
Artificial Neural Networks Elucidated the Essential Role of Mineral Nutrients versus Vitamins and Plant Growth Regulators in Achieving Healthy Micropropagated Plants

 ,
, 
 , and
, and
Abstract
:1. Introduction
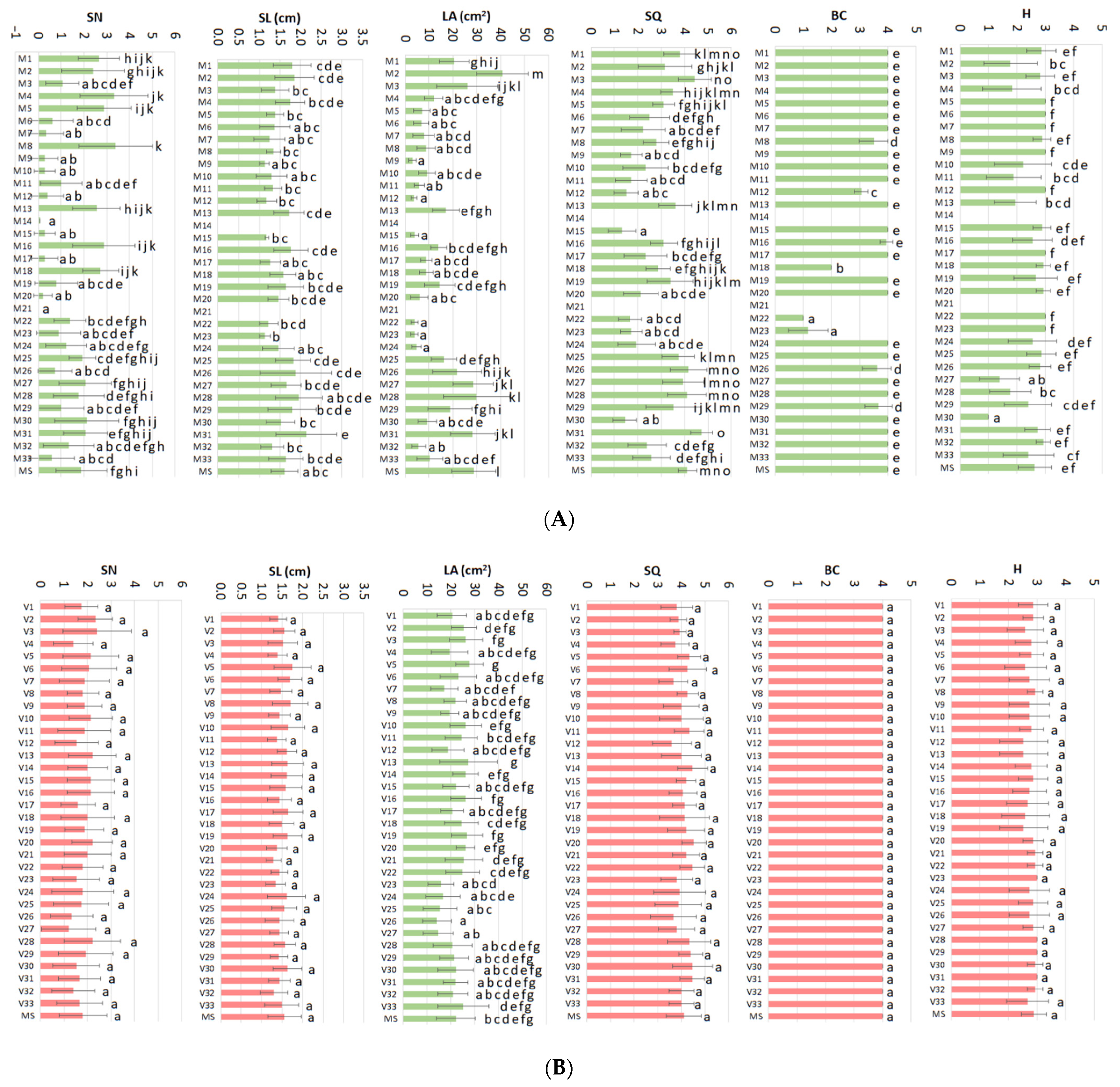
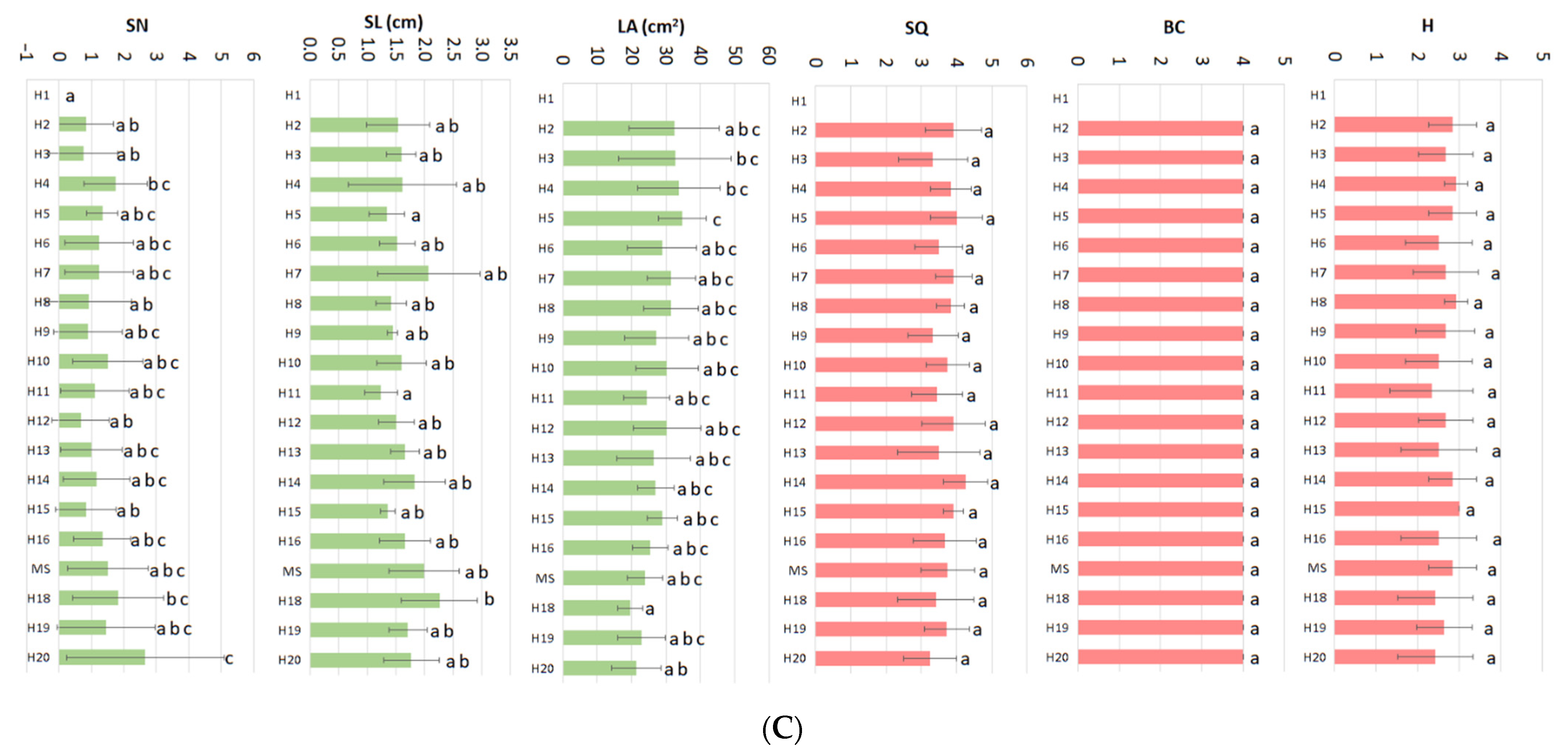
2. Results
- (i)
- Low BAP and Low GA3
- (ii)
- Mid_2 BAP and High GA3
- (iii)
- High BAP and High GA3
3. Discussion
4. Materials and Methods
4.1. Plant Material and Stock Condition
4.2. Micropropagation Culture Conditions
4.3. Experimental Design and Data Acquisition
- Shoots number (SN), number of new regenerated shoots per explant, longer than 1 cm.
- Shoot length (SL), length from the base to the tip of the new regenerated shoots longer than 1 cm.
- Leaf area (LA), the sum of areas of the leaves >1.5 cm was measured (cm2) for all the explants (the original and the new ones), using a portable laser leaf area meter (Meter CI-202, CID biosciences, WA, USA).
- Shoot quality (SQ) as indicative of shoot vigor, was visually assessed, and scored from 1 to 5 (1 very poor, 2 poor, 3 moderate, 4 good, and 5 very good).
- Basal callus (BC), callus formation at the cut edge of shoots was visually assessed and scored from 1 to 4 (1 necrotic, 2 big, 3 moderate, and 4 absent).
- Hyperhydricity (H), was visually assessed and scored from 1 to 3 (1 high, 2 low, and 3 none).
4.4. Statistical Analysis
4.5. Artificial Neural Network Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferguson, A.R.; Huang, H. Genetic Resources of Kiwifruit: Domestication and Breeding. In Horticultural Reviews; Janick, J., Ed.; Wiley: New Jersey, NJ, USA, 2007; pp. 1–121. ISBN 978-0-470-16801-1. [Google Scholar]
- Williams, M.H.; Boyd, L.M.; McNeilage, M.A.; MacRae, E.A.; Ferguson, A.R.; Beatson, R.A.; Martin, P.J. Development and Commercialization of “Baby Kiwi” (Actinidia arguta Planch.). Acta Hortic. 2003, 610, 81–86. [Google Scholar] [CrossRef]
- Latocha, P.; Debersaques, F.; Hale, I. Actinidia arguta (Kiwiberry): Botany, Production, Genetics, Nutritional Value, and Postharvest. In Horticultural Reviews; Warrington, I., Ed.; Wiley: New York, NY, USA, 2021; Volume 48, pp. 37–152. ISBN 978-1-119-75077-2. [Google Scholar]
- Hameg, R.; Arteta, T.A.; Landin, M.; Gallego, P.P.; Barreal, M.E. Modeling and Optimizing Culture Medium Mineral Composition for In Vitro Propagation of Actinidia arguta. Front. Plant Sci. 2020, 11, 554905. [Google Scholar] [CrossRef] [PubMed]
- George, E.F.; Hall, M.A.; Klerk, G.-J.D. (Eds.) Plant Propagation by Tissue Culture; Springer: Dordrecht, The Netherlands, 2007; ISBN 978-1-4020-5004-6. [Google Scholar]
- Phillips, G.C.; Garda, M. Plant Tissue Culture Media and Practices: An Overview. In Vitro Cell. Dev. Biol. Plant 2019, 55, 242–257. [Google Scholar] [CrossRef]
- Reed, B.M.; Wada, S.; DeNoma, J.; Niedz, R.P. Improving in Vitro Mineral Nutrition for Diverse Pear Germplasm. In Vitro Cell. Dev. Biol. Plant 2013, 49, 343–355. [Google Scholar] [CrossRef]
- Poothong, S.; Reed, B.M. Increased CaCl2, MgSO4, and KH2PO4 Improve the Growth of Micropropagated Red Raspberries. In Vitro Cell. Dev. Biol. Plant 2015, 51, 648–658. [Google Scholar] [CrossRef]
- Kovalchuk, I.Y.; Mukhitdinova, Z.; Turdiyev, T.; Madiyeva, G.; Akin, M.; Eyduran, E.; Reed, B.M. Nitrogen Ions and Nitrogen Ion Proportions Impact the Growth of Apricot (Prunus armeniaca) Shoot Cultures. Plant Cell Tissue Organ Cult. 2018, 133, 263–273. [Google Scholar] [CrossRef]
- Akin, M.; Eyduran, E.; Niedz, R.P.; Reed, B.M. Developing Hazelnut Tissue Culture Medium Free of Ion Confounding. Plant Cell Tissue Organ Cult. 2017, 130, 483–494. [Google Scholar] [CrossRef]
- Akin, M.; Eyduran, E.; Reed, B.M. Use of RSM and CHAID Data Mining Algorithm for Predicting Mineral Nutrition of Hazelnut. Plant Cell Tissue Organ Cult. 2017, 128, 303–316. [Google Scholar] [CrossRef]
- Landín, M.; Rowe, R.C.; York, P. Advantages of Neurofuzzy Logic against Conventional Experimental Design and Statistical Analysis in Studying and Developing Direct Compression Formulations. Eur. J. Pharm. Sci. 2009, 38, 325–331. [Google Scholar] [CrossRef]
- Gago, J.; Martínez-Núñez, L.; Landín, M.; Gallego, P.P. Artificial Neural Networks as an Alternative to the Traditional Statistical Methodology in Plant Research. J. Plant Physiol. 2010, 167, 23–27. [Google Scholar] [CrossRef]
- Nezami-Alanagh, E.; Garoosi, G.-A.; Maleki, S.; Landín, M.; Gallego, P.P. Predicting Optimal In Vitro Culture Medium for Pistacia vera Micropropagation Using Neural Networks Models. Plant Cell Tissue Organ Cult. 2017, 129, 19–33. [Google Scholar] [CrossRef]
- Gallego, P.P.; Gago, J.; Landin, M. Artificial Neural Networks Technology to Model and Predict Plant Biology Process. In Artificial Neural Networks—Methodological Advances and Biomedical Applications; Suzuki, K., Ed.; IntechOpen: Rijeka, Croatia, 2011; pp. 197–216. ISBN 978-953-307-243-2. [Google Scholar]
- Gago, J.; Pérez-Tornero, O.; Landín, M.; Burgos, L.; Gallego, P.P. Improving Knowledge of Plant Tissue Culture and Media Formulation by Neurofuzzy Logic: A Practical Case of Data Mining Using Apricot Databases. J. Plant Physiol. 2011, 168, 1858–1865. [Google Scholar] [CrossRef] [PubMed]
- Gago, J.; Landín, M.; Gallego, P.P. Artificial Neural Networks Modeling the In Vitro Rhizogenesis and Acclimatization of Vitis vinifera L. J. Plant Physiol. 2010, 167, 1226–1231. [Google Scholar] [CrossRef] [PubMed]
- Landin, M.; Rowe, R.C. Artificial Neural Networks Technology to Model, Understand, and Optimize Drug Formulations. In Formulation Tools for Pharmaceutical Development; Aguilar, J.E., Ed.; Elsevier: Cambridge, UK, 2013; pp. 7–37. ISBN 978-1-907568-99-2. [Google Scholar]
- Nezami-Alanagh, E.; Garoosi, G.-A.; Landín, M.; Gallego, P.P. Computer-Based Tools Provide New Insight into the Key Factors That Cause Physiological Disorders of Pistachio Rootstocks Cultured In Vitro. Sci. Rep. 2019, 9, 9740. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Fitzpatrick, T.B.; Chapman, L.M. The Importance of Thiamine (Vitamin B1) in Plant Health: From Crop Yield to Biofortification. J. Biol. Chem. 2020, 295, 12002–12013. [Google Scholar] [CrossRef]
- Arteta, T.A.; Hameg, R.; Landin, M.; Gallego, P.P.; Barreal, M.E. Deciphering the Effect of Vitamins and Mineral Nutrients on Kiwiberry Micropropagation Using Computer-Based Tools. Acta Hortic. 2022, 1332, 31–38. [Google Scholar] [CrossRef]
- Roberts, J.A.; Hooley, R. Introduction—The Challenge of PGR Research. In Plant Growth Regulators; Roberts, J.A., Hooley, R., Eds.; Springer: Boston, MA, USA, 1988; pp. 1–3. ISBN 978-1-4615-7594-8. [Google Scholar]
- Revathi, J.; Manokari, M.; Priyadharshini, S.; Shekhawat, M.S. Effects of Plant Growth Regulators on in Vitro Morphogenic Response in Oldenlandia herbacea (L.) Roxb. Vegetos 2020, 33, 800–804. [Google Scholar] [CrossRef]
- Abdullah, M.; Sliwinska, E.; Góralski, G.; Latocha, P.; Tuleja, M.; Widyna, P.; Popielarska-Konieczna, M. Effect of Medium Composition, Genotype and Age of Explant on the Regeneration of Hexaploid Plants from Endosperm Culture of Tetraploid Kiwiberry (Actinidia arguta). Plant Cell Tissue Organ Cult. 2021, 147, 569–582. [Google Scholar] [CrossRef]
- Zhou, R.J.; Liu, M.J. Effect of Plant Growth Regulators on Tissue Culture in Chinese Jujube. Acta Hortic. 2009, 840, 309–314. [Google Scholar] [CrossRef]
- Bhojwani, S.S.; Dantu, P.K. Micropropagation. In Plant Tissue Culture: An Introductory Text; Springer: New Delhi, India, 2013; pp. 245–274. ISBN 978-81-322-1025-2. [Google Scholar]
- Nezami-Alanagh, E.; Garoosi, G.-A.; Landín, M.; Gallego, P.P. Combining DOE with Neurofuzzy Logic for Healthy Mineral Nutrition of Pistachio Rootstocks In Vitro Culture. Front. Plant Sci. 2018, 9, 1474. [Google Scholar] [CrossRef] [PubMed]
- Moncaleán, P.; Cañal, M.J.; Feito, I.; Rodríguez, A.; Fernández, B. Cytokinins and Mineral Nutrition in Actinidia deliciosa (Kiwi) Shoots Cultured In Vitro. J. Plant Physiol. 1999, 155, 606–612. [Google Scholar] [CrossRef]
- Moncaleán, P.; Cañal, M.J.; Fernández, H.; Fernández, B.; Rodríguez, A. Nutritional and Gibberellic Acid Requirements in Kiwifruit Vitroponic Cultures. In Vitro Cell. Dev. Biol. Plant 2003, 39, 49–55. [Google Scholar] [CrossRef]
- Monette, P.L. Micropropagation of Kiwifruit Using Non-Axenic Shoot Tips. Plant Cell Tissue Organ Cult. 1986, 6, 73–82. [Google Scholar] [CrossRef]
- Akbas, F.A.; Isikalan, C.; Namli, S.; Basaran, D. Micropropagation of Kiwifruit (Actinidia deliciosa). Int. J. Agric. Biol. 2007, 9, 489–493. [Google Scholar]
- Nasib, A.; Ali, K.; Khan, S. An Optimized and Improved Method for the In Vitro Propagation of Kiwifruit (Actinidia deliciosa) Using Coconut Water. Pak. J. Bot. 2008, 40, 2355–2360. [Google Scholar]
- White, P.R. Plant Tissue Cultures. Sci. Am. 1950, 182, 48–51. [Google Scholar] [CrossRef]
- Niedz, R.P.; Evens, T.J. Design of Experiments (DOE)—History, Concepts, and Relevance to in Vitro Culture. In Vitro Cell. Dev. Biol. Plant 2016, 52, 547–562. [Google Scholar] [CrossRef]
- Niedz, R.P.; Evens, T.J. A Solution to the Problem of Ion Confounding in Experimental Biology. Nat. Methods 2006, 3, 417. [Google Scholar] [CrossRef]
- Niedz, R.P.; Evens, T.J. Regulating Plant Tissue Growth by Mineral Nutrition. In Vitro Cell. Dev. Biol. Plant 2007, 43, 370–381. [Google Scholar] [CrossRef]
- Silvestri, C.; Rugini, E.; Cristofori, V. The Effect of CuSO4 for Establishing In Vitro Culture, and the Role Nitrogen and Iron Sources in In Vitro Multiplication of Corylus avellana L. Cv. Tonda Gentile Romana. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2020, 154, 17–23. [Google Scholar] [CrossRef]
- Fries, L. Vitamin B12 in Pisum sativum (L.). Physiol. Plant. 1962, 15, 566–571. [Google Scholar] [CrossRef]
- Chauhan, M.; Kothari, S.L. Optimization of Nutrient Levels in the Medium Increases the Efficiency of Callus Induction and Plant Regeneration in Recalcitrant Indian Barley (Hordeum vulgare L.) In Vitro. In Vitro Cell. Dev. Biol. Plant 2004, 40, 520–527. [Google Scholar] [CrossRef]
- Sonnewald, U. Physiology of Metabolism. In Strasburger’s Plant Sciences; Bresinsky, A., Körner, C., Kadereit, J.W., Neuhaus, G., Sonnewald, U., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 239–409. ISBN 978-3-642-15517-8. [Google Scholar]
- Ibrahim, I.A.; Nasr, M.I.; Mohammedm, B.R.; El-Zefzafi, M.M. Nutrient Factors Affecting in Vitro Cultivation of Stevia rebaudiana. Sugar Tech 2008, 10, 248–253. [Google Scholar] [CrossRef]
- Poothong, S.; Reed, B.M. Modeling the Effects of Mineral Nutrition for Improving Growth and Development of Micropropagated Red Raspberries. Sci. Hortic. 2014, 165, 132–141. [Google Scholar] [CrossRef]
- Hunková, J.; Gajdošová, A.; Szabóová, M. Effect of Mesos Components (MgSO4, CaCl2, KH2PO4) on In Vitro Shoot Growth of Blackberry, Blueberry, and Saskatoon. Plants 2020, 9, 935. [Google Scholar] [CrossRef]
- Reed, B.M.; Wada, S.; DeNoma, J.; Niedz, R.P. Mineral Nutrition Influences Physiological Responses of Pear In Vitro. In Vitro Cell. Dev. Biol. Plant 2013, 49, 699–709. [Google Scholar] [CrossRef]
- Akin, M.; Eyduran, S.P.; Eyduran, E.; Reed, B.M. Analysis of Macro Nutrient Related Growth Responses Using Multivariate Adaptive Regression Splines. Plant Cell Tissue Organ Cult. 2020, 140, 661–670. [Google Scholar] [CrossRef]
- Eaton, F.M. Deficiency, Toxicity, and Accumulation of Boron in Plants. J. Agric. Res. 1944, 69, 237–277. [Google Scholar]
- Brdar-Jokanović, M. Boron Toxicity and Deficiency in Agricultural Plants. Int. J. Mol. Sci. 2020, 21, 1424. [Google Scholar] [CrossRef] [Green Version]
- Dalton, C.C.; Iqbal, K.; Turner, D.A. Iron Phosphate Precipitation in Murashige and Skoog Media. Physiol. Plant. 1983, 57, 472–476. [Google Scholar] [CrossRef]
- Kothari, S.L.; Agarwal, K.; Kumar, S. Inorganic Nutrient Manipulation for Highly Improved in Vitro Plant Regeneration in Finger Millet—Eleusine coracana (L.) Gaertn. In Vitro Cell. Dev. Biol. Plant 2004, 40, 515–519. [Google Scholar] [CrossRef]
- Niedz, R.P.; Hyndman, S.E.; Evens, T.J.; Weathersbee, A.A. Mineral Nutrition and in Vitro Growth of Gerbera hybrida (Asteraceae). In Vitro Cell. Dev. Biol. Plant 2014, 50, 458–470. [Google Scholar] [CrossRef]
- Rinallo, C.; Modi, G. Content of Oxalate in Actinidia deliciosa Plants Grown in Nutrient Solutions with Different Nitrogen Forms. Biol. Plant. 2002, 45, 137–139. [Google Scholar] [CrossRef]
- Antonopoulou, C.; Dimassi, K.; Therios, I.; Chatzissavvidis, C.; Papadakis, I. The Effect of Fe-EDDHA and of Ascorbic Acid on in Vitro Rooting of the Peach Rootstock GF-677 Explants. Acta Physiol. Plant. 2007, 29, 559–561. [Google Scholar] [CrossRef]
- Garrison, W.; Dale, A.; Saxena, P.K. Improved Shoot Multiplication and Development in Hybrid Hazelnut Nodal Cultures by Ethylenediamine Di-2-Hydroxy-Phenylacetic Acid (Fe-EDDHA). Can. J. Plant Sci. 2013, 93, 511–521. [Google Scholar] [CrossRef]
- Greenboim-Wainberg, Y.; Maymon, I.; Borochov, R.; Alvarez, J.; Olszewski, N.; Ori, N.; Eshed, Y.; Weiss, D. Cross Talk between Gibberellin and Cytokinin: The Arabidopsis GA Response Inhibitor SPINDLY Plays a Positive Role in Cytokinin Signaling. Plant Cell 2005, 17, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Jasinski, S.; Piazza, P.; Craft, J.; Hay, A.; Woolley, L.; Rieu, I.; Phillips, A.; Hedden, P.; Tsiantis, M. KNOX Action in Arabidopsis Is Mediated by Coordinate Regulation of Cytokinin and Gibberellin Activities. Curr. Biol. 2005, 15, 1560–1565. [Google Scholar] [CrossRef] [Green Version]
- Poddar, K.; Vishnoi, R.K.; Kothari, S.L. Plant Regeneration from Embryogenic Callus of Finger Millet Eleusine coracana (L.) Gaertn. on Higher Concentrations of NH4NO3 as a Replacement of NAA in the Medium. Plant Sci. 1997, 129, 101–106. [Google Scholar] [CrossRef]
- Kothari-Chajer, A.; Sharma, M.; Kachhwaha, S.; Kothari, S.L. Micronutrient Optimization Results into Highly Improved in Vitro Plant Regeneration in Kodo (Paspalum scrobiculatum L.) and Finger (Eleusine coracana (L.) Gaertn.) Millets. Plant Cell Tissue Organ Cult. 2008, 94, 105–112. [Google Scholar] [CrossRef]
- Ivanova, M.; van Staden, J. Effect of Ammonium Ions and Cytokinins on Hyperhydricity and Multiplication Rate of In Vitro Regenerated Shoots of Aloe polyphylla. Plant Cell Tissue Organ Cult. 2008, 92, 227–231. [Google Scholar] [CrossRef]
- Sreelekshmi, R.; Siril, E.A. Effective Reversal of Hyperhydricity Leading to Efficient Micropropagation of Dianthus chinensis L. 3 Biotech 2021, 11, 95. [Google Scholar] [CrossRef] [PubMed]
- Munné-Bosch, S.; Alegre, L. The Function of Tocopherols and Tocotrienols in Plants. Crit. Rev. Plant Sci. 2002, 21, 31–57. [Google Scholar] [CrossRef]
- Saad, A.I.M.; Elshahed, A.M. Plant Tissue Culture Media. In Recent Advances in Plant In Vitro Culture; Leva, A., Rinaldi, L., Eds.; IntecOpen: London, UK, 2012; ISBN 978-953-51-0787-3. [Google Scholar]
- Cheng, T.-Y. Adventitious Bud Formation in Culture of Douglas Fir (Pseudotsuga menziesii (MIRB.) Franco). Plant Sci. Lett. 1975, 5, 97–102. [Google Scholar] [CrossRef]
- Design-Expert, Stat-Ease Inc.: Minneapois, MI, USA, 2010.
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- FormRules; Intelligensys Ltd.: Stokesley, UK, 2008.
- Shao, Q.; Rowe, R.C.; York, P. Comparison of Neurofuzzy Logic and Neural Networks in Modelling Experimental Data of an Immediate Release Tablet Formulation. Eur. J. Pharm. Sci. 2006, 28, 394–404. [Google Scholar] [CrossRef]
- García-Pérez, P.; Lozano-Milo, E.; Landín, M.; Gallego, P.P. Combining Medicinal Plant in Vitro Culture with Machine Learning Technologies for Maximizing the Production of Phenolic Compounds. Antioxidants 2020, 9, 210. [Google Scholar] [CrossRef] [Green Version]
- Kavli, T.; Weyer, E. On ASMOD—An Algorithm for Empirical Modelling Using Spline Functions. In Neural Network Engineering in Dynamic Control Systems; Hunt, K.J., Irwin, G.R., Warwick, K., Eds.; Advances in Industrial Control; Springer: London, UK, 1995; pp. 83–104. ISBN 978-1-4471-3068-0. [Google Scholar]
- Hesami, M.; Jones, A.M.P. Application of Artificial Intelligence Models and Optimization Algorithms in Plant Cell and Tissue Culture. Appl. Microbiol. Biotechnol. 2020, 104, 9449–9485. [Google Scholar] [CrossRef]
- Vapnik, V.N. The Nature of Statistical Learning Theory, 2nd ed.; Statistics for Engineering and Information Science; Springer: New York, NY, USA, 2010; ISBN 978-1-4419-3160-3. [Google Scholar]
- Chen, Q.; Mynett, A.E. Integration of Data Mining Techniques and Heuristic Knowledge in Fuzzy Logic Modelling of Eutrophication in Taihu Lake. Ecol. Model. 2003, 162, 55–67. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Outputs | Submodel | Train Set R2 (%) | f-Ratio | df1 | df2 | f-Critical (α = 0.01) | Critical Factors |
|---|---|---|---|---|---|---|---|
| SN | 1 | 82.3 | 19.14 | 17 | 87 | 2.18 | Fe2+ × Na+ |
| 2 | GA3 | ||||||
| 3 | K+ × SO42− | ||||||
| 4 | BAP | ||||||
| SL | 1 | 70.3 | 7.56 | 20 | 84 | 2.10 | Na+− |
| 2 | Mg2+ | ||||||
| 3 | NO3− × K+ | ||||||
| 4 | Vitamin E | ||||||
| 5 | BO3− | ||||||
| 6 | GA3 × BAP | ||||||
| 7 | Co2+ | ||||||
| 8 | Myo-inositol | ||||||
| LA | 1 | 77.7 | 38.34 | 7 | 84 | 2.86 | Na+ |
| 2 | GA3 | ||||||
| 3 | K+ × NO3− | ||||||
| 4 | SO42− | ||||||
| SQ | 1 | 85.6 | 49.47 | 9 | 84 | 2.63 | NO3− |
| 2 | K+ | ||||||
| 3 | NH4+ | ||||||
| 4 | Fe2+ | ||||||
| 5 | MoO42− | ||||||
| 6 | BAP | ||||||
| BC | 1 | 96.0 | 120.91 | 14 | 84 | 2.30 | PO43− × NH4+ |
| 2 | SO42− | ||||||
| H | 1 | 84.4 | 19.76 | 18 | 84 | 2.16 | Co2+ × NH4+ |
| 2 | I− | ||||||
| 3 | SO42− × NO3− | ||||||
| 4 | Ca2+ × Fe2+ | ||||||
| 5 | BAP |
| Rules | [NO3−] | [K+] | [Na+] | [SO42−] | [Fe2+] | [BO3−] | [Mg2+] | Vit E | [Co2+] | Myo | BAP | GA3 | SN | SL | LA | MD | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Low | Low | High | 1.00 | ||||||||||||||
| 2 | High | Low | Low | 1.00 | ||||||||||||||
| 3 | Low | Mid | High | 1.00 | ||||||||||||||
| 4 | High | Mid | Low | 1.00 | ||||||||||||||
| 5 | Low | High | High | 1.00 | ||||||||||||||
| 6 | High | High | High | 0.79 | ||||||||||||||
| 7 | Low | Low | 1.00 | |||||||||||||||
| 8 | Mid | Low | 1.00 | |||||||||||||||
| 9 | IF | High | THEN | High | 0.58 | |||||||||||||
| 10 | Low | Low | Low | 1.00 | ||||||||||||||
| 11 | Low | Mid | Low | 1.00 | ||||||||||||||
| 12 | Low | High | High | 1.00 | ||||||||||||||
| 13 | Mid | Low | Low | 0.75 | ||||||||||||||
| 14 | Mid | Mid | High | 1.00 | ||||||||||||||
| 15 | Mid | High | Low | 1.00 | ||||||||||||||
| 16 | High | Low | High | 1.00 | ||||||||||||||
| 17 | High | Mid | Low | 1.00 | ||||||||||||||
| 18 | High | High | Low | 1.00 | ||||||||||||||
| 19 | Low | Low | 1.00 | |||||||||||||||
| 20 | High | Low | 0.80 | |||||||||||||||
| 21 | Low | High | 1.00 | |||||||||||||||
| 22 | Mid | High | 1.00 | |||||||||||||||
| 23 | High | Low | 1.00 | |||||||||||||||
| 24 | Low | Low | 1.00 | |||||||||||||||
| 25 | High | High | 1.00 | |||||||||||||||
| 26 | Low | Low | Low | 1.00 | ||||||||||||||
| 27 | Low | High | High | 1.00 | ||||||||||||||
| 28 | High | Low | High | 1.00 | ||||||||||||||
| 29 | High | High | Low | 1.00 | ||||||||||||||
| 30 | Low | High | 0.94 | |||||||||||||||
| 31 | IF | High | THEN | Low | 0.91 | |||||||||||||
| 32 | Low | Low | 1.00 | |||||||||||||||
| 33 | Mid | High | 1.00 | |||||||||||||||
| 34 | High | High | 1.00 | |||||||||||||||
| 35 | Low_1 | Low | High | 1.00 | ||||||||||||||
| 36 | Mid_2 | Low | Low | 1.00 | ||||||||||||||
| 37 | Mid_3 | Low | Low | 1.00 | ||||||||||||||
| 38 | High_4 | Low | Low | 1.00 | ||||||||||||||
| 39 | Low_1 | High | Low | 1.00 | ||||||||||||||
| 40 | Mid_2 | High | High | 1.00 | ||||||||||||||
| 41 | Mid_3 | High | Low | 0.50 | ||||||||||||||
| 42 | High_4 | High | High | 1.00 | ||||||||||||||
| 43 | Low | High | 1.00 | |||||||||||||||
| 44 | Mid | Low | 1.00 | |||||||||||||||
| 45 | High | Low | 1.00 | |||||||||||||||
| 46 | Low | High | 0.83 | |||||||||||||||
| 47 | High | Low | 0.79 | |||||||||||||||
| 48 | Low | High | 1.00 | |||||||||||||||
| 49 | High | Low | 1.00 | |||||||||||||||
| 50 | Low | High | 0.97 | |||||||||||||||
| 51 | High | Low | 1.00 | |||||||||||||||
| 52 | IF | Low | Low | THEN | Low | 1.00 | ||||||||||||
| 53 | High | Low | High | 1.00 | ||||||||||||||
| 54 | Low | High | Low | 0.72 | ||||||||||||||
| 55 | High | High | High | 0.57 | ||||||||||||||
| 56 | Low | Low | 1.00 | |||||||||||||||
| 57 | High | High | 1.00 |
| Rules | [NO3−] | [NH4+] | [K+] | [SO42−] | [Ca2+] | [Co2+] | [I−] | [Fe2+] | [MoO42−] | [PO43−] | BAP | SQ | BC | H | MD | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Low | High | 1.00 | ||||||||||||||
| 2 | High | Low | 1.00 | ||||||||||||||
| 3 | Low | Low | 1.00 | ||||||||||||||
| 4 | High | High | 1.00 | ||||||||||||||
| 5 | Low | Low | 1.00 | ||||||||||||||
| 6 | High | High | 1.00 | ||||||||||||||
| 7 | IF | Low | THEN | Low | 1.00 | ||||||||||||
| 8 | Mid | High | 1.00 | ||||||||||||||
| 9 | High | Low | 1.00 | ||||||||||||||
| 10 | Low | High | 1.00 | ||||||||||||||
| 11 | Mid | High | 1.00 | ||||||||||||||
| 12 | High | Low | 1.00 | ||||||||||||||
| 13 | Low | High | 0.93 | ||||||||||||||
| 14 | High | Low | 1.00 | ||||||||||||||
| 15 | Low | Low_1 | Low | 1.00 | |||||||||||||
| 16 | Mid | Low_1 | Low | 1.00 | |||||||||||||
| 17 | High | Low_1 | Low | 1.00 | |||||||||||||
| 18 | Low | Mid_2 | Low | 0.58 | |||||||||||||
| 19 | Mid | Mid_2 | Low | 1.00 | |||||||||||||
| 20 | High | Mid_2 | Low | 0.97 | |||||||||||||
| 21 | Low | Mid_3 | High | 1.00 | |||||||||||||
| 22 | IF | Mid | Mid_3 | THEN | High | 1.00 | |||||||||||
| 23 | High | Mid_3 | High | 1.00 | |||||||||||||
| 24 | Low | High_4 | High | 1.00 | |||||||||||||
| 25 | Mid | High_4 | High | 1.00 | |||||||||||||
| 26 | High | High_4 | High | 1.00 | |||||||||||||
| 27 | Low | High | 1.00 | ||||||||||||||
| 28 | Mid | High | 0.52 | ||||||||||||||
| 29 | High | High | 0.78 | ||||||||||||||
| 30 | Low | Low | 1.00 | ||||||||||||||
| 31 | High | THEN | High | 1.00 | |||||||||||||
| 32 | Low | Low | Low | 1.00 | |||||||||||||
| 33 | High | Low | Low | 1.00 | |||||||||||||
| 34 | Low | Mid | Low | 1.00 | |||||||||||||
| 35 | High | Mid | Low | 1.00 | |||||||||||||
| 36 | Low | High | High | 1.00 | |||||||||||||
| 37 | High | High | High | 1.00 | |||||||||||||
| 38 | Low_1 | Low | High | 1.00 | |||||||||||||
| 39 | IF | Low_1 | High | THEN | High | 1.00 | |||||||||||
| 40 | Mid_2 | Low | High | 1.00 | |||||||||||||
| 41 | Mid_2 | High | High | 1.00 | |||||||||||||
| 42 | Mid_3 | Low | Low | 1.00 | |||||||||||||
| 43 | Mid_3 | High | Low | 1.00 | |||||||||||||
| 44 | High_4 | Low | Low | 1.00 | |||||||||||||
| 45 | High_4 | High | Low | 1.00 | |||||||||||||
| 46 | Low | High | 0.75 | ||||||||||||||
| 47 | High | Low | 1.00 | ||||||||||||||
| 48 | Low | Low | High | 1.00 | |||||||||||||
| 49 | High | Low | High | 1.00 | |||||||||||||
| 50 | Low | High | Low | 1.00 | |||||||||||||
| 51 | High | High | Low | 1.00 |
| Input | Level | Range |
|---|---|---|
| NH4+ (mM) | High | 12.37–20.61 |
| NO3− (mM) | Mid–High | 14.35–39.41 |
| K+ (mM) | Mid | 7.28–17.46 |
| Ca2+ (mM) | Low–Mid_2 | 0.75–5.89 |
| Mg2+ (mM) | High | 2.44–4.50 |
| PO43− (mM) | Mid_3–High_4 | 1.60–3.75 |
| SO42− (mM) | High | 2.85–5.20 |
| Fe2+ (mM) | Low | 0.10–0.30 |
| BO3− (mM) | Mid–High | 0.05–0.15 |
| MoO42− (mM) | Mid | 0.0005–0.0012 |
| Na+ (mM) | Low | 0.20–0.60 |
| Co2+ (mM) | Low | 0.00001–0.00008 |
| I− (mM) | High | 0.0040–0.0075 |
| Myo (mg L−1) | Low | 0–500 |
| Vit. E (mg L−1) | Low | 0.00–0.50 |
| GA3 (mg L−1) | Low | 0.00–0.50 |
| BAP (mg L−1) | Low | 0.50–1.50 |
| Mineral Nutrient Factors | Media Salts | Range (× MS) |
|---|---|---|
| Factor 1 | NH4NO3 | 0.2–1× |
| Factor 2 | KNO3 | 0.1–1× |
| Factor 3 (Mesos) | CaCl2·2H2O | 0.25–3× |
| MgSO4·7H2O | ||
| KH2PO4 | ||
| Factor 4 (Micros) | MnSO4·4H2O | 0.1–1.5× |
| ZnSO4·7H2O | ||
| H3BO3 | ||
| KI | ||
| CuSO4·5H2O | ||
| Na2MoO4·2H2O | ||
| CoCl2·6H2O | ||
| Factor 5 (Iron) | FeSO4·7H2O | 1–5× |
| Na2·EDTA | ||
| Vitamin Factors | Vitamins | Range (× MS) |
| Factor 1 | Myo-inositol | 0–10× |
| Factor 2 | Thiamine | 0–10× |
| Factor 3 | Nicotinic acid | 0–10× |
| Factor 4 | Pyridoxine | 0–3× |
| Factor 5 | Vitamin E | – 1 |
| FormRules® v4.03 |
|---|
| Minimization parameters (ASMOD) |
| Ridge Regression Factor: 1 × 10−6 |
| Model Selection Criteria |
| Structural Risk Minimization (SRM) |
| C1LA, SQ, BC = 0.970 |
| C1SN, H = 0.868 |
| C1SL = 0.750 |
| C2 = 4.8 |
| Number of Set Densities: 2 |
| Set Densities: 2, 3 |
| Adapt Nodes: TRUE |
| Max. Inputs Per SubModel: 2 |
| Max. Nodes Per Input: 15 |
| Minimization parameters (ASMOD) |
| Ridge Regression Factor: 1 × 10−6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arteta, T.A.; Hameg, R.; Landin, M.; Gallego, P.P.; Barreal, M.E. Artificial Neural Networks Elucidated the Essential Role of Mineral Nutrients versus Vitamins and Plant Growth Regulators in Achieving Healthy Micropropagated Plants. Plants 2022, 11, 1284. https://doi.org/10.3390/plants11101284
Arteta TA, Hameg R, Landin M, Gallego PP, Barreal ME. Artificial Neural Networks Elucidated the Essential Role of Mineral Nutrients versus Vitamins and Plant Growth Regulators in Achieving Healthy Micropropagated Plants. Plants. 2022; 11(10):1284. https://doi.org/10.3390/plants11101284
Chicago/Turabian StyleArteta, Tomás A., Radhia Hameg, Mariana Landin, Pedro P. Gallego, and M. Esther Barreal. 2022. "Artificial Neural Networks Elucidated the Essential Role of Mineral Nutrients versus Vitamins and Plant Growth Regulators in Achieving Healthy Micropropagated Plants" Plants 11, no. 10: 1284. https://doi.org/10.3390/plants11101284
APA StyleArteta, T. A., Hameg, R., Landin, M., Gallego, P. P., & Barreal, M. E. (2022). Artificial Neural Networks Elucidated the Essential Role of Mineral Nutrients versus Vitamins and Plant Growth Regulators in Achieving Healthy Micropropagated Plants. Plants, 11(10), 1284. https://doi.org/10.3390/plants11101284