
Plant-Derived Smoke Solution Alleviates Cellular Oxidative Stress Caused by Arsenic and Mercury by Modulating the Cellular Antioxidative Defense System in Wheat

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Solutions
2.2. Seed Source and Sterilization
2.3. Measurement of H2O2 and Lipid Peroxidation
2.4. Quantification of Antioxidant Enzymes
2.4.1. Measurement of Superoxide Dismutase (SOD) Activity
2.4.2. Quantification of Peroxidase (POD) and Ascorbate Peroxidase (APX) Activity
2.4.3. Statistical Evaluation
3. Result and Discussions
3.1. PDS Promotes Wheat Seed Germination under As and Hg Stress
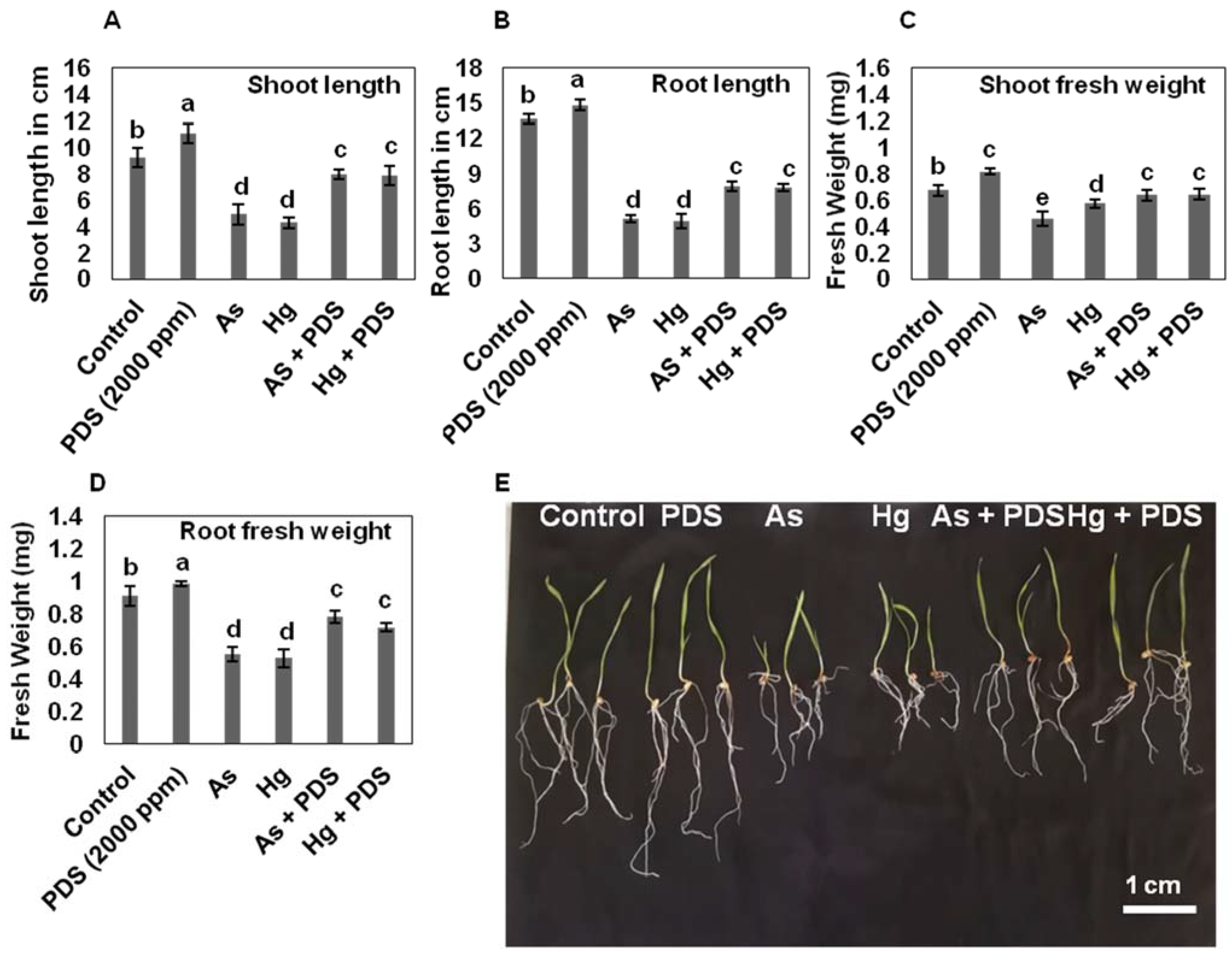
3.2. PDS Enhances Wheat Seedling Growth under As or Hg Stress
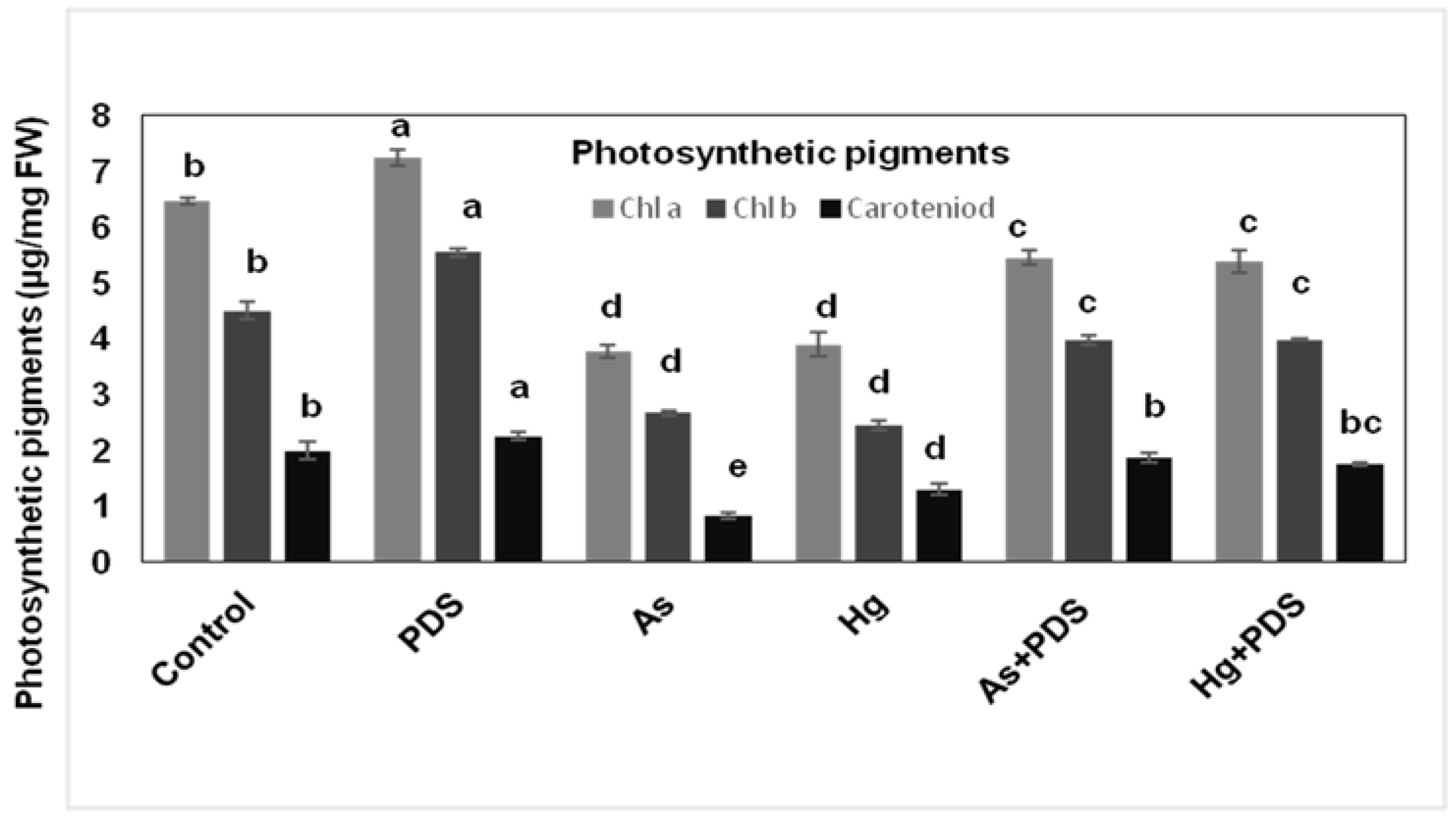
3.3. Combined Effect of As, Hg Stress and PDS (2000 ppm) Solution on Photosynthetic Pigments of Wheat
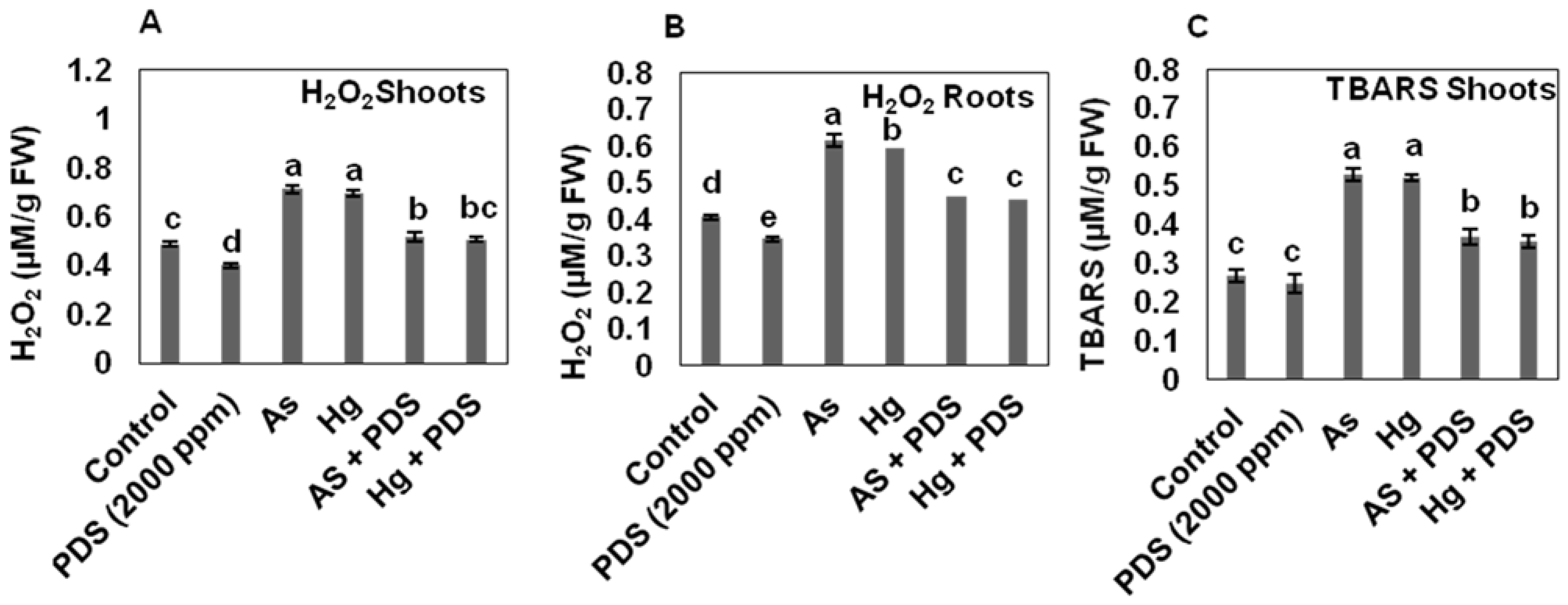
3.4. Effect of PDS on H2O2 and TBRS under As and Hg Stress
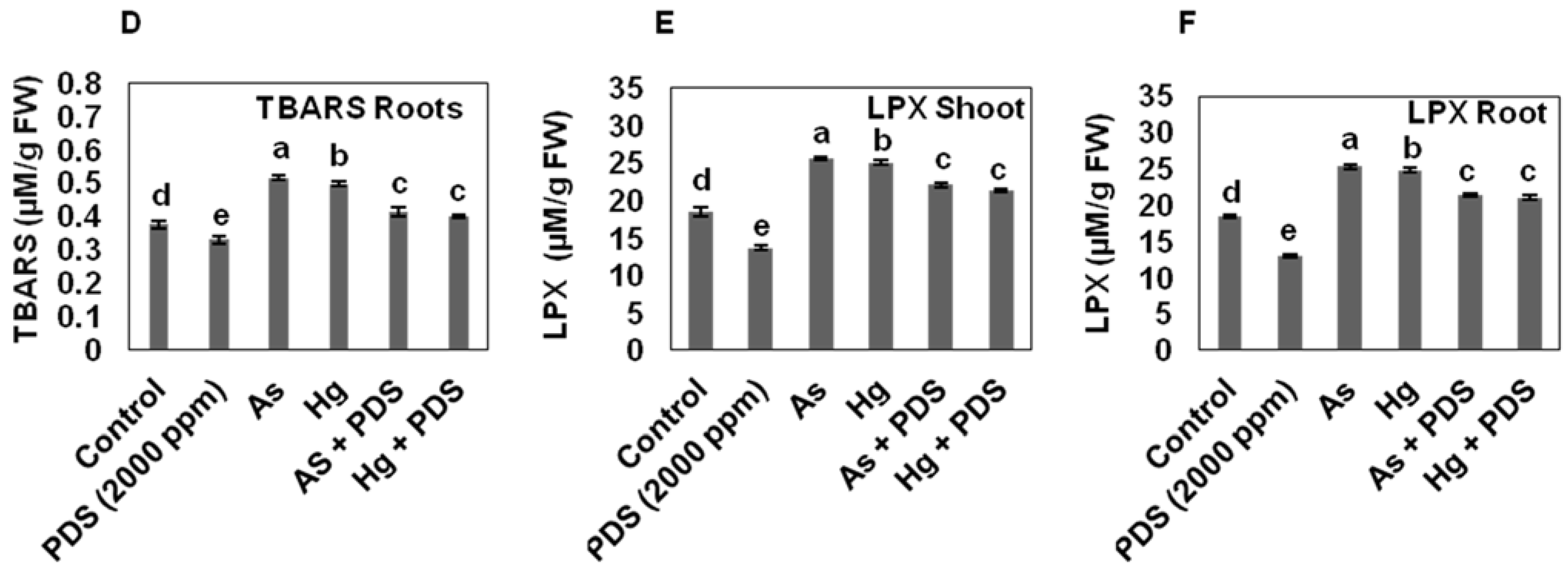
3.5. PDS Reduces Levels of Lipid Peroxidation Production under As and Hg Stress
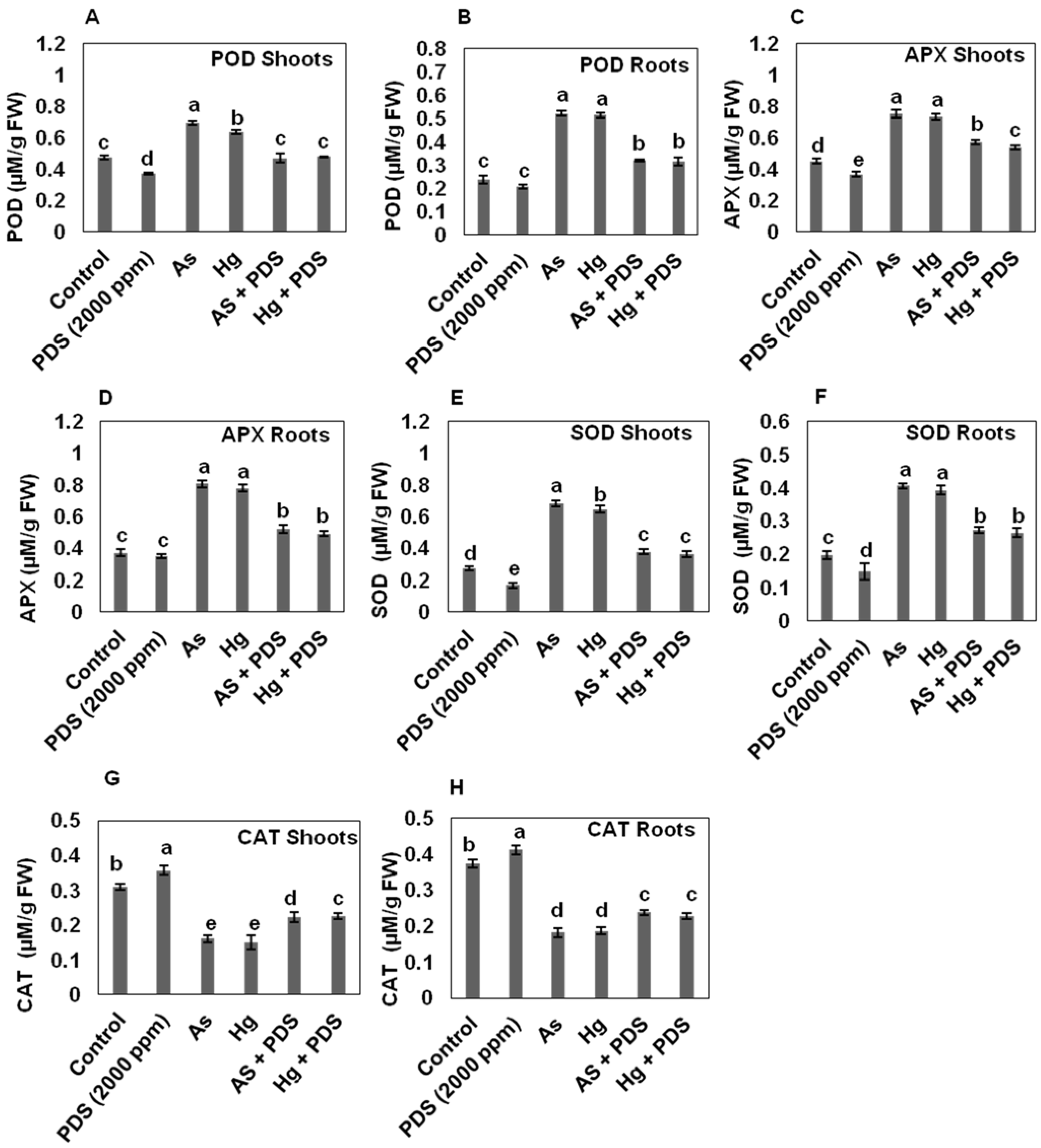
3.6. PDS Modulates the Activity of Antioxidant Enzymes under As and Hg Stress
3.6.1. PDS Lowers the Activity of Peroxidase (POD) under As and Hg Stress
3.6.2. PDS Decreases the Activity of Ascorbate Peroxidase (APX) under As and Hg Stress
3.6.3. PDS Decreases the Activity of Superoxide Dismutase (SOD) under As and Hg Stress
3.6.4. PDS Induces CAT Activities under As and Hg Stress
4. Conclusions and Future Recommendations
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ullah, R.; Alqahtani, A.S. GC-MS Analysis, Heavy Metals, Biological, and Toxicological Evaluation of Reseda muricata and Marrubium vulgare Methanol Extracts. Evid.-Based Complement. Altern. Med. 2022, 2022, 2284328. [Google Scholar] [CrossRef] [PubMed]
- Alqahtani, A.S.; Ullah, R.; Shahat, A.A. Bioactive Constituents and Toxicological Evaluation of Selected Antidiabetic Medicinal Plants of Saudi Arabia. Evid.-Based Complement. Altern. Med. 2022, 2022, 7123521. [Google Scholar] [CrossRef] [PubMed]
- Mandal, B.K.; Suzuki, K.T. Arsenic round the world: A review. Talanta 2002, 58, 201–235. [Google Scholar] [CrossRef]
- Loska, K.; Wiechula, D.; Barska, B.; Cebula, E.; Chojnecka, A. Assessment of arsenic enrichment of cultivated soils in Southern Poland. Pol. J. Environ. Stud. 2003, 12, 187–192. [Google Scholar]
- Arain, M.B.; Kazi, T.G.; Baig, J.A.; Jamali, M.K.; Afridi, H.I.; Shah, A.Q.; Jalbani, N.; Sarfraz, R.A. Determination of arsenic levels in lake water, sediment, and foodstuff from selected area of Sindh, Pakistan: Estimation of daily dietary intake. Food Chem. Toxicol. 2009, 47, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-X.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Liu, X.-L.; Zhang, S.-Z. Intraspecific differences in effects of co-contamination of cadmium and arsenate on early seedling growth and metal uptake by wheat. J. Environ. Sci. 2007, 19, 1221–1227. [Google Scholar] [CrossRef]
- Abedin, M.D.; Meharg, A.A. Relative toxicity of arsenite and arsenate on germination and early seedling growth of rice (Oryza sativa L.). Plant Soil 2002, 243, 57–66. [Google Scholar] [CrossRef]
- Yunsheng, Z.; Wei, S.; Qianli, C.; Lin, C. Synthesis and heavy metal immobilization behaviors of slag based geopolymer. J. Hazard. Mater. 2007, 143, 206–213. [Google Scholar] [CrossRef]
- Cao, X.; Ma, L.Q. Effects of compost and phosphate on plant arsenic accumulation from soils near pressure-treated wood. Environ. Pollut. 2004, 132, 435–442. [Google Scholar] [CrossRef]
- Patra, M.; Sharma, A. Mercury toxicity in plants. Bot. Rev. 2000, 66, 379–422. [Google Scholar] [CrossRef]
- Laurier, P.; Pilote, M.; Xu, X.; Zhang, H.; Beauvais, C. Atmospheric mercury speciation and deposition in the Bay St. Francois wetlands. J. Geophys. Res. Atmos. 2004, 109, D11. [Google Scholar]
- Hassett, D.J.; Eylands, K.E. Mercury capture on coal combustion fly ash. Fuel 1999, 78, 243–248. [Google Scholar] [CrossRef]
- Lodenius, M.; Tulisalo, E.; Soltanpour-Gargari, A. Exchange of mercury between atmosphere and vegetation under contaminated conditions. Sci. Total Environ. 2003, 304, 169–174. [Google Scholar] [CrossRef]
- Rea, A.W.; Lindberg, S.E.; Keeler, G.J. Dry deposition and foliar leaching of mercury and selected trace elements in deciduous forest throughfall. Atmos. Environ. 2001, 35, 3453–3462. [Google Scholar] [CrossRef]
- Schwesig, D.; Krebs, O. The role of ground vegetation in the uptake of mercury and methylmercury in a forest ecosystem. Plant Soil 2003, 253, 445–455. [Google Scholar] [CrossRef]
- Sharma, P.; Allison, J.P. The future of immune checkpoint therapy. Science 2015, 348, 56–61. [Google Scholar] [CrossRef]
- Sharma, P.; Rama, S.D. Lead toxicity in plants. Braz. J. Plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef] [Green Version]
- Patra, M.; Bhowmik, N.; Bandopadhyay, B.; Sharma, A. Comparison of mercury, lead and arsenic with respect to genotoxic effects on plant systems and the development of genetic tolerance. Environ. Exp. Bot. 2004, 52, 199–223. [Google Scholar] [CrossRef]
- Przedpelska-Wasowicz, E.M.; Malgorzata, W. Gating of aquaporins by heavy metals in Allium cepa L. epidermal cells. Protoplasma 2011, 248, 663–671. [Google Scholar] [CrossRef] [Green Version]
- Sparks, D.L. Toxic Metals in the Environment: The Role of Surfaces. Elements 2005, 1, 193–197. [Google Scholar] [CrossRef]
- Ortega-Villasante, C.; Rellán-Álvarez, R.; Del Campo, F.F.; Carpena-Ruiz, R.O.; Hernández, L.E. Cellular damage induced by cadmium and mercury in Medicago sativa. J. Exp. Bot. 2005, 56, 2239–2251. [Google Scholar] [CrossRef] [PubMed]
- Israr, M.; Sahi, S.; Datta, R.; Sarkar, D. Bioaccumulation and physiological effects of mercury in Sesbania drummondii. Chemosphere 2006, 65, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M. Vicia faba bioassay for environmental toxicity monitoring: A review. Chemosphere 2015, 144, 785–802. [Google Scholar] [CrossRef]
- Iqbal, M.; Asif, S.; Ilyas, N.; Raja, N.I.; Hussain, M.; Shabir, S.; Faz, M.N.A.; Rauf, A. Effect of Plant Derived Smoke on Germination and Post Germination Expression of Wheat (Triticum aestivum L.). Am. J. Plant Sci. 2016, 7, 806–813. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, M.G.; Light, M.E.; Van Staden, J. Plant-derived smoke: Old technology with possibilities for economic applications in agriculture and horticulture. S. Afr. J. Bot. 2011, 77, 972–979. [Google Scholar] [CrossRef] [Green Version]
- Demir, I.; Ozuaydın, I.; Yasar, F.; Van Staden, J. Effect of smoke-derived butenolide priming treatment on pepper and salvia seeds in relation to transplant quality and catalase activity. S. Afr. J. Bot. 2012, 78, 83–87. [Google Scholar] [CrossRef] [Green Version]
- Abraha, B.; Yohannes, G. The role of seed priming in improving seedling growth of maize (Zea mays L.) under salt stress at field conditions. Agric. Sci. 2013, 4, 40842. [Google Scholar]
- Waheed, M.A.; Jamil, M.; Khan, M.D.; Shakir, S.K.; Rehman, S.U. Effect of plant-derived smoke solutions on physiological and biochemical attributes of maize (Zea mays L.) under salt stress. Pak. J. Bot 2016, 48, 1763–1774. [Google Scholar]
- Khatoon, A.; Rehman, S.U.; Aslam, M.M.; Jamil, M.; Komatsu, S. Plant-Derived Smoke Affects Biochemical Mechanism on Plant Growth and Seed Germination. Int. J. Mol. Sci. 2020, 21, 7760. [Google Scholar] [CrossRef]
- Faheem Afzal, S.; Wei, X.; Wang, Q.; Liu, W.; Wang, D.; Yao, Y.; Hu, H.; Chen, X.; Huang, S.; Hou, J.; et al. Karrikin improves osmotic and salt stress tolerance via the regulation of the redox homeostasis in the oil plant Sapium sebiferum. Front. Plant Sci. 2020, 11, 216. [Google Scholar]
- Ghebrehiwot, H.M.; Kulkarni, M.G.; Kirkman, K.P.; van Staden, J. Smoke-Water and a Smoke-Isolated Butenolide Improve Germination and Seedling Vigour of Eragrostis tef (Zucc.) Trotter under High Temperature and Low Osmotic Potential. J. Agron. Crop Sci. 2008, 194, 270–277. [Google Scholar] [CrossRef]
- Nelson, D.C.; Scaffidi, A.; Dun, E.A.; Waters, M.T.; Flematti, G.R.; Dixon, K.W.; Beveridge, C.A.; Ghisalberti, E.L.; Smith, S.M. F-box protein MAX2 has dual roles in karrikin and strigolactone signaling in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2011, 108, 8897–8902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunmonu, T.; Kulkarni, M.; Van Staden, J. Smoke-water, karrikinolide and gibberellic acid stimulate growth in bean and maize seedlings by efficient starch mobilization and suppression of oxidative stress. S. Afr. J. Bot. 2016, 102, 4–11. [Google Scholar] [CrossRef]
- Shabir, S.; Noshin, I.; Saira, A.; Muhammad, I.; Sidra, K.; Zeshan, A. Deciphering the Role of Plant-Derived Smoke Solution in Ameliorating Saline Stress and Improving Physiological, Biochemical, and Growth Responses of Wheat. J. Plant Growth Regul. 2021, 40, 1–18. [Google Scholar] [CrossRef]
- Sun, R.; Yang, J.; Xia, P.; Wu, S.; Lin, T.; Yi, Y. Contamination features and ecological risks of heavy metals in the farmland along shoreline of Caohai plateau wetland, China. Chemosphere 2020, 254, 126828. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, P.; Benavente, E.; Manzano-Agugliaro, F.; Gimenez, E. Worldwide Research Trends on Wheat and Barley: A Bibliometric Comparative Analysis. Agronomy 2019, 9, 352. [Google Scholar] [CrossRef] [Green Version]
- Smol, J.P. Climate Change: A planet in flux. Nature 2012, 483, S12–S15. [Google Scholar] [CrossRef]
- Sun, Z.W.; Ren, L.K.; Fan, J.W.; Li, Q.; Wang, K.J.; Guo, M.M.; Wang, L.; Li, J.; Zhang, G.X.; Yang, Z.Y.; et al. Salt response of photosynthetic electron transport system in wheat cultivars with contrasting tolerance. Plant Soil Environ. 2016, 62, 515–521. [Google Scholar]
- Hendry, G.A.F. Evolutionary origins and natural functions of fructans–a climatological, biogeographic and mechanistic appraisal. New Phytol. 1993, 123, 3–14. [Google Scholar] [CrossRef]
- Bailly, C. Active oxygen species and antioxidants in seed biology. Seed Sci. Res. 2004, 14, 93–107. [Google Scholar] [CrossRef]
- Tieu, A.; Plummer, J.A.; Dixon, K.A.; Sivasithamparam, K.; Sieler, I.M. Germination of Four Species of Native Western Australian Plants using Plant-derived Smoke. Aust. J. Bot. 1999, 47, 207–219. [Google Scholar] [CrossRef]
- Hayat, N.; Afroz, N.; Rehman, S.; Bukhari, S.H.; Iqbal, K.; Khatoon, A.; Taimur, N.; Sakhi, S.; Ahmad, N.; Ullah, R.; et al. Plant-Derived Smoke Ameliorates Salt Stress in Wheat by Enhancing Expressions of Stress-Responsive Genes and Antioxidant Enzymatic Activity. Agronomy 2021, 12, 28. [Google Scholar] [CrossRef]
- Willekens, H.; Chamnongpol, S.; Davey, M.; Schraudner, M.; Langebartels, C.; Van Montagu, M.; Inzé, D.; Van Camp, W. Catalase is a sink for H2O2 and is indispensable for stress defence in C3 plants. EMBO J. 1997, 16, 4806–4816. [Google Scholar] [CrossRef] [Green Version]
- Mark, H.D.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Jun, N.; Wang, Q.; Afzal Shah, F.; Liu, W.; Wang, D.; Huang, S.; Fu, S.; Wu, L. Exogenous melatonin confers cadmium tolerance by counterbalancing the hydrogen peroxide homeostasis in wheat seedlings. Molecules 2018, 23, 799. [Google Scholar]
- Mukherjee, S.; Basak, B.; Bhunia, B.; Dey, A.; Mondal, B. Potential use of polyphenol oxidases (PPO) in the bioremediation of phenolic contaminants containing industrial wastewater. Rev. Environ. Sci. Bio/Technol. 2012, 12, 61–73. [Google Scholar] [CrossRef]
- Jamal, S.N.; Iqbal, Z.; Athar, M. Evaluation of two wheat varieties for phytotoxic effect of mercury on seed germination and seedling growth. Agric. Conspec. Sci. 2006, 71, 41–44. [Google Scholar]
- Talukdar, D. Effect of arsenic-induced toxicity on morphological traits of Trigonella foenum-graecum L. and Lathyrus sativus L. during germination and early seedling growth. Curr. Res. J. Biol. Sci. 2011, 3, 116–123. [Google Scholar]
- Shankar, D.; Shenoi, S.S.C.; Nayak, R.K.; Vinayachandran, P.N.; Nampoothiri, G.; Almeida, A.M.; Michael, G.S.; Kumar, M.R.R.; Sundar, D.; Sreejith, O.P. Hydrography of the eastern Arabian Sea during summer monsoon 2002. J. Earth Syst. Sci. 2005, 114, 459–474. [Google Scholar] [CrossRef]
- Munzuroglu, O.; Geckil, H. Effects of metals on seed germination, root elongation, and coleoptile and hypocotyl growth in Triticum aestivum and Cucumis sativus. Arch. Environ. Contam. Toxicol. 2002, 43, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Espen, L.; Pirovano, L.; Cocucci, S.M. Effects of Ni2+ during the early phases of radish (Raphanus sativus) seed germination. Environ. Exp. Bot. 1997, 38, 187–1977. [Google Scholar] [CrossRef]
- Jain, N.; Kulkarni, M.G.; Van Staden, J. A butenolide, isolated from smoke, can overcome the detrimental effects of extreme temperatures during tomato seed germination. Plant Growth Regul. 2006, 49, 263–267. [Google Scholar] [CrossRef]
- Malook, I.; Atlas, A.; Rehman, S.U.; Wang, W.; Jamil, M. Smoke alleviates adverse effects induced by stress on rice. Toxicol. Environ. Chem. 2014, 96, 755–767. [Google Scholar] [CrossRef]
- Ling, T.; Fangke, Y.; Jun, R. Effect of mercury to seed germination, coleoptile growth and root elongation of four vegetables. Res. J. Phytochem. 2010, 4, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Cenkci, S.; Ciğerci, İ.H.; Yıldız, M.; Özay, C.; Bozdağ, A.; Terzi, H. Lead contamination reduces chlorophyll biosynthesis and genomic template stability in Brassica rapa L. Environ. Exp. Bot. 2010, 67, 467–473. [Google Scholar] [CrossRef]
- Van Staden, J.; Sparg, S.G.; Kulkarni, M.G.; Light, M.E. Post-germination effects of the smoke-derived compound 3-methyl-2H-furo[2,3-c]pyran-2-one, and its potential as a preconditioning agent. Field Crops Res. 2006, 98, 98–105. [Google Scholar] [CrossRef]
- Blank, R.R.; Young, J.A. Heated Substrate and Smoke: Influence on Seed Emergence and Plant Growth. Rangel. Ecol. Manag. 1998, 51, 577. [Google Scholar] [CrossRef]
- Sparg, S.; Kulkarni, M.; Light, M.; Vanstaden, J. Improving seedling vigour of indigenous medicinal plants with smoke. Bioresour. Technol. 2005, 96, 1323–1330. [Google Scholar] [CrossRef]
- Light, M.; van Staden, J.; Bornman, C. The potential of smoke in seed technology. S. Afr. J. Bot. 2004, 70, 97–101. [Google Scholar] [CrossRef] [Green Version]
- Brown, N.; van Staden, J.; Daws, M.; Johnson, T. Patterns in the seed germination response to smoke in plants from the Cape Floristic Region, South Africa. S. Afr. J. Bot. 2003, 69, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Sparg, S.G.; Kulkarni, M.G.; Van Staden, J. Aerosol Smoke and Smoke-Water Stimulation of Seedling Vigor of a Commercial Maize Cultivar. Crop Sci. 2006, 46, 1336–1340. [Google Scholar] [CrossRef]
- Jamil, M.; Kanwal, M.; Aslam, M.M.; Khan, S.U.; Malook, I.; Tu, J.; Rehman, S.U. Effect of plant-derived smoke priming on physiological and biochemical characteristics of rice under salt stress condition. Aust. J. Crop Sci. 2014, 8, 159–170. [Google Scholar]
- McNear, D.H.; Afton, S.E.; Caruso, J.A. Exploring the structural basis for selenium/mercury antagonism in Allium fistulosum. Metallomics 2012, 4, 267–276. [Google Scholar] [CrossRef]
- Farooq, M.A.; Hong, Z.; Islam, F.; Noor, Y.; Hannan, F.; Zhang, Y.; Ayyaz, A.; Mwamba, T.M.; Zhou, W.; Song, W. Comprehensive proteomic analysis of arsenic induced toxicity reveals the mechanism of multilevel coordination of efficient defense and energy metabolism in two Brassica napus cultivars. Ecotoxicol. Environ. Saf. 2020, 208, 111744. [Google Scholar] [CrossRef]
- Michael, R.; Novak, O.; Strnad, M.; Schmülling, T. Arabidopsis cytokinin receptor mutants reveal functions in shoot growth, leaf senescence, seed size, germination, root development, and cytokinin metabolism. Plant Cell 2006, 18, 40–54. [Google Scholar]
- Muhammad, F.; Muhammad, N.; Niazi, K.U.K.; Hu, Y.; Wang, Y.; Wu, Y.; Sun, H.; Liu, R.; Dong, W.; Zhang, W.; et al. CO-releasing materials: An emphasis on therapeutic implications, as release and subsequent cytotoxicity are the part of therapy. Materials 2019, 12, 1643. [Google Scholar]
- Zhang, X.F.; Dong, X.L.; Huang, H.; Lv, B.; Lei, J.P.; Choi, C.J. Microstructure and microwave absorption properties of carbon-coated iron nanocapsules. J. Phys. D: Appl. Phys. 2007, 40, 5383–5387. [Google Scholar] [CrossRef]
- Stoeva, N.; Berova, M.; Zlatev, Z. Physiological Response of Maize to Arsenic Contamination. Biol. Plant. 2003, 46, 449–452. [Google Scholar] [CrossRef]
- Geetika, S.; Mir, M.A.; Abd-Allah, E.F.; Ahmad, P.; Gucel, S. Jasmonic acid modulates the physio-biochemical attributes, antioxidant enzyme activity, and gene expression in Glycine max under nickel toxicity. Front. Plant Sci. 2016, 7, 591. [Google Scholar]
- Manna, I.; Bandyopadhyay, M. Engineered Nickel Oxide Nanoparticle Causes Substantial Physicochemical Perturbation in Plants. Front. Chem. 2017, 5, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kevers, C.; Franck, T.; Strasser, R.J.; Dommes, J.; Gaspar, T. Hyperhydricity of Micropropagated Shoots: A Typically Stress-induced Change of Physiological State. Plant Cell Tissue Organ Cult. (PCTOC) 2004, 77, 181–191. [Google Scholar] [CrossRef]
- Zhou, X.; Wheeler, M.M.; Oi, F.M.; Scharf, M.E. RNA interference in the termite Reticulitermes flavipes through ingestion of double-stranded RNA. Insect Biochem. Mol. Biol. 2008, 38, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Hartley-Whitaker, J.; Woods, C.; Meharg, A.A. Is differential phytochelatin production related to decreased arsenate influx in arsenate tolerant Holcus lanatus? New Phytol. 2002, 155, 219–225. [Google Scholar] [CrossRef]
- Gajewska, E.; Skłodowska, M. Differential biochemical responses of wheat shoots and roots to nickel stress: Antioxidative reactions and proline accumulation. Plant Growth Regul. 2007, 54, 179–188. [Google Scholar] [CrossRef]
- Nandita, S.; Ma, L.Q.; Srivastava, M.; Rathinasabapathi, B. Metabolic adaptations to arsenic-induced oxidative stress in Pteris vittata L. and Pteris ensiformis L. Plant Sci. 2006, 170, 274–282. [Google Scholar]
- Homaee, B.M.; Ehsanpour, A.A. Silver nanoparticles and silver ions: Oxidative stress responses and toxicity in potato (Solanum tuberosum L.) grown in vitro. Hortic. Environ. Biotechnol. 2016, 57, 544–553. [Google Scholar] [CrossRef]
- Manju, S.; Kumar, S.; Chakrabarty, D.; Trivedi, P.K.; Mallick, S.; Misra, P.; Shukla, D.; Mishra, S.; Srivastava, S.; Tripathi, R.D.; et al. Effect of arsenic on growth, oxidative stress, and antioxidant system in rice seedlings. Ecotoxicol. Environ. Saf. 2009, 72, 1102–1110. [Google Scholar]
- Elbaz, A.; Wei, Y.Y.; Meng, Q.; Zheng, Q.; Yang, Z.M. Mercury-induced oxidative stress and impact on antioxidant enzymes in Chlamydomonas reinhardtii. Ecotoxicology 2010, 19, 1285–1293. [Google Scholar] [CrossRef]
- Verma, S.; Dubey, R. Lead toxicity induces lipid peroxidation and alters the activities of antioxidant enzymes in growing rice plants. Plant Sci. 2003, 164, 645–655. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Klaus, A.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar]
- Hsu, Y.T.; Kao, C.H. Cadmium toxicity is reduced by nitric oxide in rice leaves. Plant Growth Regul. 2004, 42, 227–238. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, M.; Nawaz, S.; Iqbal, K.; Rehman, S.; Ullah, R.; Nawaz, G.; Almeer, R.; Sayed, A.A.; Peluso, I. Plant-Derived Smoke Solution Alleviates Cellular Oxidative Stress Caused by Arsenic and Mercury by Modulating the Cellular Antioxidative Defense System in Wheat. Plants 2022, 11, 1379. https://doi.org/10.3390/plants11101379
Ibrahim M, Nawaz S, Iqbal K, Rehman S, Ullah R, Nawaz G, Almeer R, Sayed AA, Peluso I. Plant-Derived Smoke Solution Alleviates Cellular Oxidative Stress Caused by Arsenic and Mercury by Modulating the Cellular Antioxidative Defense System in Wheat. Plants. 2022; 11(10):1379. https://doi.org/10.3390/plants11101379
Chicago/Turabian StyleIbrahim, Muhammad, Sadam Nawaz, Khalid Iqbal, Shafiq Rehman, Riaz Ullah, Ghazala Nawaz, Rafa Almeer, Amany A. Sayed, and Ilaria Peluso. 2022. "Plant-Derived Smoke Solution Alleviates Cellular Oxidative Stress Caused by Arsenic and Mercury by Modulating the Cellular Antioxidative Defense System in Wheat" Plants 11, no. 10: 1379. https://doi.org/10.3390/plants11101379
APA StyleIbrahim, M., Nawaz, S., Iqbal, K., Rehman, S., Ullah, R., Nawaz, G., Almeer, R., Sayed, A. A., & Peluso, I. (2022). Plant-Derived Smoke Solution Alleviates Cellular Oxidative Stress Caused by Arsenic and Mercury by Modulating the Cellular Antioxidative Defense System in Wheat. Plants, 11(10), 1379. https://doi.org/10.3390/plants11101379