
Benefits of Cardamom (Elettaria cardamomum (L.) Maton) and Turmeric (Curcuma longa L.) Extracts for Their Applications as Natural Anti-Inflammatory Adjuvants


 , , , ,
, , , ,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Material and Extraction
2.3. Qualitative Phytochemical Tests
2.4. GC and GC-MS/MS Analysis of the Aqueous Extracts
2.5. Gene Expression of Anti-Inflammatory Cytokines on Peritoneal Macrophages Stimulated with Aqueous Extracts and Challenged with Lipopolysaccharide
2.5.1. Obtaining Peritoneal Cells
2.5.2. RNA Extraction
2.5.3. Complementary DNA (cDNA) Synthesis
2.5.4. Oligonucleotide Design
2.5.5. Real Time—qPCR (RT-qPCR)
2.6. Index of Cytotoxicity of the Aqueous Extract of E. cardamomum and C. longa on Cell Cultures
2.7. Nitric Oxide Assay
2.8. Statistical Analysis
3. Results
3.1. Phytochemical Tests
3.2. Compound Identification
3.3. Gene Expression of Anti-Inflammatory Cytokines
3.4. Cytotoxic Activity of the Aqueous Extract
3.5. Nitric Oxide Depletion in Peritoneal Macrophages
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sánchez-Ramos, M.; Alvarez, L.; Romero-Estrada, A.; Bernabé-Antonio, A.; Marquina-Bahena, S.; Cruz-Sosa, F. Establishment of a cell suspension culture of ageratina pichinchensis (Kunth) for the improved production of anti-inflammatory compounds. Plants 2020, 9, 1398. [Google Scholar] [CrossRef] [PubMed]
- Elizondo-Luévano, J.H.; Castro-Ríos, R.; Sánchez-García, E.; Hernández-García, M.E.; Vargas-Villarreal, J.; Rodríguez-Luis, O.E.; Chávez-Montes, A. In vitro study of antiamoebic activity of methanol extracts of argemone mexicana on trophozoites of entamoeba histolytica HM1-IMSS. Can. J. Infect. Dis. Med. Microbiol. 2018, 2018, 7453787. [Google Scholar] [CrossRef] [PubMed]
- Frencken, J.E.; Sharma, P.; Stenhouse, L.; Green, D.; Laverty, D.; Dietrich, T. Global epidemiology of dental caries and severe periodontitis—A comprehensive review. J. Clin. Periodontol. 2017, 44, S94–S105. [Google Scholar] [CrossRef]
- Soehnlein, O.; Steffens, S.; Hidalgo, A.; Weber, C. Neutrophils as protagonists and targets in chronic inflammation. Nat. Rev. Immunol. 2017, 17, 248–261. [Google Scholar] [CrossRef]
- Konkel, J.E.; O’Boyle, C.; Krishnan, S. Distal consequences of oral inflammation. Front. Immunol. 2019, 10, 1403. [Google Scholar] [CrossRef] [Green Version]
- Scannapieco, F.A.; Cantos, A. Oral inflammation and infection, and chronic medical diseases: Implications for the elderly. Periodontology 2016, 72, 153–175. [Google Scholar] [CrossRef] [PubMed]
- Rider, P.; Carmi, Y.; Cohen, I. Biologics for targeting inflammatory cytokines, clinical uses, and limitations. Int. J. Cell Biol. 2016, 2016, 9259646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, E.M.; Reis, C.; Manzanares-Céspedes, M.C. Chronic periodontitis, inflammatory cytokines, and interrelationship with other chronic diseases. Postgrad. Med. 2018, 130, 98–104. [Google Scholar] [CrossRef]
- Dini, M. Peptic ulcer disease and non-steroidal anti-inflammatory drugs. Aust. Prescr. 2017, 40, 91–93. [Google Scholar]
- Little, P. Non-steroidal anti-inflammatory drugs and covid-19. BMJ 2020, 368, m1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanović, M.; Makoter, K.; Razboršek, M.I. Comparative study of chemical composition and antioxidant activity of essential oils and crude extracts of four characteristic zingiberaceae herbs. Plants 2021, 10, 501. [Google Scholar] [CrossRef]
- Rajan, A.; Rajan, A.R.; Philip, D. Elettaria cardamomum seed mediated rapid synthesis of gold nanoparticles and its biological activities. Open Nano 2017, 2, 1–8. [Google Scholar] [CrossRef]
- Souissi, M.; Azelmat, J.; Chaieb, K.; Grenier, D. Antibacterial and anti-inflammatory activities of cardamom (Elettaria cardamomum) extracts: Potential therapeutic benefits for periodontal infections. Anaerobe 2020, 61, 102089. [Google Scholar] [CrossRef]
- Tanvir, E.M.; Hossen, M.S.; Hossain, M.F.; Afroz, R.; Gan, S.H.; Khalil, M.I.; Karim, N. Antioxidant properties of popular turmeric (Curcuma longa) varieties from Bangladesh. J. Food Qual. 2017, 2017, 8471785. [Google Scholar] [CrossRef] [Green Version]
- Soleimani, V.; Sahebkar, A.; Hosseinzadeh, H. Turmeric (Curcuma longa) and its major constituent (curcumin) as nontoxic and safe substances: Review. Phyther. Res. 2018, 32, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Elizondo-Luévano, J.H.; Hernández-García, M.E.; Pérez-Narváez, O.A.; Castro-Ríos, R.; Chávez-Montes, A. Berberina, curcumina y quercetina como potenciales agentes con capacidad antiparasitaria. Rev. Biol. Trop. 2020, 68, 1241–1249. [Google Scholar] [CrossRef]
- White, C.M.; Pasupuleti, V.; Roman, Y.M.; Li, Y.; Hernandez, A.V. Oral turmeric/curcumin effects on inflammatory markers in chronic inflammatory diseases: A systematic review and meta-analysis of randomized controlled trials. Pharmacol. Res. 2019, 146, 104280. [Google Scholar] [CrossRef]
- Elizondo-Luevano, J.H.; Verde-Star, J.; González-Horta, A.; Castro-Ríos, R.; Hernández-García, M.E.; Chávez-Montes, A. In Vitro Effect of Methanolic Extract of Argemone mexicana against Trichomonas vaginalis. Korean J. Parasitol. 2020, 58, 135–145. [Google Scholar] [CrossRef]
- Parekh, J.; Chanda, S.V. In vitro antimicrobial activity and phytochemical analysis of some Indian medicinal plants. Turkish J. Biol. 2007, 31, 53–58. [Google Scholar]
- Bekinbo, M.; Tariah FS, A.; Dapper, D. Comparative GC-MS determination of bioactive constituents of the methanolic extracts of Curcuma longa rhizome and Spondias mombin leaves Bekinbo MT, Amah-Tariah FS and Dapper DV. J. Med. Plants Stud. 2020, 8, 1–6. [Google Scholar]
- Masoumi-Ardakani, Y.; Mandegary, A.; Esmaeilpour, K.; Najafipour, H.; Sharififar, F.; Pakravanan, M.; Ghazvini, H. Chemical composition, anticonvulsant activity, and toxicity of essential oil and methanolic extract of Elettaria cardamomum. Planta Med. 2016, 82, 1482–1486. [Google Scholar] [CrossRef]
- Salvesen, Ø.; Reiten, M.R.; Espenes, A.; Bakkebø, M.K.; Tranulis, M.A.; Ersdal, C. LPS-induced systemic inflammation reveals an immunomodulatory role for the prion protein at the blood-brain interface. J. Neuroinflam. 2017, 14, 106. [Google Scholar] [CrossRef] [Green Version]
- Humberto, C.H.-M.; Ricardo, G.-F.; Patricia, T.-G.; Ramiro, Q.-L.; Mario, A.S.E.; Enriqueta, M.-C.; Reyes, T.-G.; Cristina, R.-P. Antitumor activity of Pachycereus marginatus (DC.) Britton Rose extracts against murine lymphoma L5178Y-R and skin melanoma B16F10 cells. J. Med. Plants Res. 2016, 10, 635–639. [Google Scholar] [CrossRef] [Green Version]
- Castillo-Velázquez, U.; Aranday-Cortés, E.; Gutiérrez-Pabello, J.A. Alternative activation modifies macrophage resistance to Mycobacterium bovis. Vet. Microbiol. 2011, 151, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Nevárez-Garza, A.M.; Castillo-Velázquez, U.; Soto-Domínguez, A.; Montes-de-Oca-Luna, R.; Zamora-Ávila, D.E.; Wong-González, A.; Rodríguez-Tovar, L.E. Quantitative analysis of TNF-α IL-4, and IL-10 expression, nitric oxide response, and apoptosis in Encephalitozoon cuniculi–infected rabbits. Dev. Comp. Immunol. 2018, 81, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Delgado, A.J.M.; Velázquez, U.C.; González, J.G.B.; Montes, A.C.; Villarreal, S.M.L.; García, L.E.V.; Casas, R.M.S.; Luis, O.E.R. Evaluation of the Essential Oil of Citrus paradisi as an Alternative Treatment against Candida albicans. Open J. Stomatol. 2020, 10, 258–270. [Google Scholar] [CrossRef]
- Adler, B.; Adler, H.; Jungi, T.W.; Peterhans, E. Interferon-α primes macrophages for lipopolysaccharide-induced apoptosis. Biochem. Biophys. Res. Commun. 1995, 215, 921–927. [Google Scholar] [CrossRef]
- Chandran, B.; Goel, A. A randomized, pilot study to assess the efficacy and safety of curcumin in patients with active rheumatoid arthritis. Phyther. Res. 2012, 26, 1719–1725. [Google Scholar] [CrossRef]
- Sandner, G.; Heckmann, M.; Weghuber, J. Immunomodulatory activities of selected essential oils. Biomolecules 2020, 10, 1139. [Google Scholar] [CrossRef]
- Noumi, E.; Snoussi, M.; Alreshidi, M.M.; Rekha, P.D.; Saptami, K.; Caputo, L.; De Martino, L.; Souza, L.F.; Msaada, K.; Mancini, E.; et al. Chemical and biological evaluation of essential oils from cardamom species. Molecules 2018, 23, 2818. [Google Scholar] [CrossRef] [Green Version]
- Githinji, C.G.; Mbugua, P.M.; Kanui, T.I.; Kariuki, D.K. Analgesic and anti-inflammatory activities of 9-Hexacosene and Stigmasterol isolated from Mondia whytei. Phytopharmacology 2012, 2, 212–223. [Google Scholar]
- Nogueira, M.N.M.; Aquino, S.G.; Rossa, C.; Spolidorio, D.M.P. Terpinen-4-ol and alpha-terpineol (tea tree oil components) inhibit the production of IL-1β, IL-6 and IL-10 on human macrophages. Inflamm. Res. 2014, 63, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Abe, S.; Maruyama, N.; Hayama, K.; Ishibashi, H.; Inoue, S.; Oshima, H.; Yamaguchi, H. Suppression of tumor necrosis factor-alpha-induced neutrophil adherence responses by essential oils. Mediat. Inflamm. 2003, 12, 323–328. [Google Scholar] [CrossRef]
- Hamdan, D.I.; Abdulla, R.H.; Mohamed, M.E.; El-Shazly, A.M. Chemical composition and biological activity of essential oils of cleopatra mandarin (citrus reshni) cultivated in Egypt. J. Pharmacogn. Phyther. 2013, 5, 83–90. [Google Scholar] [CrossRef]
- Djouahri, A.; Boualem, S.; Boudarene, L.; Baaliouamer, A. Geographic’s variation impact on chemical composition, antioxidant and anti-inflammatory activities of essential oils from wood and leaves of Tetraclinis articulata (Vahl) Masters. Ind. Crops Prod. 2015, 63, 138–146. [Google Scholar] [CrossRef]
- Valente, J.; Resende, R.; Zuzarte, M.; Gonçalves, M.J.; Lopes, M.C.; Cavaleiro, C.; Pereira, C.; Salgueiro, L.; Cruz, M.T. Bioactivity and safety profile of Daucus carota subsp. maximus essential oil. Ind. Crops Prod. 2015, 77, 218–224. [Google Scholar] [CrossRef]
- Angel, G.R.; Menon, N.; Vimala, B.; Nambisan, B. Essential oil composition of eight starchy Curcuma species. Ind. Crop. Prod. 2014, 60, 233–238. [Google Scholar] [CrossRef]
- Niranjan, A.; Prakash, D. Chemical constituents and biological activities of turmeric (Curcuma longa L.)—A review. J. Food Sci. Technol. 2008, 45, 109–116. [Google Scholar]
- Liju, V.; Jeena, K.; Kuttan, R. An evaluation of antioxidant, anti-inflammatory, and antinociceptive activities of essential oil from Curcuma longa L. Indian J. Pharmacol. 2011, 43, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Han, A.R.; Park, H.R.; Jang, E.J.; Kim, H.K.; Jeong, M.G.; Song, H.; Park, G.H.; Seo, E.K.; Hwang, E.S. Suppression of inflammatory cytokine production by ar-turmerone isolated from Curcuma phaeocaulis. Chem. Biodivers. 2014, 11, 1034–1041. [Google Scholar] [CrossRef]
- Guillamón, E. Effect of phytochemical compounds of the genus Allium on the immune system and the inflammatory response. Ars Pharm. 2018, 59, 185–196. [Google Scholar] [CrossRef]
- Freire-González, R.A.; Vistel-Vigo, M. Caracterizacion fitoquimica de la curcuma. Rev. Cuba. Quim. 2015, 27, 9–18. [Google Scholar]
- Dávila-Martínez, C.; Castillo-Velázquez, U.; Soto-Domínguez, A.; Nevárez-Garza, A.M.; Arce-Mendoza, A.Y.; Hernandez-Vidal, G.; Zamora-Avila, D.E.; Rodriguez-Tovar, L.E. Immunohistochemical localization of TNF-α and IL-4 in granulomas of immunocompetent and immunosuppressed New Zealand white rabbits infected with Encephalitozoon cuniculi. Cytokine 2020, 130, 155055. [Google Scholar] [CrossRef] [PubMed]
- Koppula, S.; Kumar, H.; Kim, I.S.; Choi, D.K. Reactive oxygen species and inhibitors of inflammatory enzymes, NADPH oxidase, and iNOS in experimental models of parkinsons disease. Mediat. Inflamm. 2012, 823902. [Google Scholar] [CrossRef] [Green Version]
- Baranipour, S.; Kadijani, A.A.; Qujeq, D.; Shahrokh, S.; Haghazali, M.; Mirzaei, A.; Asadzadeh-Aghdaei, H. Inducible nitric oxide synthase as a potential blood-based biomarker in inflammatory bowel diseases. Gastroenterol. Hepatol. Bed Bench 2018, 11, S124–S128. [Google Scholar] [CrossRef] [PubMed]
- Witte, M.B.; Barbul, A. Arginine physiology and its implication for wound healing. Wound Repair Regen. 2003, 11, 419–423. [Google Scholar] [CrossRef]
- Yoshino, F.; Yoshida, A.; Nakajima, A.; Wada-Takahashi, S.; Takahashi, S.; Lee, M.C. Il Alteration of the redox state with reactive oxygen species for 5-fluorouracil-induced oral mucositis in hamsters. PLoS ONE 2013, 8, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Curra, M.; Martins, M.A.T.; Lauxen, I.S.; Pellicioli, A.C.A.; Sant’Ana Filho, M.; Pavesi, V.C.S.; Carrard, V.C.; Martins, M.D. Effect of topical chamomile on immunohistochemical levels of IL-1β and TNF-α in 5-fluorouracil-induced oral mucositis in hamsters. Cancer Chemother. Pharmacol. 2013, 71, 293–299. [Google Scholar] [CrossRef]
- Majdalawieh, A.F.; Carr, R.I. In vitro investigation of the potential immunomodulatory and anti-cancer activities of black pepper (Piper nigrum) and cardamom (Elettaria cardamomum). J. Med. Food 2010, 13, 371–381. [Google Scholar] [CrossRef]
- Yadav, N.; Chandra, H. Suppression of inflammatory and infection responses in lung macrophages by eucalyptus oil and its constituent 1,8-cineole: Role of pattern recognition receptors TREM-1 and NLRP3, the MAP kinase regulator MKP-1, and NFκB. PLoS ONE 2017, 12, e0188232. [Google Scholar] [CrossRef] [Green Version]
- Adcock, I.M.; Mumby, S. Glucocorticoids. Handb. Exp. Pharmacol. 2017, 237, 171–196. [Google Scholar] [CrossRef]
- Conlon, K.C.; Miljkovic, M.D.; Waldmann, T.A. Cytokines in the treatment of cancer. J. Interferon Cytokine Res. J. Int. Soc. Interferon Cytokine Res. 2019, 39, 6–21. [Google Scholar] [CrossRef]
- Billiau, A. Anti-inflammatory properties of Type I interferons. Antivir. Res. 2006, 71, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Holdsworth, S.R.; Can, P.Y. Cytokines: Names and numbers you should care about. Clin. J. Am. Soc. Nephrol. 2015, 10, 2243–2254. [Google Scholar] [CrossRef]
- Elizondo-Luévano, J.H.; Castro-Ríos, R.; Vicente, B.; Fernández-Soto, P.; López-Aban, J.; Muro, A.; Chávez-Montes, A. In vitro antischistosomal activity of the argemone mexicana methanolic extract and its main component berberine. Iran. J. Parasitol. 2021, 16, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Elizondo-Luévano, J.H.; Castro-Ríos, R.; López-Abán, J.; Gorgojo-Galindo, O.; Fernández-Soto, P.; Vicente, B.; Muro, A.; Chávez-Montes, A. Berberine: A nematocidal alkaloid from Argemone mexicana against Strongyloides venezuelensis. Exp. Parasitol. 2021, 220, 108043. [Google Scholar] [CrossRef] [PubMed]
- Malek, S.N.A.; Phang, C.W.; Ibrahim, H.; Wahab, N.A.; Sim, K.S. Phytochemical and cytotoxic investigations of Alpinia mutica rhizomes. Molecules 2011, 16, 583–589. [Google Scholar] [CrossRef] [Green Version]
- Akram, M.; Ahmed, A.; Usmanghani, K.; Hannan, A.; Mohiuddin, E.; Asif, M. Curcuma longa and curcumin: A review article. Rom. J. Biol. 2010, 55, 65–70. [Google Scholar]
- Berbudi, A.; Rahmi, N.; Atik, N.; Wikayani, T.; Qomarilla, N.; Rahayu, N.S.; Putri, A.C. The administration of low-dose curcuma longa extract induces m2 polarization in peritoneal macrophage culture. Res. J. Pharm. Technol. 2021, 14, 1079–1084. [Google Scholar] [CrossRef]
- Figueira, L.W.; de Oliveira, J.R.; Camargo, S.E.A.; de Oliveira, L.D. Curcuma longa L. (turmeric), Rosmarinus officinalis L. (rosemary), and Thymus vulgaris L. (thyme) extracts aid murine macrophages (RAW 264.7) to fight Streptococcus mutans during in vitro infection. Arch. Microbiol. 2020, 202, 2269–2277. [Google Scholar] [CrossRef]
- Okuda-Hanafusa, C.; Uchio, R.; Fuwa, A.; Kawasaki, K.; Muroyama, K.; Yamamoto, Y.; Murosaki, S. Turmeronol A and turmeronol B from Curcuma longa prevent inflammatory mediator production by lipopolysaccharide-stimulated RAW264.7 macrophages, partially via reduced NF-κB signaling. Food Funct. 2019, 10, 5779–5788. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, M.D.; Blázquez, M.A. Curcuma longa l. Rhizome essential oil from extraction to its agri-food applications. A review. Plants 2021, 10, 44. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.W.; Kim, H.; Park, J.Y.; Kim, J.E.; Moon, J.Y.; Park, S.D.; Park, W.H. Amomum cardamomum L. ethyl acetate fraction protects against carbon tetrachloride-induced liver injury via an antioxidant mechanism in rats. BMC Complement. Altern. Med. 2016, 16, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.Y.; Cho, S.S.; Li, Y.C.; Bae, C.S.; Park, K.M.; Park, D.H. Anti-inflammatory Effect of Curcuma longa and Allium hookeri Co-treatment via NF-κB and COX-2 Pathways. Sci. Rep. 2020, 10, 5718. [Google Scholar] [CrossRef]
- Lee, T.K.; Trinh, T.A.; Lee, S.R.; Kim, S.; So, H.M.; Moon, E.; Hwang, G.S.; Kang, K.S.; Kim, J.H.; Yamabe, N.; et al. Bioactivity-based analysis and chemical characterization of anti-inflammatory compounds from Curcuma zedoaria rhizomes using LPS-stimulated RAW264.7 cells. Bioorg. Chem. 2019, 82, 26–32. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aqueous Extract | Concentration (µg/mL) |
|---|---|
| E. cardamomum | 100 |
| C. longa | 70 |
| Gene | Genebank ID | Oligonucleotides |
|---|---|---|
| IL-4 | NM_021283.2 | F: 5′ TTG AGA GAG ATC ATC GGC ATT T -3′ R: 5′ CTC ACT CTC TGT GGT GTT CTT C -3′ |
| IL-6 | NM_031168.2 | F: 5′ CTT CCA TCC AGT TGC CTT CT -3′ R: 5′ CTC CGA CTT GTG AAG TGG TAT AG -3′ |
| IL-10 | NM_010548.2 | F: 5′ TTG AAT TCC CTG GGT GAG AAG -3′ R: 5′ TCC ACT GCC TTG CTC TTA TTT -3′ |
| Gpd1 | NM_010271.3 | F: 5′ CCT ACT GCT GAC CTT TCT TCT C -3′ R: 5′ GCC CTG AGG ACG ATA AAC TAT AA -3′ |
| TNF-α | NM_013693.3 | F: 5′ TTG TCT ACT CCC AGG TTC TCT -3′ R: 5′ GAG GTT GAC TTT CTC CTG GTA TG -3′ |
| β-actin | NM_07393.5 | F: 5′- GAG GTA TCC TGA CCC TGA AGT A -3′ R: 5′- CAC ACG CAG CTC ATT GTA GA -3′ |
| Test | Chemical Groups | E. cardamomum | C. longa |
|---|---|---|---|
| Lieberman Burchard | Sterols, triterpenes | + | + |
| Shinoda | Flavonoids | + | + |
| Baljet | Sesquiterpene lactones | + | + |
| Sulfuric acid | Quinones | − | − |
| Ferric chloride | Tannins | + | − |
| Yield % | 5.64 | 9.83 |
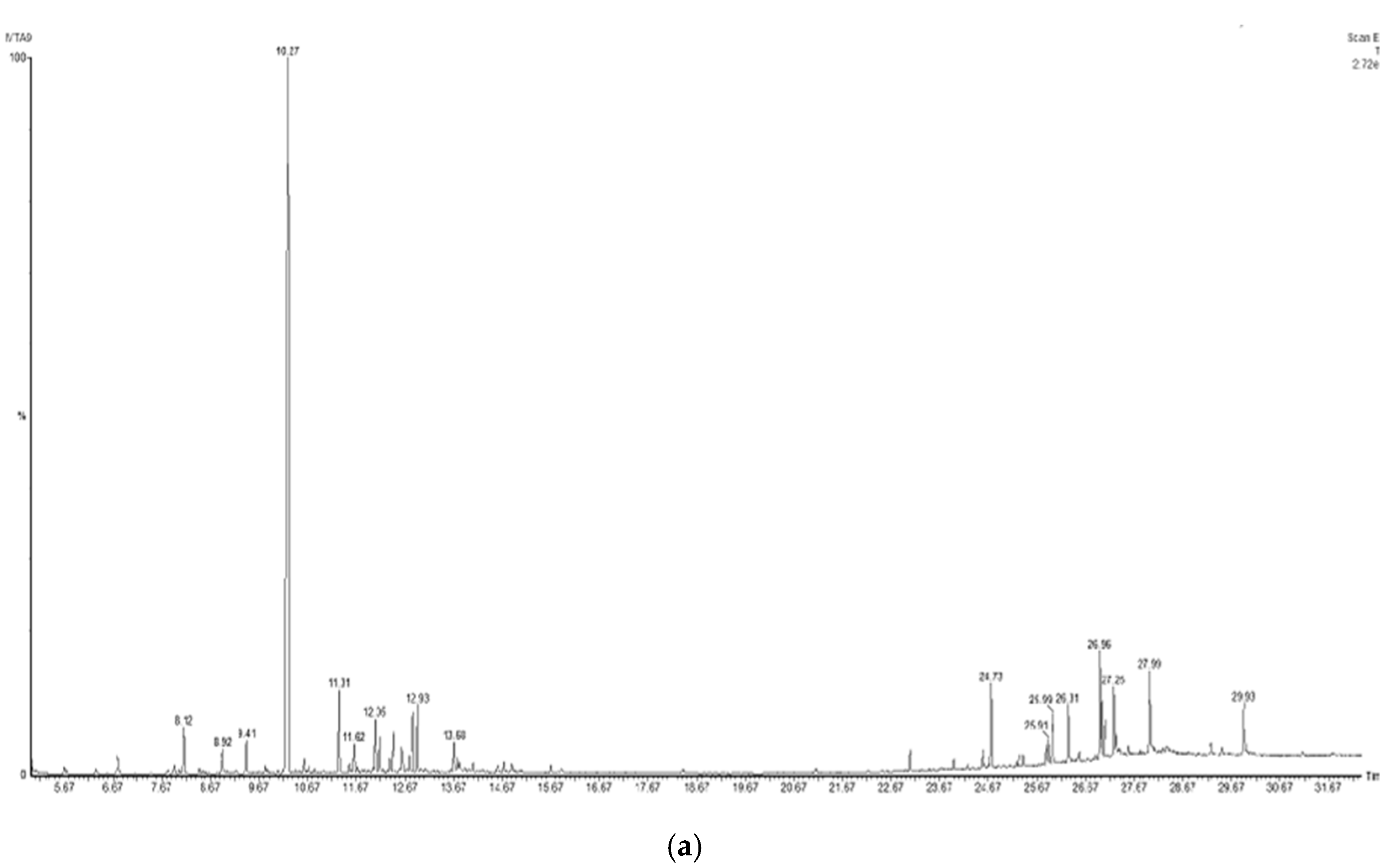
| Plant Extract | Chemical Compound | R.T. | P.A. % | M.W. (g/mol) |
|---|---|---|---|---|
| E. cadamomum (Cardamom) | α-terpinyl acetate | 10.27 | 54.46 | 196.29 |
| 2-((1R,4R)-4-hydroxy-4-methylcyclohex-2-enyl) propan-2-yl acetate | 11.31 | 3.55 | 212.28 | |
| 9-hexacosene | 26.96 | 3.52 | 364.69 | |
| γ-sitoesterol | 29.93 | 3.38 | 414.71 | |
| Heneicosane | 24.73 | 3.04 | 296.57 | |
| 8-acetoxycarvotanacetone | 12.93 | 2.57 | 210.27 | |
| Geranyl oleate | 27.24 | 2.28 | 418.70 | |
| (Z)-3,7-dimethylocta-2,6-dien-1-yl palmitate | 26.31 | 2.08 | 392.70 | |
| Naphthalene, decahydro-4a-methyl-1-methylene-7-(1-methylethenyl)-, [4aR-(4aα,7α,8aα)]- | 12.05 | 2.03 | 204.35 | |
| α-terpineol | 8.12 | 1.95 | 154.25 | |
| Pentacosane | 25.99 | 1.85 | 352.68 | |
| C. longa (Turmeric) | β-turmerone | 15.51 | 33.45 | 218.33 |
| α-turmerone | 15.95 | 21.30 | 218.33 | |
| Ar-turmerone | 15.57 | 19.85 | 216.32 | |
| 16-kauren-19-yl acetate | 27.08 | 18.72 | 330.26 | |
| α-atlantone | 16.80 | 2.77 | 218.33 | |
| β-sesquiphellandrene | 13.84 | 1.98 | 204.35 | |
| Benzene | 13.31 | 1.93 | 78.11 |
| Cytokine | C. longa | E. cardamomum | LPS |
|---|---|---|---|
| IL-4 | 5.07 ± 1.6 | 0.30 ± 0.185 | 0.00 |
| IL-6 | 2.21 ± 0.04 | 1.05 ± 0.012 | 5.61 ± 0.284 |
| IL-10 | 1.84 ± 0.87 | 2.02 ± 0.026 | 0.90 ± 0.073 |
| TNF-α | 1.46 ± 0.97 | 0.086 ± 0.070 | 4.59 ± 2.132 |
| Cell Cytotoxicity Percent (%) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Elettaria cardamomum | Curcuma longa | |||||||
| Concentration µg/mL | HeLa | J774A.1 | Vero E6 | Balb/C Peritoneal Cells | HeLa | J774A.1 | Vero E6 | Balb/C Peritoneal Cells |
| 3.125 | 8.26 a | 4.11 a | 4.14 a | 11.73 a | 2.18 a | 11.74 a | 8.34 a | 2.18 a |
| 6.25 | 10.17 a | 10.28 b | 4.57 a | 14.07 a | 8.23 b | 14.06 a | 10.18 a | 8.23 b |
| 12.5 | 13.03 a | 13.73 b | 8.39 b | 14.33 a | 9.59 b | 14.35 a | 19.03 b | 9.59 b |
| 25 | 13.51 a | 22.26 c | 12.71 b | 16.23 a | 13.85 b | 16.24 a | 21.76 b | 13.85 b |
| 50 | 19.47 b | 31.31 c | 13.04 b | 17.60 a | 14.63 b | 17.62 ab | 22.44 b | 14.63 b |
| 100 | 20.58 b | 37.94 c | 19.54 c | 20.37 ab | 22.16 c | 20.39 ab | 25.81 b | 22.16 c |
| 200 | 21.93 b | 45.33 d | 35.21 d | 27.20 b | 22.92 c | 27.21 b | 26.41 b | 22.92 c |
| EC50 µg/mL | 424.01 | 237.36 | 257.51 | 431.16 | 351.17 | 430.96 | 396.24 | 362.86 |
| LL | 372.09 | 190.79 | 191.02 | 287.35 | 321.20 | 381.60 | 349.78 | 269.66 |
| UL | 473.84 | 261.68 | 301.23 | 582.74 | 382.14 | 480.33 | 442.71 | 509.85 |
| EC90 µg/mL | 890.09 | 423.17 | 436.37 | 838.33 | 721.65 | 957.64 | 931.97 | 705.80 |
| LL | 811.96 | 412.91 | 402.75 | 690.10 | 705.80 | 934.98 | 905.02 | 596.56 |
| UL | 988.53 | 434.32 | 478.67 | 983.15 | 757.78 | 982.90 | 962.03 | 831.34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cárdenas Garza, G.R.; Elizondo Luévano, J.H.; Bazaldúa Rodríguez, A.F.; Chávez Montes, A.; Pérez Hernández, R.A.; Martínez Delgado, A.J.; López Villarreal, S.M.; Rodríguez Rodríguez, J.; Sánchez Casas, R.M.; Castillo Velázquez, U.; et al. Benefits of Cardamom (Elettaria cardamomum (L.) Maton) and Turmeric (Curcuma longa L.) Extracts for Their Applications as Natural Anti-Inflammatory Adjuvants. Plants 2021, 10, 1908. https://doi.org/10.3390/plants10091908
Cárdenas Garza GR, Elizondo Luévano JH, Bazaldúa Rodríguez AF, Chávez Montes A, Pérez Hernández RA, Martínez Delgado AJ, López Villarreal SM, Rodríguez Rodríguez J, Sánchez Casas RM, Castillo Velázquez U, et al. Benefits of Cardamom (Elettaria cardamomum (L.) Maton) and Turmeric (Curcuma longa L.) Extracts for Their Applications as Natural Anti-Inflammatory Adjuvants. Plants. 2021; 10(9):1908. https://doi.org/10.3390/plants10091908
Chicago/Turabian StyleCárdenas Garza, Gustavo R., Joel H. Elizondo Luévano, Aldo F. Bazaldúa Rodríguez, Abelardo Chávez Montes, Raymundo A. Pérez Hernández, Ameyalli J. Martínez Delgado, Sonia M. López Villarreal, José Rodríguez Rodríguez, Rosa M. Sánchez Casas, Uziel Castillo Velázquez, and et al. 2021. "Benefits of Cardamom (Elettaria cardamomum (L.) Maton) and Turmeric (Curcuma longa L.) Extracts for Their Applications as Natural Anti-Inflammatory Adjuvants" Plants 10, no. 9: 1908. https://doi.org/10.3390/plants10091908
APA StyleCárdenas Garza, G. R., Elizondo Luévano, J. H., Bazaldúa Rodríguez, A. F., Chávez Montes, A., Pérez Hernández, R. A., Martínez Delgado, A. J., López Villarreal, S. M., Rodríguez Rodríguez, J., Sánchez Casas, R. M., Castillo Velázquez, U., & Rodríguez Luis, O. E. (2021). Benefits of Cardamom (Elettaria cardamomum (L.) Maton) and Turmeric (Curcuma longa L.) Extracts for Their Applications as Natural Anti-Inflammatory Adjuvants. Plants, 10(9), 1908. https://doi.org/10.3390/plants10091908