
Genome-Wide Identification and Expression Profiling of Potassium Transport-Related Genes in Vigna radiata under Abiotic Stresses

 ,
,  , ,
, ,  , ,
, ,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
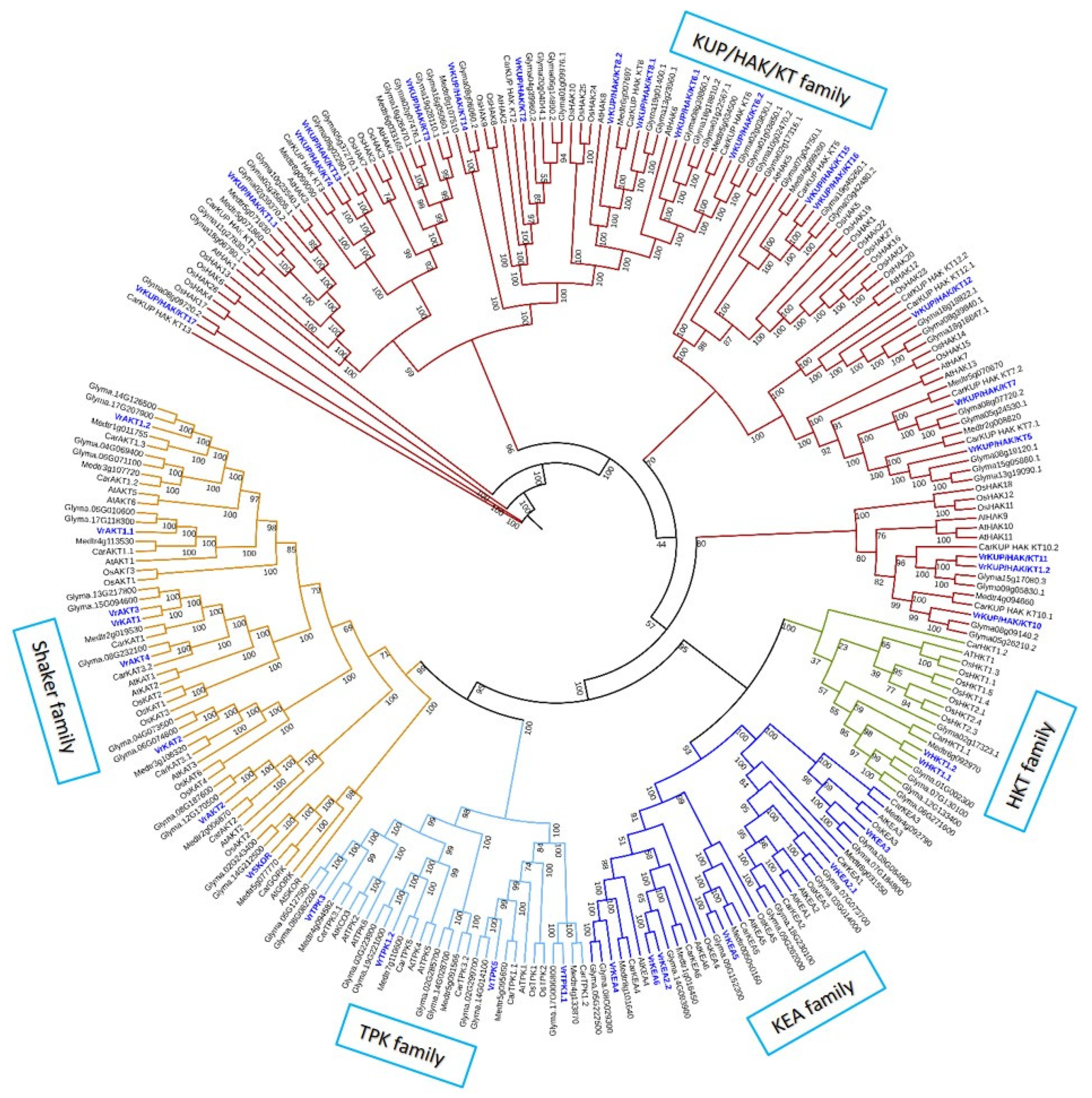
2.1. Identification and Phylogenetics of K+ Transporters and Channels
2.1.1. The K+ Transporters in V. radiata
2.1.2. K+ Channels in the V. radiata Genome
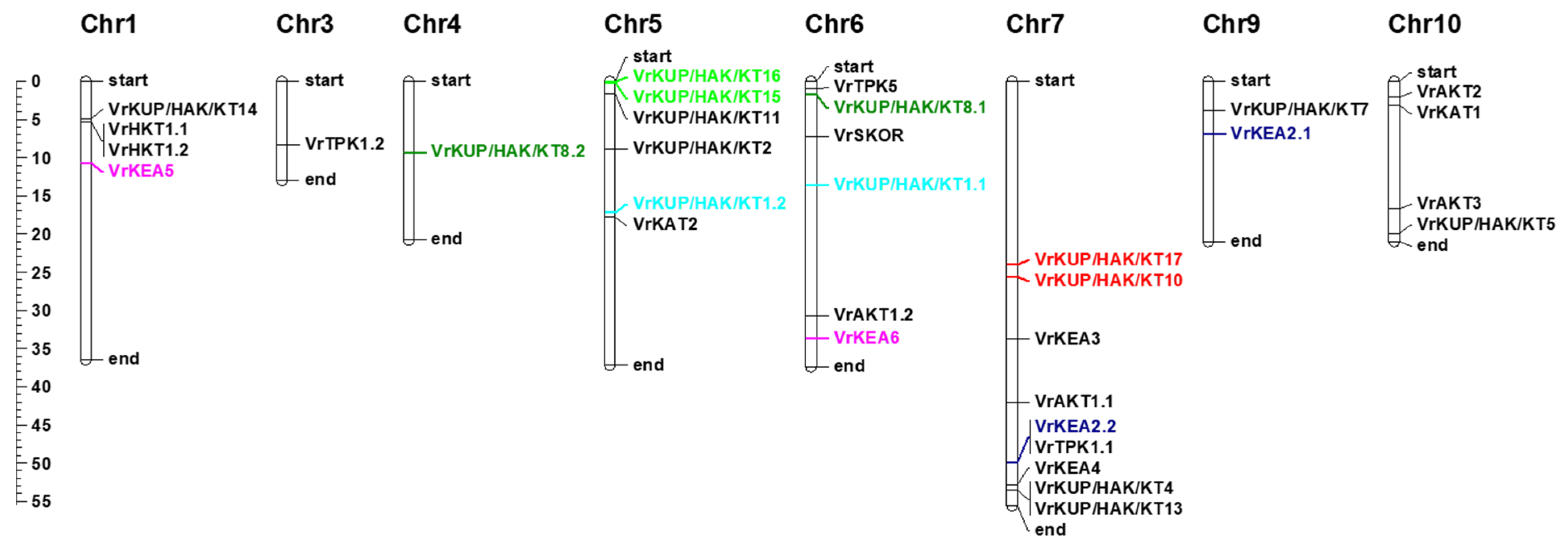
2.2. Gene Structure and Gene Duplications of K+ Transport-Related Genes in V. radiata
2.3. Promoter Analysis of Putative V. radiata K+ Transporter and Channels
2.4. Physiological and Biochemical Response of V. radiata to Drought, Heat, and Salt Stresses
2.5. Effect of Abiotic Stress on Na+ and K+ Concentration in V. radiata
2.6. Expression of K+ Transporters in Drought, Heat, and Salt Stress Conditions
3. Discussion
3.1. K+ Transporters and Channels in V. radiata Are Similar to Other Plant Species
3.2. Abiotic Stress Significantly Affects V. radiata Seedling Growth
3.3. Abiotic Stress Modulates the Expression of K+ Transporters and Channels in V. radiata
4. Materials and Methods
4.1. Data Retrieval and Identification of Potassium Transporters and Channels
4.2. Phylogeny, Gene Structure, Physical Mapping, and Duplication Analyses
4.3. Plant Material and Stress Imposition
4.4. Physiological and Biochemical Analyses
4.4.1. Chlorophyll Concentration Measurements
4.4.2. Carotenoid Concentrations Measurement
4.4.3. Biochemical Studies
4.4.4. Determination of Na+ and K+ Concentrations
4.4.5. Statistical Analysis
4.5. In-silico Expression of Potassium Transport Genes in Different Tissues
4.6. RNA Extraction, cDNA Synthesis, and qRT-PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ashley, M.K.; Grant, M.; Grabov, A. Plant responses to potassium deficiencies: A role for potassium transport proteins. J. Exp. Bot. 2006, 57, 425–436. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wu, W.-H. Potassium Transport and Signaling in Higher Plants. Annu. Rev. Plant Biol. 2013, 64, 451–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The Critical Role of Potassium in Plant Stress Response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Ishitani, M.; Kim, C.; Zhu, J.K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6896–6901. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.S.; Berkowitz, G.A.; Pier, P.A. Maintenance of photosynthesis at low leaf water potential in wheat: Role of potassium status and irrigation history. Plant Physiol. 1989, 89, 1358–1365. [Google Scholar] [CrossRef] [Green Version]
- Leigh, R.A. Potassium homeostasis and membrane transport. J. Plant Nutr. Soil Sci. 2001, 164, 193–198. [Google Scholar] [CrossRef]
- Maser, P. Phylogenetic Relationships within Cation Transporter Families of Arabidopsis. Plant Physiol. 2001, 126, 1646–1667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Very, A.-A.; Sentenac, H. Molecular mechanisms and regulation of K+ transport in higher plants. Annu. Rev. Plant Biol. 2003, 54, 575–603. [Google Scholar] [CrossRef]
- Hedrich, R. Ion channels in plants. Physiol. Rev. 2012, 92, 1777–1811. [Google Scholar] [CrossRef] [PubMed]
- Lebaudy, A.; Véry, A.-A.; Sentenac, H. K+ channel activity in plants: Genes, regulations and functions. FEBS Lett. 2007, 581, 2357–2366. [Google Scholar] [CrossRef] [PubMed]
- Nieves-Cordones, M.; Chavanieu, A.; Jeanguenin, L.; Alcon, C.; Szponarski, W.; Estaran, S.; Cherel, I.; Zimmermann, S.; Sentenac, H.; Gaillard, I. Distinct amino acids in the C-linker domain of the Arabidopsis K+ channel KAT2 determine its subcellular localization and activity at the plasma membrane. Plant Physiol. 2014, 164, 1415–1429. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Xu, G.; Alli, A.; Yu, L. Plant HAK/KUP/KT K+ transporters: Function and regulation. Semin. Cell Dev. Biol. 2018, 74, 133–141. [Google Scholar] [CrossRef]
- Sze, H.; Chanroj, S. Plant Endomembrane Dynamics: Studies of K+ /H+ Antiporters Provide Insights on the Effects of pH and Ion Homeostasis. Plant Physiol. 2018, 177, 875–895. [Google Scholar] [CrossRef] [Green Version]
- Riedelsberger, J.; Miller, J.K.; Valdebenito-Maturana, B.; Piñeros, M.A.; González, W.; Dreyer, I. Plant HKT Channels: An Updated View on Structure, Function and Gene Regulation. Int. J. Mol. Sci. 2021, 22, 1892. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Sicilia, M.N.; Cagnac, O.; Chanroj, S.; Sze, H.; Rodríguez-Rosales, M.P.; Venema, K. Arabidopsis KEA2, a homolog of bacterial KefC, encodes a K+/H+ antiporter with a chloroplast transit peptide. Biochim. Biophys. Acta Biomembr. 2012, 1818, 2362–2371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanroj, S.; Wang, G.; Venema, K.; Zhang, M.W.; Delwiche, C.F.; Sze, H. Conserved and Diversified Gene Families of Monovalent Cation/H+ Antiporters from Algae to Flowering Plants. Front. Plant Sci. 2012, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-McSweeney, A.; González-Gordo, S.; Aranda-Sicilia, M.N.; Rodríguez-Rosales, M.P.; Venema, K.; Palma, J.M.; Corpas, F.J. Loss of function of the chloroplast membrane K+/H+ antiporters AtKEA1 and AtKEA2 alters the ROS and NO metabolism but promotes drought stress resilience. Plant Physiol. Biochem. 2021, 160, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Pan, T.; Zhang, X.; Fan, L.; Quintero, F.J.; Zhao, H.; Su, X.; Li, X.; Villalta, I.; Mendoza, I.; et al. K+ Efflux Antiporters 4, 5, and 6 Mediate pH and K+ Homeostasis in Endomembrane Compartments. Plant Physiol. 2018, 178, 1657–1678. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Tang, R.-J.; Yang, X.; Zheng, X.; Shao, Q.; Tang, Q.-L.; Fu, A.; Luan, S. Golgi-localized cation/proton exchangers regulate ionic homeostasis and skotomorphogenesis in Arabidopsis. Plant Cell Environ. 2019, 42, 673–687. [Google Scholar] [CrossRef]
- Rodríguez-Navarro, A.; Rubio, F. High-affinity potassium and sodium transport systems in plants. J. Exp. Bot. 2006, 57, 1149–1160. [Google Scholar] [CrossRef] [Green Version]
- Nair, R.M.; Yang, R.Y.; Easdown, W.J.; Thavarajah, D.; Thavarajah, P.; Hughes, J.d.A.; Keatinge, J.D.H. Biofortification of mungbean (Vigna radiata) as a whole food to enhance human health. J. Sci. Food Agric. 2013, 93, 1805–1813. [Google Scholar] [CrossRef]
- Sehrawat, N.; Yadav, M.; Sharma, A.K.; Kumar, V.; Bhat, K.V. Salt stress and mungbean [Vigna radiata (L.) Wilczek]: Effects, physiological perspective and management practices for alleviating salinity. Arch. Agron. Soil Sci. 2019, 65, 1287–1301. [Google Scholar] [CrossRef]
- Pataczek, L.; Zahir, Z.A.; Ahmad, M.; Rani, S.; Nair, R.; Schafleitner, R.; Cadisch, G.; Hilger, T. Beans with Benefits—The Role of Mungbean (Vigna radiata) in a Changing Environment. Am. J. Plant Sci. 2018, 9, 1577–1600. [Google Scholar] [CrossRef] [Green Version]
- Kant, S.; Kafkafi, U. Potassium and Abiotic Stresses in Plants. In Potassium for Sustainable Crop Production. 2002, 233, 251. [Google Scholar]
- Azeem, F.; Ahmad, B.; Atif, R.M.; Ali, M.A.; Nadeem, H.; Hussain, S.; Manzoor, H.; Azeem, M.; Afzal, M. Genome-Wide Analysis of Potassium Transport-Related Genes in Chickpea (Cicer arietinum L.) and Their Role in Abiotic Stress Responses. Plant Mol. Biol. Report. 2018, 36, 451–468. [Google Scholar] [CrossRef]
- Rehman, H.M.; Nawaz, M.A.; Shah, Z.H.; Daur, I.; Khatoon, S.; Yang, S.H.; Chung, G. In-Depth Genomic and Transcriptomic Analysis of Five K+ Transporter Gene Families in Soybean Confirm Their Differential Expression for Nodulation. Front. Plant Sci. 2017, 8, 804. [Google Scholar] [CrossRef] [Green Version]
- Amrutha, R.N.; Sekhar, P.N.; Varshney, R.K.; Kishor, P.B.K. Genome-wide analysis and identification of genes related to potassium transporter families in rice (Oryza sativa L.). Plant Sci. 2007, 172, 708–721. [Google Scholar] [CrossRef] [Green Version]
- Gierth, M.; Mäser, P. Potassium transporters in plants—Involvement in K+ acquisition, redistribution and homeostasis. FEBS Lett. 2007, 581, 2348–2356. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Liu, X.; Mao, W.; Zhang, X.; Chen, S.; Zhan, K.; Bi, H.; Xu, H. Genome-Wide Identification and Analysis of HAK/KUP/KT Potassium Transporters Gene Family in Wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2018, 19, 3969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Véry, A.-A.; Nieves-Cordones, M.; Daly, M.; Khan, I.; Fizames, C.; Sentenac, H. Molecular biology of K+ transport across the plant cell membrane: What do we learn from comparison between plant species? J. Plant Physiol. 2014, 171, 748–769. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Ródenas, R.; Chavanieu, A.; Rivero, R.M.; Martinez, V.; Gaillard, I.; Rubio, F. Uneven HAK/KUP/KT Protein Diversity Among Angiosperms: Species Distribution and Perspectives. Front. Plant Sci. 2016, 7, 127. [Google Scholar] [CrossRef] [Green Version]
- Lerouge, P.; Cabanes-Macheteau, M.; Rayon, C.; Fischette-Lainé, A.C.; Gomord, V.; Faye, L. N-glycoprotein biosynthesis in plants: Recent developments and future trends. Plant Mol. Biol. 1998, 38, 31–48. [Google Scholar] [CrossRef]
- Koizumi, N.; Ujino, T.; Sano, H.; Chrispeels, M.J. Overexpression of a gene that encodes the first enzyme in the biosynthesis of asparagine-linked glycans makes plants resistant to tunicamycin and obviates the tunicamycin-induced unfolded protein response. Plant Physiol. 1999, 121, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Tsujii, M.; Kera, K.; Hamamoto, S.; Kuromori, T.; Shikanai, T.; Uozumi, N. Evidence for potassium transport activity of Arabidopsis KEA1-KEA6. Sci. Rep. 2019, 9, 10040. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7. 0 for Bigger Datasets Brief communication. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamb, A.; Iverson, L.E.; Tanouye, M.A. Molecular characterization of Shaker, a Drosophila gene that encodes a potassium channel. Cell 1987, 50, 405–413. [Google Scholar] [CrossRef]
- Cuéllar, T.; Pascaud, F.; Verdeil, J.-L.; Torregrosa, L.; Adam-Blondon, A.-F.; Thibaud, J.-B.; Sentenac, H.; Gaillard, I. A grapevine Shaker inward K(+) channel activated by the calcineurin B-like calcium sensor 1-protein kinase CIPK23 network is expressed in grape berries under drought stress conditions. Plant J. 2010, 61, 58–69. [Google Scholar] [CrossRef]
- Corratgé-Faillie, C.; Ronzier, E.; Sanchez, F.; Prado, K.; Kim, J.-H.; Lanciano, S.; Leonhardt, N.; Lacombe, B.; Xiong, T.C. The Arabidopsis guard cell outward potassium channel GORK is regulated by CPK33. FEBS Lett. 2017, 591, 1982–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, D.a. The Structure of the Potassium Channel: Molecular Basis of K+ Conduction and Selectivity. Science 1998, 280, 69–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czempinski, K.; Gaedeke, N.; Zimmermann, S.; Müller-Röber, B.; Mu, B. Molecular mechanisms and regulation of plant ion channels. J. Exp. Bot. 1999, 50, 955–966. [Google Scholar] [CrossRef]
- Hamamoto, S.; Marui, J.; Matsuoka, K.; Higashi, K.; Igarashi, K.; Nakagawa, T.; Kuroda, T.; Mori, Y.; Murata, Y.; Nakanishi, Y.; et al. Characterization of a tobacco TPK-type K+ channel as a novel tonoplast K+ channel using yeast tonoplasts. J. Biol. Chem. 2008, 283, 1911–1920. [Google Scholar] [CrossRef] [Green Version]
- Isayenkov, S.; Isner, J.-C.; Maathuis, F.J.M. Rice two-pore K+ channels are expressed in different types of vacuoles. Plant Cell 2011, 23, 756–768. [Google Scholar] [CrossRef] [Green Version]
- Marcel, D.; Müller, T.; Hedrich, R.; Geiger, D. K+ transport characteristics of the plasma membrane tandem-pore channel TPK4 and pore chimeras with its vacuolar homologs. FEBS Lett. 2010, 584, 2433–2439. [Google Scholar] [CrossRef] [Green Version]
- Ijaz, U.; Pervaiz, T.; Ahmed, T.; Seemab, R.; Shahid, M.; Noman, M.; Nadeem, M.; Azeem, F. Plant Cis-regulatory elements: Methods of identification and applications. Asian J. Agric. Biol. 2020, 8, 207–222. [Google Scholar] [CrossRef]
- Huang, G.-T.; Ma, S.-L.; Bai, L.-P.; Zhang, L.; Ma, H.; Jia, P.; Liu, J.; Zhong, M.; Guo, Z.-F. Signal transduction during cold, salt, and drought stresses in plants. Mol. Biol. Rep. 2012, 39, 969–987. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.-S.; Kuo, T.C.-Y.; Ko, C.-Y.; Wu, D.-C.; Li, K.-Y.; Lin, W.-J.; Lin, C.-P.; Wang, Y.-W.; Schafleitner, R.; Lo, H.-F.; et al. Genomic and transcriptomic comparison of nucleotide variations for insights into bruchid resistance of mungbean (Vigna radiata [L.] R. Wilczek). BMC Plant Biol. 2016, 16, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.; Li, S.; Liu, Y.; Liu, X. Transcriptomic profiling reveals metabolic and regulatory pathways in the desiccation tolerance of Mungbean (Vigna radiata [L.] R. Wilczek). Front. Plant Sci. 2016, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Marschner, H.; Kirkby, E.A.; Cakmak, I. Effect of mineral nutritional status on shoot-root partitioning of photoassimilates and cycling of mineral nutrients. J. Exp. Bot. 1996, 47, 1255–1263. [Google Scholar] [CrossRef]
- Wang, S.; Wan, C.; Wang, Y.; Chen, H.; Zhou, Z.; Fu, H.; Sosebee, R.E. The characteristics of Na+, K+ and free proline distribution in several drought-resistant plants of the Alxa Desert, China. J. Arid Environ. 2004, 56, 525–539. [Google Scholar] [CrossRef]
- Ahmad, I.; Mian, A.; Maathuis, F.J.M. Overexpression of the rice AKT1 potassium channel affects potassium nutrition and rice drought tolerance. J. Exp. Bot. 2016, 67, 2689–2698. [Google Scholar] [CrossRef] [Green Version]
- Schachtman, D.P.; Schroeder, J.I.; Lucas, W.J.; Anderson, J.A.; Gaber, R.F. Expression of an inward-rectifying potassium channel by the Arabidopsis KAT1 cDNA. Science 1992, 258, 1654–1658. [Google Scholar] [CrossRef]
- Sentenac, H.; Bonneaud, N.; Minet, M. Cloning and expression in yeast of a plant potassium ion transport system. Science 1992, 256. [Google Scholar] [CrossRef]
- Ketchum, K.A.; Slayman, C.W. Isolation of an ion channel gene from Arabidopsis thaliana using the H5 signature sequence from voltage-dependent K+ channels. FEBS Lett. 1996, 378, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Uozumi, N.; Gassmann, W.; Cao, Y.; Schroeder, J.I. Identification of strong modifications in cation selectivity in an Arabidopsis inward rectifying potassium channel by mutant selection in yeast. J. Biol. Chem. 1995, 270, 24276–24281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butt, A.D.; Blatt, M.R.; Ainsworth, C.C. Expression, evolution and genomic complexity of potassium ion channel genes of Arabidopsis thaliana. J. Plant Physiol. 1997, 150, 652–660. [Google Scholar] [CrossRef]
- Reintanz, B.; Szyroki, A.; Ivashikina, N.; Ache, P.; Godde, M.; Becker, D.; Palme, K.; Hedrich, R. AtKC1, a silent Arabidopsis potassium channel alpha -subunit modulates root hair K+ influx. Proc. Natl. Acad. Sci. USA 2002, 99, 4079–4084. [Google Scholar] [CrossRef] [Green Version]
- Ketehouli, T.; Idrice Carther, K.F.; Noman, M.; Wang, F.-W.; Li, X.-W.; Li, H.-Y. Adaptation of Plants to Salt Stress: Characterization of Na+ and K+ Transporters and Role of CBL Gene Family in Regulating Salt Stress Response. Agronomy 2019, 9, 687. [Google Scholar] [CrossRef] [Green Version]
- Sangar, V.; Blankenberg, D.J.; Altman, N.; Lesk, A.M. Quantitative sequence-function relationships in proteins based on gene ontology. BMC Bioinform. 2007, 8, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keisham, M.; Mukherjee, S.; Bhatla, S.C. Mechanisms of Sodium Transport in Plants—Progresses and Challenges. Int. J. Mol. Sci. 2018, 19, 647. [Google Scholar] [CrossRef] [Green Version]
- Nair, R.M.; Pandey, A.K.; War, A.R.; Hanumantharao, B.; Shwe, T.; Alam, A.; Pratap, A.; Malik, S.R.; Karimi, R.; Mbeyagala, E.K.; et al. Biotic and Abiotic Constraints in Mungbean Production—Progress in Genetic Improvement. Front. Plant Sci. 2019, 10, 1340. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and Salinity Stress Responses and Microbe-Induced Tolerance in Plants. Front. Plant Sci. 2020, 11, 1750. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef] [Green Version]
- Taïbi, K.; Taïbi, F.; Ait Abderrahim, L.; Ennajah, A.; Belkhodja, M.; Mulet, J.M. Effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidant defence systems in Phaseolus vulgaris L. S. Afr. J. Bot. 2016, 105, 306–312. [Google Scholar] [CrossRef]
- Walker, B.J.; Drewry, D.T.; Slattery, R.A.; VanLoocke, A.; Cho, Y.B.; Ort, D.R. Chlorophyll Can Be Reduced in Crop Canopies with Little Penalty to Photosynthesis. Plant Physiol. 2018, 176, 1215–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopsell, D.E.; Kopsell, D.A.; Randle, W.M.; Coolong, T.W.; Sams, C.E.; Curran-Celentano, J. Kale carotenoids remain stable while flavor compounds respond to changes in sulfur fertility. J. Agric. Food Chem. 2003, 51, 5319–5325. [Google Scholar] [CrossRef]
- Silva, E.N.; Vieira, S.A.; Ribeiro, R.V.; Ponte, L.F.A.; Ferreira-Silva, S.L.; Silveira, J.A.G. Contrasting Physiological Responses of Jatropha curcas Plants to Single and Combined Stresses of Salinity and Heat. J. Plant Growth Regul. 2013, 32, 159–169. [Google Scholar] [CrossRef]
- Kim, S.H.; Ahn, Y.O.; Ahn, M.-J.; Lee, H.-S.; Kwak, S.-S. Down-regulation of β-carotene hydroxylase increases β-carotene and total carotenoids enhancing salt stress tolerance in transgenic cultured cells of sweetpotato. Phytochemistry 2012, 74, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Strzałka, K.; Kostecka-Gugała, A.; Latowski, D. Carotenoids and Environmental Stress in Plants: Significance of Carotenoid-Mediated Modulation of Membrane Physical Properties. Russ. J. Plant Physiol. 2003, 50, 168–173. [Google Scholar] [CrossRef]
- Waqas, M.A.; Kaya, C.; Riaz, A.; Farooq, M.; Nawaz, I.; Wilkes, A.; Li, Y. Potential Mechanisms of Abiotic Stress Tolerance in Crop Plants Induced by Thiourea. Front. Plant Sci. 2019, 10, 1336. [Google Scholar] [CrossRef]
- Perveen, A.; Wahid, A.; Mahmood, S.; Hussain, I.; Rasheed, R. Possible mechanism of medium-supplemented thiourea in improving growth, gas exchange, and photosynthetic pigments in cadmium-stressed maize (Zea mays). Brazilian J. Bot. 2015, 38, 71–79. [Google Scholar] [CrossRef]
- Lakhdar, A.; Hafsi, C.; Rabhi, M.; Debez, A.; Montemurro, F.; Abdelly, C.; Jedidi, N.; Ouerghi, Z. Application of municipal solid waste compost reduces the negative effects of saline water in Hordeum maritimum L. Bioresour. Technol. 2008, 99, 7160–7167. [Google Scholar] [CrossRef] [PubMed]
- Dagar, J.C.; Tomar, O.S.; Kumar, Y.; Yadav, R.K. Growing three aromatic grasses in different alkali soils in semi-arid regions of northern India. L. Degrad. Dev. 2004, 15, 143–151. [Google Scholar] [CrossRef]
- Sawada, Y.; Oyama, T.; Yamazaki, I. Preparation and physicochemical properties of green pea superoxide dismutase. Biochim. Biophys. Acta 1972, 268, 305–312. [Google Scholar] [CrossRef]
- Abedi, T.; Pakniyat, H. Antioxidant enzymes changes in response to drought stress in ten cultivars of oilseed rape (Brassica napus L.). Czech J. Genet. Plant Breed. 2010, 46, 27–34. [Google Scholar] [CrossRef]
- Rached-kanouni, M.; Kehal, L.; Touaba, C.; Bouzobra, R.; Belghit, F.; Alatou, D. Change in activity of antioxidative enzymes in leaves of Acacia retinodes, Biota orientalis and Casuarina equisetifolia under heat stress condition. Eur. Sci. J. 2013, 9, 402–410. [Google Scholar]
- Sarker, U.; Oba, S. The Response of Salinity Stress-Induced A. tricolor to Growth, Anatomy, Physiology, Non-Enzymatic and Enzymatic Antioxidants. Front. Plant Sci. 2020, 11, 559876. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Gao, Y.; Mao, T.; Zhang, X.; Zhang, S.; Zhang, H.; Song, Z. Characterization and Expression of KT/HAK/KUP Transporter Family Genes in Willow under Potassium Deficiency, Drought, and Salt Stresses. Biomed Res. Int. 2020, 2020, 2690760. [Google Scholar] [CrossRef]
- Ragel, P.; Raddatz, N.; Leidi, E.O.; Quintero, F.J.; Pardo, J.M. Regulation of K+ Nutrition in Plants. Front. Plant Sci. 2019, 10, 281. [Google Scholar] [CrossRef] [Green Version]
- Sharma, T.; Dreyer, I.; Riedelsberger, J. The role of K+ channels in uptake and redistribution of potassium in the model plant Arabidopsis thaliana. Front. Plant Sci. 2013, 4, 224. [Google Scholar] [CrossRef] [Green Version]
- HanumanthaRao, B.; Nair, R.M.; Nayyar, H. Salinity and High Temperature Tolerance in Mungbean [Vigna radiata (L.) Wilczek] from a Physiological Perspective. Front. Plant Sci. 2016, 7, 957. [Google Scholar] [CrossRef] [Green Version]
- Abhinandan, K.; Skori, L.; Stanic, M.; Hickerson, N.M.N.; Jamshed, M.; Samuel, M.A. Abiotic Stress Signaling in Wheat—An Inclusive Overview of Hormonal Interactions During Abiotic Stress Responses in Wheat. Front. Plant Sci. 2018, 9, 734. [Google Scholar] [CrossRef] [PubMed]
- James, R.A.; Blake, C.; Byrt, C.S.; Munns, R. Major genes for Na+ exclusion, Nax1 and Nax2 (wheat HKT1;4 and HKT1;5), decrease Na+ accumulation in bread wheat leaves under saline and waterlogged conditions. J. Exp. Bot. 2011, 62, 2939–2947. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Qi, J.; Sun, S.; Yang, L.; Li, M.; Ma, F.; Zou, Y. Potassium Uptake and Transport in Apple Roots Under Drought Stress. Hortic. Plant J. 2019, 5, 10–16. [Google Scholar] [CrossRef]
- Vacic, V.; Iakoucheva, L.M.; Radivojac, P. Two Sample Logo: A graphical representation of the differences between two sets of sequence alignments. Bioinformatics 2006, 22, 1536–1537. [Google Scholar] [CrossRef] [Green Version]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, Y.Y.; Ichida, H.; Abe, T.; Suzuki, Y.; Sugano, S.; Obokata, J. Differentiation of core promoter architecture between plants and mammals revealed by LDSS analysis. Nucleic Acids Res. 2007, 35, 6219–6226. [Google Scholar] [CrossRef] [Green Version]
- Rose, A.B.; Elfersi, T.; Parra, G.; Korf, I. Promoter-Proximal Introns in Arabidopsis thaliana Are Enriched in Dispersed Signals that Elevate Gene Expression. Plant Cell 2008, 20, 543–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, A.B.; Carter, A.; Korf, I.; Kojima, N. Intron sequences that stimulate gene expression in Arabidopsis. Plant Mol. Biol. 2016, 92, 337–346. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Lotfi, R. The impact of salicylic acid and silicon on chlorophyll a fluorescence in mung bean under salt stress. Russ. J. Plant Physiol. 2015, 62, 611–616. [Google Scholar] [CrossRef]
- Kono, Y. Generation of superoxide radical during autoxidation of hydroxylamine and an assay for superoxide dismutase. Arch. Biochem. Biophys. 1978, 186, 189–195. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalases and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar] [CrossRef]
- Aebi, H.E. Catalase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Verlag Chemie: Weinheim, Germany, 1982; pp. 273–286. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Farasat, M. Salt Tolerance in Rice Cultivars and Changes in Sodium and Potassium Ions. J. Crop Nutr. Sci. 2017, 3, 1–13. [Google Scholar]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools-an integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr# | Locus Tag | Gene Name | Protein ID | Isoelectric Point | Molecular Weight | TM Domains | Domains | Protein Length | Chr # |
|---|---|---|---|---|---|---|---|---|---|
| 1 | LOC106764953 | VrKUP/HAK/KT1.1 | XP_014504897.1 | 6.89 | 84.60425 | 13 | K_trans | 759 | 6 |
| 2 | LOC106785321 | VrKUP/HAK/KT1.2 | XP_014501656.1 | 6.12 | 91.40354 | 14 | K_trans | 791 | 5 |
| 3 | LOC106762218 | VrKUP/HAK/KT2 | XP_014501480.1 | 6.7 | 88.31495 | 12 | K_trans | 791 | 5 |
| 4 | LOC106753929 | VrKUP/HAK/KT3 | XP_014491302.1 | 8.66 | 87.40831 | 13 | PLN | 784 | Unknown |
| 5 | LOC106767047 | VrKUP/HAK/KT4 | XP_014507349.1 | 9.28 | 88.27794 | 11 | K_trans | 790 | 7 |
| 6 | LOC106775695 | VrKUP/HAK/KT5 | XP_014518335.1 | 5.79 | 93.4367 | 11 | PLN | 840 | 10 |
| 7 | LOC106779868 | VrKUP/HAK/KT6.1 | XP_014523566.1 | 8.65 | 85.72046 | 12 | K_trans | 772 | Unknown |
| 8 | LOC106779674 | VrKUP/HAK/KT6.2 | XP_014523333.1 | 8.66 | 86.64245 | 13 | PotE, K_trans | 776 | Unknown |
| 9 | LOC106773994 | VrKUP/HAK/KT7 | XP_014516251.1 | 5.88 | 94.30594 | 10 | PotE, PLN | 846 | 9 |
| 10 | LOC106771412 | VrKUP/HAK/KT8.1 | XP_014512874.1 | 7.83 | 87.44141 | 12 | PotE, PLN | 779 | 6 |
| 11 | LOC106759282 | VrKUP/HAK/KT8.2 | XP_014497862.1 | 7.29 | 87.06509 | 13 | PotE, PLN | 775 | 4 |
| 12 | LOC106768780 | VrKUP/HAK/KT10 | XP_014509587.1 | 8.27 | 88.70751 | 13 | K_trans, PotE | 791 | 7 |
| 13 | LOC106762322 | VrKUP/HAK/KT11 | XP_022636150.1 | 7.59 | 84.73469 | 12 | K_trans | 758 | 5 |
| 14 | LOC106779912 | VrKUP/HAK/KT12 | XP_014523613.1 | 6.63 | 92.41383 | 11 | PLN, PotE | 833 | Unknown |
| 15 | LOC106767048 | VrKUP/HAK/KT13 | XP_014507351.1 | 9.44 | 88.0777 | 12 | K_trans | 790 | 7 |
| 16 | LOC106759816 | VrKUP/HAK/KT14 | XP_014498681.1 | 8.2 | 88.92217 | 11 | K_trans | 796 | 1 |
| 17 | LOC106760231 | VrKUP/HAK/KT15 | XP_014499185.1 | 9.02 | 89.50289 | 12 | K_trans, | 804 | 5 |
| 18 | LOC106760017 | VrKUP/HAK/KT16 | XP_014498941.1 | 6.48 | 68.47955 | 7 | K_trans | 610 | 5 |
| 19 | LOC106766977 | VrKUP/HAK/KT17 | XP_014507267.1 | 8.17 | 80.93343 | 12 | PotE, K_trans | 723 | 7 |
| 20 | LOC106756241 | VrHKT1.1 | XP_014494069.1 | 9.39 | 57.12935 | 9 | TrkH | 507 | 1 |
| 21 | LOC106763952 | VrHKT1.2 | XP_014503627.1 | 9.39 | 58.81727 | 9 | TrkH | 518 | 1 |
| 22 | LOC106774167 | VrKEA2.1 | XP_014516532.1 | 4.96 | 78.72034 | 10 | TrkA_N, RILP | 1195 | 9 |
| 23 | LOC106764844 | VrKEA2.2 | XP_022638221.1 | 4.55 | 56.79963 | 6 | Na_H_Exchanger | 527 | 7 |
| 24 | LOC106768449 | VrKEA3 | XP_014509103.1 | 5.52 | 85.86885 | 1 | KefB, TrkA_N | 790 | 7 |
| 25 | LOC106769002 | VrKEA4 | XP_014509921.1 | 5.77 | 62.90096 | 12 | Na_H_Exchanger | 586 | 7 |
| 26 | LOC106771900 | VrKEA5 | XP_014513420.1 | 6.17 | 62.48549 | 11 | Na_H_Exchanger | 576 | 1 |
| 27 | LOC107634854 | VrKEA6 | XP_022638220.1 | 5.55 | 56.79963 | 11 | Na_H_Exchanger | 595 | 6 |
| 28 | LOC106766559 | VrTPK1.1 | XP_014506766.1 | 5.5 | 38.65153 | 5 | Ion_trans_2 | 344 | 7 |
| 29 | LOC106756712 | VrTPK1.2 | XP_014494736.1 | 9.01 | 43.24297 | 5 | Ion_trans_2 | 389 | 3 |
| 30 | LOC106752883 | VrTPK3 | XP_014490151.1 | 8.76 | 47.3163 | 5 | Ion_trans_2, EF-hand_7 | 425 | Unknown |
| 31 | LOC106764594 | VrTPK5 | XP_014504362.1 | 6.11 | 38.82113 | 5 | Ion_trans_2, EFh | 348 | 6 |
| 32 | LOC106765548 | VrAKT1.1 | XP_014505698.1 | 7.04 | 97.42988 | 5 | ANK, KHA, PLN03192, Ion_trans_2 | 869 | 7 |
| 33 | LOC106763693 | VrAKT1.2 | XP_014503345.1 | 6.36 | 99.25835 | 5 | PLN03192, CAP_ED, ANK, Ion_trans_2 | 875 | 6 |
| 34 | LOC106776053 | VrAKT2 | XP_014518839.1 | 6.2 | 95.02182 | 7 | PLN03192, CAP_ED, KHA, ANK | 832 | 10 |
| 35 | LOC106775235 | VrAKT3 | XP_014517810.1 | 6.51 | 89.20001 | 5 | PLN03192, Ion_trans_2, KHA | 776 | 10 |
| 36 | LOC106752947 | VrAKT4 | XP_014490211.1 | 6.84 | 83.63201 | 6 | PLN03192, KHA, Ion_trans, CAP_ED | 717 | Unknown |
| 37 | LOC106775884 | VrKAT1 | XP_014518605.1 | 6.27 | 89.45222 | 5 | PLN03192, Ion_trans, cNMP_binding, KHA | 778 | 10 |
| 38 | LOC106761761 | VrKAT2 | XP_022636536.1 | 8.95 | 71.49999 | 5 | PLN03192, Ank_2, KHA | 623 | 5 |
| 39 | LOC106765054 | VrSKOR | XP_014505028.1 | 6.46 | 97.56075 | 5 | PLN03192, ANK, KHA, Ion_trans_2 | 851 | 6 |
| Regulatory Element | Core Sequence | VrAKT1.1 | VrAKT1.2 | VrAKT1.3 | VrAKT2 | VrKAT1.1 | VrKAT1.2 | VrKAT1 | VrSKOR | VrHKT1.1 | KT1.2 | Function |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ABRE | CACGTG | 1 | 1 | 1 | 1 | Response to abscisic acid signals | ||||||
| ACGTG | 1 | 2 | 1 | 3 | 3 | 1 | ||||||
| MYB | TAACCA | 2 | 4 | 2 | 4 | 2 | 2 | 2 | Response to drought stress and ABA signals | |||
| CAACCA | 1 | 1 | 1 | 2 | 1 | 2 | 2 | 1 | ||||
| MYC | CATTTG | 4 | 2 | 2 | 6 | 2 | 3 | 4 | 2 | 3 | Response to drought, ABA, and cold signals | |
| W-box | TTGACC | 1 | 1 | 2 | Response to SA, GA, and pathogenesis signals | |||||||
| GT-1 motif | GGTTAA | 1 | 1 | 1 | 3 | 4 | 1 | 1 | 4 | Light-responsive element | ||
| G-box | CACGTG | 1 | 3 | 3 | 3 | 1 | Involved in the light response | |||||
| GARE | TCTGTTG | 1 | Gibberellin-responsive element | |||||||||
| MBS | CAACTG | 3 | 7 | 1 | 2 | 4 | 8 | Involved in drought-inducibility | ||||
| ARE | AAACCA | 1 | 1 | 1 | 1 | 4 | 1 | Essential for the anaerobic induction | ||||
| TCA-element | CCATCTTTTT | 2 | 1 | 1 | 1 | Response to salicylic acid | ||||||
| TC-rich repeats | ATTCTCTAAC | 2 | 2 | 4 | 10 | 5 | 2 | Involved in defense and the stress response | ||||
| P-box | CCTTTTG | 1 | 1 | Gibberellin-responsive element | ||||||||
| LTR | CCGAAA | 4 | 2 | 1 | Response to low temperature | |||||||
| I-Box | GATAA | 10 | 4 | 4 | 10 | 6 | 4 | 3 | 4 | 1 | Response to SA, GA, and pathogenesis signals | |
| W-Box | TGAC | 18 | 12 | 10 | 12 | 10 | 3 | 7 | 9 | 6 | Response to drought, ABA, and cold signals |
| Treatments | Na+ Concentration (mg g−1) | K+ Concentration (mg g−1) |
|---|---|---|
| Control | 38.97 ± 1.29 | 60.74 ± 2.27 |
| Salt | 51.71 ± 1.49 ** | 45.70 ± 1.82 ** |
| Drought | 40.21 ± 1.39 | 56.16 ± 1.41 |
| Heat | 39.76 ± 1.12 | 55.270 ± 1.38 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azeem, F.; Ijaz, U.; Ali, M.A.; Hussain, S.; Zubair, M.; Manzoor, H.; Abid, M.; Zameer, R.; Kim, D.-S.; Golokhvast, K.S.; et al. Genome-Wide Identification and Expression Profiling of Potassium Transport-Related Genes in Vigna radiata under Abiotic Stresses. Plants 2022, 11, 2. https://doi.org/10.3390/plants11010002
Azeem F, Ijaz U, Ali MA, Hussain S, Zubair M, Manzoor H, Abid M, Zameer R, Kim D-S, Golokhvast KS, et al. Genome-Wide Identification and Expression Profiling of Potassium Transport-Related Genes in Vigna radiata under Abiotic Stresses. Plants. 2022; 11(1):2. https://doi.org/10.3390/plants11010002
Chicago/Turabian StyleAzeem, Farrukh, Usman Ijaz, Muhammad Amjad Ali, Sabir Hussain, Muhammad Zubair, Hamid Manzoor, Muhammad Abid, Roshan Zameer, Dong-Seon Kim, Kirill S. Golokhvast, and et al. 2022. "Genome-Wide Identification and Expression Profiling of Potassium Transport-Related Genes in Vigna radiata under Abiotic Stresses" Plants 11, no. 1: 2. https://doi.org/10.3390/plants11010002
APA StyleAzeem, F., Ijaz, U., Ali, M. A., Hussain, S., Zubair, M., Manzoor, H., Abid, M., Zameer, R., Kim, D. -S., Golokhvast, K. S., Chung, G., Sun, S., & Nawaz, M. A. (2022). Genome-Wide Identification and Expression Profiling of Potassium Transport-Related Genes in Vigna radiata under Abiotic Stresses. Plants, 11(1), 2. https://doi.org/10.3390/plants11010002