Pausas, J.G. Physiological differences explain the co-existence of different regeneration strategies in Mediterranean ecosystems. New Phytol. 2014, 201, 1277–1288. [Google Scholar] [CrossRef]
- Hernandéz, E.I.; Vilagrosa, A.; Pausas, J.G.; Bellot, J. Morphological traits and water use strategies in seedlings of Mediterranean coexisting species. Plant Ecol. 2010, 207, 233–244. [Google Scholar] [CrossRef] [Green Version]
- West, A.G.; Dawson, T.E.; February, E.C.; Midgley, G.F.; Bond, W.J.; Aston, T.L. Diverse functional responses to drought in a Mediterranean-type shrubland in South Africa. New Phytol. 2012, 195, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Nardini, A.; Lo Gullo, M.A.; Trifilò, P.; Salleo, S. The challenge of the Mediterranean climate to plant hydraulics: Responses and adaptations. Environ. Exp. Bot. 2014, 103, 68–79. [Google Scholar] [CrossRef]
- Gazol, A.; Sanguesa-Barreda, G.; Granda, E.; Camarero, J.J. Tracking the impact of drought on functionally different woody plants in a Mediterranean scrubland ecosystem. Plant Ecol. 2017, 218, 1009–1020. [Google Scholar] [CrossRef]
- McDowell, N.G.; Brodribb, T.J.; Nardini, A. Hydraulics in the 21st century. New Phytol. 2019, 224, 537–542. [Google Scholar] [CrossRef] [Green Version]
- Anderegg, W.R.L.; Klein, T.; Bartlett, M.; Sack, L.; Pellegrini, A.F.A.; Choat, B.; Jansen, S. Meta-analysis reveals that hydraulic traits explain cross-species patterns of drought-induced tree mortality across the globe. Proc. Natl. Acad. Sci. USA 2016, 113, 5024–5029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursuma, R.A.; López, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.; Zeppel, M.J.B.; Anderegg, W.R.L.; Bloemen, J.; De Kauwe, M.G.; Hudson, P.; Ruehr, N.K.; Powell, T.L.; von Arx, G.; Nardini, A. Xylem embolism refilling and resilience against drought-induced mortality in woody plants: Processes and trade-offs. Ecol. Res. 2018, 33, 839–855. [Google Scholar] [CrossRef]
- Martinez-Vilalta, J.; Anderegg, W.R.L.; Sapes, G.; Sala, A. Greater focus on water pools may improve our ability to understand and anticipate drought-induced mortality in plants. New Phytol. 2019, 223, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Sapes, G.; Roskilly, B.; Dobrowski, S.; Maneta, M.; Anderegg, W.R.L.; Martinez-Vilalta, J.; Sala, A. Plant water content integrates hydraulics and carbon depletion to predict drought-induced seedling mortality. Tree Physiol. 2019, 39, 1300–1312. [Google Scholar] [CrossRef]
- Sapes, G.; Sala, A. Relative water content consistently predicts drought mortality risk in seedling populations with different morphology, physiology, and times to death. Plant. Cell Environ. 2021, in press. [Google Scholar] [CrossRef]
- Rosner, S.; Heinze, B.; Savi, T.; Dalla-Salda, G. Prediction of hydraulic conductivity loss from relative water loss: New insights into water storage of tree stems and branches. Physiol. Plant. 2019, 165, 843–854. [Google Scholar] [CrossRef] [Green Version]
- Mantova, M.; Menezes-Silva, P.E.; Badel, E.; Cochard, H.; Torres-Ruiz, J.M. The interplay of hydraulic failure and cell vitality explains tree capacity to recover from drought. Physiol. Plant. 2021, 172, 247–257. [Google Scholar] [CrossRef]
- Crisafulli, A.; Cannavò, S.; Maiorca, G.; Musarella, C.M.; Signorino, G.; Spampinato, G. Aggiornamenti floristici per la Calabria. Inf. Bot. Ital. 2010, 42, 437–448. [Google Scholar]
- Spampinato, G.; Crisafulli, A.; Marino, A.; Signorino, G. Salvia ceratophylloides Ard. Inf. Bot. Ital. 2011, 43, 381–458. [Google Scholar]
- Vescio, R.; Abenavoli, M.R.; Araniti, F.; Musarella, C.M.; Sofo, A.; Laface, V.L.A.; Spampinato, G.; Sorgonà, A. The assessment and the within-plant variation of the morpho-physiological traits and VOCs profile in endemic and rare Salvia ceratophylloides Ard. (Lamiaceae). Plants 2021, 10, 474. [Google Scholar] [CrossRef]
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italy, 2002. [Google Scholar]
- Raimondo, F.; Trifilò, P.; Lo Gullo, M.A.; Andri, S.; Savi, T.; Nardini, A. Plant performance on Mediterranean green roofs: Interaction of species-specific hydraulic strategies and substrate water relations. AoB Plants 2015, 7, plv007. [Google Scholar] [CrossRef]
- Savi, T.; Marin, M.; Luglio, J.; Petruzzellis, F.; Mayr, S.; Nardini, A. Leaf hydraulic vulnerability protects stem functionality under drought stress in Salvia officinalis. Funct. Plant Biol. 2016, 43, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Blackman, C.J.; Gleason, S.M.; Chang, Y.; Cook, A.M.; Laws, C.; Westoby, M. Leaf hydraulic vulnerability to drought is linked to site water availability across a broad range of species and climates. Ann. Bot. 2014, 114, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Nardini, A.; Luglio, J. Leaf hydraulic capacity and drought vulnerability: Possible trade-offs and correlations with climate across three major biomes. Funct. Ecol. 2014, 28, 810–818. [Google Scholar] [CrossRef]
- Fang, J.; Lutz, J.A.; Wang, L.; Shugart, H.H.; Yan, X. Using climate-driven leaf phenology and growth to improve predictions of gross primary productivity in North American forests. Glob. Chang. Biol. 2020, 26, 6974–6988. [Google Scholar] [CrossRef] [PubMed]
- Abate, E.; Nardini, A.; Petruzzellis, F.; Trifilò, P. Too dry to survive: Leaf hydraulic failure in two Salvia species can be predicted on the basis of water content. Plant Physiol. Biochem. 2021, 166, 215–224. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Brouillette, L.C.; Mason, C.M.; Shirk, R.Y.; Donovan, L.A. Adaptive differentiation of traits related to resource use in a desert annual along a resource gradient. New Phytol. 2014, 201, 1316–1327. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, U.; Poorter, L.; Wright, J.J.; Villar, R. Causes and consequences of variation in leaf massa area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef]
- Forner, A.; Valladeres, F.; Aranda, I. Mediterranean trees coping with severe drought: Avoidance might not be safe. Environ. Exp. Bot. 2014, 155, 529–540. [Google Scholar] [CrossRef]
- North, G.B.; Nobel, P.S. Changes in hydraulic conductivity and anatomy caused by drying and rewetting roots of Agave deserti. Am. J. Bot. 1991, 78, 906–915. [Google Scholar] [CrossRef]
- North, G.B.; Nobel, P.S. Radial hydraulic conductivity of individual root yissues of Opuntia ficus-indica (L.) Miller as soil moisture varies. Ann. Bot. 1996, 77, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Trifilò, P.; Raimondo, F.; Nardini, A.; Lo Gullo, M.A.; Salleo, S. Drought resistance of Ailanthus altissima: Root hydraulics and water relations. Tree Physiol. 2004, 24, 107–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuneo, I.F.; Barrios-Masias, F.; Knipfer, T.; Uretsky, J.; Reyes, C.; Lenain, P.; Brodersen, C.R.; Walker, M.A.; McElrone, A.J. Differences in grapevine rootstock sensitivity and recovery from drought are linked to fine root cortical lacunae and root tip function. New Phytol. 2021, 229, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Nobel, P.S.; Sanderson, J. Rectifier-like activities of roots of two desert succulents. J. Exp. Bot. 1984, 35, 727–737. [Google Scholar] [CrossRef]
- Padilla, F.M.; Miranda, J.D.; Jorquera, M.J.; Pugnaire, F.I. Variability in amount and frequency of water supply affects roots but not growth of arid shrubs. Plant Ecol. 2009, 204, 261–270. [Google Scholar] [CrossRef]
- Markesteijn, L.; Poorter, L. Seedling root morphology and biomass allocation of 62 tropical tree species in relation to drought- and shade-tolerance. J. Ecol. 2009, 97, 311–325. [Google Scholar] [CrossRef]
- Lopez_Iglesis, B.; Villar, R.; Poorter, L. Functional traits predict drought performance and distribution of Mediterranean woody species. Acta Oecol. 2014, 56, 10–18. [Google Scholar] [CrossRef]
- Creek, D.; Blackmann, C.J.; Brodribb, T.J.; Choat, B. Coordination between leaf, stem, and root hydraulics and gas exchange in three arid-zone angiosperms during severe drought and recovery. Plant Cell Environ. 2018, 12, 2869–2881. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Domiguez, C.M.; Brodribb, T.J. Declining root water transport drives stomatal closure in olive under moderate water stress. New Phytol. 2019, 225, 126–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourbia, I.; Pritzkow, C.; Brodribb, T.J. Herb and conifer roots show similar high sensitivity to water deficit. Plant Physiol. 2021, 186, 1908–1918. [Google Scholar] [CrossRef]
- Weigelt, A.; Mommer, L.; Andraczek, K.; Iversen, C.M.; Bergmann, J.; Bruelheide, H.; Fan, Y.; Freschet, G.T.; Guerrero-Ramírez, N.R.; Kattge, J.; et al. An integrated framework of plant form and function: The belowground perspective. New Phytol. 2021, 232, 42–59. [Google Scholar] [CrossRef]
- Pivovaroff, A.L.; Sack, L.; Santiago, L.S. Coordination of stem and leaf hydraulic conductance in southern California shrubs: A test of the hydraulic segmentation hypothesis. New Phytol. 2014, 203, 842–850. [Google Scholar] [CrossRef] [Green Version]
- Trifilò, P.; Petruzzellis, F.; Abate, E.; Nardini, A. The extra-vascular water pathway regulates dynamic leaf hydraulic decline and recovery in Populus nigra. Physiol. Plant. 2021, 172, 29–40. [Google Scholar] [CrossRef]
- Tsuda, M.; Tyree, M.T. Plant hydraulic conductance measured by the high pressure flow meter in crop plants. J. Exp. Bot. 2000, 51, 823–828. [Google Scholar] [CrossRef]
- Trifilò, P.; Nardini, A.; Raimondo, F.; Lo Gullo, M.A.; Salleo, S. Ion-mediated compensation for drought-induced loss of xylem hydraulic conductivity in field-growing plants of Laurus nobilis. Funct. Plant Biol. 2010, 38, 606–613. [Google Scholar] [CrossRef]
- Aranda, I.; Gil, L.; Pardos, J.A. Seasonal changes in apparent hydraulic conductance and their implications for water use of European beech (Fagus sylvatica L.) and sessile oak [Quercus petraea (Matt.) Liebl] in South Europe. Plant Ecol. 2005, 179, 155–167. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | S. ceratophylloides | S. officinalis | p Value |
|---|---|---|---|
| Structural and biomass data | |||
| N leaves/plant | 47.7 ± 7.7 | 164.7 ± 59.3 | <0.001 |
| AL (cm2) | 776 ± 80 | 2788 ± 976 | <0.001 |
| LDMC | 0.16 ± 0.01 | 0.25 ± 0.03 | <0.001 |
| DW leaves, g | 2.2 ± 0.9 | 12.7 ± 1.9 | <0.001 |
| N shoots/plant | 2.4 ± 0.5 | 2.5 ± 1.3 | 0.411 |
| DW stem, g | 0.3 ± 0.1 | 1.1 ± 0.2 | <0.001 |
| SDMC | 0.22 ± 0.03 | 0.31 ± 0.03 | <0.001 |
| Stem/leaf ratio | 1.78 ± 0.95 | 0.95 ± 0.02 | 0.007 |
| DW root, g | 1.1 ± 0.4 | 1.4 ± 0.4 | 0.104 |
| RDMC | 0.14 ± 0.03 | 0.18 ± 0.01 | 0.004 |
| Root/shoot ratio | 2.2 ± 0.9 | 0.6 ± 0.3 | <0.001 |
| Water storage traits | |||
| SWCleaf, g g−1 | 4.8 ± 0.4 | 2.7 ± 0.3 | <0.001 |
| SWCstem, g g−1 | 3.3 ± 0.6 | 2.4 ± 0.6 | <0.001 |
| SWCroot, g g−1 | 5.2 ± 1.4 | 4.4 ± 0.5 | 0.061 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abate, E.; Azzarà, M.; Trifilò, P. When Water Availability Is Low, Two Mediterranean Salvia Species Rely on Root Hydraulics. Plants 2021, 10, 1888. https://doi.org/10.3390/plants10091888
Abate E, Azzarà M, Trifilò P. When Water Availability Is Low, Two Mediterranean Salvia Species Rely on Root Hydraulics. Plants. 2021; 10(9):1888. https://doi.org/10.3390/plants10091888
Chicago/Turabian StyleAbate, Elisa, Maria Azzarà, and Patrizia Trifilò. 2021. "When Water Availability Is Low, Two Mediterranean Salvia Species Rely on Root Hydraulics" Plants 10, no. 9: 1888. https://doi.org/10.3390/plants10091888
APA StyleAbate, E., Azzarà, M., & Trifilò, P. (2021). When Water Availability Is Low, Two Mediterranean Salvia Species Rely on Root Hydraulics. Plants, 10(9), 1888. https://doi.org/10.3390/plants10091888