
Germination and the Initial Seedling Growth of Lettuce, Celeriac and Wheat Cultivars after Micronutrient and a Biological Application Pre-Sowing Seed Treatment

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results and Discussion
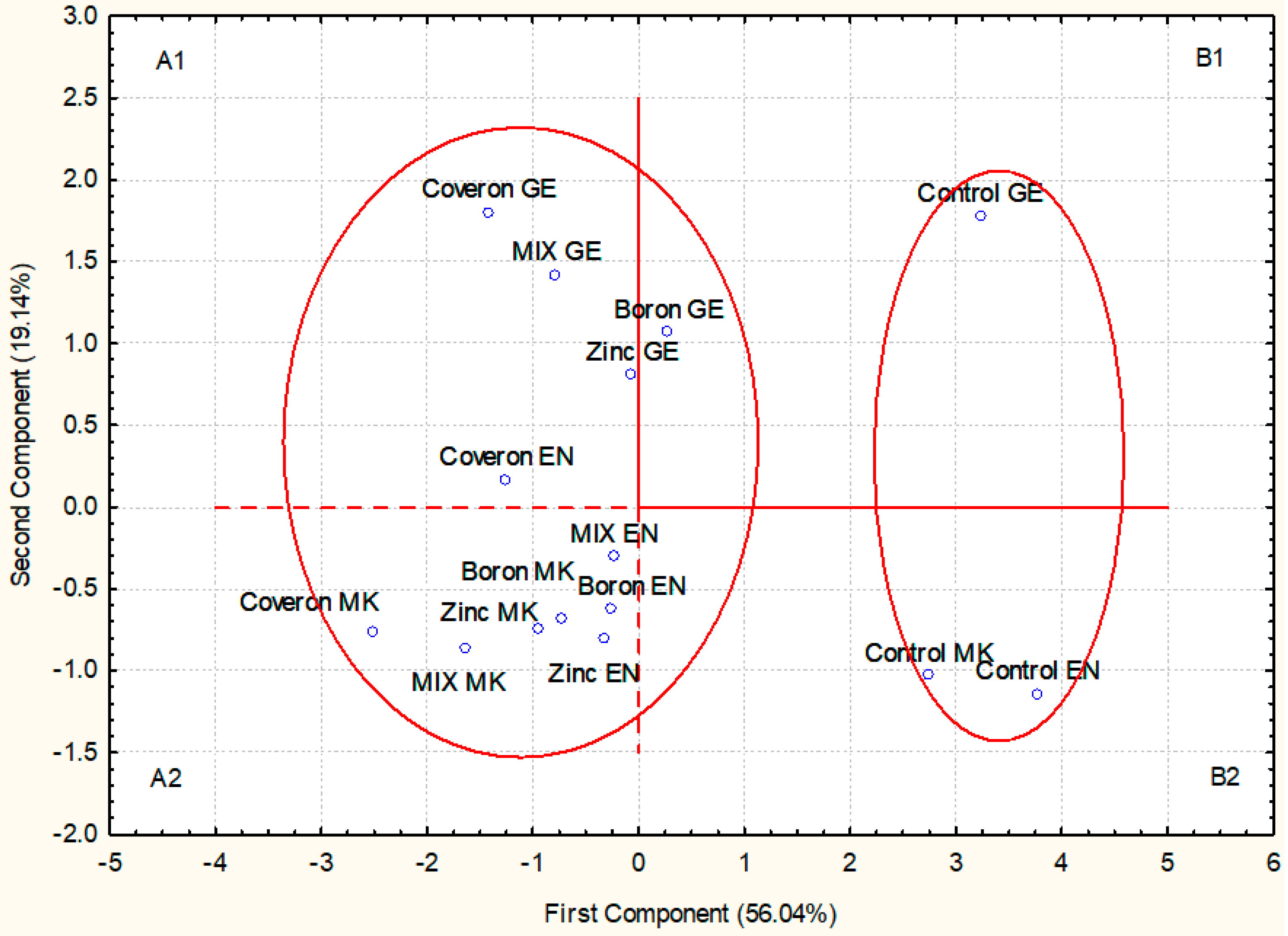
2.1. Lettuce Seed
2.1.1. Germination
2.1.2. Shoot Length
2.1.3. Root Length
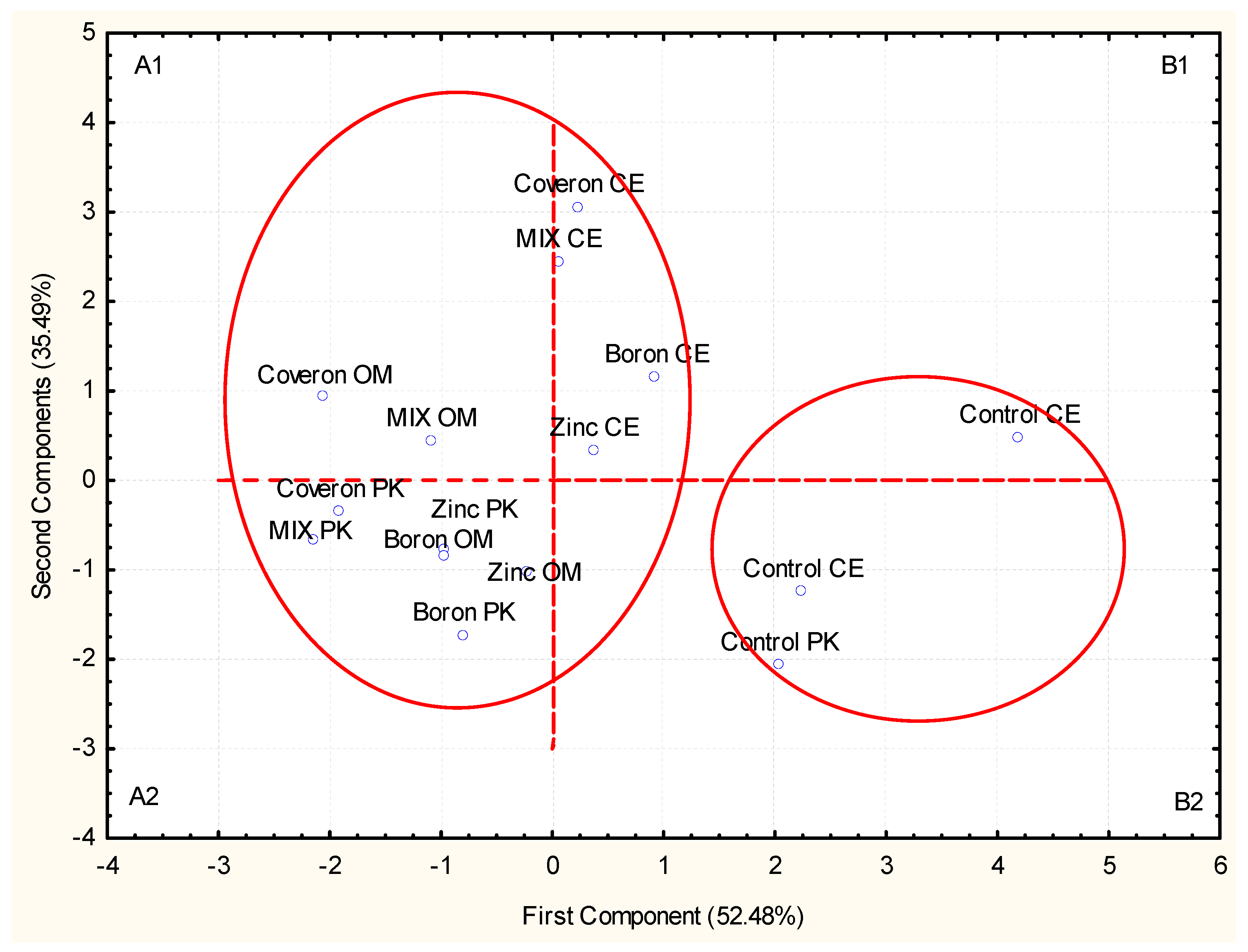
2.2. Celeriac Seed
2.2.1. Germination
2.2.2. Shoot Length
2.2.3. Root Length
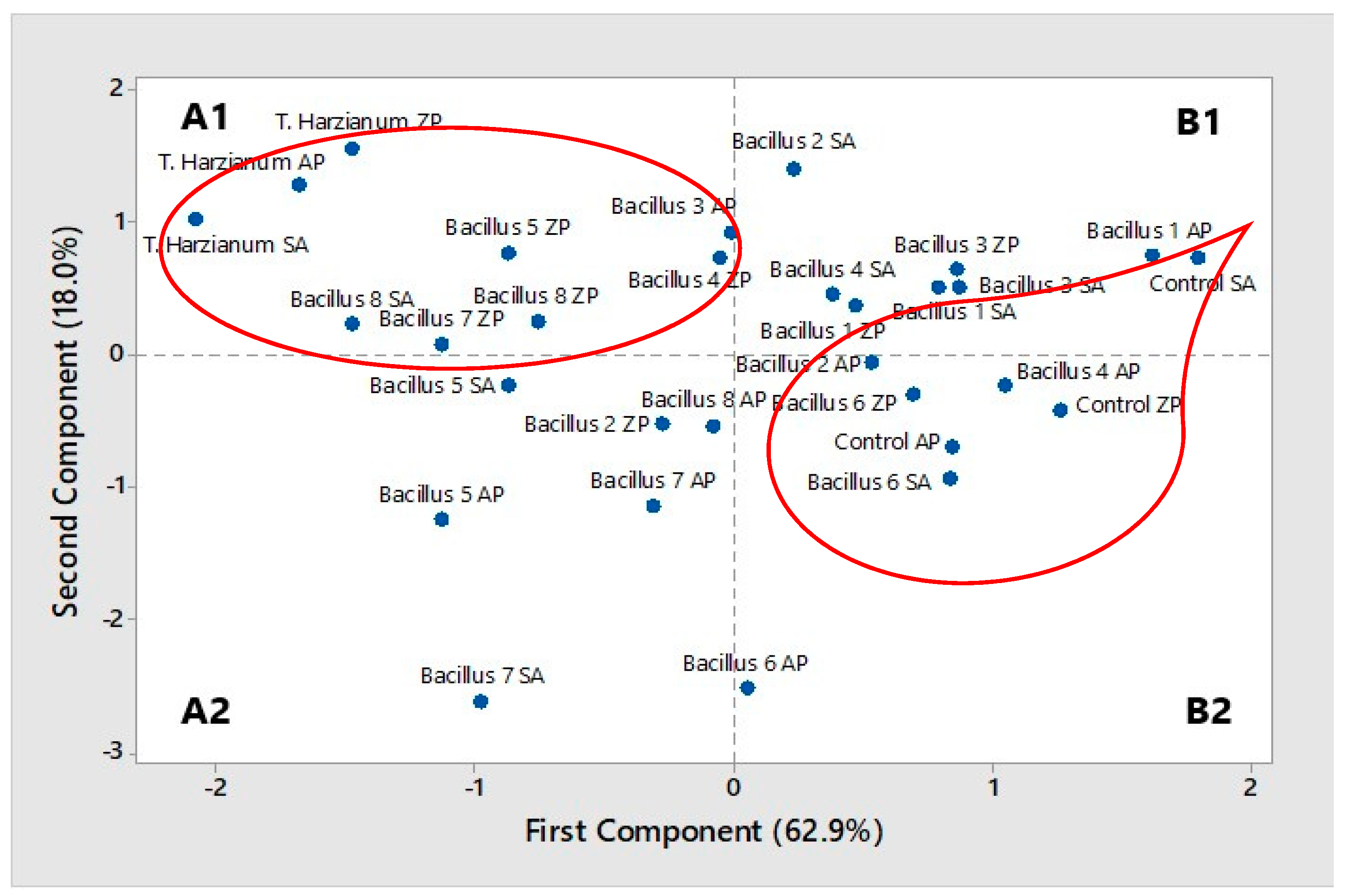
2.3. Wheat Seed
2.3.1. Effects of Factors: Years (Y)/Cultivars (C)/Trichoderma harzianum, Bacillus spp. and Treatment (T)
2.3.2. Germination
2.3.3. Seedling Infection and Growth
2.3.4. Correlation, PCA, and Cluster Analysis
3. Materials and Methods
3.1. Plant Material
3.2. Preparation of Plant Material to Germination
3.3. Laboratory Condition of Germination
3.4. Stress Factors
3.5. Determination of Shoot and Root Length in Laboratory Conditions and Greenhouses
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shatilov, M.; Razin, A.; Ivanova, M. Analysis of the World Lettuce Market. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2019; p. 012053. [Google Scholar]
- Thomas, J.A.; Vasiliev, M.; Alameh, K. Increasing the yield of Lactuca sativa, L. in glass greenhouses through illumination spectral filtering and fevelopment of an optical thin film filter. Sustainability 2020, 12, 3740. [Google Scholar] [CrossRef]
- Sarmanovna, T.Z. Phytochemical study of odorous celery root (Apium graveolens L.) grown in the north caucasus. Pharmacogn. J. 2019, 11, 527–530. [Google Scholar] [CrossRef] [Green Version]
- Aćimović, M.G.; Milić, N.B. Perspectives of the Apiaceae hepatoprotective effects—A review. Nat. Prod. Commun. 2017, 12, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Wahid, A.; Siddique, K.H. Micronutrient application through seed treatments: A review. J. Soil Sci. Plant Nutr. 2012, 12, 125–142. [Google Scholar] [CrossRef]
- Tsonev, T.; Cebola Lidon, F.J. Zinc in plants-an overview. Emir. J. Food Agric. (EJFA) 2012, 24, 322–333. [Google Scholar]
- Tanaka, M.; Fujiwara, T. Physiological roles and transport mechanisms of boron: Perspectives from plants. Pflugers Arch. 2008, 456, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]
- Erdinc, C.; Durak, E.D.; Ekincialp, A.; Şensoy, S.; Demir, S. Variations in response of determinate common bean (Phaseolus vulgaris L.) genotypes to arbuscular mycorrhizal fungi (AMF) inoculation. Turk. J. Agric. For. 2017, 41, 1–9. [Google Scholar] [CrossRef]
- Pathak, N.; Zaidi, R.K. Fungi associated with wheat seed discolouration and abnormalities in in-vitro study. Agric. Sci. 2013, 4, 516–520. [Google Scholar]
- Zafar, M.; Jamal, A.; Tahira, R.; Zakria, M.; Naeemullah, M. Incidence of seed-borne mycoflora in wheat and rice germplasm. Int. J. Agric. Innov. Res. 2014, 2, 720–722. [Google Scholar]
- Wiese, M.V. Compendium of Wheat Diseases; American Phytopathological Society: St. Paul, MN, USA, 1987. [Google Scholar]
- Moura, A.B.; Santos, A.G.; Farias, C.R.J.d.; Ludwig, J.; Corrêa, B.O.; Soares, V.N. Combination of biocontrol bacteria for rice seeds microbiolization: Incidence and transmission of Bipolaris oryzae, and plant growth. J.Seed Sci. 2018, 40, 378–387. [Google Scholar] [CrossRef] [Green Version]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codon, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar]
- Ji, P.; Li, W.; Zheng, Y.; Wang, Z.; Huo, Q.; Hua, C.; Han, C. Isolation and identification of four novel biocontrol Bacillus strains against wheat sharp eyespot and their growth-promoting effect on wheat seedling. Int. J. Agric. Biol. 2019, 21, 282–288. [Google Scholar]
- Zrcková, M.; Capouchová, I.; Paznocht, L.; Eliášová, M.; Dvořák, P.; Konvalina, P.; Janovská, D.; Orsák, M.; Bečková, L. Variation of the total content of polyphenols and phenolic acids in einkorn, emmer, spelt and common wheat grain as a function of genotype, wheat species and crop year. Plant Soil Environ. 2019, 65, 260–266. [Google Scholar] [CrossRef] [Green Version]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Singh, B.; Singh, A.; Singh, B.; Singh, H. Trichoderma harzianum elicits induced resistance in sunflower challenged by Rhizoctonia solani. J. Appl. Microbiol. 2014, 116, 654–666. [Google Scholar] [CrossRef]
- Keswani, C.; Mishra, S.; Sarma, B.K.; Singh, S.P.; Singh, H.B. Unraveling the efficient applications of secondary metabolites of various Trichoderma spp. Appl. Microbiol. Biotechnol. 2014, 98, 533–544. [Google Scholar] [CrossRef]
- Woo, S.L.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef] [Green Version]
- Jain, A.; Singh, A.; Chaudhary, A.; Singh, S.; Singh, H.B. Modulation of nutritional and antioxidant potential of seeds and pericarp of pea pods treated with microbial consortium. Food Res. Int. 2014, 64, 275–282. [Google Scholar] [CrossRef]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Degani, O.; Rabinovitz, O.; Becher, P.; Gordani, A.; Chen, A. Trichoderma longibrachiatum and Trichoderma asperellum confer growth promotion and protection against late wilt disease in the field. J. Fungi 2021, 7, 444. [Google Scholar] [CrossRef]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [Green Version]
- Hampton, J.G.; Boelt, B.; Rolston, M.; Chastain, T. Effects of elevated CO2 and temperature on seed quality. J. Agric. Sci. 2013, 151, 154–162. [Google Scholar] [CrossRef] [Green Version]
- Hampton, J. Producing quality seed: The problem of seed vigour. Curr. Res. Seeds N. Z. 2000, 7, 53–67. [Google Scholar]
- Dornbos, D.L. Production environment and seed quality. In Seed Quality; CRC Press: Boca Raton, FL, USA, 2020; pp. 119–152. [Google Scholar]
- Febri, D.; Anizan, I.; Che, R.; Ahmad, H.S.; Muhammad, H.R.; Wan, M. Enhancement of rice seed germination and vigour by Trichoderma spp. Res. J. Appl. Sci. Eng. Tech. 2014, 7, 4547–4552. [Google Scholar]
- Okoth, S.A.; Otadoh, J.A.; Ochanda, J.O. Improved seedling emergence and growth of maize and beans by Trichoderma harziunum. Trop. Subtrop. Agroecosyst. 2011, 13, 65–71. [Google Scholar]
- Luz, W.C. Effect of bioprotectors on seed pathogens, seed emergence, and corn yield. Fitopatol. Bras. 2001, 26, 16–20. [Google Scholar] [CrossRef] [Green Version]
- Shoresh, M.; Harman, G.E.; Mastouri, F. Induced systemic resistance and plant responses to fungal biocontrol agents. Annu. Rev. Phytopathol. 2010, 48, 21–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, R.G.; de Sá, E.L.S.; Damasceno, R.G.; Hahn, L.; Almeida, D.; Moraes, T.; de Oliveira Camargo, F.A.; Reartes, D.S. Promoção de crescimento de Lotus corniculatus L. e Avena strigosa schreb pela inoculação conjunta de Trichoderma harzianum e rizóbio. Ciênc. Nat. 2011, 33, 111–126. [Google Scholar]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Del-Val, E.; Larsen, J. Ecological functions of Trichoderma spp. and their secondary metabolites in the rhizosphere: Interactions with plants. FEMS Microbiol. Ecol. 2016, 92, fiw036. [Google Scholar] [CrossRef] [Green Version]
- Imran, M.; Neumann, G.; Romheld, V. Nutrient Seed Priming Improves Germination Rate and Seedling Growth under Submergence Stress at Low Temperature. In Competition for Resources in a Changing World: New Drive for Rural Development; University of Hohenheim: Stuttgart, Germany, 2008. [Google Scholar]
- Iqbal, S.; Farooq, M.; Nawaz, A.; Rehman, A. Optimizing boron seed priming treatments for improving the germination and early seedling growth of wheat. J. Agric. Soc. Sci. 2012, 8, 57–61. [Google Scholar]
- Mondal, S.; Bose, B. Impact of micronutrient seed priming on germination, growth, development, nutritional status and yield aspects of plants. J. Plant Nutr. 2019, 42, 2577–2599. [Google Scholar] [CrossRef]
- Aboutalebian, M.; Ekbatani, G.Z.; Sepehri, A. Effects of on-farm seed priming with zinc sulfate and urea solutions on emergence properties, yield and yield components of three rainfed wheat cultivars. Ann. Biol. Res. 2012, 3, 4790–4796. [Google Scholar]
- Foti, R.; Abureni, K.; Tigere, A.; Gotosa, J.; Gere, J. The efficacy of different seed priming osmotica on the establishment of maize (Zea mays L.) caryopses. J. Arid Environ. 2008, 72, 1127–1130. [Google Scholar] [CrossRef]
- Soleimanzadeh, H. Effect of seed priming on germination and yield of corn. Int. J. Agric. Crop Sci. IJACS 2013, 5, 366–369. [Google Scholar]
- Harris, D.; Rashid, A.; Miraj, G.; Arif, M.; Shah, H. ‘On-farm’seed priming with zinc sulphate solution—A cost-effective way to increase the maize yields of resource-poor farmers. Field Crops Res. 2007, 102, 119–127. [Google Scholar] [CrossRef]
- Atique-ur-Rehman; Farooq, M.; Cheema, Z.A.; Wahid, A. Role of boron in leaf elongation and tillering dynamics in fine-grain aromatic rice. J. Plant Nutr. 2013, 36, 42–54. [Google Scholar] [CrossRef]
- Afrayeem, S.M.; Chaurasia, A. Effect of zinc oxide nanoparticles on seed germination and seed vigour in chilli (Capsicum annuum L.). J. Pharmacogn. Phytochem. 2017, 6, 1564–1566. [Google Scholar]
- Poštić, D.; Štrbanović, R.; Stanojković-Sebić, A.; Tabaković, M.; Milivojević, M.; Jovanović, S.; Stanisavljević, R. Increasing the pepper seed quality using mycorrhizal fungi. J. Process. Energy Agric. 2019, 23, 66–68. [Google Scholar] [CrossRef]
- Štrbanović, R.; Poštić, D.; Tabaković, M.; Knežević, J.; Živanović, L.; Stanisavljević, R. Effects of pre-sowing seed treatments for improving germination and the growth of pepper and tomato seedlings. Acta Sci. Pol. Hortorum Cultus 2021, 20, 101–109. [Google Scholar] [CrossRef]
- Tavares, L.C.; Fonseca, D.Â.R.; Brunes, A.P.; Rufino, C.d.A.; Meneguello, G.E.; Barros, A.C.S.A. Performance of rice seeds treated with zinc, boron, and molybdenum. J. Seed Sci. 2013, 35, 532–539. [Google Scholar] [CrossRef] [Green Version]
- Freiberg, J.A.; Ludwig, M.P.; Avelar, S.A.G.; Girotto, E. Seed treatment and its impact on wheat crop yield potential. J. Seed Sci. 2017, 39, 280–287. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, J.B.; Muniz, P.; Peixoto, G.H.S.; de Oliveira, T.A.; Duarte, E.A.; Rodrigues, F.; Carvalho, D.D.C. Promotion of seedling growth and production of wheat by using Trichoderma spp. J. Agric. Sci. 2018, 10, 267–276. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Roldán, A.; Albacete, A.; Pascual, J.A. The interaction with arbuscular mycorrhizal fungi or Trichoderma harzianum alters the shoot hormonal profile in melon plants. Phytochemistry 2011, 72, 223–229. [Google Scholar] [CrossRef]
- Sofo, A.; Scopa, A.; Manfra, M.; De Nisco, M.; Tenore, G.; Troisi, J.; Di Fiori, R.; Novellino, E. Trichoderma harzianum strain T-22 induces changes in phytohormone levels in cherry rootstocks (Prunus cerasus × P. canescens). Plant Growth Regul. 2011, 65, 421–425. [Google Scholar] [CrossRef] [Green Version]
- Harman, G.E. Myths and dogmas of biocontrol changes in perceptions derived from research on Trichoderma harzinum T-22. Plant Dis. 2000, 84, 377–393. [Google Scholar] [CrossRef] [Green Version]
- Hoyos-Carvajal, L.; Orduz, S.; Bissett, J. Growth stimulation in bean (Phaseolus vulgaris L.) by Trichoderma. Biol. Control 2009, 51, 409–416. [Google Scholar] [CrossRef]
- Feledyn-Szewczyk, B.; Cacak-Pietrzak, G.; Lenc, L.; Stalenga, J. Rating of spring wheat varieties (Triticum aestivum L.) according to their suitability for organic agriculture. Agronomy 2020, 10, 1900. [Google Scholar] [CrossRef]
- Banjac, N.; Stanisavljević, R.; Dimkić, I.; Velijević, N.; Soković, M.; Ćirić, A. Trichoderma harzianum IS005–12 promotes germination, seedling growth and seedborne fungi suppression in Italian ryegrass forage. Plant Soil Environ. 2021, 67, 130–136. [Google Scholar] [CrossRef]
- Gravel, V.; Antoun, H.; Tweddell, R.J. Growth stimulation and fruit yield improvement of greenhouse tomato plants by inoculation with Pseudomonas putida or Trichoderma atroviride: Possible role of indole acetic acid (IAA). Soil Biol. Biochem. 2007, 39, 1968–1977. [Google Scholar] [CrossRef]
- Silva, F.F.d.; Castro, E.d.M.; Moreira, S.I.; Ferreira, T.C.; Lima, A.E.d.; Alves, E. Emergência e análise ultraestrutural de plântulas de soja inoculadas com Sclerotinia sclerotiorum sob efeito da aplicação de Trichoderma harzianum. Summa Phytopathol. 2017, 43, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Shukla, N.; Awasthi, R.; Rawat, L.; Kumar, J. Seed biopriming with drought tolerant isolates of Trichoderma harzianum promote growth and drought tolerance in Triticum aestivum. Ann. Appl. Biol. 2015, 166, 171–182. [Google Scholar] [CrossRef]
- Wang, J.; Hasanalieva, G.; Wood, L.; Anagnostopoulos, C.; Ampadogiannis, G.; Bempelou, E.; Kiousi, M.; Markellou, E.; Iversen, P.O.; Seal, C.; et al. Effect of wheat species (Triticum aestivum vs. T. spelta), farming system (organic vs. conventional) and flour type (whole grain vs. white) on composition of wheat flour–results of a retail survey in the UK and Germany–3. Pesticide residue content. Food Chem. X 2020, 7, 100089. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Baranski, M.; Korkut, R.; Kalee, H.A.; Wood, L.; Bilsborrow, P.; Janovska, D.; Leifert, A.; Winter, S.; Willson, A.; et al. Performance of modern and traditional spelt wheat (Triticum spelta) varieties in rain-fed and irrigated, organic and conventional production systems in a semi-arid environment; results from exploratory field experiments in Crete, Greece. Agronomy 2021, 11, 890. [Google Scholar] [CrossRef]
- Guardado-Valdivia, L.; Tovar-Pérez, E.; Chacón-López, A.; López-García, U.; Gutiérrez-Martínez, P.; Stoll, A.; Aguilera, S. Identification and characterization of a new Bacillus atrophaeus strain B5 as biocontrol agent of postharvest anthracnose disease in soursop (Annona muricata) and avocado (Persea americana). Microbiol. Res. 2018, 210, 26–32. [Google Scholar] [CrossRef]
- Abdel-Monaim, M.F.; Abdel-Gaid, M.A.; Zayan, S.A.; Nassef, D.M. Enhancement of growth parameters and yield components in eggplant using antagonism of Trichoderma spp. against Fusarium wilt disease. Int. J. Phytopathol. 2014, 3, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Mukhtar, I.; Hannan, A.; Atiq, M.; Nawaz, A. Impact of Trichoderma species on seed germination in soybean. Pak. J. Phytopathol. 2012, 24, 159–162. [Google Scholar]
- Nikolić, I.; Berić, T.; Dimkić, I.; Popović, T.; Lozo, J.; Fira, D.; Stanković, S. Biological control of Pseudomonas syringae pv. aptata on sugar beet with Bacillus pumilus SS-10.7 and Bacillus amyloliquefaciens (SS-12.6 and SS-38.4) strains. J. Appl. Microbiol. 2019, 126, 165–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Seed Testing Association (ISTA). Rules for Testing Seeds; ISTA: Zurich, Switzerland, 2020. [Google Scholar]
- Stanisavljevic, R.; Ðjokic, D.; Milenkovic, J.; Ðukanovic, L.; Stevovic, V.; Simic, A.; Dodig, D. Seed germination and seedling vigour of Italian ryegrass, cocksfoot and timothy following harvest and storage. Cienc. Agrotecnol. 2011, 35, 1141–1148. [Google Scholar] [CrossRef]
- Gianinetti, A. Basic features of the analysis of germination data with generalized linear mixed models. Data 2020, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R-Statistics, a Language and Environment for Statistical Computing (Version 3.4.4); R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Minitab Inc. Version 16.1.0. State College, Pennsylvania, USA, Was Used to Process Data (Free Version). Available online: https://www.minitab.com/en-us/ (accessed on 17 November 2020).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Results Obtained in Germination Cabinets (a) | ||||||
|---|---|---|---|---|---|---|
| Traits | Treatment | Cultivar | Average | CV (%) | ||
| Genius | Majska Kraljica | Endivia | ||||
| Germination | ||||||
| (%) | Control | 88 ± 0.58 b | 86 ± 0.19 b | 86 ± 0.12 b | 87 | 1.33 |
| Zinc | 89 ± 0.33 ab | 89 ± 0.33 ab | 91 ± 0.18 a | 90 | 1.29 | |
| Boron | 89 ± 0.28 ab | 88 ± 0.48 ab | 91 ± 0.42 a | 89 | 1.71 | |
| MIX | 91 ± 0.72 a | 90 ± 0.63 a | 90 ± 0.60 a | 90 | 0.64 | |
| Coveron | 92 ± 0.52 a | 91 ± 0.51 a | 89 ± 0.50 ab | 91 | 1.68 | |
| Average | 89 | 88 | 90 | - | - | |
| CV % | 1.41 | 1.94 | 2.66 | - | - | |
| Results Obtained in Greenhouses (b) | ||||||
| Control | 74 ± 0.54 b | 79 ± 0.12 c | 83 ± 0.58 b | 79 | 5.73 | |
| Zinc | 87 ± 0.63 a | 95 ± 0.33 a | 89 ± 0.58 a | 90 | 4.61 | |
| Boron | 88 ± 0.81 a | 94 ± 0.58 a | 90 ± 0.58 a | 91 | 3.37 | |
| MIX | 90 ± 0.45 a | 93 ± 0.50 ab | 88 ± 0.58 a | 90 | 2.79 | |
| Coveron | 88 ± 0.16 a | 91 ± 0.76 b | 89 ± 0.58 a | 89 | 1.71 | |
| Average | 85 | 90 | 85 | - | - | |
| CV % | 7.57 | 7.24 | 4.53 | - | - | |
| Results Obtained in Germination Cabinets (a) | ||||||
|---|---|---|---|---|---|---|
| Traits | Treatment | Cultivar | Average | CV (%) | ||
| Genius | Majska Kraljica | Endivia | ||||
| Shoot length | ||||||
| (cm) | Control | 4.5 ± 0.16 b | 3.7 ± 0.11 b | 3.1 ± 0.13 b | 3.8 | 18.6 |
| Zinc | 4.6 ± 0.28 ab | 4.2 ± 0.55 a | 3.7 ± 0.16 a | 4.2 | 10.8 | |
| Boron | 4.7 ± 0.58 ab | 4.3 ± 0.68 a | 3.8 ± 0.72 a | 4.3 | 10.6 | |
| MIX | 4.8 ± 0.62 a | 4.1 ± 0.19 a | 3.6 ± 0.55 ab | 4.2 | 14.5 | |
| Coveron | 5.0 ± 0.50 a | 4.2 ± 0.38 a | 4.3 ± 0.14 a | 4.5 | 9.68 | |
| Average | 4.7 | 4.1 | 3.7 | - | - | |
| CV % | 4.08 | 5.72 | 11.6 | - | - | |
| Results Obtained in Greenhouses (b) | ||||||
| Control | 5.9 ± 0.76 b | 5.5 ± 0.14 b | 5.6 ± 0.50 c | 5.7 | 3.67 | |
| Zinc | 7.2 ± 0.32 a | 6.9 ± 0.59 a | 6.6 ± 0.14 b | 6.9 | 4.35 | |
| Boron | 6.9 ± 0.12 a | 7.0 ± 0.12 a | 6.5 ± 0.16 b | 6.8 | 3.89 | |
| MIX | 7.3 ± 0.45 a | 7.3 ± 0.14 a | 7.8 ± 0.25 a | 7.5 | 3.87 | |
| Coveron | 7.6 ± 0.65 a | 7.7 ± 0.62 a | 8.1 ± 0.36 a | 7.8 | 3.39 | |
| Average | 7.0 | 6.9 | 6.9 | - | - | |
| CV % | 9.36 | 12.1 | 14.8 | - | - | |
| Results Obtained in Germination Cabinets (a) | ||||||
|---|---|---|---|---|---|---|
| Traits | Treatment | Cultivar | Average | CV (%) | ||
| Genius | Majska Kraljica | Endivia | ||||
| Root length | ||||||
| (cm) | Control | 1.1 ± 0.11 b | 1.8 ± 0.72 b | 0.6 ± 0.42 b | 1.2 | 51.7 |
| Zinc | 1.2 ± 0.56 ab | 2.1 ± 0.58 ab | 1.3 ± 0.55 a | 1.5 | 32.2 | |
| Boron | 1.3 ± 0.12 ab | 2.1 ± 0.14 ab | 1.0 ± 0.18 ab | 1.5 | 38.8 | |
| MIX | 1.5 ± 0.62 a | 2.2 ± 0.33 a | 1.1 ± 0.14 ab | 1.6 | 34.8 | |
| Coveron | 1.4 ± 0.50 a | 2.4 ± 0.51 a | 1.4 ± 0.56 a | 1.7 | 33.3 | |
| Average | 1.3 | 2.1 | 1.1 | - | - | |
| CV % | 12.2 | 10.2 | 28.8 | - | - | |
| Results Obtained in Greenhouses (b) | ||||||
| Control | 0.4 ± 0.48 b | 0.6 ± 0.50 b | 0.5 ± 0.77 b | 0.5 | 20.0 | |
| Zinc | 0.7 ± 0.58 a | 0.7 ± 0.66 ab | 0.8 ± 0.65 a | 0.7 | 7.87 | |
| Boron | 0.6 ± 0.52 a | 0.7 ± 0.17 ab | 0.8 ± 0.14 a | 0.7 | 14.3 | |
| MIX | 0.6 ± 0.13 a | 0.8 ± 0.53 a | 0.7 ± 0.18 a | 0.7 | 14.3 | |
| Coveron | 0.7 ± 0.55 a | 0.9 ± 0.88 a | 0.7 ± 0.22 a | 0.8 | 15.1 | |
| Average | 0.6 | 0.7 | 0.7 | - | - | |
| CV % | 20.4 | 15.4 | 17.7 | - | - | |
| Results Obtained in Germination Cabinets (F-Test) | |||
|---|---|---|---|
| Source | Genius | Majska Kraljica | Endivia |
| Cultivar (A) | ** | * | * |
| Treatment (B) | ** | * | * |
| Location (C) | ns | ns | ns |
| Year (D) | ns | ns | ns |
| A × B | * | * | * |
| A × C | ns | ns | ns |
| A × D | ns | ns | ns |
| B × C | ns | ns | ns |
| B × D | ns | ns | ns |
| C × D | ns | ns | ns |
| Results Obtained in Greenhouses (F-Test) | |||
| Cultivar (A) | * | * | * |
| Treatment (B) | ** | * | * |
| Location (C) | ns | ns | ns |
| Year (D) | ns | ns | ns |
| A × B | * | * | * |
| A × C | ns | ns | ns |
| A × D | ns | ns | ns |
| B × C | ns | ns | ns |
| B × D | ns | ns | ns |
| C × D | ns | ns | ns |
| Results Obtained in Germination Cabinets (a) | ||||||
|---|---|---|---|---|---|---|
| Traits | Treatment | Cultivar | Average | CV (%) | ||
| Cezar | Omega | Praški Krupni | ||||
| Germination | ||||||
| (%) | Control | 64 ± 0.13 c | 79 ± 0.54 b | 80 ± 0.50 b | 74 | 12.1 |
| Zinc | 87 ± 0.28 a | 86 ± 0.28 a | 92 ± 0.45 a | 88 | 3.64 | |
| Boron | 79 ± 0.58 ab | 89 ± 0.34 a | 95 ± 0.84 a | 88 | 9.22 | |
| MIX | 82 ± 0.92 a | 86 ± 0.22 a | 95 ± 0.67 a | 88 | 7.60 | |
| Coveron | 78 ± 0.19 b | 87 ± 0.77 a | 95 ± 0.55 a | 87 | 9.81 | |
| Average | 78 | 85 | 91 a | - | - | |
| CV % | 11.0 | 4.43 | 7.12 | - | - | |
| Results Obtained in Greenhouses (b) | ||||||
| Control | 57 ± 0.19 c | 69 ± 0.10 b | 79 ± 0.41 c | 68 | 16.1 | |
| Zinc | 82 ± 0.11 a | 81 ± 0.18 a | 87 ± 0.63 b | 83 | 3.86 | |
| Boron | 72 ± 0.71 b | 84 ± 0.84 a | 91 ± 0.17 a | 82 | 11.7 | |
| MIX | 72 ± 0.63 b | 79 ± 0.52 ab | 86 ± 0.63 b | 79 | 8.86 | |
| Coveron | 69 ± 0.54 b | 82 ± 0.32 a | 90 ± 0.48 a | 80 | 13.2 | |
| Average | 70 | 79 | 87 | - | - | |
| CV % | 12.7 | 7.44 | 5.45 | - | - | |
| Results Obtained in Germination Cabinets (a) | ||||||
|---|---|---|---|---|---|---|
| Traits | Treatment | Cultivar | Average | CV (%) | ||
| Cezar | Omega | Praški krupni | ||||
| Shoot length | ||||||
| (cm) | Control | 1.0 ± 0.63 b | 1.8 ± 0.55 b | 1.8 ± 0.44 c | 1.5 | 30.1 |
| Zinc | 1.2 ± 0.18 a | 2.5 ± 0.63 a | 2.3 ± 0.56 b | 2.0 | 35.0 | |
| Boron | 1.3 ± 0.77 a | 2.4 ± 0.98 a | 2.2 ± 0.25 b | 2.0 | 29.8 | |
| MIX | 1.3 ± 0.28 a | 2.3 ± 0.13 a | 2.7 ± 0.15 a | 2.1 | 34.3 | |
| Coveron | 1.2 ± 0.14 a | 2.6 ± 0.42 a | 2.8 ± 0.76 a | 2.2 | 39.6 | |
| Average | 1.2 | 2.3 | 2.4 | - | - | |
| CV % | 10.2 | 13.4 | 17.1 | - | - | |
| Results Obtained in Greenhouses (b) | ||||||
| Control | 1.7 ± 0.67 c | 1.3 ± 0.09 c | 1.2 ± 0.72 b | 1.4 | 18.9 | |
| Zinc | 2.4 ± 0.53 b | 1.8 ± 0.17 b | 2.0 ± 0.67 a | 2.1 | 14.8 | |
| Boron | 2.3 ± 0.45 b | 1.8 ± 0.54 b | 1.8 ± 0.14 a | 2.0 | 14.7 | |
| MIX | 3.0 ± 0.62 a | 2.1 ± 0.82 ab | 1.9 ± 0.17 a | 2.3 | 25.1 | |
| Coveron | 3.1 ± 0.13 a | 2.4 ± 0.43 a | 2.0 ± 0.43 a | 2.5 | 22.3 | |
| Average | 2.5 | 1.9 | 1.8 | - | - | |
| CV % | 22.8 | 21.7 | 18.8 | - | - | |
| Results Obtained in Germination Cabinets (a) | ||||||
|---|---|---|---|---|---|---|
| Traits | Treatment | Cultivar | Average | CV (%) | ||
| Cezar | Omega | Praški krupni | ||||
| Root length | ||||||
| (cm) | Control | 1.2 ± 0.43 b | 1.1 ± 0.63 b | 1.0 ± 0.64 c | 1.1 | 9.09 |
| Zinc | 1.4 ± 0.16 ab | 1.4 ± 0.78 ab | 1.3 ± 0.53 b | 1.4 | 4.22 | |
| Boron | 1.5 ± 0.75 a | 1.6 ± 0.25 a | 1.3 ± 0.17 b | 1.5 | 10.4 | |
| MIX | 1.5 ± 0.19 a | 1.7 ± 0.65 a | 1.8 ± 0.13 a | 1.7 | 9.17 | |
| Coveron | 1.6 ± 0.55 a | 1.8 ± 0.13 a | 1.7 ± 0.26 a | 1.7 | 5.88 | |
| Average | 1.4 | 1.5 | 1.4 | - | - | |
| CV % | 10.5 | 18.3 | 23.0 | - | - | |
| Results Obtained in Greenhouses (b) | ||||||
| Control | 2.6 ± 0.38 c | 2.9 ± 0.19 c | 2.7 ± 0.11 c | 2.7 | 5.59 | |
| Zinc | 3.3 ± 0.56 b | 3.3 ± 0.11 b | 3.8 ± 0.55 a | 3.5 | 8.33 | |
| Boron | 3.6 ± 0.13 ab | 3.5 ± 0.77 b | 3.2 ± 0.61 b | 3.4 | 6.06 | |
| MIX | 4.3 ± 0.10 a | 4.0 ± 0.39 ab | 3.7 ± 0.13 a | 4.0 | 7.50 | |
| Coveron | 4.4 ± 0.17 a | 4.4 ± 0.46 a | 3.9 ± 0.52 a | 4.2 | 6.82 | |
| Average | 3.6 | 3.6 | 3.5 | - | - | |
| CV % | 20.4 | 16.3 | 14.5 | - | - | |
| Results Obtained in Germination Cabinets (F-Test) | |||
|---|---|---|---|
| Source | Cezar | Omega | Praški krupni |
| Cultivar (A) | * | * | * |
| Treatment (B) | ** | * | * |
| Location (C) | ns | ns | ns |
| Year (D) | ns | ns | ns |
| A × B | * | * | * |
| A × C | ns | ns | ns |
| A × D | ns | ns | ns |
| B × C | ns | ns | ns |
| B × D | ns | ns | ns |
| C × D | ns | ns | ns |
| Results Obtained in Greenhouses (F-Test) | |||
| Cultivar (A) | * | * | * |
| Treatment (B) | ** | * | * |
| Location (C) | ns | ns | ns |
| Year (D) | ns | ns | ns |
| A × B | * | * | * |
| A × C | ns | ns | ns |
| A × D | ns | ns | ns |
| B × C | ns | ns | ns |
| B × D | ns | ns | ns |
| C × D | ns | ns | ns |
| Factor | Feature | ||||||
|---|---|---|---|---|---|---|---|
| G | F | A | P | M | SL | RL | |
| D | ns | ns | ns | ns | ns | ns | ns |
| A | * | ** | ** | ** | ** | * | * |
| B | *** | *** | *** | *** | *** | * | * |
| D × B | ns | ns | ns | ns | ns | ns | ns |
| D × A | ns | ns | ns | ns | ns | ns | ns |
| A × B | * | * | * | ** | ** | * | * |
| C | Treatment | F | A | P | M | G | SL | RL |
|---|---|---|---|---|---|---|---|---|
| Apicol | Control | 4 ± 0.96 b | 4 ± 0.00 a | 1 ± 0.58 a | 1 ±0.00 ab | 86 ± 0.48 b | 6.01 ± 0.31 b | 10.6 ± 0.95 b |
| Trichoderma harzianum | 1 ± 0.58 c | 1 ± 0.58 b | 0 ± 0.00 b | 0 ± 0.00 b | 90 ± 0.29 a | 7.56 ± 0.33 a | 13.4 ± 0.66 a | |
| Bacillus 1 | 4 ± 0.96 b | 3 ±0.58 ab | 0 ± 0.50 b | 0 ± 0.50 b | 83 ± 0.31 b | 6.03 ± 0.81 b | 11.1 ± 0.75 ab | |
| Bacillus 2 | 4 ± 0.82 b | 3 ±0.82 ab | 0 ± 0.00 b | 0 ± 0.00 b | 86 ± 0.19 b | 6.52 ± 0.48 ab | 11.4 ± 0.89 ab | |
| Bacillus 3 | 2 ± 0.96 c | 2 ± 0.96 b | 1 ± 0.00 a | 1 ± 0.58 ab | 87 ± 0.18 ab | 6.67 ±0.51 ab | 11.2 ± 0.88 ab | |
| Bacillus 4 | 4 ± 0.96 b | 4 ± 0.82 a | 0 ± 0.50 b | 0 ± 0.00 b | 85 ± 0.09 b | 6.56 ± 0.67 ab | 10.8 ± 0.81 b | |
| Bacillus 5 | 5 ± 0.78 ab | 2 ± 0.96 b | 0 ± 0.00 b | 0 ± 0.00 b | 90 ± 0.22 a | 7.52 ± 0.51 a | 12.8 ± 0.55 a | |
| Bacillus 6 | 6 ± 0.65 a | 4 ± 0.50 a | 0 ± 0.00 b | 2 ± 0.96 a | 88 ± 0.36 ab | 6.99 ± 0.80 ab | 11.9 ± 0.46 ab | |
| Bacillus 7 | 3 ± 0.82 bc | 4 ± 0.50 a | 0 ± 0.50 b | 3 ± 0.58 a | 88 ± 0.21 ab | 7.03 ± 0.71 a | 12.6 ± 0.81 a | |
| Bacillus 8 | 4 ± 0.00 b | 3 ± 0.58 ab | 1 ± 0.58 a | 1 ± 0.58 ab | 87 ± 0.36 ab | 6.98 ± 0.55 a | 12.9 ± 0.92 a | |
| Salazar | Control | 4 ± 0.96 b | 3 ± 0.00 ab | 0 ± 0.00 b | 0 ± 0.00 b | 83 ± 0.26 c | 6.06 ± 0.52 b | 10.1 ± 0.86 b |
| Trichoderma harziaum | 1 ± 0.58 c | 1 ± 0.00 b | 0 ± 0.00 b | 0 ± 0.00 b | 91 ± 0.26 a | 7.77 ± 0.44 a | 13.7 ± 0.38 a | |
| Bacillus 1 | 5 ± 0.29 ab | 1 ± 0.58 b | 0 ± 0.00 b | 0 ± 0.00 b | 85 ± 0.38 bc | 6.65 ± 0.55 ab | 11.2 ± 1.08 ab | |
| Bacillus 2 | 2 ± 0.00 c | 2 ± 0.00 b | 0 ± 0.00 b | 0 ± 0.00 b | 86 ± 0.22 b | 6.72 ± 031 ab | 11.5 ± 0.55 ab | |
| Bacillus 3 | 3 ± 0.82 bc | 3 ± 0.58 ab | 0 ± 0.50 b | 1 ± 0.00 ab | 85 ± 0.26 bc | 6.69 ± 0.48 ab | 10.9 ± 0.28 ab | |
| Bacillus 4 | 3 ± 0.82 bc | 3 ± 0.58 ab | 1 ± 0.00 a | 0 ± 0.50 b | 86 ± 0.38 b | 6.71 ± 0.68 ab | 11.7 ± 0.77 ab | |
| Bacillus 5 | 2 ± 0.96 c | 4 ± 0.00 a | 0 ± 0.00 b | 0 ± 0.00 b | 89 ± 0.17 ab | 7.21 ± 0.78 a | 12.9 ± 0.81 a | |
| Bacillus 6 | 4 ± 0.82 b | 4 ± 0.96 a | 0 ± 0.50 b | 2 ± 0.00 a | 86 ± 0.32 b | 6.66 ± 0.66 ab | 10.4 ± 0.89 b | |
| Bacillus 7 | 6 ± 0.46 a | 4 ± 0.82 a | 0 ± 0.50 b | 0 ± 0.00 b | 90 ± 0.24 a | 7.71 ± 0.51 a | 13.3 ± 0.98 a | |
| Bacillus 8 | 3 ± 0.00 bc | 1 ± 0.58 b | 1 ± 0.58 a | 0 ± 0.00 b | 90 ± 0.22 a | 7.68 ± 0.63 a | 13.2 ± 0.75 a | |
| Zemunska Rosa | Control | 4 ± 0.96 a | 4 ± 0.96 a | 1 ± 0.58 a | 1 ± 0.00 ab | 85 ± 0.32 c | 5.59 ± 0.67 b | 10.3 ± 0.62 b |
| Trichoderma harzianum | 1 ± 0.00 c | 1 ± 0.00 b | 0 ± 0.00 b | 0 ± 0.00 b | 89 ± 0.41 a | 7.59 ± 0.28 a | 14.1 ± 0.46 a | |
| Bacillus 1 | 4 ± 0.96 a | 2 ± 0.00 b | 0 ± 0.00 b | 0 ± 0.00 b | 86 ± 0.32 bc | 6.35 ± 0.31 ab | 11.3 ± 0.35 ab | |
| Bacillus 2 | 3 ± 0.82 ab | 4 ± 0.50 a | 0 ± 0.50 b | 0 ± 0.00 b | 88 ± 0.26 ab | 7.55 ± 0.52 ab | 11.7 ± 0.68 ab | |
| Bacillus 3 | 4 ± 0.00 a | 2 ± 0.96 b | 0 ± 0.00 b | 0 ± 0.00 b | 85 ± 0.31 c | 6.08 ± 0.83 b | 11.1 ± 0.70 ab | |
| Bacillus 4 | 2 ± 0.96 b | 3 ± 0.58 ab | 0 ± 0.00 b | 0 ± 0.50 b | 87 ± 0.36 b | 6.62 ± 0.74 ab | 11.8 ± 0.68 ab | |
| Bacillus 5 | 3 ± 0.82 ab | 1 ± 0.58 b | 0 ± 0.50 b | 0 ± 0.50 b | 88 ± 0.48 ab | 7.58 ± 0.51 a | 13.5 ± 0.87 a | |
| Bacillus 6 | 4 ± 0.96 a | 3 ± 0.82 ab | 0 ± 0.00 b | 1 ± 0.58 ab | 86 ± 0.31 bc | 6.59 ± 0.43 ab | 10.6 ± 0.53 b | |
| Bacillus 7 | 3 ± 0.00 ab | 2 ± 0.96 b | 0 ± 0.00 b | 0 ± 0.00 b | 89 ± 0.17 a | 7.62 ± 0.49 a | 13.6 ± 0.88 a | |
| Bacillus 8 | 4 ± 0.82 a | 1 ± 0.00 b | 0 ± 0.50 b | 0 ± 0.00 b | 88 ± 0.21 ab | 7.53 ± 0.82 a | 13.4 ± 0.81 a |
| Cultivar | Examined Feature | F | A | P | M | PSL | PRL |
|---|---|---|---|---|---|---|---|
| Apicol | Germination | −0.181 ns | −0.488 ns | −0.106 ns | −0.199 ns | 0.911 *** | 0.785 ** |
| Fusarium sp. | - | 0.595 ns | −0.178 ns | 0.106 ns | −0.174 ns | −0.522 ns | |
| Aternaria sp. | - | 0.000 ns | 0.510 ns | −0.535 ns | −0.522 ns | ||
| Penicillium sp. | - | 0.134 ns | −0.301 ns | −0.211 ns | |||
| Mucor sp. | - | −0.087 ns | −0.146 ns | ||||
| Primary shoot length | - | 0.893 *** | |||||
| Salazar | Germination | −0.257 ns | −0.150 ns | −0.177 ns | −0.264 ns | 0.983 *** | 0.960 *** |
| Fusarium sp. | - | 0.306 ns | −0.106 ns | 0.121 ns | −0.165 ns | −0.268 ns | |
| Aternaria sp. | - | −0.250 ns | 0.416 ns | −0.213 ns | −0.240 ns | ||
| Penicillium sp. | - | −0.234 ns | 0.191 ns | 0.228 ns | |||
| Mucor sp. | - | −0.271 ns | −0.505 ns | ||||
| Primary shoot length | - | 0.955 *** | |||||
| Zemunska rosa | Germination | −0.649 * | −0.459 ns | −0.484 ns | −0.553 ns | 0.954 *** | 0.903 *** |
| Fusarium sp. | - | 0.223 ns | 0.272 ns | 0.408 ns | −0.495 ns | −0.589 ns | |
| Aternaria sp. | - | 0.515 ns | 0.545 ns | −0.500 ns | −0.562 ns | ||
| Penicillium sp. | - | 0.667 * | −0.614 ns | −0.467 ns | |||
| Mucor sp. | - | −0.572 ns | −0.643 * | ||||
| Primary shoot length | - | 0.863 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poštić, D.; Štrbanović, R.; Tabaković, M.; Popović, T.; Ćirić, A.; Banjac, N.; Trkulja, N.; Stanisavljević, R. Germination and the Initial Seedling Growth of Lettuce, Celeriac and Wheat Cultivars after Micronutrient and a Biological Application Pre-Sowing Seed Treatment. Plants 2021, 10, 1913. https://doi.org/10.3390/plants10091913
Poštić D, Štrbanović R, Tabaković M, Popović T, Ćirić A, Banjac N, Trkulja N, Stanisavljević R. Germination and the Initial Seedling Growth of Lettuce, Celeriac and Wheat Cultivars after Micronutrient and a Biological Application Pre-Sowing Seed Treatment. Plants. 2021; 10(9):1913. https://doi.org/10.3390/plants10091913
Chicago/Turabian StylePoštić, Dobrivoj, Ratibor Štrbanović, Marijenka Tabaković, Tatjana Popović, Ana Ćirić, Nevena Banjac, Nenad Trkulja, and Rade Stanisavljević. 2021. "Germination and the Initial Seedling Growth of Lettuce, Celeriac and Wheat Cultivars after Micronutrient and a Biological Application Pre-Sowing Seed Treatment" Plants 10, no. 9: 1913. https://doi.org/10.3390/plants10091913
APA StylePoštić, D., Štrbanović, R., Tabaković, M., Popović, T., Ćirić, A., Banjac, N., Trkulja, N., & Stanisavljević, R. (2021). Germination and the Initial Seedling Growth of Lettuce, Celeriac and Wheat Cultivars after Micronutrient and a Biological Application Pre-Sowing Seed Treatment. Plants, 10(9), 1913. https://doi.org/10.3390/plants10091913