
N-Acetylcysteine Priming Alleviates the Transplanting Injury of Machine-Transplanted Rice by Comprehensively Promoting Antioxidant and Photosynthetic Systems


 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Phenotype, Yield, and Yield Components
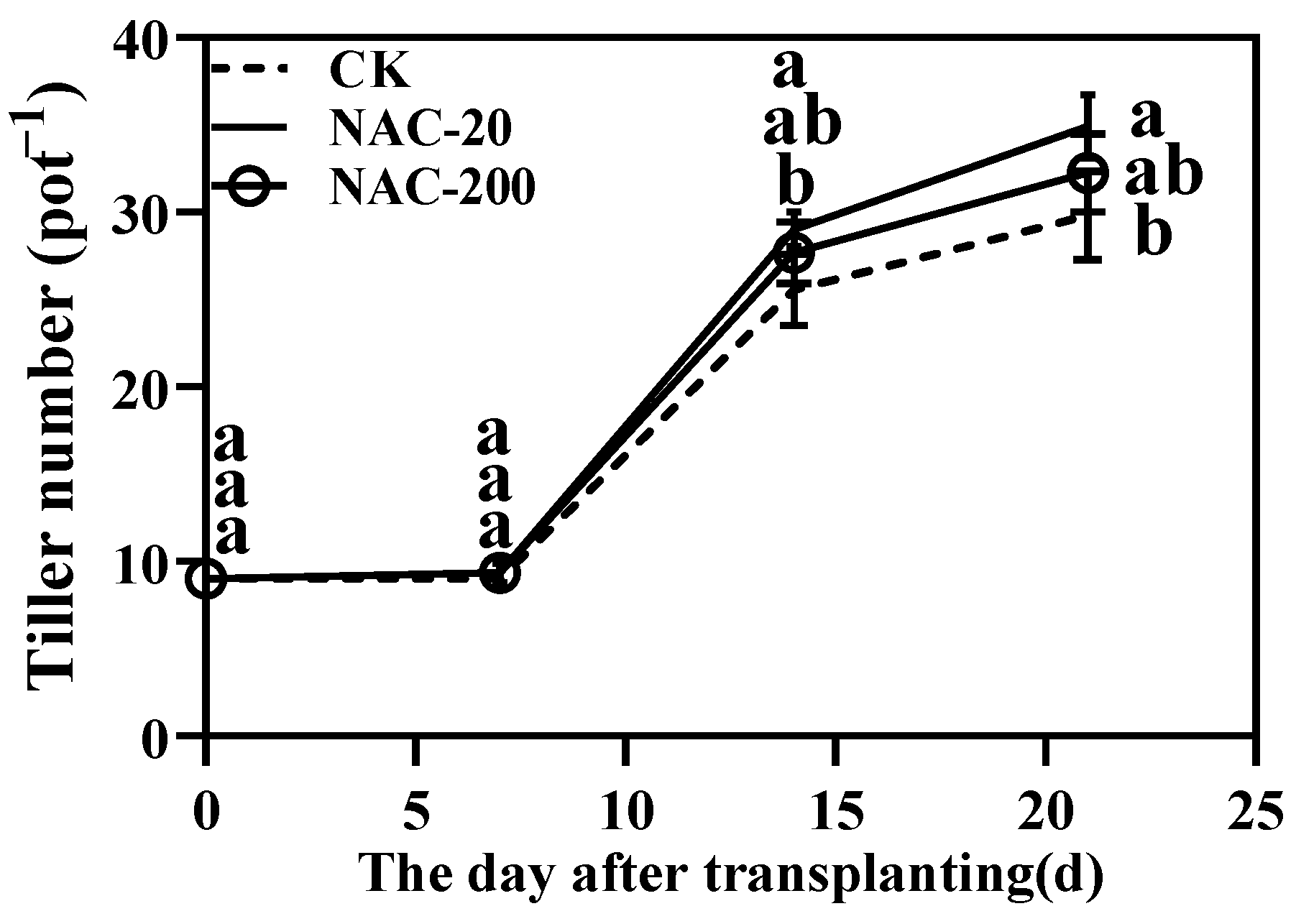
2.2. Tiller Dynamics
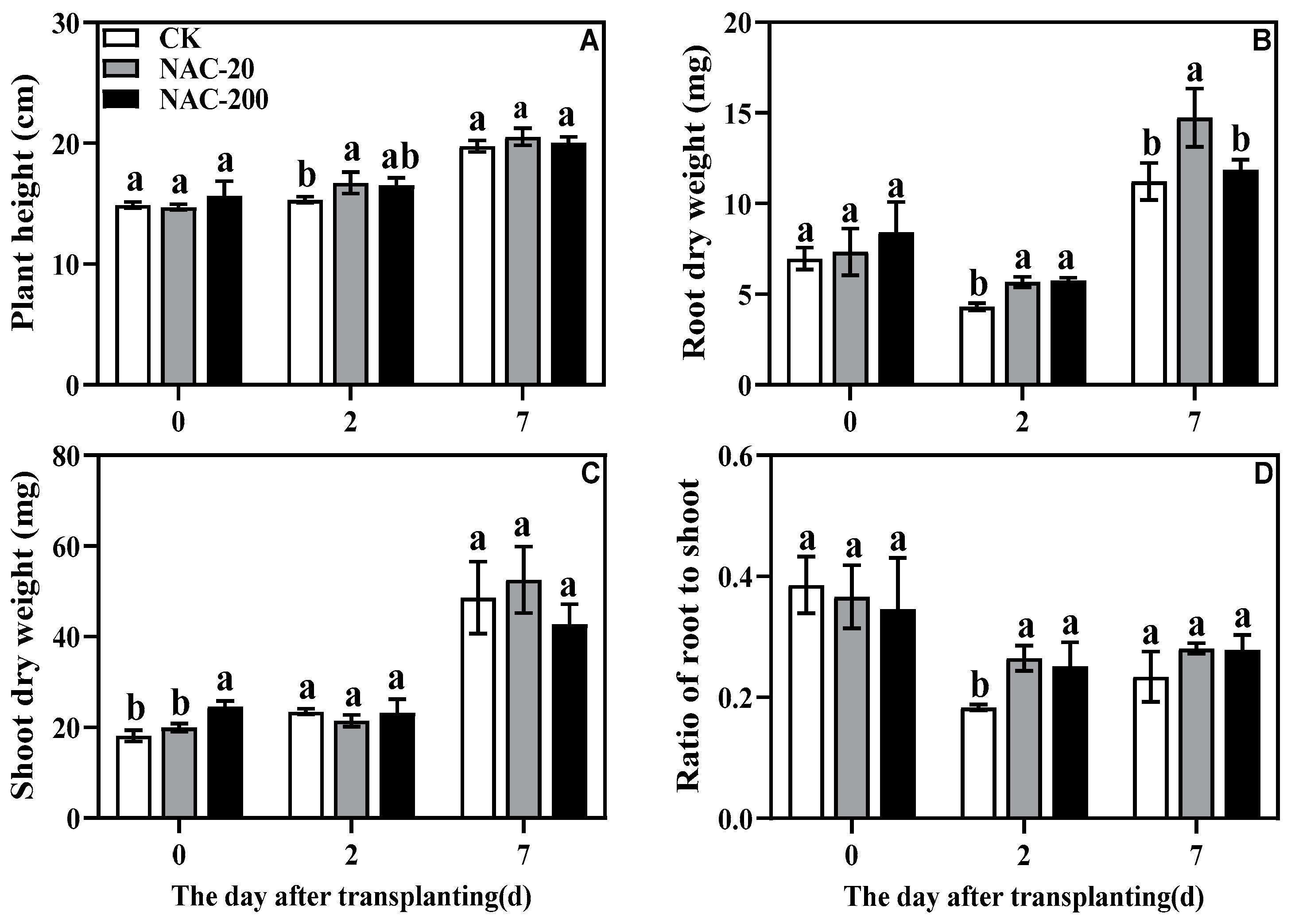
2.3. Plant Growth Parameters
2.4. Chloroplast Pigment Content
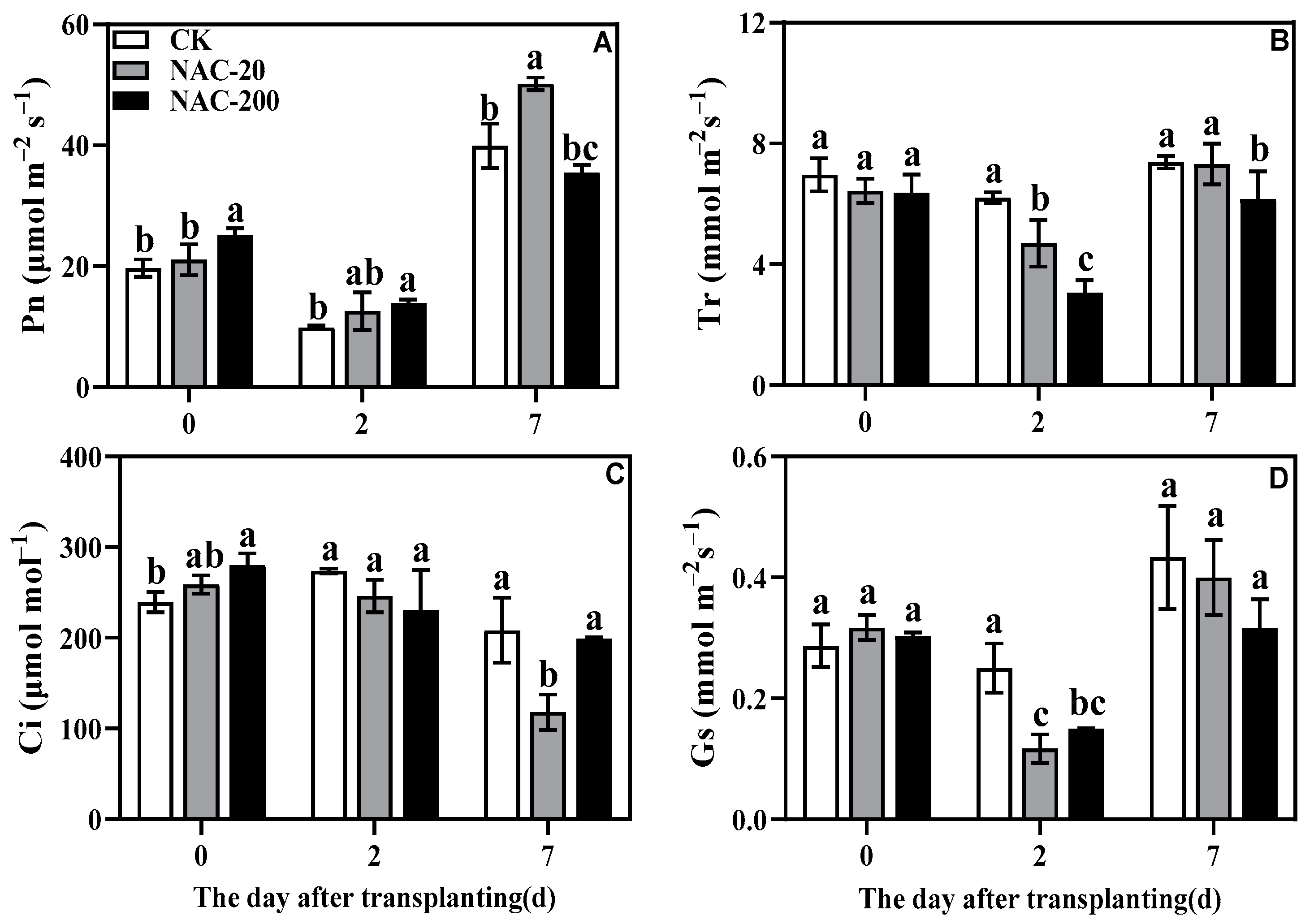
2.5. Gas Exchange Parameters
2.6. Sucrose Content
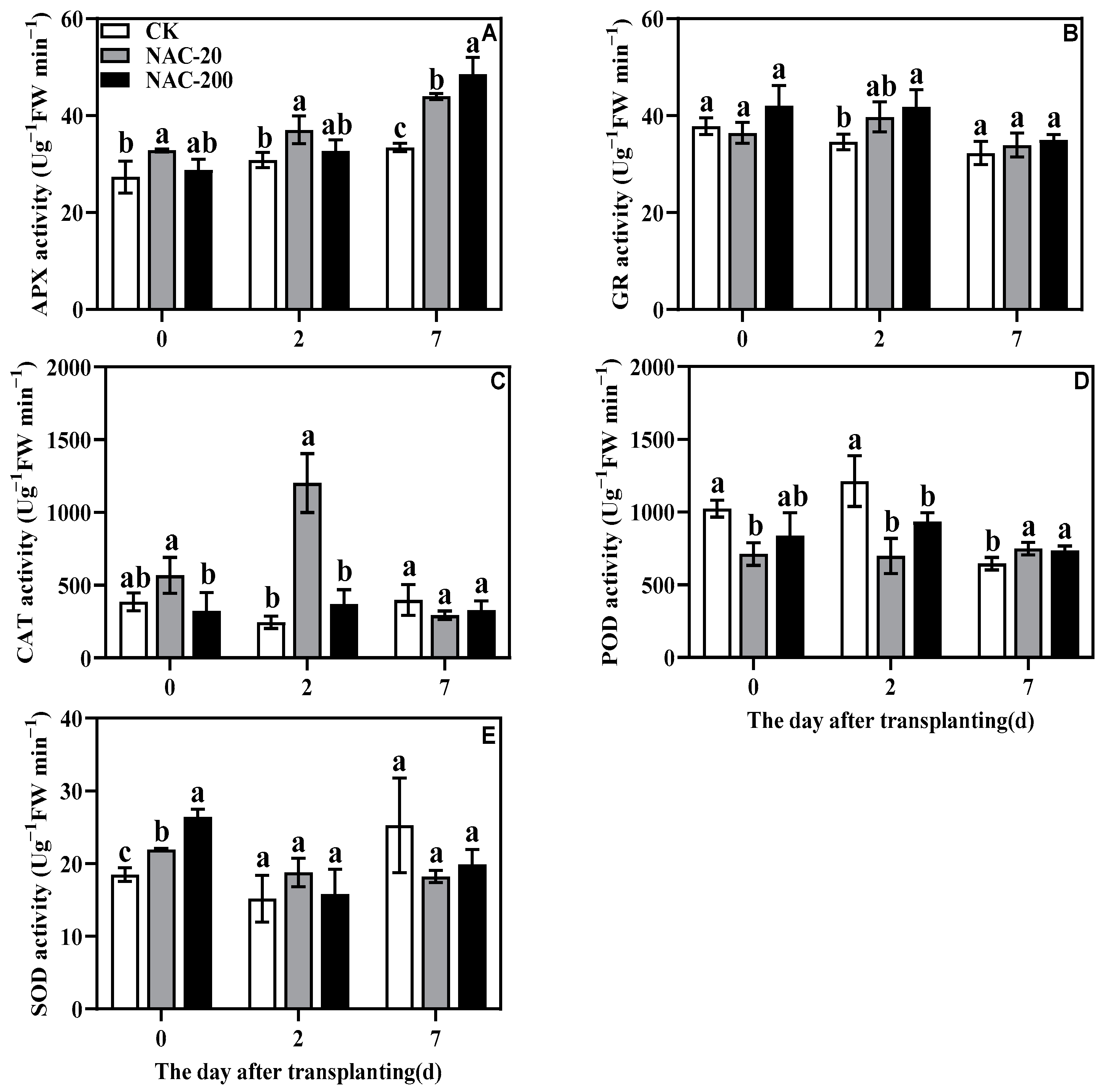
2.7. Antioxidant Enzyme Activity
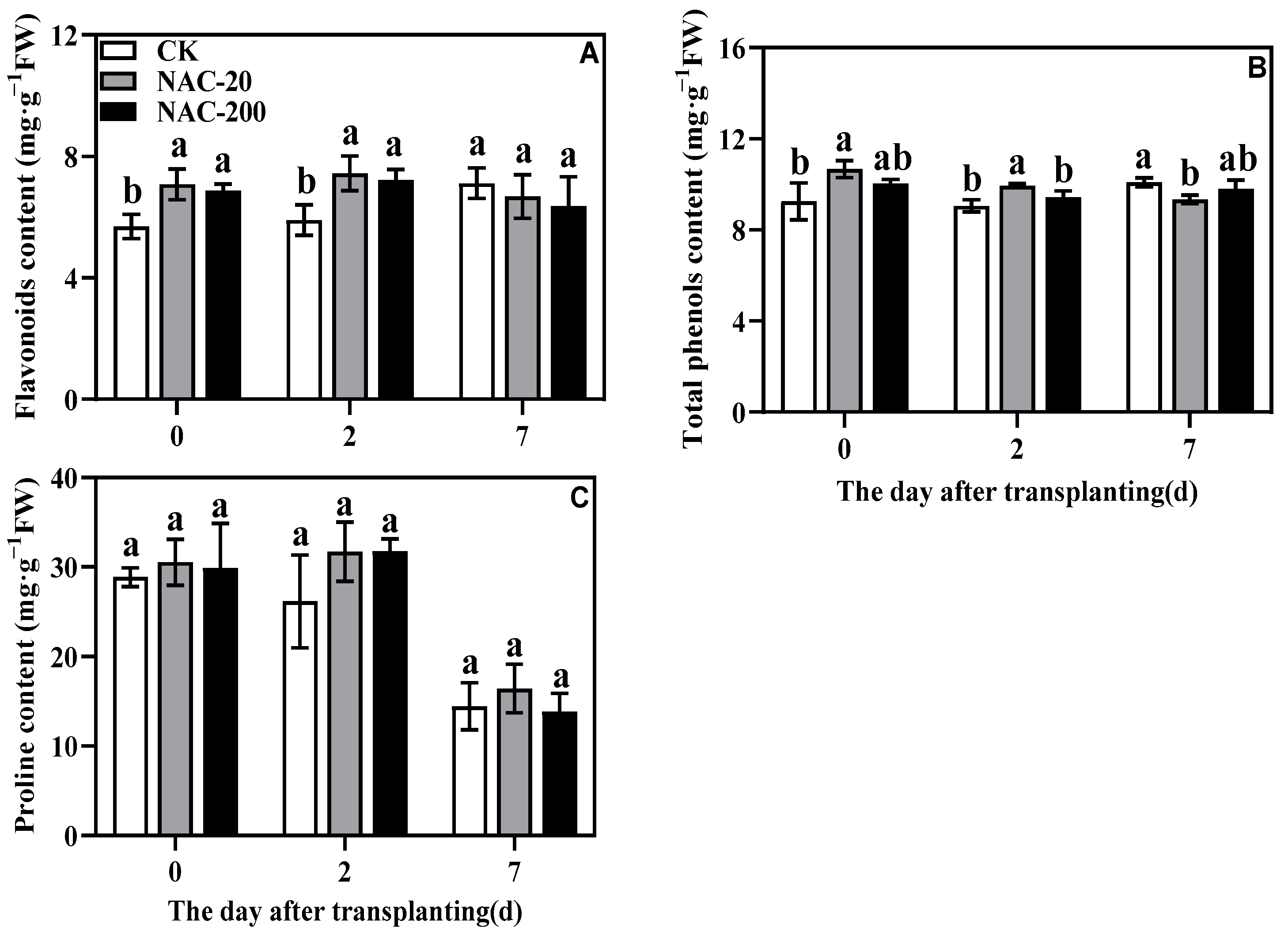
2.8. Non-Enzymatic Antioxidant Content
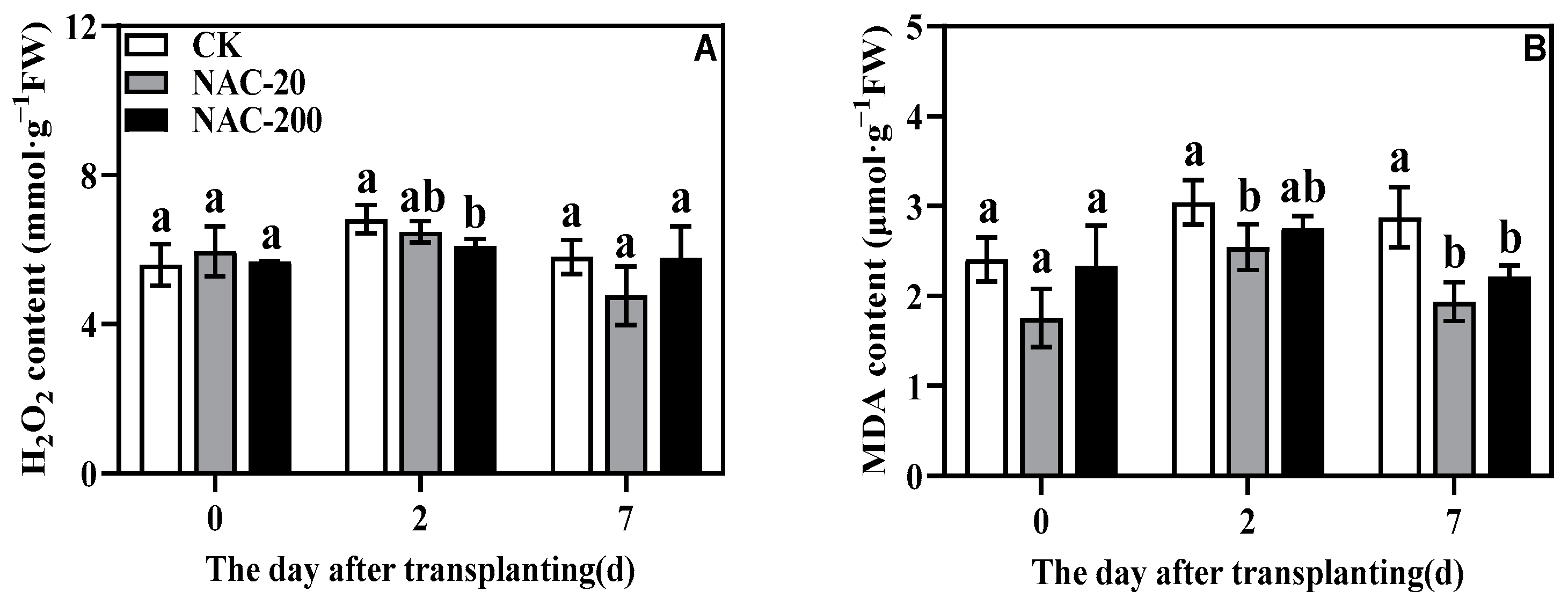
2.9. H2O2 and MDA Content
3. Discussion
3.1. The Concentration of 20 μM NAC Priming Is More Effective in Promoting Tillering and Yield of Machine-Transplanted Rice under Transplanting Injury
3.2. NAC Priming Improves the Photosynthetic Capacity and Promotes the Accumulation of Sucrose and Dry Matter of Machine-Transplanted Rice under Transplanting Injury
3.3. NAC Priming Improves the Antioxidant Capacity to Reduce Transplanting Injury of Machine-Transplanted Rice
3.4. The Mechanism of NAC Priming Mitigating the Transplanting Injury of Machine-Transplanted Rice
4. Materials and Methods
4.1. Experiment Site
4.2. Experimental Design
4.3. Parameter Measurements
4.4. Statistic Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Abid, M.; Khan, I.; Mahmood, F.; Ashraf, U.; Imran, M.; Anjum, S.A. Response of Hybrid Rice to Various Transplanting Dates and Nitrogen Application Rates. Philippine Agric. Sci. 2015, 98, 98–104. [Google Scholar]
- Kong, L.L.; Ashraf, U.; Cheng, S.R.; Rao, G.S.; Mo, Z.W.; Tian, H.; Pan, S.G.; Tang, X.R. Short-term water management at early filling stage improves early-season rice performance under high temperature stress in South China. Eur. J. Agron. 2017, 90, 117–126. [Google Scholar] [CrossRef]
- Chen, S.; Liu, S.W.; Zheng, X.; Yin, M.; Chu, G.; Xu, C.M.; Yan, J.X.; Chen, L.P.; Wang, D.Y.; Zhang, X.F. Effect of various crop rotations on rice yield and nitrogen use efficiency in paddy-upland systems in southeastern China. Crop. J. 2018, 6, 576–588. [Google Scholar] [CrossRef]
- Xing, Z.P.; Hu, Y.J.; Qian, H.J.; Cao, W.W.; Guo, B.W.; Wei, H.Y.; Xu, K.; Huo, Z.Y.; Zhou, G.S.; Dai, Q.G.; et al. Comparison of yield traits in rice among three mechanized planting methods in a rice-wheat rotation system. J. Integr. Agric. 2017, 16, 1451–1466. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Khan, A.L.; Waqas, M.; Shahzad, R.; Lee, I.J. Silicon-mediated Mitigation of Wounding Stress Acts by Up-regulating the Rice Antioxidant System. Cereal Res. Commun. 2016, 44, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Consales, F.; Schweizer, F.; Erb, M.; Gouhier-Darimont, C.; Bodenhausen, N.; Bruessow, F.; Sobhy, I.; Reymond, P. Insect oral secretions suppress wound-induced responses in Arabidopsis. J. Exp. Bot. 2012, 63, 727–737. [Google Scholar] [CrossRef] [Green Version]
- Cui, F.; Brosche, M.; Sipari, N.; Tang, S.; Overmyer, K. Regulation of ABA dependent wound induced spreading cell death by MYB108. New Phytol. 2013, 200, 634–640. [Google Scholar] [CrossRef]
- Savatin, D.V.; Gramegna, G.; Modesti, V.; Cervone, F. Wounding in the plant tissue: The defense of a dangerous passage. Front. Plant Sci. 2014, 5, 470. [Google Scholar] [CrossRef] [Green Version]
- Leon, J.; Rojo, E.; Sanchez-Serrano, J.J. Wound signalling in plants. J. Exp. Bot. 2001, 52, 1–9. [Google Scholar] [CrossRef]
- Lin, C.C.; Kao, C.H. Effect of NaCl stress on H2O2 metabolism in rice leaves. Plant Growth Regul. 2000, 30, 151–155. [Google Scholar] [CrossRef]
- Zhu, Z.J.; Wei, G.Q.; Li, J.; Qian, Q.Q.; Yu, J.Q. Silicon alleviates salt stress and increases antioxidant enzymes activity in leaves of salt-stressed cucumber (Cucumis sativus L.). Plant Sci. 2004, 167, 527–533. [Google Scholar] [CrossRef]
- Lee, A.; Cho, K.; Jang, S.; Rakwal, R.; Iwahashi, H.; Agrawal, G.K.; Shim, J.; Han, O. Inverse correlation between jasmonic acid and salicylic acid during early wound response in rice. Biochem. Biophys. Res. Commun. 2004, 318, 734–738. [Google Scholar] [CrossRef] [PubMed]
- Suttle, J.C.; Lulai, E.C.; Huckle, L.L.; Neubauer, J.D. Wounding of potato tubers induces increases in ABA biosynthesis and catabolism and alters expression of ABA metabolic genes. J. Plant Physiol. 2013, 170, 560–566. [Google Scholar] [CrossRef]
- Liu, Y.; Pan, Q.H.; Yang, H.R.; Liu, Y.Y.; Huang, W.D. Relationship between H2O2 and Jasmonic Acid in Pea Leaf Wounding Response. Russian J. Plant Physiol. 2008, 55, 765–775. [Google Scholar] [CrossRef]
- Castrillon-Arbelaez, P.A.; Martinez-Gallardo, N.; Arnaut, H.A.; Tiessen, A.; Delano-Frier, J.P. Metabolic and enzymatic changes associated with carbon mobilization, utilization and replenishment triggered in grain amaranth (Amaranthus cruentus) in response to partial defoliation by mechanical injury or insect herbivory. BMC Plant Biol. 2012, 12, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bari, R.; Jones, J.D. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Mulyati; Bell, R.; Huang, L.B. Root pruning and transplanting increase zinc requirements of canola (Brassica napus). Plant Soil 2009, 314, 11–24. [Google Scholar] [CrossRef]
- Savvides, A.; Ali, S.; Tester, M.; Fotopoulos, V. Chemical Priming of Plants Against Multiple Abiotic Stresses: Mission Possible? Trends Plant Sci. 2016, 21, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Anfossi, G.; Russo, I.; Massucco, P.; Mattiello, L.; Cavalot, F.; Trovati, M. N-acetyl-L-cysteine exerts direct anti-aggregating effect on human platelets. Eur. J. Clin. Investig. 2001, 31, 452–461. [Google Scholar] [CrossRef]
- Muranaka, L.S.; Giorgiano, T.E.; Takita, M.A.; Forim, M.R.; Silva, L.F.C.; Coletta-Filho, H.D.; Machado, M.A.; de Souza, A.A. N-Acetylcysteine in Agriculture, a Novel Use for an Old Molecule: Focus on Controlling the Plant-Pathogen Xylella fastidiosa. PLoS ONE 2013, 8, e72937. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Zhang, X.; He, X.; Ahmed, I.M.; Cao, F.; Zhang, G.; Wu, F. N-acetyl-cysteine alleviates Cd toxicity and reduces Cd uptake in the two barley genotypes differing in Cd tolerance. Plant Growth Regul. 2014, 74, 93–105. [Google Scholar] [CrossRef]
- Deng, X.P.; Xia, Y.; Hu, W.; Zhang, H.X.; Shen, Z.G. Cadmium-induced oxidative damage and protective effects of N-acetyl-L-cysteine against cadmium toxicity in Solanum nigrum L. J. Hazardous Mater. 2010, 180, 722–729. [Google Scholar] [CrossRef] [PubMed]
- He, Y.Y.; Hader, D.P. UV-B-induced formation of reactive oxygen species and oxidative damage of the cyanobacterium Anabaena sp.: Protective effects of ascorbic acid and N-acetyl-L-cysteine. J. Photochem. Photobiol. B 2002, 66, 115–124. [Google Scholar] [CrossRef]
- Malanga, G.; Kozak, R.G.; Puntarulo, S. N-acetylcysteine-dependent protection against UV-B damage in two photosynthetic organisms. Plant Sci. 1999, 141, 129–137. [Google Scholar] [CrossRef]
- Iriti, M.; Castorina, G.; Picchi, V.; Faoro, F.; Gomarasca, S. Acute exposure of the aquatic macrophyte Callitriche obtusangula to the herbicide oxadiazon: The protective role of N-acetylcysteine. Chemosphere 2009, 74, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.-J.; Wu, P.; Zhang, H.-C.; Dai, Q.-G.; Huo, Z.-Y.; Xu, K.; Gao, H.; Wei, H.-Y.; Guo, B.-W.; Cui, P.-Y. Comparison of agronomic performance between inter-sub-specific hybrid and inbred japonica rice under different mechanical transplanting methods. J. Integr. Agric. 2018, 17, 806–816. [Google Scholar] [CrossRef] [Green Version]
- Cui, Z.; Dou, Z.; Chen, X.; Ju, X.; Zhang, F. Managing Agricultural Nutrients for Food Security in China: Past, Present, and Future. Agron. J. 2014, 106, 191–198. [Google Scholar] [CrossRef]
- Aslam, M.M.; Rashid, M.A.R.; Siddiqui, M.A.; Khan, M.T.; Farhat, F.; Yasmeen, S.; Khan, I.A.; Raja, S.; Rasool, F.; Ali Sial, M.; et al. Recent Insights into Signaling Responses to Cope Drought Stress in Rice. Rice Sci. 2022, 29, 105–117. [Google Scholar] [CrossRef]
- Colak, N.; Torun, H.; Gruz, J.; Strnad, M.; Ayaz, F.A. Exogenous N-Acetylcysteine alleviates heavy metal stress by promoting phenolic acids to support antioxidant defence systems in wheat roots. Ecotoxicol. Environ. Saf. 2019, 181, 49–59. [Google Scholar] [CrossRef]
- Zhang, C.M. The key techniques of “standardizing seedlings, precise transplanting, steady growing, earlier drainage, optimizing middle-stage, strengthening later-stage” model for precise quantitative high-yielding cultivation of mechanical transplanting rice. China Rice 2010, 16, 6. [Google Scholar]
- Takahashi, N.; Sunohara, Y.; Fujiwara, M.; Matsumoto, H. Improved tolerance to transplanting injury and chilling stress in rice seedlings treated with orysastrobin. Plant Physiol. Biochem. 2017, 113, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Reid, J.F.; Zhang, Q.; Noguchi, N.; Dickson, M. Agricultural automatic guidance research in North America. Comput. Electron. Agric. 2000, 25, 155–167. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N.; et al. Phytohormones and plant responses to salinity stress: A review. Plant Growth Regul. 2015, 75, 391–404. [Google Scholar] [CrossRef]
- Muthuchelian, K.; Murugan, C.; Nedunchezhian, N.; Kulandaivelu, G. Photosynthesis and growth of Erythrina variegata as affected by water stress and triacontanol. Photosynthetica 1997, 33, 241–248. [Google Scholar] [CrossRef]
- Weerasinghe, O.R.; Costa, W.D.; Perera, A. Evaluation of different genotypes of tomato under well watered and water stressed conditions on the basis of yield and some selected physiological parameters. Trop. Agric. Res. 2003, 144–156. [Google Scholar]
- Munirah, N.; Mat, N.; Jahan, M.S. N-acetylcysteine and Zn regulate corn yield. Scienceasia 2015, 41, 246–250. [Google Scholar] [CrossRef] [Green Version]
- Nozulaidi, M.; Jahan, M.S.; Khairi, M.; Khandaker, M.M.; Nashriyah, M.; Khanif, Y.M. N-acetylcysteine increased rice yield. Turk J. Agric. For. 2015, 39, 204–211. [Google Scholar] [CrossRef]
- Colak, N.; Tarkowski, P.; Ayaz, F.A. Effect of N-acetyl-L-cysteine (NAC) on soluble sugar and polyamine content in wheat seedlings exposed to heavy metal stress (Cd, Hg and Pb). Bot. Serb. 2020, 44, 191–201. [Google Scholar] [CrossRef]
- Bell, R.W.; Lu, Z.G.; Li, J.; Hu, D.J.; Xie, Z.C. Response of transplanted oilseed rape to zinc placement and root pruning. J. Plant Nutr. 2004, 27, 427–439. [Google Scholar] [CrossRef]
- Dumanovic, J.; Nepovimova, E.; Natic, M.; Kuca, K.; Jacevic, V. The Significance of Reactive Oxygen Species and Antioxidant Defense System in Plants: A Concise Overview. Front. Plant Sci. 2021, 11, 2106. [Google Scholar] [CrossRef]
- Flowers, T.J. Sun, Salt, and Halophytes—A Citation-Classic Commentary on the Mechanism of Salt Tolerance in Halophytes by Flowers, T.J., Troke, P.F., Yeo, A.R. Curr. Content Agric. Biol. Environ. 1993, 8. [Google Scholar]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Ann. Rev. Plant Phys. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; He, Z.; Li, X.; Ding, Y.; Li, G.; Liu, Z.; Tang, S.; Wang, S. Quality and Field Growth Characteristics of Hydroponically Grown Long-Mat Seedlings. Agron. J. 2016, 108, 1581–1591. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24. [Google Scholar] [CrossRef] [Green Version]
- Hendrix, D.L. Rapid Extraction and Analysis of Nonstructural Carbohydrates in Plant Tissues. Crop Sci. 1993, 33, 1306–1311. [Google Scholar] [CrossRef]
- Chen, Y.E.; Cui, J.M.; Su, Y.Q.; Zhang, C.M.; Ma, J.; Zhang, Z.W.; Yuan, M.; Liu, W.J.; Zhang, H.Y.; Yuan, S. Comparison of phosphorylation and assembly of photosystem complexes and redox homeostasis in two wheat cultivars with different drought resistance. Sci. Rep. 2017, 7, 12718. [Google Scholar] [CrossRef] [Green Version]
- Jia, Z.; Tang, M.C.; Wu, J.M. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Djeridane, A.; Yousfi, M.; Nadjemi, B.; Boutassouna, D.; Stocker, P.; Vidal, N. Antioxidant activity of some algerian medicinal plants extracts containing phenolic compounds. Food Chem. 2006, 97, 654–660. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, J. Effect of abscisic acid on active oxygen species, antioxidative defence system and oxidative damage in leaves of maize seedlings. Plant Cell Physiol. 2001, 42, 1265–1273. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Panicles Pot−1 | Spikelets Panicle−1 | Grain Filling (%) | Grain Weight (mg) | Grain Yield (g·pot−1) |
|---|---|---|---|---|---|
| CK | 28.7 ± 1.2 b | 153.3 ± 5.5 a | 90.2 ± 1.5 b | 28.5 ± 0.2 a | 112.6 ± 2.2 b |
| NAC-20 | 31.4 ± 1.0 a | 149.1 ± 6.6 a | 92.9 ± 0.7 a | 28.8 ± 0.4 a | 125.2 ± 2.7 a |
| NAC-200 | 30.9 ± 0.7 ab | 148.4 ± 5.2 a | 92.6 ± 0.9 ab | 28.7 ± 0.5 a | 121.7 ± 4.6 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, W.; Zhong, Q.; He, B.; Wu, B.; Mohi Ud Din, A.; Han, J.; Ding, Y.; Liu, Z.; Li, W.; Jiang, Y.; et al. N-Acetylcysteine Priming Alleviates the Transplanting Injury of Machine-Transplanted Rice by Comprehensively Promoting Antioxidant and Photosynthetic Systems. Plants 2022, 11, 1311. https://doi.org/10.3390/plants11101311
He W, Zhong Q, He B, Wu B, Mohi Ud Din A, Han J, Ding Y, Liu Z, Li W, Jiang Y, et al. N-Acetylcysteine Priming Alleviates the Transplanting Injury of Machine-Transplanted Rice by Comprehensively Promoting Antioxidant and Photosynthetic Systems. Plants. 2022; 11(10):1311. https://doi.org/10.3390/plants11101311
Chicago/Turabian StyleHe, Wenjun, Qiuyi Zhong, Bin He, Boyang Wu, Atta Mohi Ud Din, Jielyv Han, Yanfeng Ding, Zhenghui Liu, Weiwei Li, Yu Jiang, and et al. 2022. "N-Acetylcysteine Priming Alleviates the Transplanting Injury of Machine-Transplanted Rice by Comprehensively Promoting Antioxidant and Photosynthetic Systems" Plants 11, no. 10: 1311. https://doi.org/10.3390/plants11101311
APA StyleHe, W., Zhong, Q., He, B., Wu, B., Mohi Ud Din, A., Han, J., Ding, Y., Liu, Z., Li, W., Jiang, Y., & Li, G. (2022). N-Acetylcysteine Priming Alleviates the Transplanting Injury of Machine-Transplanted Rice by Comprehensively Promoting Antioxidant and Photosynthetic Systems. Plants, 11(10), 1311. https://doi.org/10.3390/plants11101311