
Physiology and Gene Expression Analysis of Potato (Solanum tuberosum L.) in Salt Stress

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
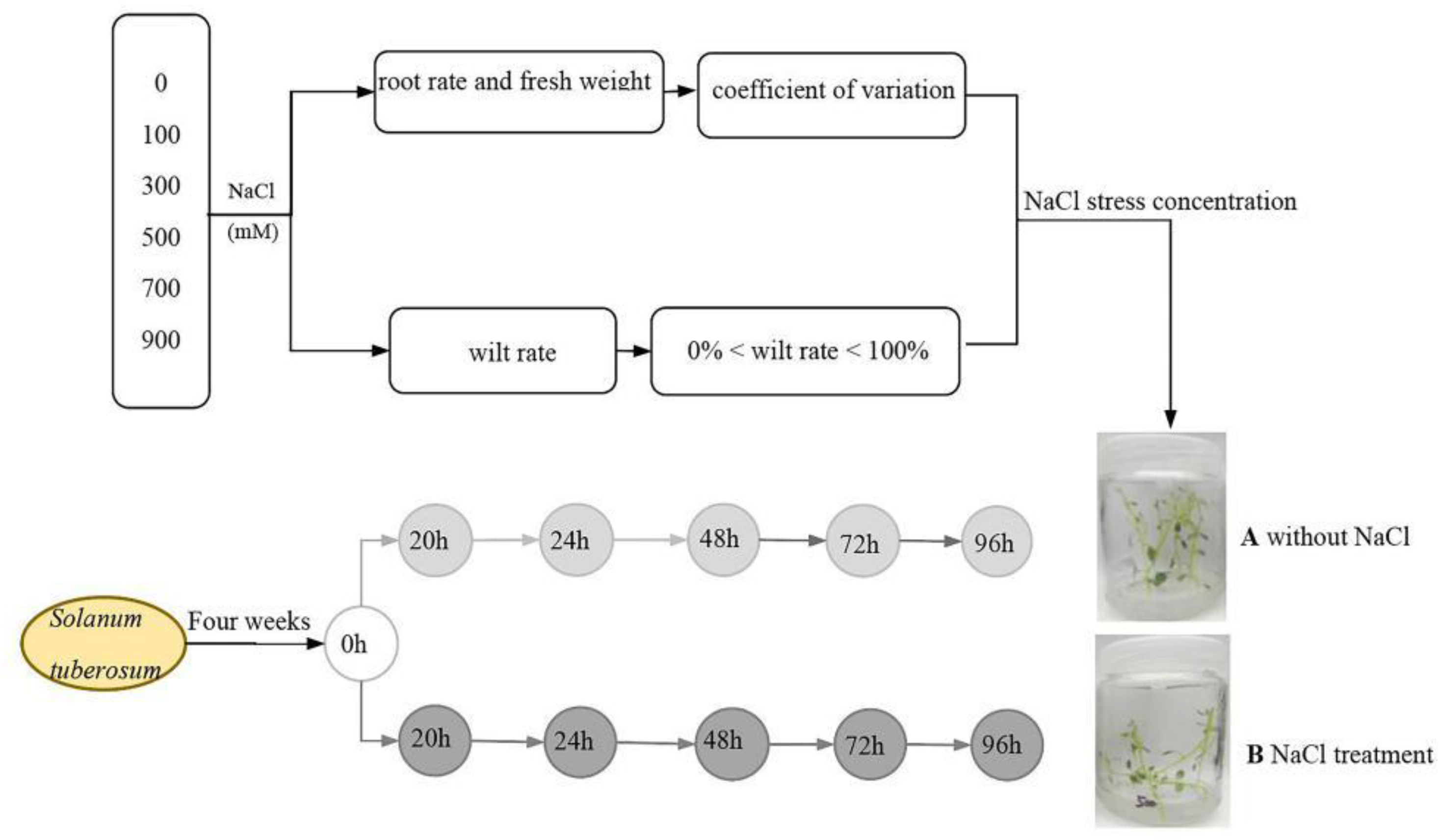
2.1. Screening the Appropriate Salt Concentration for Physiological Measurements
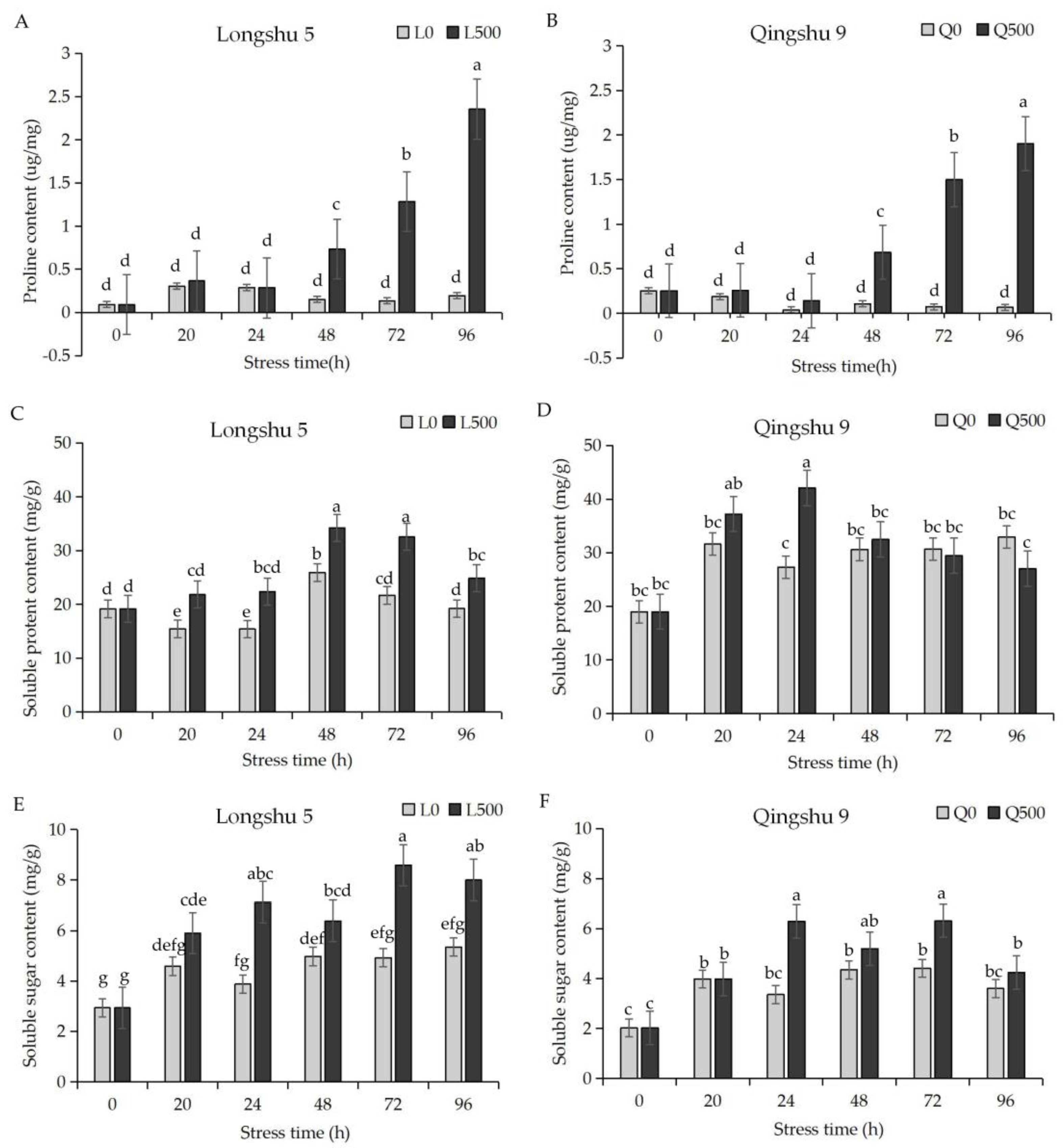
2.2. Proline, Soluble Protein, and Soluble Sugar Respond Differently to Salt Stress
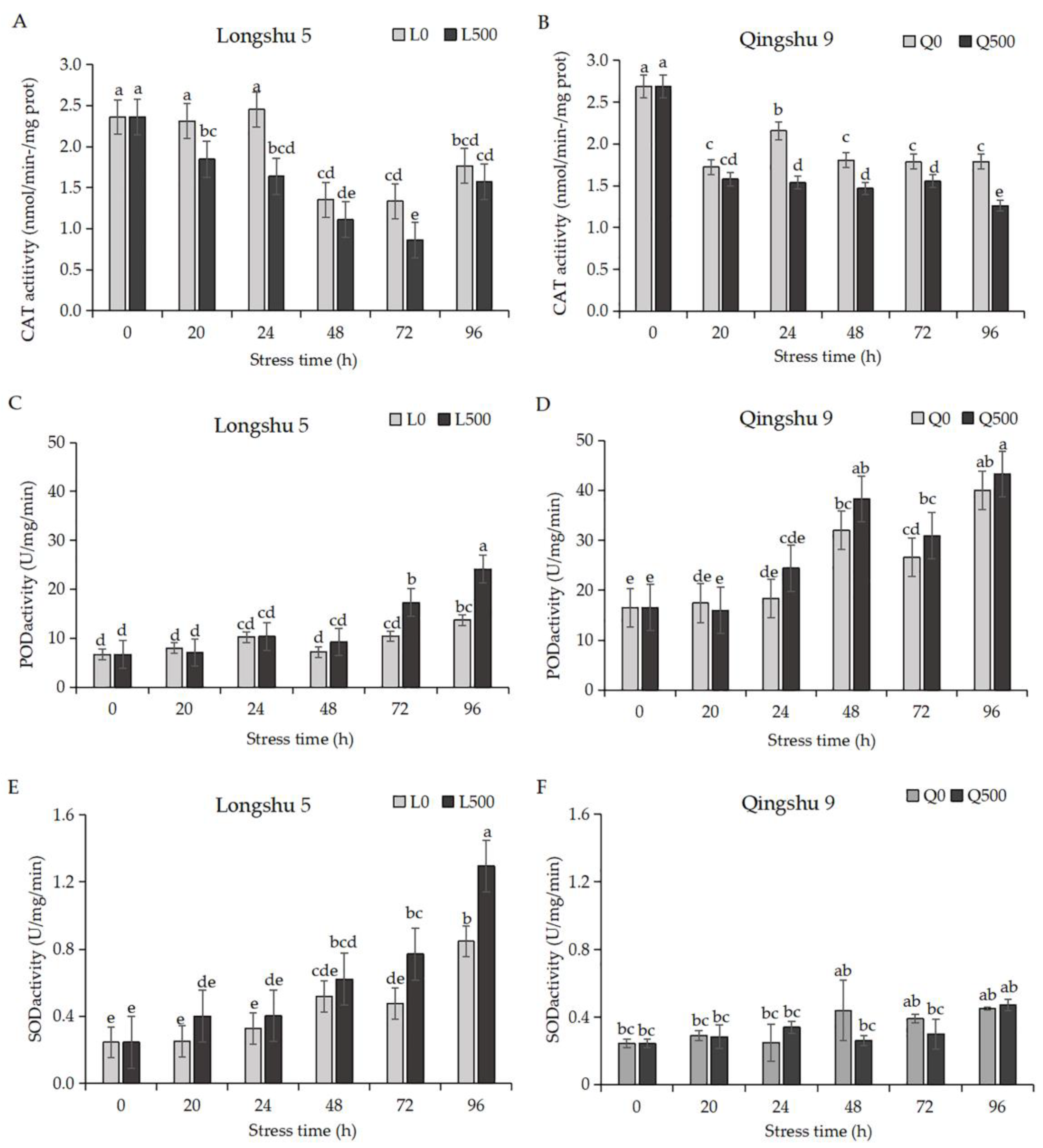
2.3. Antioxidant Enzyme Activities and MDA Respond Differently to Salt Stress
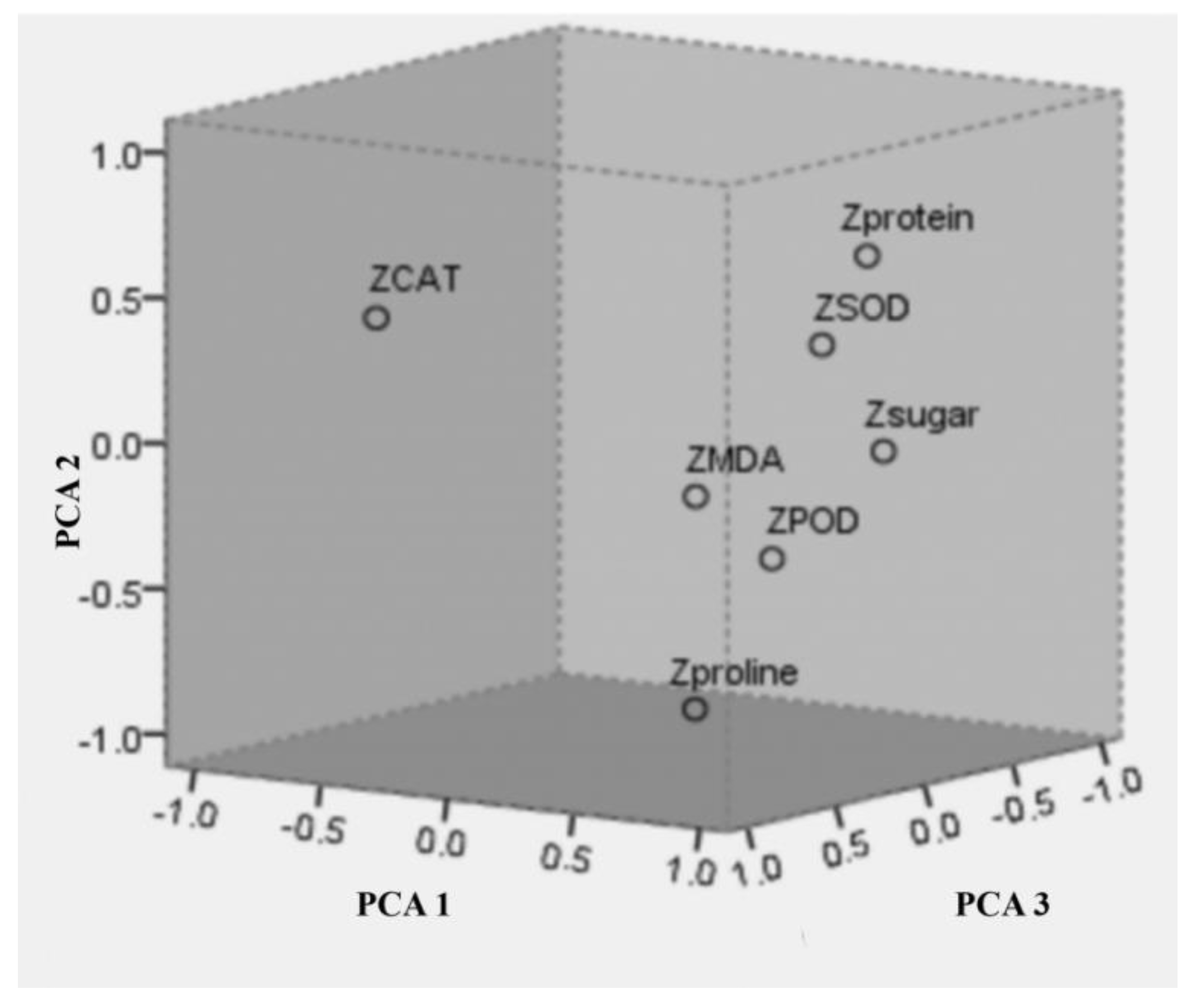
2.4. Principal Component Analysis of Salt Tolerance of Different Cultivars
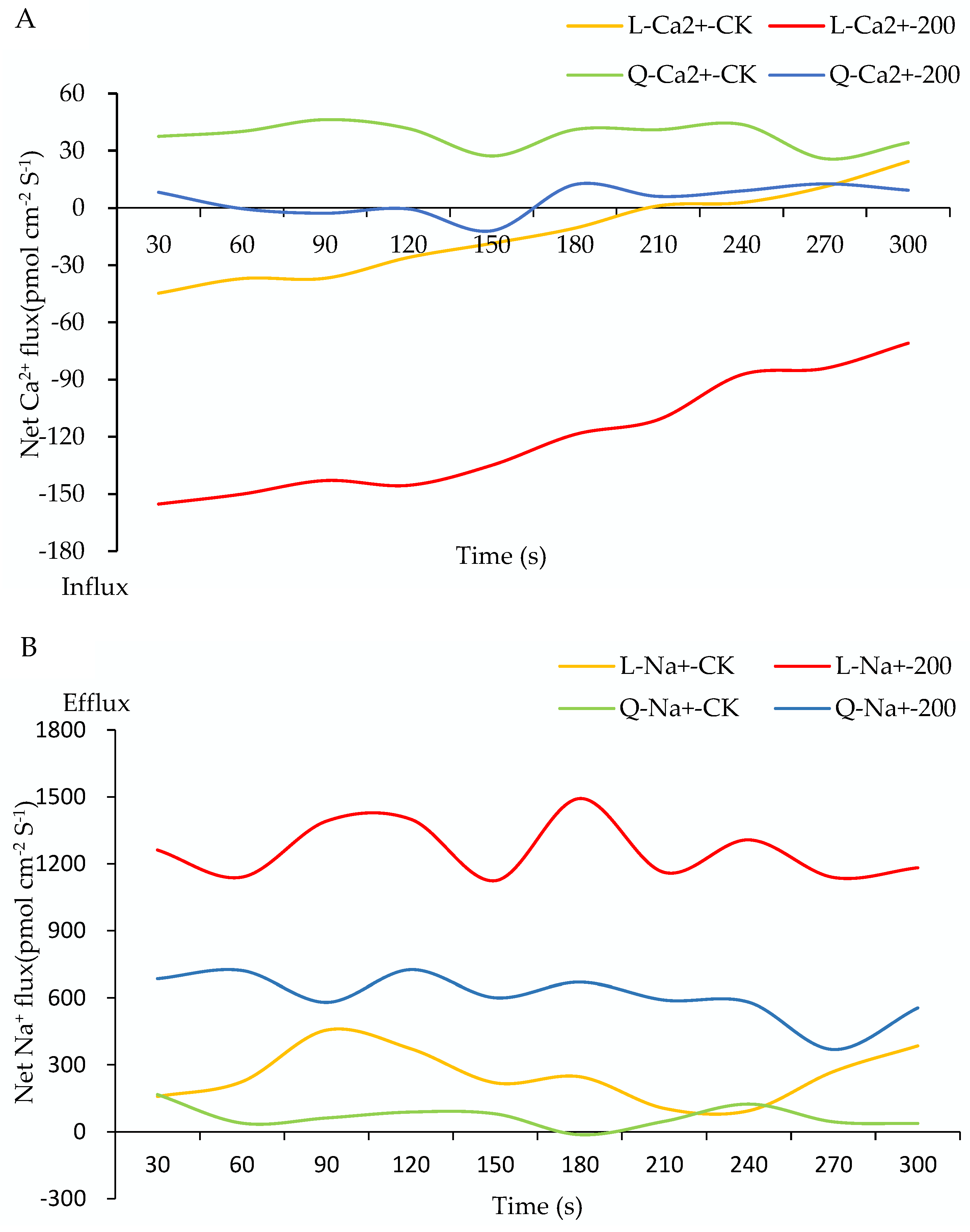
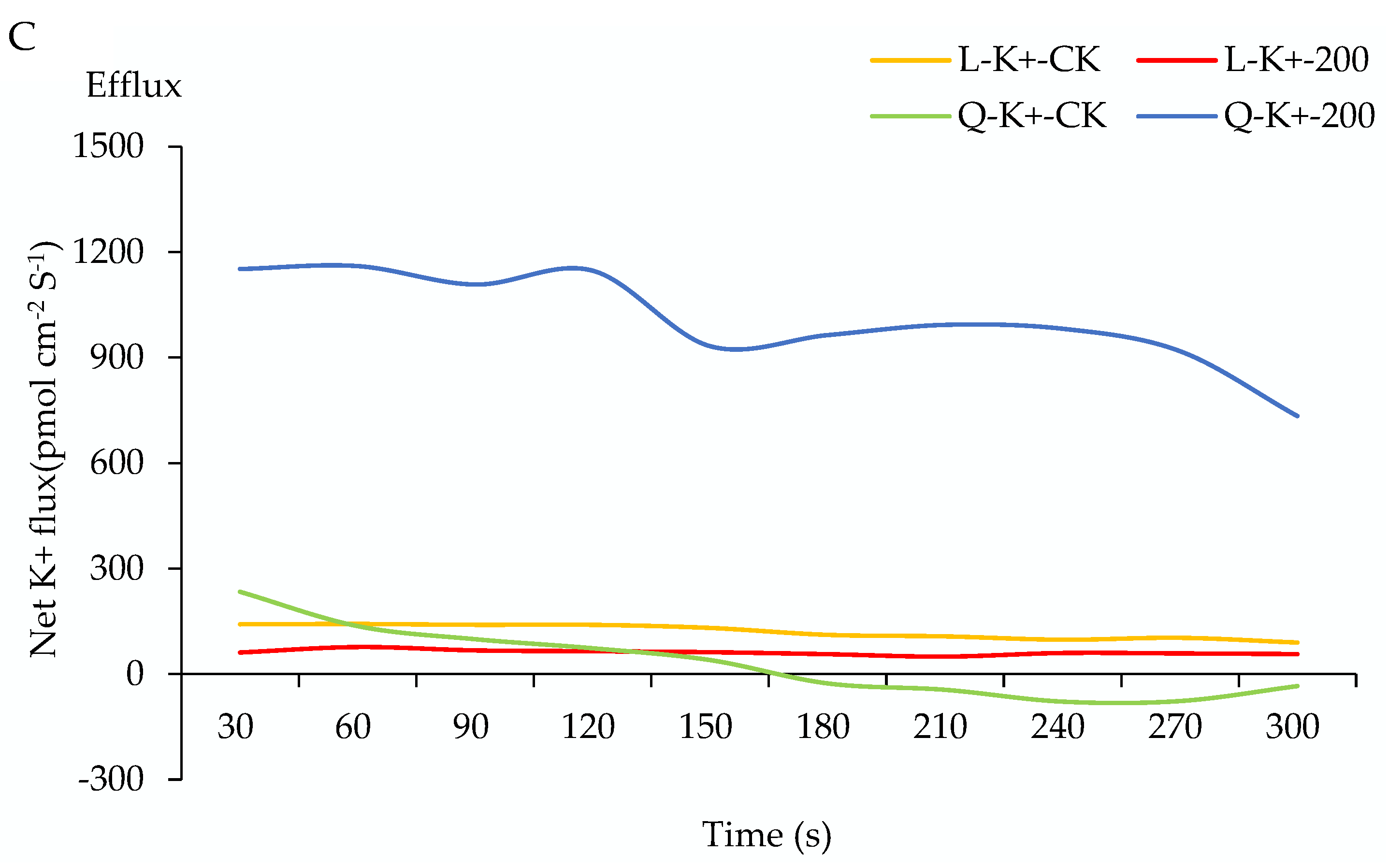
2.5. Na+, K+, and Ca2+ Flux of Potato Root under Salt Stress
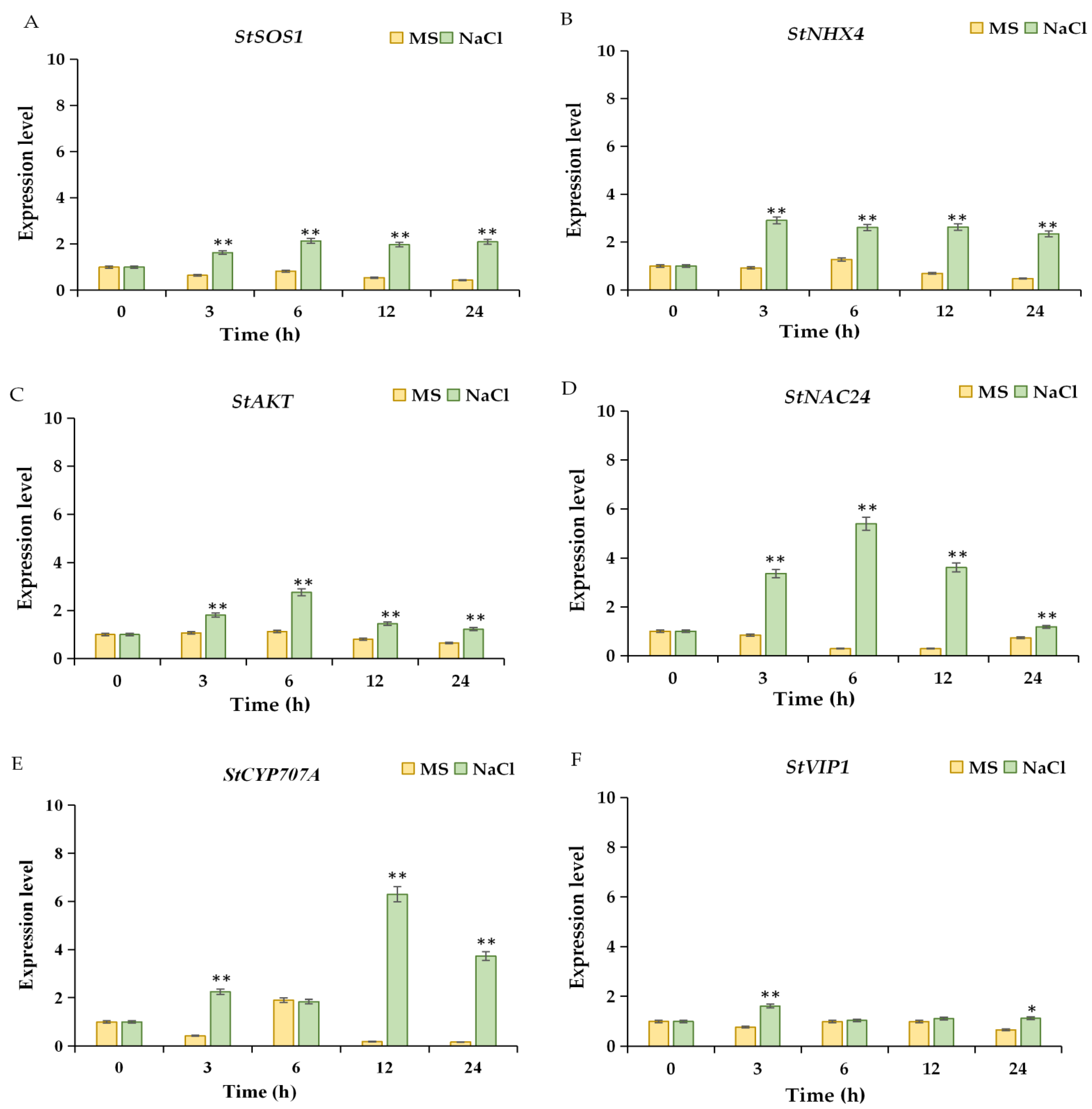
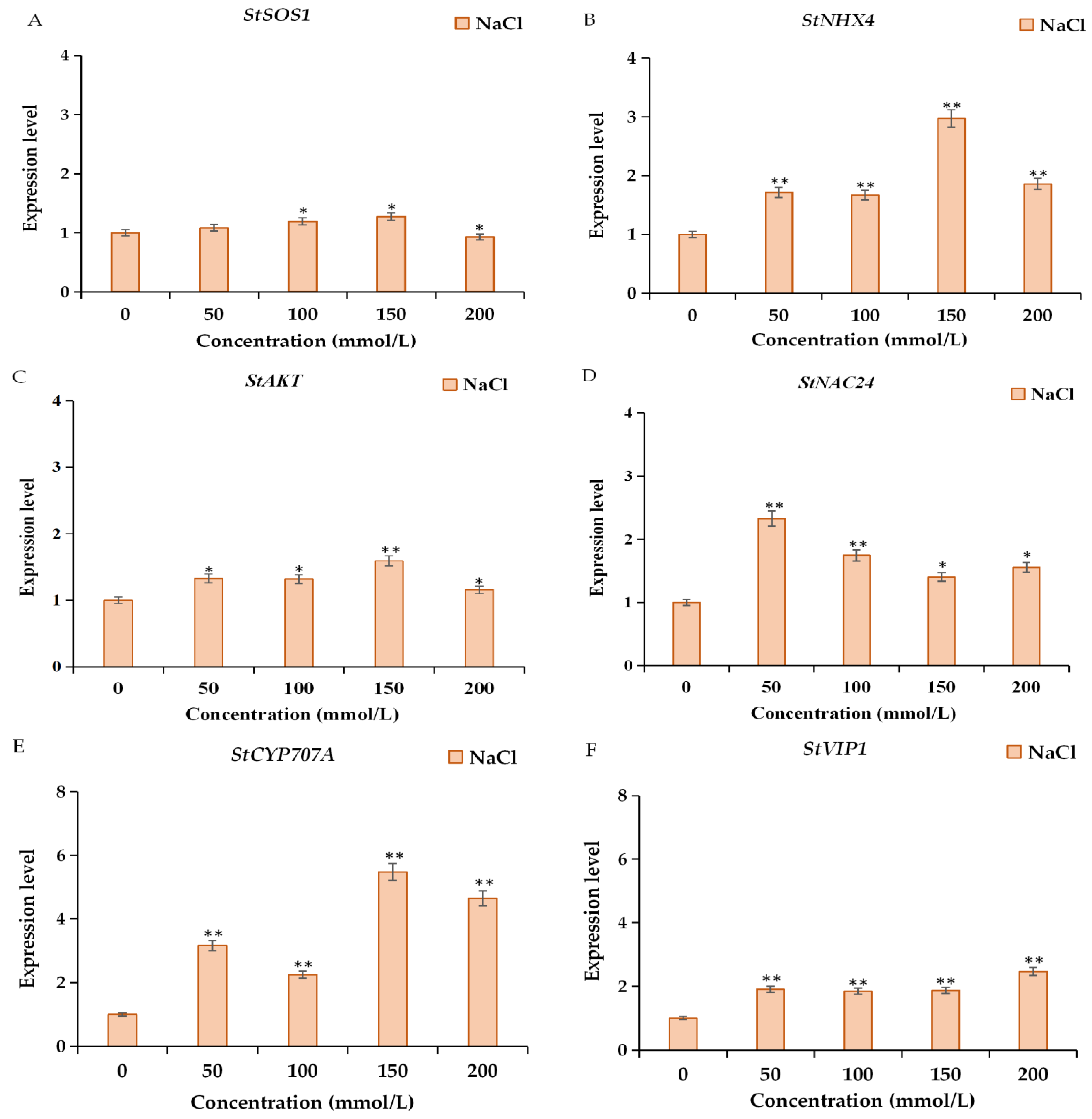
2.6. Expression Analysis of Salt-Response-Related Genes and Transcription Factors under NaCl Treatment in a Time Course
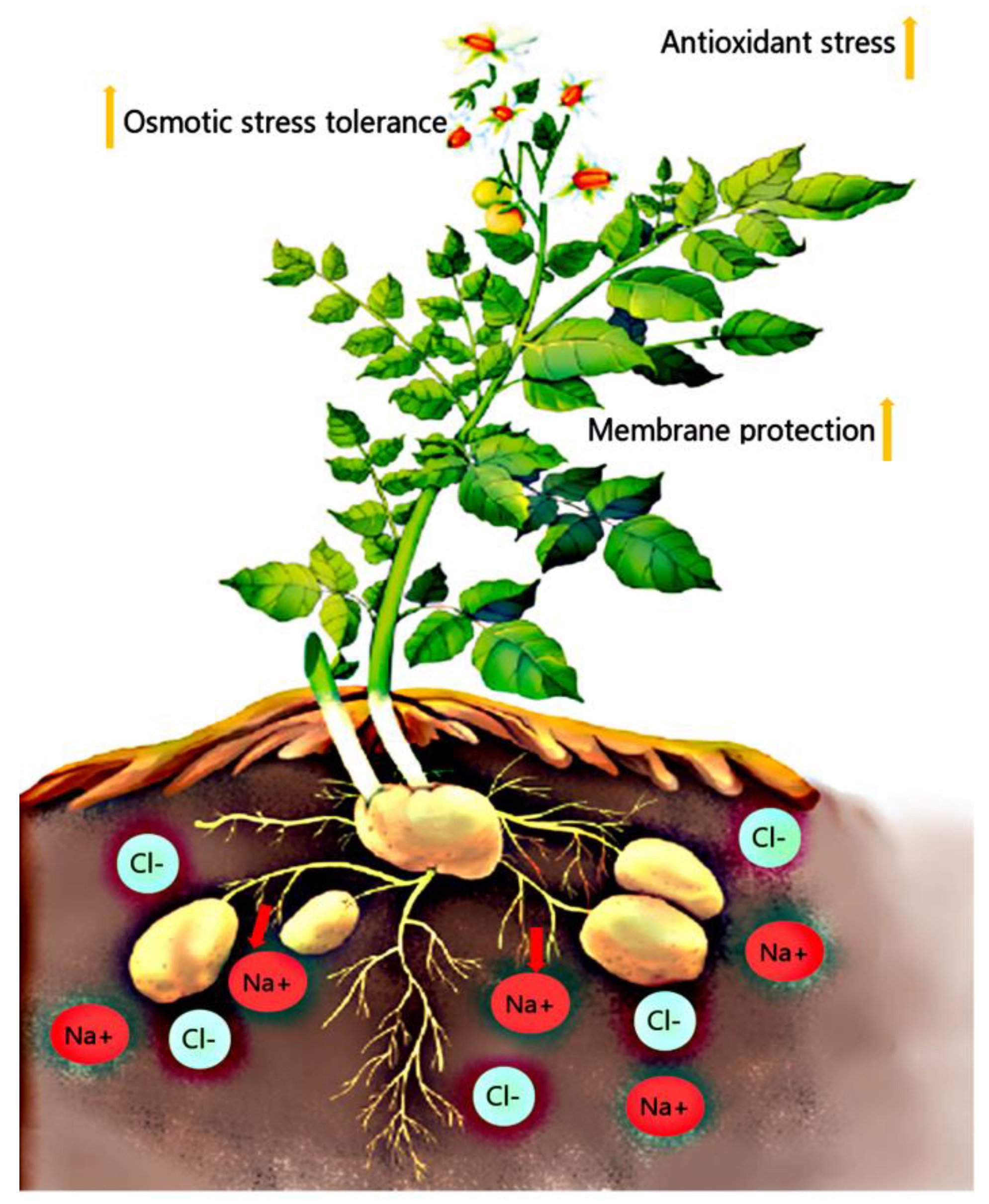
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Salt-Stress Treatments
4.2.1. Selection of Appropriate Salt-Stress Concentration for Physiological Measurement
4.2.2. Salt Treatment for Physiological Indicator Determination
4.2.3. Salt Treatment for Na+, K+, and Ca2+ Flux Determination
4.2.4. Expression Level of Ion Transport-Related Genes and Transcription Factors in ‘Longshu 5’ under NaCl
4.3. Determination of Physiological and Biochemical Indicators
4.3.1. Homogenized Protein Extraction
4.3.2. Proline Content, Soluble Protein, and Soluble Sugar Determination
4.3.3. Determination of Antioxidant Enzyme Activity
4.3.4. MDA Content Determination
4.4. Principal Component Analysis
4.5. Ca2+, Na+, and K+ Flux Determination
4.6. Quantitative Real-Time PCR (qRT-PCR) of Ion Transport-Related Genes
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, S.; Li, X.W.; Fan, S.D.; Zhou, L.J.; Wang, Y. Overexpression of HcSCL13, a Halostachys caspica GRAS transcription factor, enhances plant growth and salt stress tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2020, 151, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Faried, H.N.; Ayyub, C.M.; Wattoo, F.M.; Bashir, M.; Razzaq, K.; Akhtar, G.; Hussain, A.; Ullah, S.; Wattoo, J.I.; Amin, M.; et al. Assessing salt tolerance induction in potato by salicylic acid using morpho-physio-biochemical, ionic, and yield indices. Potato Res. 2022. [Google Scholar] [CrossRef]
- Dahal, K.; Li, X.Q.; Tai, H.; Creelman, A.; Bizimungu, B. Improving potato stress tolerance and tuber yield under a climate change scenario—A current overview. Front. Plant Sci. 2019, 10, 563. [Google Scholar] [CrossRef] [PubMed]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.S.; Zhang, Q.K.; Liu, M.Y.; Zhou, H.P.; Ma, C.L.; Wang, P.P. Regulation of plant responses to salt stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- Carillo, P.; Cirillo, C.; De Micco, V.; Arena, C.; De Pascale, S.; Rouphael, Y. Morpho-anatomical, physiological and biochemical adaptive responses to saline water of Bougainvillea spectabilis Willd. trained to different canopy shapes. Agric. Water Manag. 2019, 212, 12–22. [Google Scholar] [CrossRef]
- Gao, H.J.; Yang, H.Y.; Bai, J.P.; Liang, X.Y.; Lou, Y.; Zhang, J.L.; Wang, D.; Zhang, J.L.; Niu, S.Q.; Chen, Y.L. Ultrastructural and physiological responses of potato (Solanum tuberosum L.) plantlets to gradient saline stress. Front. Plant Sci. 2015, 5, 787. [Google Scholar] [CrossRef]
- Chourasia, K.N.; Lal, M.K.; Tiwari, R.K.; Dev, D.; Kardile, H.B.; Patil, V.U.; Kumar, A.; Vanishree, G.; Kumar, D.; Bhardwaj, V.; et al. Salinity stress in potato: Understanding physiological, biochemical and molecular responses. Life 2021, 11, 545. [Google Scholar] [CrossRef]
- Jaarsma, R.; de Boer, A.H. Salinity tolerance of two potato cultivars (Solanum tuberosum) correlates with differences in vacuolar transport activity. Front. Plant Sci. 2018, 9, 737. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Fan, Y.; Shabala, S.; Li, C.D.; Lv, C.; Guo, B.J.; Xu, R.G.; Zhou, M.X. Understanding mechanisms of salinity tolerance in Barley by proteomic and biochemical analysis of near-isogenic lines. Int. J. Mol. Sci. 2020, 21, 1516. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Li, Z.Y.; Jiang, X.M.; Qin, Y. Effects of salt stress on photosynthetic fluorescence characteristics, antioxidant system, and osmoregulation of Coreopsis tinctoria Nutt. Hortscience 2021, 56, 1066–1072. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, A.; Qayyum, M.F.; Shahzad, A.N.; Gul, M.; Wakeel, A. Interactive effect of boron and salinity on growth, physiological and biochemical attributes of wheat (Triticum aestivum). Int. J. Agric. Biol. 2016, 18, 238–244. [Google Scholar] [CrossRef]
- Liu, J.H.; Fu, C.C.; Li, G.J.; Khan, M.N.; Wu, H.H. ROS homeostasis and plant salt tolerance: Plant nanobiotechnology updates. Sustainability 2021, 13, 3552. [Google Scholar] [CrossRef]
- Shaterian, J.; Waterer, D.R.; de Jong, H.; Tanino, K.K. Methodologies and traits for evaluating the salt tolerance in diploid potato clones. Am. J. Potato Res. 2008, 85, 93–100. [Google Scholar] [CrossRef]
- Velásquez, B.; Balzarini, M.; Taleisnik, E. Salt tolerance variability amongst Argentine Andean potatoes (Solanum tuberosum L. subsp. andigena). Potato Res. 2005, 48, 59–67. [Google Scholar] [CrossRef]
- Li, Q.; Qin, Y.Z.; Hu, X.X.; Ding, H.Y.; Xiong, X.Y.; Wang, W.X. Establishment of an optimized bioassay being valuable for determining salt tolerance in 52 potato germplasm accessions. J. Plant Genet. Resour. 2018, 19, 587–597. [Google Scholar]
- Bari, M.A.; Islam, S.M.S.; Rashid, M.H.O. In vitro screening for salt stress tolerance of native and exotic potato genotypes by morphological and physiological parameters. J. Bio-Sci. 2019, 28, 21–32. [Google Scholar]
- Li, Q.; Qin, Y.Z.; Hu, X.X.; Li, G.C.; Ding, H.Y.; Xiong, X.Y.; Wang, W.X. Transcriptome analysis uncovers the gene expression profile of salt-stressed potato (Solanum tuberosum L.). Sci. Rep. 2020, 10, 5411. [Google Scholar] [CrossRef]
- Hasan, A.; Hafiz, H.R.; Siddiqui, N.; Khatun, M.; Islam, R.; Mamun, A.A. Evaluation of wheat genotypes for salt tolerance based on some physiological traits. J. Crop Sci. Biotechnol. 2016, 18, 333–340. [Google Scholar] [CrossRef]
- Liu, R.X.; Kuang, J.; Gong, Q.; Hou, X.L. Principal component regression analysis with SPSS. Comput. Methods Programs Biomed. 2003, 71, 141–147. [Google Scholar] [CrossRef]
- Joshi, S.; Nath, J.; Singh, A.K.; Pareek, A.; Joshi, R. Ion transporters and their regulatory signal transduction mechanisms for salinity tolerance in plants. Physiol. Plant. 2022, 174, e13702. [Google Scholar] [CrossRef] [PubMed]
- Saddhe, A.A.; Mishra, A.K.; Kumar, K. Molecular insights into the role of plant transporters in salt stress response. Physiol. Plant. 2021, 173, 1481–1494. [Google Scholar] [CrossRef]
- Yu, Z.P.; Duan, X.B.; Luo, L.; Dai, S.J.; Ding, Z.J.; Xia, G.M. How plant hormones mediate salt stress responses. Trends Plant Sci. 2020, 25, 1117–1130. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.J.; Huang, J.J.; Wang, X.D.; Dang, H.; Jiang, T.B.; Han, Y.Z. Expression analysis of the NAC transcription factor family of populus in response to salt stress. Forests 2019, 10, 688. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.L.; Li, C.J.; Lu, X.D.; Zhao, X.B.; Yan, C.X.; Wang, J.; Sun, Q.X.; Shan, S.H. Comprehensive genomic characterization of NAC transcription factor family and their response to salt and drought stress in peanut. BMC Plant Biol. 2020, 20, 454. [Google Scholar] [CrossRef] [PubMed]
- He, W.J.; Yan, K.; Zhang, Y.; Bian, L.X.; Mei, H.M.; Han, G.X. Contrasting photosynthesis, photoinhibition and oxidative damage in honeysuckle (Lonicera japonica Thunb.) under iso-osmotic salt and drought stresses. Environ. Exp. Bot. 2021, 182, 104313. [Google Scholar] [CrossRef]
- Takeo, K.; Ito, T. Subcellular localization of VIP1 is regulated by phosphorylation and 14-3-3 proteins. FEBS Lett. 2017, 591, 1972–1981. [Google Scholar] [CrossRef]
- Yang, Z.J.; Wang, C.W.; Xue, Y.; Liu, X.; Chen, S.; Song, C.P.; Yang, Y.Q.; Guo, Y. Calcium-activated 14-3-3 proteins as a molecular switch in salt stress tolerance. Nat. Commun. 2019, 10, 1199. [Google Scholar] [CrossRef] [Green Version]
- Jaarsma, R.; de Vries, R.S.; de Boer, A.H. Effect of salt stress on growth, Na+ accumulation and proline metabolism in potato (Solanum tuberosum) cultivars. PLoS ONE 2013, 8, e60183. [Google Scholar]
- Jiang, F.Z.; Wang, J.; Zhang, Y.P.; Jia, H.; Shen, H.F. A new potato variety Qingshu 9 with high and stable yield, drought resistance and virus resistance. Crops 2008, 89. [Google Scholar] [CrossRef]
- Qayyum, M.A.; Akhtar, J.; Bashir, F.; Naz, T.; Iqbal, M.M.; Farooq, O.; Atique-ur, R.; Zafar, M.I.; Ali, M.; Imtiaz, M.; et al. Physiological and biochemical characterization of linseed genotypes under salinity stress. Int. J. Agric. Biol. 2020, 23, 630–636. [Google Scholar]
- D’Amelia, L.; Dell’Aversana, E.; Woodrow, P.; Ciarmiello, L.F.; Carillo, P. Metabolomics for crop improvement against salinity stress. In Salinity Responses and Tolerance in Plants; Kumar, V., Wani, S., Suprasanna, P., Tran, L.S., Eds.; Springer: Cham, Switzerland, 2018; Volume 2, pp. 267–287. [Google Scholar]
- Li, W.; Qiang, X.J.; Han, X.R.; Jiang, L.L.; Zhang, S.H.; Han, J.; He, R.; Cheng, X.G. Ectopic expression of a thellungiella salsuginea aquaporin gene, TsPIP1;1, increased the salt tolerance of rice. Int. J. Mol. Sci. 2018, 19, 2229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulen, H.; Turhan, E.; Plantarum, A.E.J.A.P. Changes in peroxidase activities and soluble proteins in strawberry varieties under salt-stress. Acta Physiol. Plant. 2006, 28, 109–116. [Google Scholar] [CrossRef]
- El Moukhtari, A.; Cabassa-Hourton, C.; Farissi, M.; Savoure, A. How does proline treatment promote salt stress tolerance during crop plant development? Front. Plant Sci. 2020, 11, 1127. [Google Scholar] [CrossRef] [PubMed]
- Kitayama, M.; Samphumphuang, T.; Tisarum, R.; Theerawitaya, C.; Cha-um, K.; Takagaki, M.; Cha-um, S. Calcium and soluble sugar enrichments and physiological adaptation to mild NaCl salt stress in sweet potato (Ipomoea batatas) genotypes. J. Hortic. Sci. Biotechnol. 2020, 95, 782–793. [Google Scholar] [CrossRef]
- Ghosh, S.C.; Asanuma, K.I.; Kusutani, A.; Toyota, M. Effect of salt stress on some chemical components and yield of potato. Soil Sci. Plant Nutr. 2001, 47, 467–475. [Google Scholar] [CrossRef]
- Ye, T.T.; Shi, H.T.; Wang, Y.P.; Yang, F.; Chan, Z.L. Contrasting proteomic and metabolomic responses of bermudagrass to drought and salt stresses. Front. Plant Sci. 2016, 7, 1694. [Google Scholar] [CrossRef] [Green Version]
- Heidari, M.; Mesri, F. Salinity effects on compatible solutes, antioxidants enzymes and ion content in three wheat cultivars. Pak. J. Biol. Sci. 2008, 11, 1385–1389. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Guo, Y. Unraveling salt stress signaling in plants. J. Integr. Plant Biol. 2018, 60, 796–804. [Google Scholar] [CrossRef] [Green Version]
- Charfeddine, M.; Charfeddine, S.; Ghazala, I.; Bouaziz, D.; Bouzid, R.G. Investigation of the response to salinity of transgenic potato plants overexpressing the transcription factor StERF94. J. Biosci. 2019, 44, 141. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, E.A.; Osama, E.; Manal, E.; Samah, A.; Salah, G.; Hazem, K.M.; Jacek, W.; Nabil, E. Impact of gamma irradiation pretreatment on biochemical and molecular responses of potato growing under salt stress. Chem. Biol. Technol. Agric. 2021, 8, 35. [Google Scholar] [CrossRef]
- Jbir-Koubaa, R.; Charfeddine, S.; Bouaziz, D.; Ben Mansour, R.; Gargouri-Bouzid, R.; Nouri-Ellouz, O. Enhanced antioxidant enzyme activities and respective gene expressions in potato somatic hybrids under NaCl stress. Biol. Plant. 2019, 63, 633–642. [Google Scholar] [CrossRef]
- Yasar, F.; Ellialtioglu, S.; Yildiz, K. Effect of salt stress on antioxidant defense systems, lipid peroxidation, and chlorophyll content in green bean. Russ. J. Plant Physiol. 2008, 55, 782–786. [Google Scholar] [CrossRef]
- Monihan, S.M.; Ryu, C.H.; Magness, C.A.; Schumaker, K.S. Linking duplication of a calcium sensor to salt tolerance in Eutrema salsugineum. Plant Physiol. 2019, 179, 1176–1192. [Google Scholar] [CrossRef] [Green Version]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.Y.; Li, J.; Wang, P.-Y.; Qin, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Jbir-Koubaa, R.; Charfeddine, S.; Ellouz, W.; Saidi, M.N.; Drira, N.; Gargouri-Bouzid, R.; Nouri-Ellouz, O. Investigation of the response to salinity and to oxidative stress of inter specific potato somatic hybrids grown in a greenhouse. Plant Cell Tissue Organ Cult. 2015, 120, 933–947. [Google Scholar] [CrossRef]
- Zhou, H.P.; Lin, H.X.; Chen, S.; Becker, K.; Yang, Y.Q.; Zhao, J.F.; Kudla, J.; Schumaker, K.S.; Guo, Y. Inhibition of the arabidopsis salt overly sensitive pathway by 14-3-3 proteins. Plant Cell 2014, 26, 1166–1182. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Ye, J.M.; Yang, Y.Q.; Lin, H.X.; Yue, L.L.; Luo, J.; Long, Y.; Fu, H.Q.; Liu, X.N.; Zhang, Y.L.; et al. The SOS2-SCaBP8 Complex Generates and Fine-Tunes an AtANN4-dependent calcium signature under salt stress. Dev. Cell 2019, 48, 697–709. [Google Scholar] [CrossRef] [Green Version]
- Tsugama, D.; Liu, S.K.; Takano, T. Analysis of functions of VIP1 and its close homologs in osmosensory responses of Arabidopsis thaliana. PLoS ONE 2014, 9, e103930. [Google Scholar] [CrossRef]
- Zheng, Y.; Huang, Y.; Xian, W.; Wang, J.; Liao, H. Identification and expression analysis of the glycine max CYP707A gene family in response to drought and salt stresses. Ann. Bot. 2012, 110, 743–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, S. Variation in antioxidant metabolism of young and mature leaves of Arabidopsis thaliana subjected to drought. Plant Sci. 2004, 166, 459–466. [Google Scholar] [CrossRef]
- Wang, B.; Guo, X.; Zhao, P.J.; Ruan, M.B.; Yu, X.L.; Zou, L.P.; Yang, Y.L.; Li, X.; Deng, D.L.; Xiao, J.X.; et al. Molecular diversity analysis, drought related marker-traits association mapping and discovery of excellent alleles for 100-day old plants by EST-SSRs in cassava germplasms (Manihot esculenta Cranz). PLoS ONE 2017, 12, e0177456. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.R.; Zhao, P.S.; Zhao, X.; Wang, X.P.; Ma, X.F.; Li, Y. Physiological adaptations to osmotic stress and characterization of a polyethylene glycol-responsive gene in Braya humilis. Acta Soc. Bot. Pol. Pol. Tow. Bot. 2016, 85, 3487. [Google Scholar]
- Xing, W.; Li, D.H.; Liu, G.H. Antioxidative responses of Elodea nuttallii (Planch.) H. St. John to short-term iron exposure. Plant Physiol. Biochem. 2010, 48, 873–878. [Google Scholar] [CrossRef]
- Zhang, S.Z.; Hua, B.Z.; Zhang, F. Induction of the activities of antioxidative enzymes and the levels of malondialdehyde in cucumber seedlings as a consequence of Bemisia tabaci (Hemiptera: Aleyrodidae) infestation. Arthropod-Plant Interact. 2008, 2, 209–213. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salt Concentration (mmol/L) | Root Percentage (%) | Fresh Weight (g) | Total CV(%) | Wilt Rate (%) | |||
|---|---|---|---|---|---|---|---|
| Longshu 5 | Qingshu 9 | Longshu 5 | Qingshu 9 | Longshu 5 | Qingshu 9 | ||
| 0 | 100 | 100 | 0.715 | 2.678 | 57.6 | 0.0 | 0.0 |
| 100 | 100 | 100 | 0.555 | 1.483 | 32.5 | 0.0 | 0.0 |
| 300 | 0.0 | 0.0 | 0.442 | 1.306 | 122.0 | 0.0 | 0.0 |
| 500 | 0.0 | 0.0 | 0.414 | 1.227 | 122.1 | 26.67 | 33.33 |
| 700 | 0.0 | 0.0 | 0.385 | 0.917 | 115.5 | 100 | 100 |
| 900 | 0.0 | 0.0 | 0.333 | 0.746 | 113.7 | 100 | 100 |
| Variety | NaCl Concentration (mmol/L) | MDA Content (nmol/g) | |||||
|---|---|---|---|---|---|---|---|
| 0 h | 20 h | 24 h | 48 h | 72 h | 96 h | ||
| Longshu 5 | 0 | 8.69 ± 0.91 d | 9.80 ± 1.36 bcd | 8.60 ± 0.59 d | 9.89 ± 0.29 bcd | 9.29 ± 0.1 cd | 9.12 ± 0.40 cd |
| 500 | 8.69 ± 0.91 d | 9.80 ± 0.93 bcd | 9.89 ± 1.05 bcd | 10.75 ± 0.39 bc | 11.10 ± 0.93 b | 12.90 ±1.29 a | |
| Qingshu 9 | 0 | 12.51 ±1.16 b | 11.86 ± 0.78 b | 12.39 ± 1.34 b | 12.25 ± 1.42 b | 12.90 ± 1.29 b | 11.22 ± 0.13 b |
| 500 | 12.51± 1.16 b | 10.92 ± 1.27 b | 13.07 ± 1.30 b | 16.17 ± 0.91 a | 12.38 ± 0.78 b | 11.61 ± 1.80 b | |
| Component | Initial Eigenvalues | Extraction Sums of Squared Loadings | ||||
|---|---|---|---|---|---|---|
| Total | % of Variance | Cumulative % | Total | % of Variance | Cumulative % | |
| PCA1 | 3.144 | 44.912 | 44.912 | 3.144 | 44.912 | 44.912 |
| PCA2 | 1.716 | 24.511 | 69.423 | 1.716 | 24.511 | 69.423 |
| PCA3 | 1.025 | 14.639 | 84.062 | 1.025 | 14.639 | 84.062 |
| PCA4 | 0.636 | 9.089 | 93.151 | |||
| PCA5 | 0.285 | 4.078 | 97.229 | |||
| PCA6 | 0.136 | 1.937 | 99.166 | |||
| PCA7 | 0.058 | 0.834 | 100 | |||
| Cultivar | Time (h) | Principal Component Scores | Composite Scores | Total Score | ||
|---|---|---|---|---|---|---|
| PCA 1 | PCA 2 | PCA 3 | ||||
| Longshu 5 | 0 | −0.245 | −0.280 | −1.376 | −0.337 | |
| 20 | 1.139 | −0.197 | −0.362 | −0.002 | ||
| 24 | 0.758 | −0.886 | 1.439 | 0.374 | ||
| 48 | −0.220 | −0.818 | 0.679 | 0.058 | 1.703 | |
| 72 | 1.246 | −0.173 | 0.115 | 0.805 | ||
| 96 | 1.211 | 1.072 | −1.177 | 0.805 | ||
| Qingshu 9 | 0 | −0.245 | −0.280 | −1.376 | −0.337 | |
| 20 | −0.498 | −0.698 | −0.433 | −0.386 | ||
| 24 | 0.133 | −0.841 | 1.025 | 0.467 | −1.703 | |
| 48 | −2.130 | −0.531 | 0.059 | 0.015 | ||
| 72 | −1.063 | 1.591 | 0.024 | −0.649 | ||
| 96 | −0.087 | 2.040 | 1.383 | −0.813 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Qin, Y.; Hu, X.; Jin, L.; Li, G.; Gong, Z.; Xiong, X.; Wang, W. Physiology and Gene Expression Analysis of Potato (Solanum tuberosum L.) in Salt Stress. Plants 2022, 11, 1565. https://doi.org/10.3390/plants11121565
Li Q, Qin Y, Hu X, Jin L, Li G, Gong Z, Xiong X, Wang W. Physiology and Gene Expression Analysis of Potato (Solanum tuberosum L.) in Salt Stress. Plants. 2022; 11(12):1565. https://doi.org/10.3390/plants11121565
Chicago/Turabian StyleLi, Qing, Yuzhi Qin, Xinxi Hu, Liping Jin, Guangcun Li, Zhenping Gong, Xingyao Xiong, and Wanxing Wang. 2022. "Physiology and Gene Expression Analysis of Potato (Solanum tuberosum L.) in Salt Stress" Plants 11, no. 12: 1565. https://doi.org/10.3390/plants11121565
APA StyleLi, Q., Qin, Y., Hu, X., Jin, L., Li, G., Gong, Z., Xiong, X., & Wang, W. (2022). Physiology and Gene Expression Analysis of Potato (Solanum tuberosum L.) in Salt Stress. Plants, 11(12), 1565. https://doi.org/10.3390/plants11121565