
Potential Impacts of Climate Change on the Habitat Suitability of the Dominant Tree Species in Greece

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
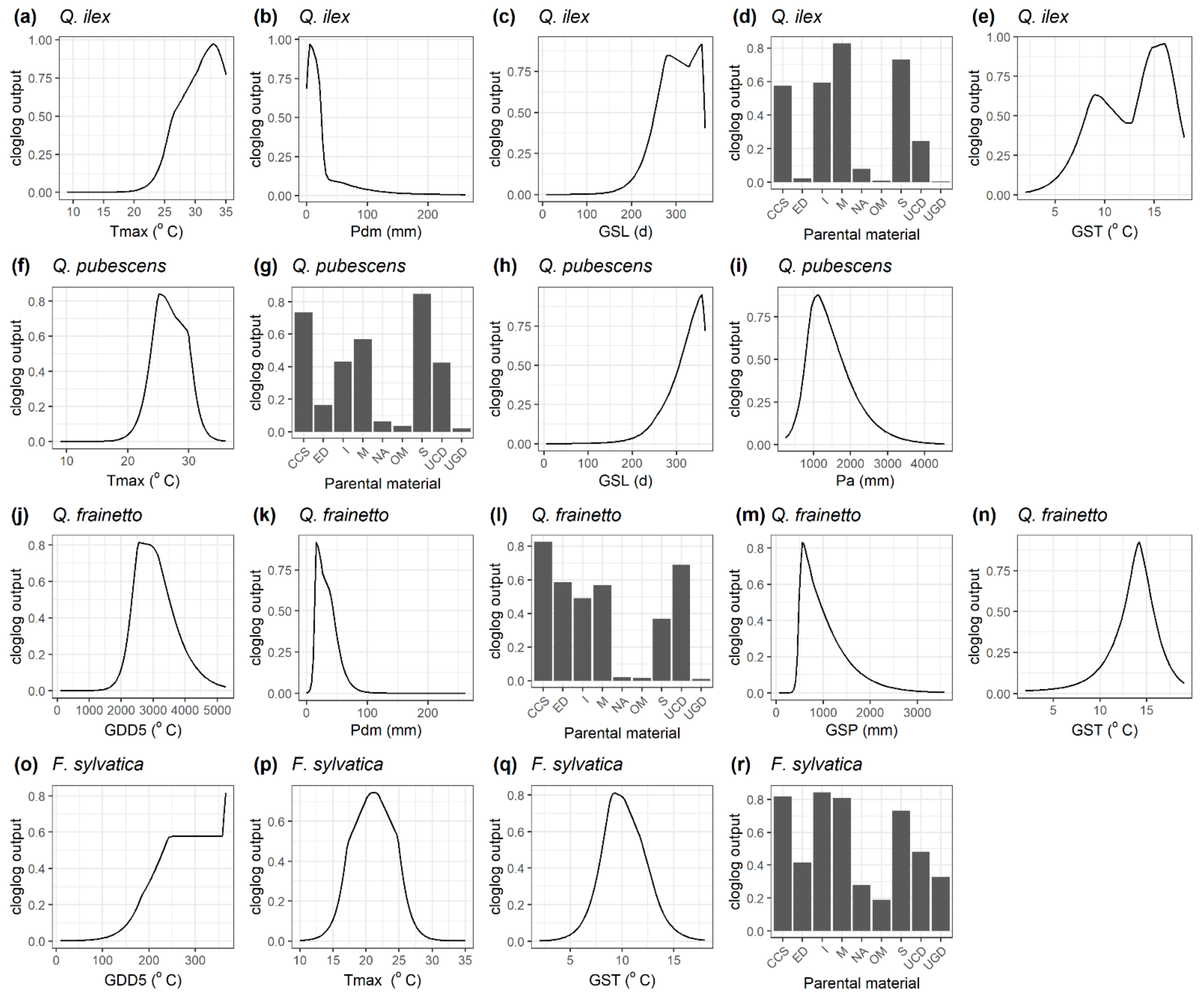
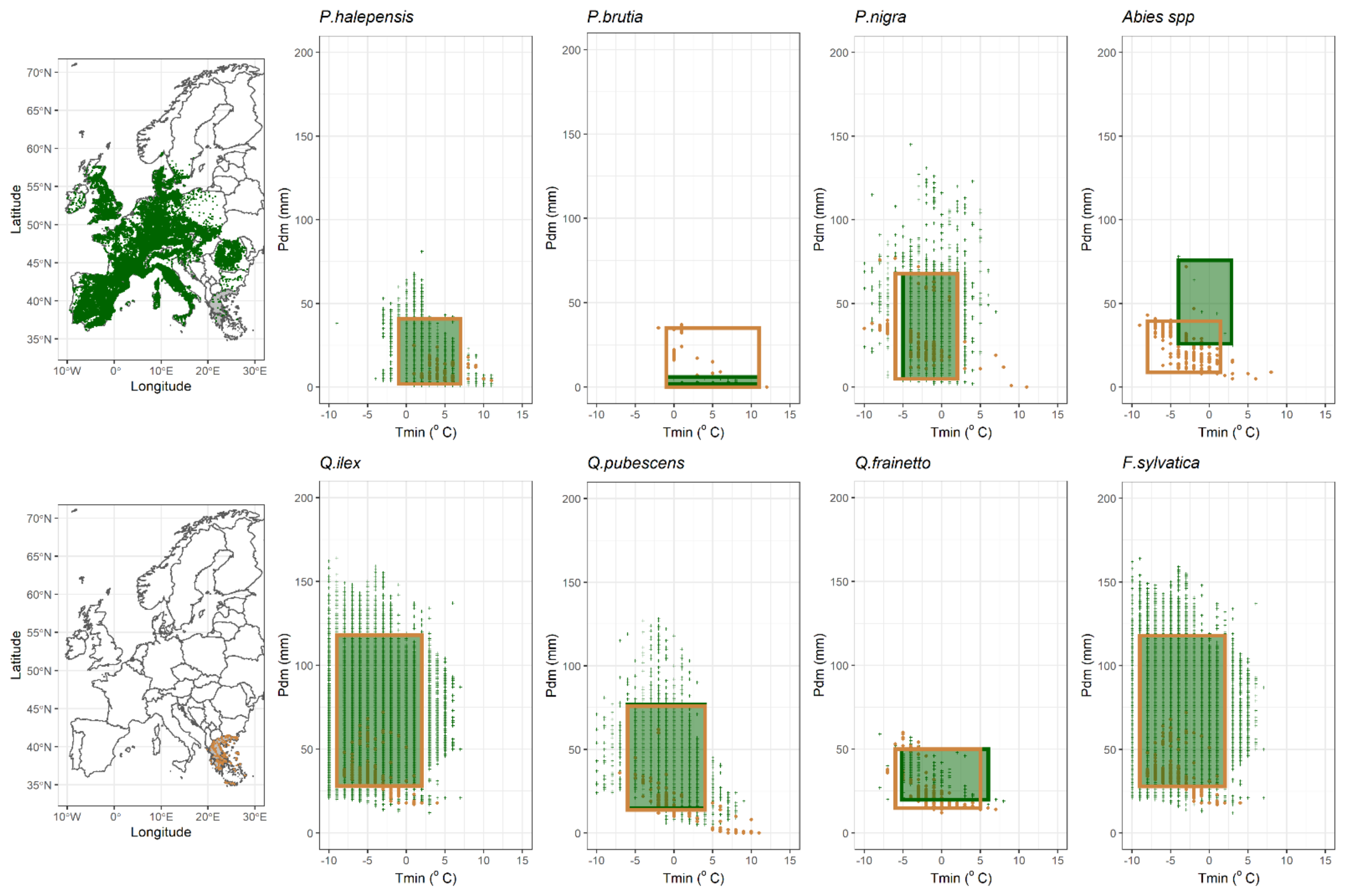
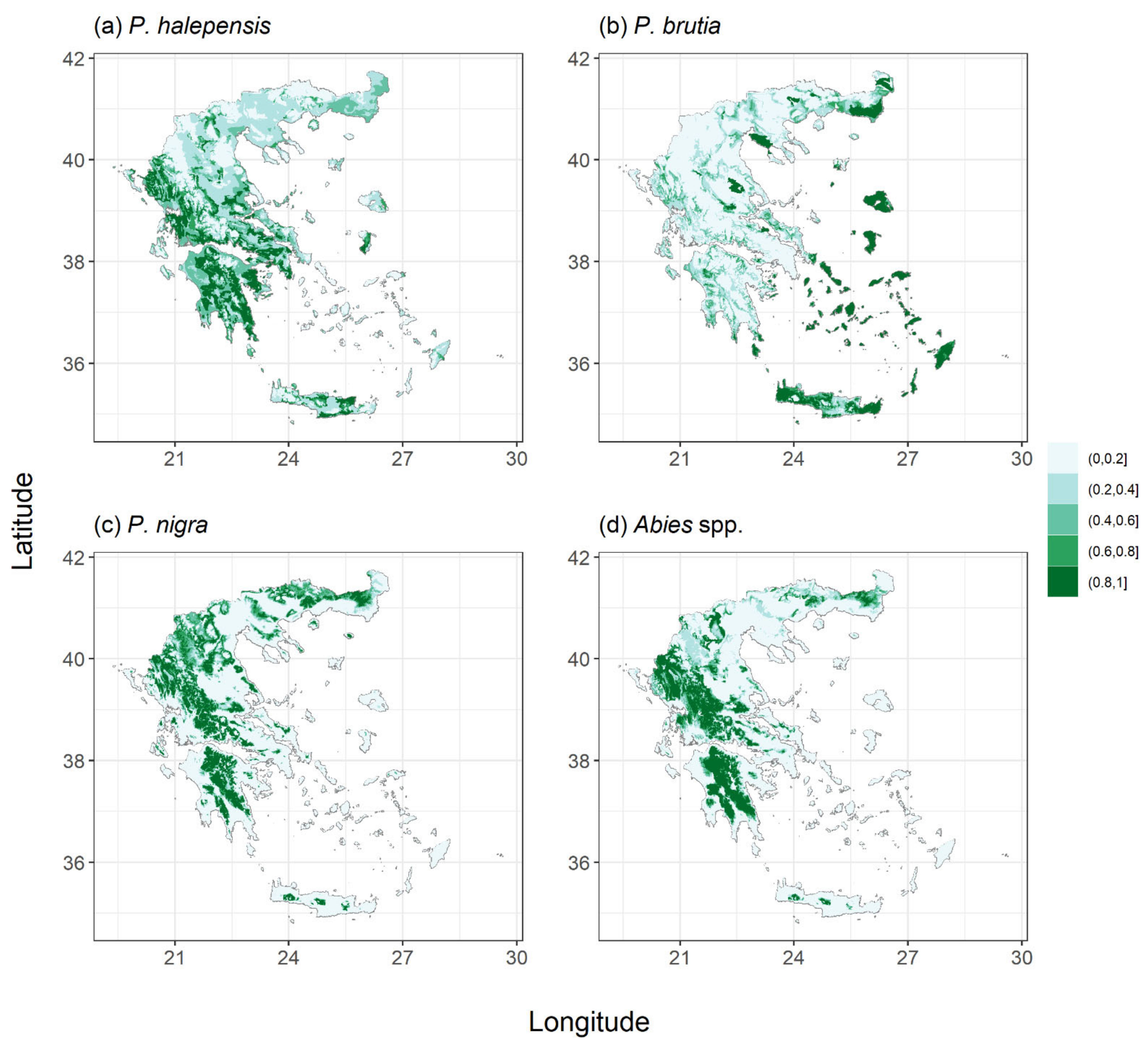
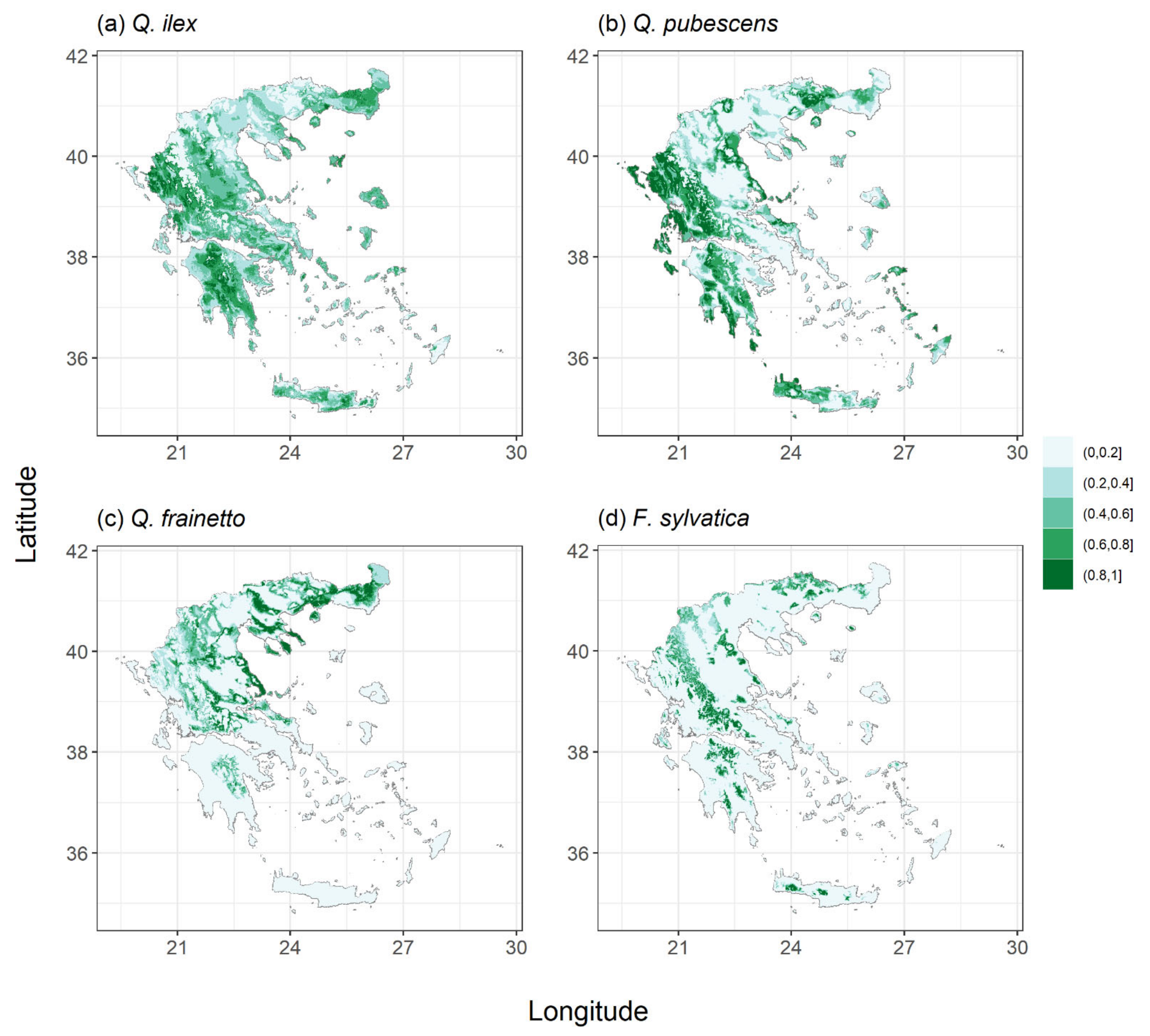
2.1. Species Habitat Suitability under Current Conditions
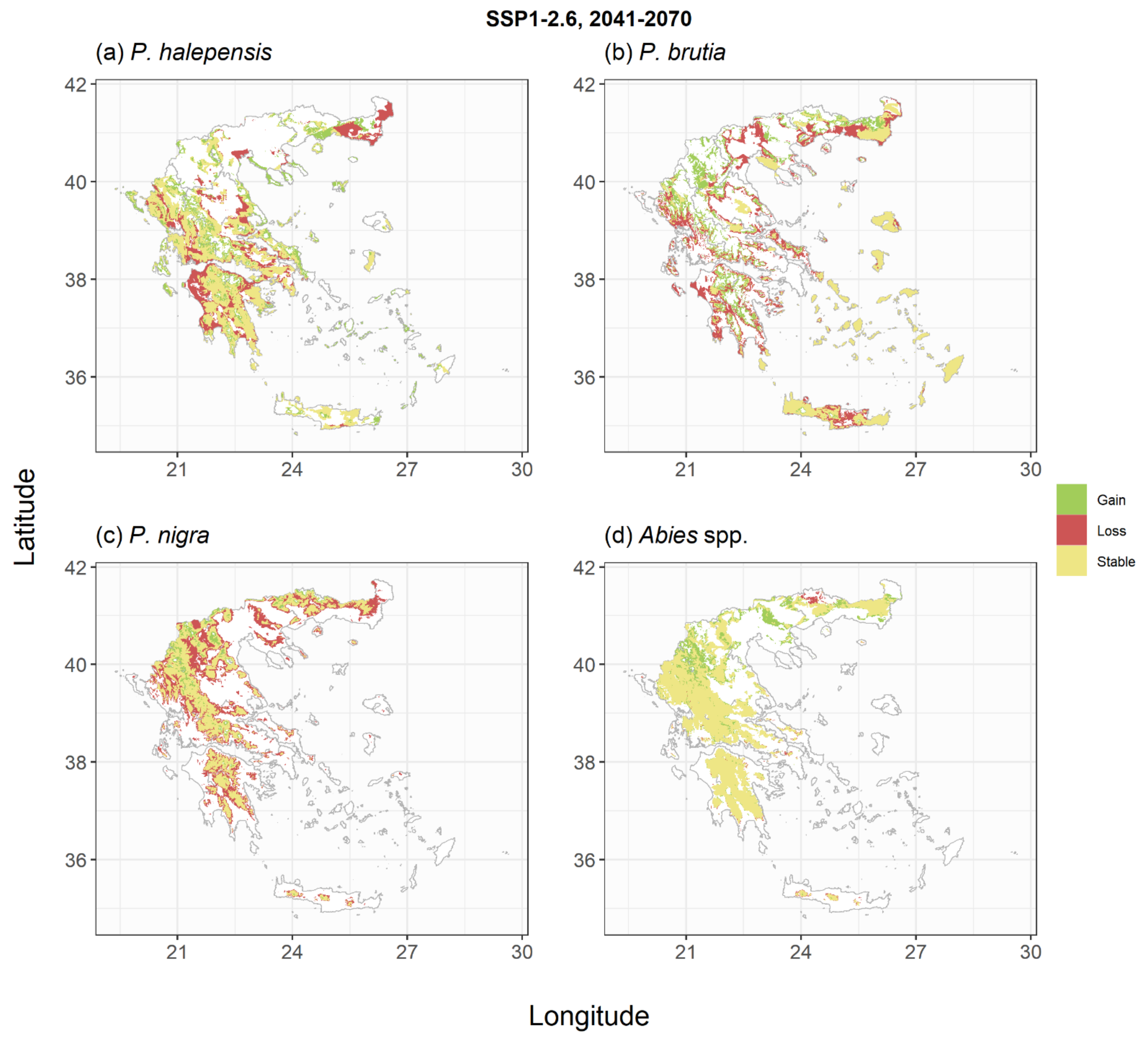
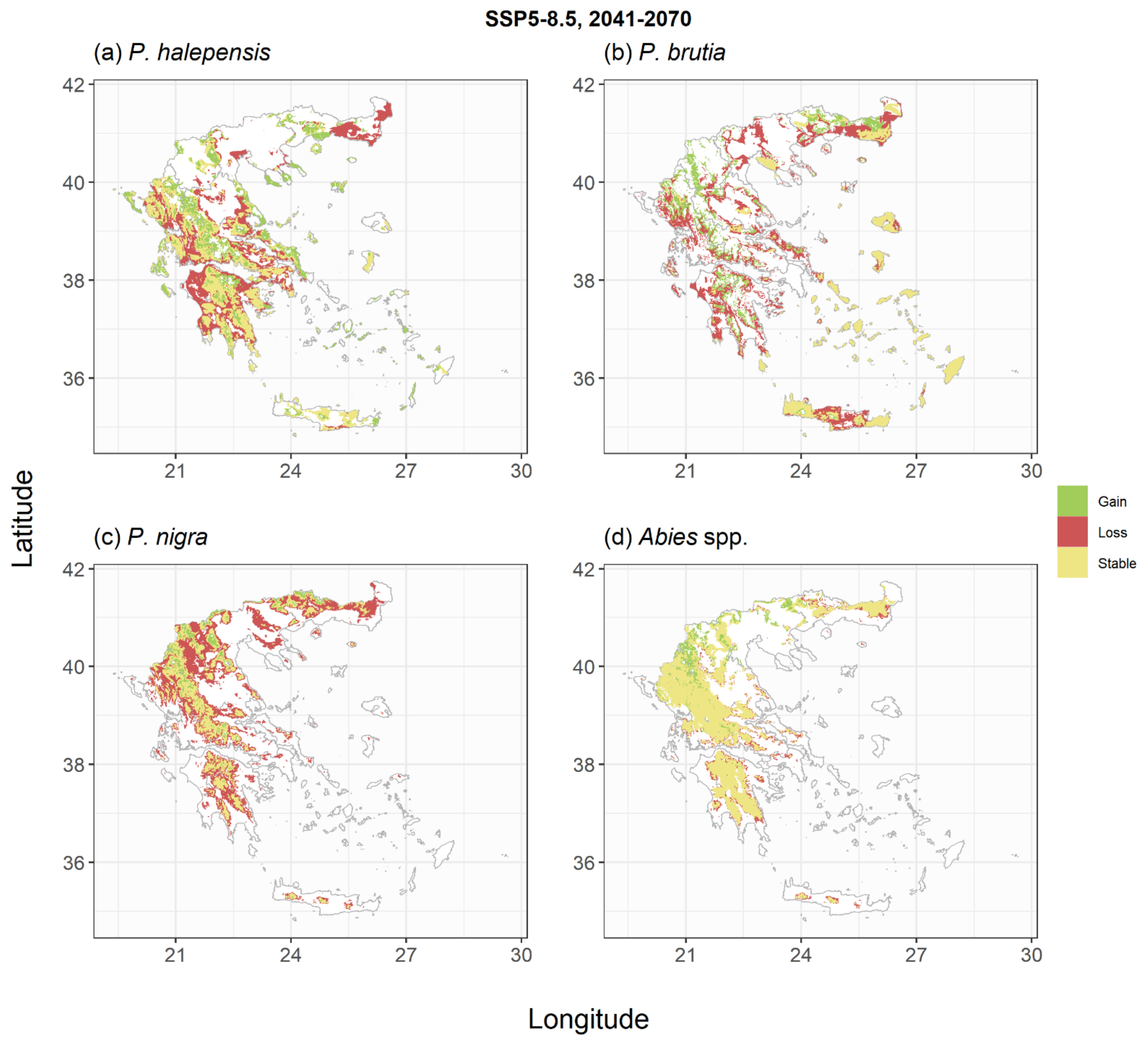
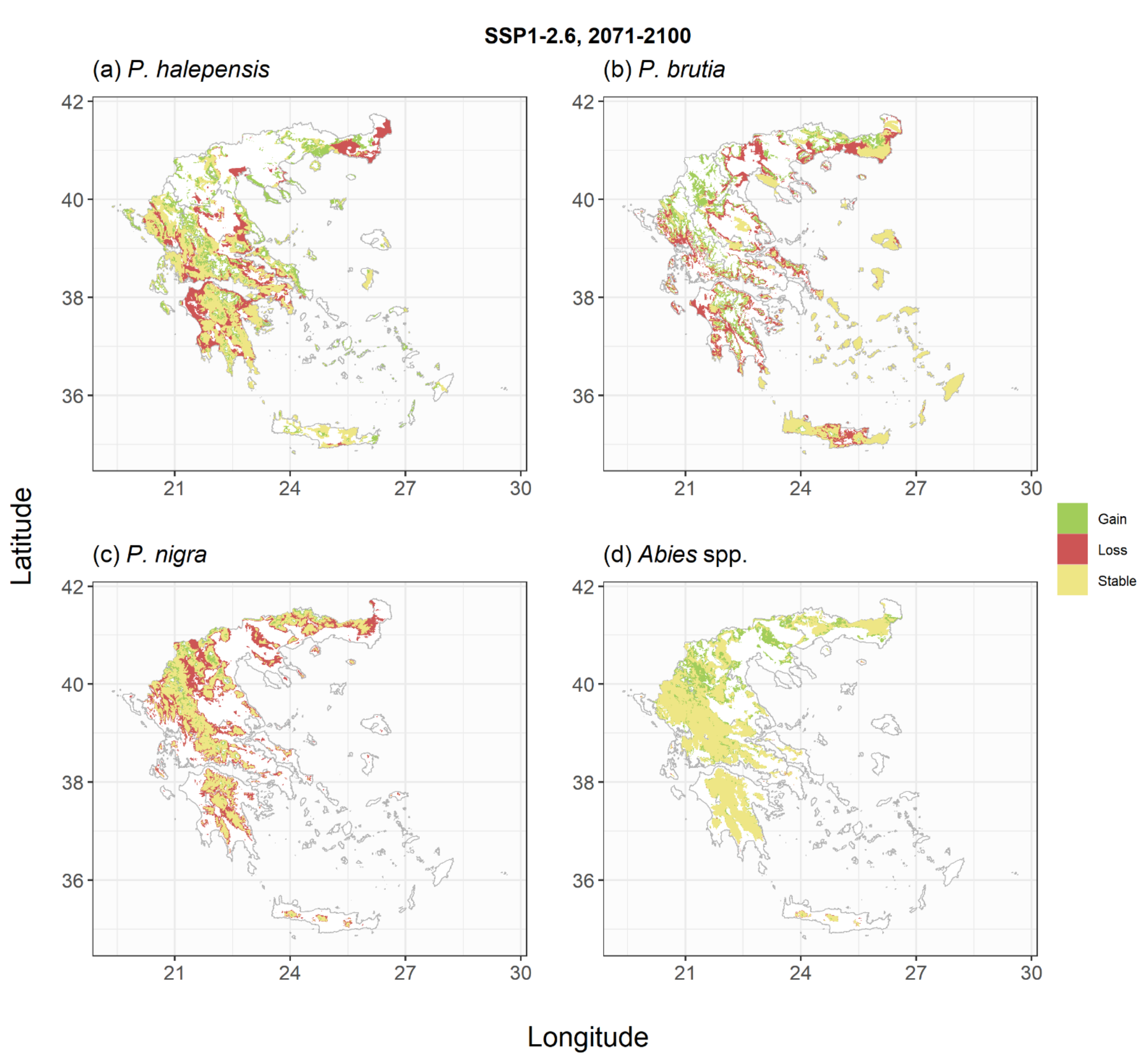
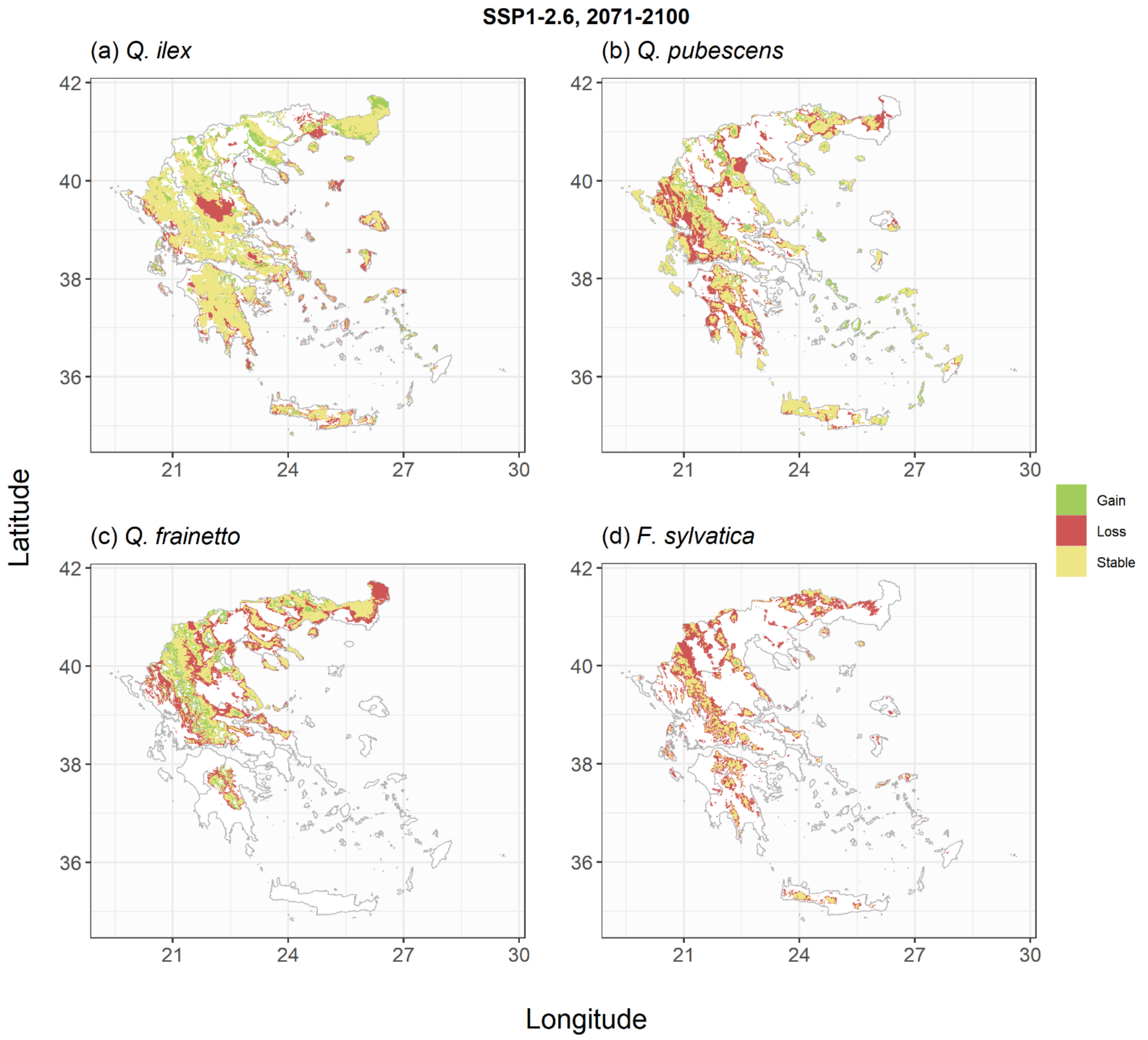
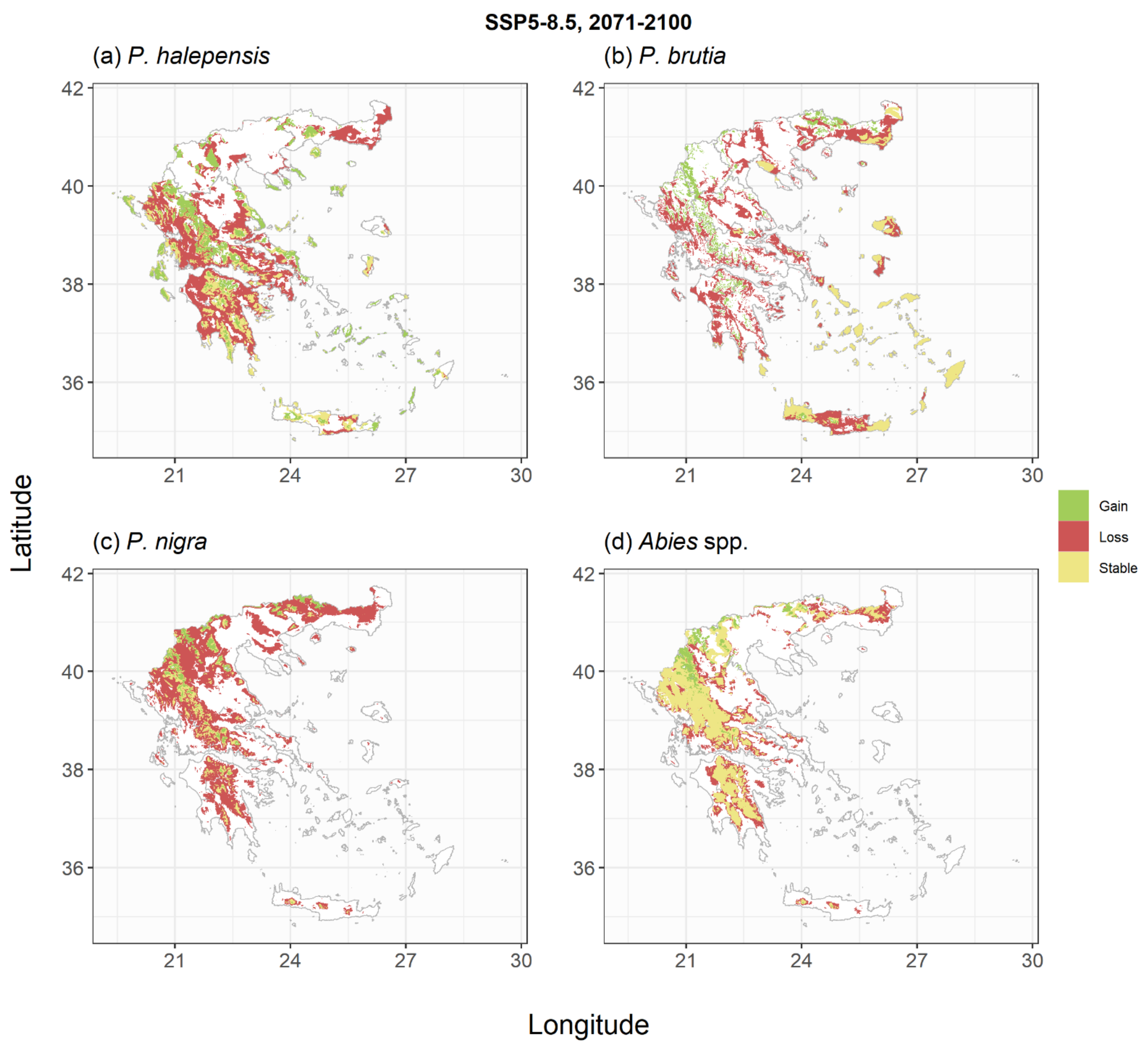
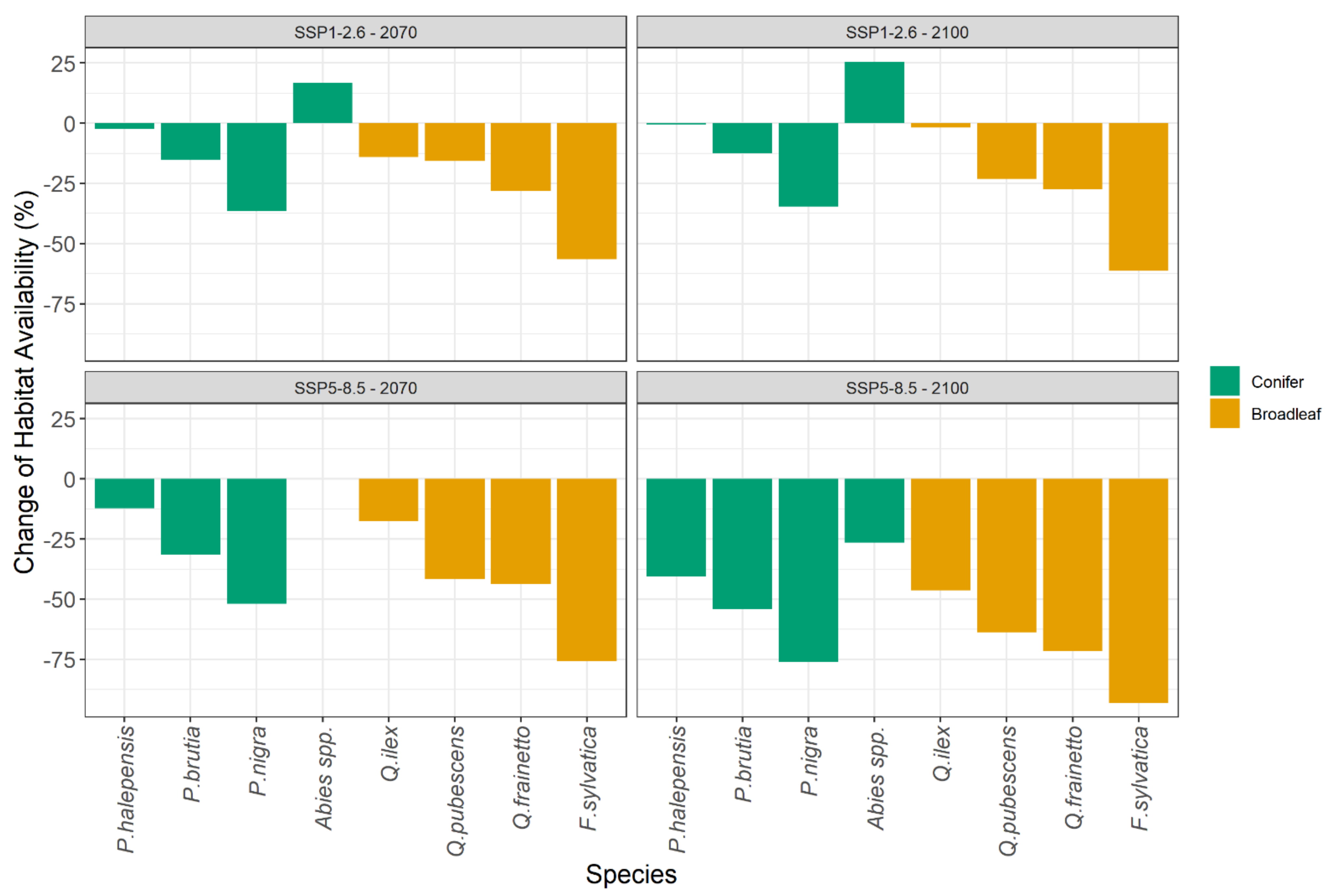
2.2. Species Habitat Suitability under Climate Change
3. Discussion
4. Materials and Methods
4.1. Study Species
4.2. Species Presence Database
4.3. Climate and Soil Data
4.4. Species Distribution Modelling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A








References
- FAO. Global Forest Resources Assessment (Desk References); FAO: Rome, Italy, 2015; ISBN 978-92-5-108826-5. [Google Scholar]
- Grammatikopoulou, I.; Vačkářová, D. The Value of Forest Ecosystem Services: A Meta-Analysis at the European Scale and Application to National Ecosystem Accounting. Ecosyst. Serv. 2021, 48, 101262. [Google Scholar] [CrossRef]
- Canadell, J.G.; Raupach, M.R. Managing Forests for Climate Change Mitigation. Science 2008, 320, 1456–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brienen, R.J.W.; Phillips, O.L.; Feldpausch, T.R.; Gloor, E.; Baker, T.R.; Lloyd, J.; Lopez-Gonzalez, G.; Monteagudo-Mendoza, A.; Malhi, Y.; Lewis, S.L.; et al. Long-Term Decline of the Amazon Carbon Sink. Nature 2015, 519, 344–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boisvenue, C.; Running, S.W. Impacts of Climate Change on Natural Forest Productivity—Evidence since the Middle of the 20th Century. Glob. Chang. Biol. 2006, 12, 862–882. [Google Scholar] [CrossRef]
- Medlyn, B.E.; Duursma, R.A.; Zeppel, M.J.B. Forest Productivity under Climate Change: A Checklist for Evaluating Model Studies. WIREs Clim. Chang. 2011, 2, 332–355. [Google Scholar] [CrossRef]
- Hickler, T.; Vohland, K.; Feehan, J.; Miller, P.A.; Smith, B.; Costa, L.; Giesecke, T.; Fronzek, S.; Carter, T.R.; Cramer, W.; et al. Projecting the Future Distribution of European Potential Natural Vegetation Zones with a Generalized, Tree Species-Based Dynamic Vegetation Model: Future Changes in European Vegetation Zones. Glob. Ecol. Biogeogr. 2012, 21, 50–63. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavergne, S.; Roquet, C.; Boulangeat, I.; Lafourcade, B.; Araujo, M.B. Consequences of Climate Change on the Tree of Life in Europe. Nature 2011, 470, 531–534. [Google Scholar] [CrossRef]
- Ruiz-Labourdette, D.; Schmitz, M.F.; Pineda, F.D. Changes in Tree Species Composition in Mediterranean Mountains under Climate Change: Indicators for Conservation Planning. Ecol. Indic. 2013, 24, 310–323. [Google Scholar] [CrossRef]
- Santini, M.; Collalti, A.; Valentini, R. Climate Change Impacts on Vegetation Andwater Cycle in the Euro-Mediterranean Region, Studied by a Likelihoodapproach. Reg. Environ. Chang. 2014, 14, 1405–1418. [Google Scholar] [CrossRef]
- Fyllas, N.M.; Phillips, O.L.; Kunin, W.E.; Matsinos, Y.G.; Troumbis, A.I. Development and Parameterization of a General Forest Gap Dynamics Simulator for the North-Eastern Mediterranean Basin (GREek FOrest Species). Ecol. Model. 2007, 204, 439–456. [Google Scholar] [CrossRef]
- Fyllas, N.M.; Christopoulou, A.; Galanidis, A.; Michelaki, C.Z.; Giannakopoulos, C.; Dimitrakopoulos, P.G.; Arianoutsou, M.; Gloor, M. Predicting Species Dominance Shifts across Elevation Gradients in Mountain Forests in Greece under a Warmer and Drier Climate. Reg. Environ. Chang. 2017, 17, 1165–1177. [Google Scholar] [CrossRef]
- Fyllas, N.M.; Troumbis, A.Y. Simulating Vegetation Shifts in North-Eastern Mediterranean Mountain Forests under Climatic Change Scenarios. Glob. Ecol. Biogeogr. 2009, 18, 64–77. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models: With Applications in R; Ecology, Biodiversity and Conservation; Cambridge University Press: Cambridge, MA, USA, 2017; ISBN 978-0-521-76513-8. [Google Scholar]
- Hutchinson, G.E. Concluding Remarks. Cold Spring Harb. Symp. Quant. Biol. 1957, 22, 415–427. [Google Scholar] [CrossRef]
- Araújo, M.B.; Guisan, A. Five (or so) Challenges for Species Distribution Modelling. J. Biogeogr. 2006, 33, 1677–1688. [Google Scholar] [CrossRef]
- Clark, J.S.; Gelfand, A.E.; Woodall, C.W.; Zhu, K. More than the Sum of the Parts: Forest Climate Response from Joint Species Distribution Models. Ecol. Appl. 2014, 24, 990–999. [Google Scholar] [CrossRef]
- Zimmermann, N.E.; Jandl, R.; Hanewinkel, M.; Kunstler, G.; Kölling, C.; Gasparini, P.; Breznikar, A.; Meier, E.S.; Normand, S.; Ulmer, U.; et al. Potential Future Ranges of Tree Species in the Alps. In Management Strategies to Adapt Alpine Space Forests to Climate Change Risks; InTech: Houston, TX, USA, 2013. [Google Scholar] [CrossRef] [Green Version]
- Noce, S.; Collalti, A.; Santini, M. Likelihood of Changes in Forest Species Suitability, Distribution, and Diversity under Future Climate: The Case of Southern Europe. Ecol. Evol. 2017, 7, 9358–9375. [Google Scholar] [CrossRef] [PubMed]
- Júnior, P.D.M.; Nóbrega, C.C. Evaluating Collinearity Effects on Species Distribution Models: An Approach Based on Virtual Species Simulation. PLoS ONE 2018, 13, e0202403. [Google Scholar] [CrossRef]
- Loehle, C.; LeBlanc, D. Model-Based Assessments of Climate Change Effects on Forests: A Critical Review. Ecol. Model. 1996, 90, 1–31. [Google Scholar] [CrossRef]
- Huey, R.B.; Kearney, M.R.; Krockenberger, A.; Holtum, J.A.M.; Jess, M.; Williams, S.E. Predicting Organismal Vulnerability to Climate Warming: Roles of Behaviour, Physiology and Adaptation. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1665–1679. [Google Scholar] [CrossRef] [Green Version]
- Evans, T.; Diamond, S.; Kelly, M. Mechanistic Species Distribution Modelling as a Link between Physiology and Conservation. Conserv. Physiol. 2015, 3, cov056. [Google Scholar] [CrossRef] [Green Version]
- Wiens, J.A.; Stralberg, D.; Jongsomjit, D.; Howell, C.A.; Snyder, M.A. Niches, Models, and Climate Change: Assessing the Assumptions and Uncertainties. Proc. Natl. Acad. Sci. USA 2009, 106, 19729–19736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benito Garzón, M.; Robson, T.M.; Hampe, A. ΔTraitSDMs: Species Distribution Models That Account for Local Adaptation and Phenotypic Plasticity. New Phytol. 2019, 222, 1757–1765. [Google Scholar] [CrossRef]
- Keenan, T.; Maria Serra, J.; Lloret, F.; Ninyerola, M.; Sabate, S. Predicting the Future of Forests in the Mediterranean under Climate Change, with Niche- and Process-Based Models: CO2 Matters! Glob. Chang. Biol. 2011, 17, 565–579. [Google Scholar] [CrossRef]
- Soberón, J.; Peterson, A.T. Interpretation of Models of Fundamental Ecological Niches and Species’ Distributional Areas. Biodivers. Inform. 2005, 2. [Google Scholar] [CrossRef] [Green Version]
- San-Miguel-Ayanz, J.; de Rigo, D.; Caudullo, G.; Durrant, T.; Mauri, A.; Tinner, W.; Ballian, D.; Beck, P.; Birks, H.; Eaton, E.; et al. European Atlas of Forest Tree Species; Publications Office of the European Union: Luxembourg, 2016; ISBN 978-92-79-36740-3. [Google Scholar]
- Mauri, A.; Strona, G.; San-Miguel-Ayanz, J. EU-Forest, a High-Resolution Tree Occurrence Dataset for Europe. Sci Data 2017, 4, 160123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A Statistical Explanation of MaxEnt for Ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample Selection Bias and Presence-Only Distribution Models: Implications for Background and Pseudo-Absence Data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Pecchi, M.; Marchi, M.; Burton, V.; Giannetti, F.; Moriondo, M.; Bernetti, I.; Bindi, M.; Chirici, G. Species Distribution Modelling to Support Forest Management. A Literature Review. Ecol. Model. 2019, 411, 108817. [Google Scholar] [CrossRef]
- Mudereri, B.T.; Mukanga, C.; Mupfiga, E.T.; Gwatirisa, C.; Kimathi, E.; Chitata, T. Analysis of Potentially Suitable Habitat within Migration Connections of an Intra-African Migrant-the Blue Swallow (Hirundo Atrocaerulea). Ecol. Inform. 2020, 57, 101082. [Google Scholar] [CrossRef]
- Tessarolo, G.; Lobo, J.M.; Rangel, T.F.; Hortal, J. High Uncertainty in the Effects of Data Characteristics on the Performance of Species Distribution Models. Ecol. Indic. 2021, 121, 107147. [Google Scholar] [CrossRef]
- Mauri, A.; Girardello, M.; Strona, G.; Beck, P.S.A.; Forzieri, G.; Caudullo, G.; Manca, F.; Cescatti, A. EU-Trees4F, a Dataset on the Future Distribution of European Tree Species. Sci. Data 2022, 9, 37. [Google Scholar] [CrossRef] [PubMed]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How Much Does Climate Change Threaten European Forest Tree Species Distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Benito Garzón, M.; Sánchez de Dios, R.; Sainz Ollero, H. Effects of Climate Change on the Distribution of Iberian Tree Species. Appl. Veg. Sci. 2008, 11, 169–178. [Google Scholar] [CrossRef]
- Ruiz-Labourdette, D.; Nogués-Bravo, D.; Ollero, H.S.; Schmitz, M.F.; Pineda, F.D. Forest Composition in Mediterranean Mountains Is Projected to Shift along the Entire Elevational Gradient under Climate Change. J. Biogeogr. 2012, 39, 162–176. [Google Scholar] [CrossRef]
- Buras, A.; Menzel, A. Projecting Tree Species Composition Changes of European Forests for 2061–2090 Under RCP 4.5 and RCP 8.5 Scenarios. Front. Plant Sci. 2019, 9. [Google Scholar]
- Takolander, A.; Hickler, T.; Meller, L.; Cabeza, M. Comparing Future Shifts in Tree Species Distributions across Europe Projected by Statistical and Dynamic Process-Based Models. Reg. Environ. Chang. 2019, 19, 251–266. [Google Scholar] [CrossRef] [Green Version]
- Pecchi, M.; Marchi, M.; Moriondo, M.; Forzieri, G.; Ammoniaci, M.; Bernetti, I.; Bindi, M.; Chirici, G. Potential Impact of Climate Change on the Forest Coverage and the Spatial Distribution of 19 Key Forest Tree Species in Italy under RCP4.5 IPCC Trajectory for 2050s. Forests 2020, 11, 934. [Google Scholar] [CrossRef]
- Arslan, E.; Örücü, Ö.K. Present and future potential distribution of the Pinus nigra Arnold. and Pinus sylvestris L. using Maxent models. Int. J. Ecosyst. Ecol. Sci. 2019, 9, 787–798. [Google Scholar] [CrossRef]
- George, J.-P.; Schueler, S.; Karanitsch-Ackerl, S.; Mayer, K.; Klumpp, R.T.; Grabner, M. Inter- and Intra-Specific Variation in Drought Sensitivity in Abies Spec. and Its Relation to Wood Density and Growth Traits. Agric. For. Meteorol. 2015, 214–215, 430–443. [Google Scholar] [CrossRef] [Green Version]
- Attorre, F.; Alfò, M.; De Sanctis, M.; Francesconi, F.; Valenti, R.; Vitale, M.; Bruno, F. Evaluating the Effects of Climate Change on Tree Species Abundance and Distribution in the Italian Peninsula. Appl. Veg. Sci. 2011, 14, 242–255. [Google Scholar] [CrossRef]
- Mauri, A.; Leo, M.D.; de Rigo, D.; Caudullo, G. Pinus halepensis and Pinus brutia. European Atlas of Forest Tree Species; Publications Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Klein, T.; Cohen, S.; Yakir, D. Hydraulic Adjustments Underlying Drought Resistance of Pinus Halepensis. Tree Physiol. 2011, 31, 637–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pausas, J.G. The Effect of Landscape Pattern on Mediterranean Vegetation Dynamics: A Modelling Approach Using Functional Types. J. Veg. Sci. 2003, 14, 365–374. [Google Scholar] [CrossRef]
- Kazanis, D.; Arianoutsou, M. Long-Term Post-Fire Vegetation Dynamics in Pinus Halepensis Forests of Central Greece: A Functional Group Approach. Plant Ecol. 2004, 171, 101–121. [Google Scholar] [CrossRef]
- Papadopoulos, A.; Serre-Bachet, F.; Tessier, L. Tree Ring to Climate Relationships of Aleppo Pine (Pinus halepensis Mill.) in Greece. Ecol. Mediterr. 2001, 27, 89–98. [Google Scholar] [CrossRef]
- Sarris, D.; Christodoulakis, D.; Körner, C. Recent Decline in Precipitation and Tree Growth in the Eastern Mediterranean. Glob. Chang. Biol. 2007, 13, 1187–1200. [Google Scholar] [CrossRef]
- Fyllas, N.M.; Christopoulou, A.; Galanidis, A.; Michelaki, C.Z.; Dimitrakopoulos, P.G.; Fulé, P.Z.; Arianoutsou, M. Tree Growth-Climate Relationships in a Forest-Plot Network on Mediterranean Mountains. Sci. Total Environ. 2017, 598, 393–403. [Google Scholar] [CrossRef]
- Sarris, D.; Christodoulakis, D.; Körner, C. Impact of Recent Climatic Change on Growth of Low Elevation Eastern Mediterranean Forest Trees. Clim. Chang. 2011, 106, 203–223. [Google Scholar] [CrossRef]
- Christopoulou, A.; Sazeides, C.I.; Fyllas, N.M. Size-Mediated Effects of Climate on Tree Growth and Mortality in Mediterranean Brutia Pine Forests. Sci. Total Environ. 2022, 812, 151463. [Google Scholar] [CrossRef]
- Sabaté, S.; Gracia, C.A.; Sánchez, A. Likely Effects of Climate Change on Growth of Quercus Ilex, Pinus Halepensis, Pinus Pinaster, Pinus Sylvestris and Fagus Sylvatica Forests in the Mediterranean Region. For. Ecol. Manag. 2002, 162, 23–37. [Google Scholar] [CrossRef]
- Dimitrakopoulos, A.P. A Statistical Classification of Mediterranean Species Based on Their Flammability Components. Int. J. Wildland Fire 2001, 10, 113–118. [Google Scholar] [CrossRef]
- Moriondo, M.; Good, P.; Durao, R.; Bindi, M.; Giannakopoulos, C.; Corte-Real, J. Potential Impact of Climate Change on Fire Risk in the Mediterranean Area. Clim. Res. 2006, 31, 85–95. [Google Scholar] [CrossRef]
- Enescu, C.; de Rigo, D.; Caudullo, G.; Durrant, T. Pinus nigra in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; Publications Office of the European Union: Luxembourg, 2016; ISBN 978-92-79-36740-3. [Google Scholar]
- Fyllas, N.M.; Dimitrakopoulos, P.G.; Troumbis, A.Y. Regeneration Dynamics of a Mixed Mediterranean Pine Forest in the Absence of Fire. For. Ecol. Manag. 2008, 256, 1552–1559. [Google Scholar] [CrossRef]
- Calama, R.; Manso, R.; Lucas-Borja, M.E.; Espelta, J.M.; Piqué, M.; Bravo, F.; del Peso, C.; Pardos, M. Natural Regeneration in Iberian Pines: A Review of Dynamic Processes and Proposals for Management. For. Syst. 2017, 26, eR02S. [Google Scholar] [CrossRef]
- Camarero, J.J.; Manzanedo, R.D.; Sanchez-Salguero, R.; Navarro-Cerrillo, R.M. Growth Response to Climate and Drought Change along an Aridity Gradient in the Southernmost Pinus nigra Relict Forests. Ann. For. Sci. 2013, 70, 769–780. [Google Scholar] [CrossRef]
- Linares, J.C.; Tíscar, P.A. Climate Change Impacts and Vulnerability of the Southern Populations of Pinus nigra subsp. salzmannii. Tree Physiol 2010, 30, 795–806. [Google Scholar] [CrossRef] [Green Version]
- Savi, T.; Casolo, V.; Dal Borgo, A.; Rosner, S.; Torboli, V.; Stenni, B.; Bertoncin, P.; Martellos, S.; Pallavicini, A.; Nardini, A. Drought-Induced Dieback of Pinus nigra: A Tale of Hydraulic Failure and Carbon Starvation. Conserv. Physiol. 2019, 7, coz012. [Google Scholar] [CrossRef]
- Deligöz, A.; Cankara, F.G. Differences in Physiological and Biochemical Responses to Summer Drought of Pinus nigra subsp. pallasiana and Pinus brutia in a Natural Mixed Stand. J. For. Res. 2020, 31, 1479–1487. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Navarro-Cerrillo, R.M.; Camarero, J.J.; Fernández-Cancio, Á. Selective Drought-Induced Decline of Pine Species in Southeastern Spain. Clim. Chang. 2012, 113, 767–785. [Google Scholar] [CrossRef]
- Móricz, N.; Garamszegi, B.; Rasztovits, E.; Bidló, A.; Horváth, A.; Jagicza, A.; Illés, G.; Vekerdy, Z.; Somogyi, Z.; Gálos, B. Recent Drought-Induced Vitality Decline of Black Pine (Pinus nigra Arn.) in South-West Hungary—Is This Drought-Resistant Species under Threat by Climate Change? Forests 2018, 9, 414. [Google Scholar] [CrossRef] [Green Version]
- González de Andrés, E.; Camarero, J.J. Disentangling Mechanisms of Drought-Induced Dieback in Pinus nigra Arn. from Growth and Wood Isotope Patterns. Forests 2020, 11, 1339. [Google Scholar] [CrossRef]
- Martin-Benito, D.; Kint, V.; del Río, M.; Muys, B.; Cañellas, I. Growth Responses of West-Mediterranean Pinus nigra to Climate Change Are Modulated by Competition and Productivity: Past Trends and Future Perspectives. For. Ecol. Manag. 2011, 262, 1030–1040. [Google Scholar] [CrossRef]
- Candel-Pérez, D.; Lucas-Borja, M.E.; García-Cervigón, A.I.; Tíscar, P.A.; Andivia, E.; Bose, A.K.; Sánchez-Salguero, R.; Camarero, J.J.; Linares, J.C. Forest Structure Drives the Expected Growth of Pinus nigra along Its Latitudinal Gradient under Warming Climate. For. Ecol. Manag. 2022, 505, 119818. [Google Scholar] [CrossRef]
- Christopoulou, A.; Fyllas, N.M.; Andriopoulos, P.; Koutsias, N.; Dimitrakopoulos, P.G.; Arianoutsou, M. Post-Fire Regeneration Patterns of Pinus nigra in a Recently Burned Area in Mount Taygetos, Southern Greece: The Role of Unburned Forest Patches. For. Ecol. Manag. 2014, 327, 148–156. [Google Scholar] [CrossRef]
- Guehl, J.M.; Aussenac, G.; Bouachrine, J.; Zimmermann, R.; Pennes, J.M.; Ferhi, A.; Grieu, P. Sensitivity of Leaf Gas Exchange to Atmospheric Drought, Soil Drought, and Water-Use Efficiency in Some Mediterranean Abies Species. Can. J. For. Res. 1991, 21, 1507–1515. [Google Scholar] [CrossRef]
- Aussenac, G. Ecology and Ecophysiology of Circum-Mediterranean Firs in the Context of Climate Change. Ann. For. Sci. 2002, 59, 823–832. [Google Scholar] [CrossRef]
- Markalas, S. Site and Stand Factors Related to Mortality Rate in a Fir Forest after a Combined Incidence of Drought and Insect Attack. For. Ecol. Manag. 1992, 47, 367–374. [Google Scholar] [CrossRef]
- Heliotis, F.D.; Karandinos, M.G.; Whiton, J.C. Air Pollution and the Decline of the Fir Forest in Parnis National Park, near Athens, Greece. Environ. Pollut. 1988, 54, 29–40. [Google Scholar] [CrossRef]
- Raftoyannis, Y.; Spanos, I.; Radoglou, K. The Decline of Greek Fir (Abies cephalonica Loudon): Relationships with Root Condition. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2008, 142, 386–390. [Google Scholar] [CrossRef]
- Raftoyannis, Y.; Spanos, I. Regeneration of Abies cephalonica Loudon after a Large Fire in Central Greece. South-East Eur. For. SEEFOR 2015, 6, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, A. Tree-Ring Patterns and Climate Response of Mediterranean Fir Populations in Central Greece. Dendrochronologia 2016, 40, 17–25. [Google Scholar] [CrossRef]
- Koutavas, A. CO2 Fertilization and Enhanced Drought Resistance in Greek Firs from Cephalonia Island, Greece. Glob. Chang. Biol. 2013, 19, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Daskalakou, E.N.; Koutsovoulou, K.; Ioannidis, K.; Koulelis, P.P.; Ganatsas, P.; Thanos, C.A. Masting and Regeneration Dynamics of Abies cephalonica, the Greek Endemic Silver Fir. Seed Sci. Res. 2019, 29, 227–237. [Google Scholar] [CrossRef]
- Christopoulou, A.; Kazanis, D.; Fyllas, N.M.; Arianoutsou, M. Post-Fire Recovery of Abies cephalonica Forest Communities: The Case of Mt Parnitha National Park, Attica, Greece. IForest 2018, 11, 757–764. [Google Scholar] [CrossRef] [Green Version]
- Bantis, F.; Radoglou, K.; Brüggemann, W. Differential Ecophysiological Responses to Seasonal Drought of Three Co-Existing Oak Species in Northern Greece. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2019, 153, 378–384. [Google Scholar] [CrossRef]
- Baquedano, F.J.; Castillo, F.J. Drought Tolerance in the Mediterranean Species Quercus coccifera, Quercus ilex, Pinus halepensis, and Juniperus phoenicea. Photosynthetica 2007, 45, 229. [Google Scholar] [CrossRef]
- de Rigo, D.; Caudullo, G. Quercus ilex in Europe: Distribution, Habitat, Usage and Threats; Publications Office of the European Union: Luxembourg, 2016; ISBN 978-92-79-36740-3. [Google Scholar]
- Früchtenicht, E.; Neumann, L.; Klein, N.; Bonal, D.; Brüggemann, W. Response of Quercus robur and Two Potential Climate Change Winners—Quercus Pubescens and Quercus Ilex—To Two Years Summer Drought in a Semi-Controlled Competition Study: I—Tree Water Status. Environ. Exp. Bot. 2018, 152, 107–117. [Google Scholar] [CrossRef]
- Bantis, F.; Graap, J.; Früchtenicht, E.; Bussotti, F.; Radoglou, K.; Brüggemann, W. Field Performances of Mediterranean Oaks in Replicate Common Gardens for Future Reforestation under Climate Change in Central and Southern Europe: First Results from a Four-Year Study. Forests 2021, 12, 678. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Colangelo, M.; Matías, L.; Ripullone, F.; Camarero, J.J. Shifts in Growth Responses to Climate and Exceeded Drought-Vulnerability Thresholds Characterize Dieback in Two Mediterranean Deciduous Oaks. Forests 2020, 11, 714. [Google Scholar] [CrossRef]
- Colangelo, M.; Camarero, J.J.; Borghetti, M.; Gazol, A.; Gentilesca, T.; Ripullone, F. Size Matters a Lot: Drought-Affected Italian Oaks Are Smaller and Show Lower Growth Prior to Tree Death. Front. Plant Sci. 2017, 8, 135. [Google Scholar] [CrossRef] [Green Version]
- Colangelo, M.; Camarero, J.J.; Borghetti, M.; Gentilesca, T.; Oliva, J.; Redondo, M.-A.; Ripullone, F. Drought and Phytophthora Are Associated With the Decline of Oak Species in Southern Italy. Front. Plant Sci. 2018, 9, 1595. [Google Scholar] [CrossRef] [PubMed]
- Geßler, A.; Keitel, C.; Kreuzwieser, J.; Matyssek, R.; Seiler, W.; Rennenberg, H. Potential Risks for European Beech (Fagus sylvatica L.) in a Changing Climate. Trees 2007, 21, 1–11. [Google Scholar] [CrossRef]
- Piovesan, G.; Biondi, F.; Filippo, A.D.; Alessandrini, A.; Maugeri, M. Drought-Driven Growth Reduction in Old Beech (Fagus sylvatica L.) Forests of the Central Apennines, Italy. Glob. Chang. Biol. 2008, 14, 1265–1281. [Google Scholar] [CrossRef]
- Scharnweber, T.; Manthey, M.; Criegee, C.; Bauwe, A.; Schröder, C.; Wilmking, M. Drought Matters—Declining Precipitation Influences Growth of Fagus sylvatica L. and Quercus robur L. in North-Eastern Germany. For. Ecol. Manag. 2011, 262, 947–961. [Google Scholar] [CrossRef]
- Tegel, W.; Seim, A.; Hakelberg, D.; Hoffmann, S.; Panev, M.; Westphal, T.; Büntgen, U. A Recent Growth Increase of European Beech (Fagus sylvatica L.) at Its Mediterranean Distribution Limit Contradicts Drought Stress. Eur. J. For. Res. 2014, 133, 61–71. [Google Scholar] [CrossRef]
- Leuschner, C. Drought Response of European Beech (Fagus sylvatica L.)—A Review. Perspect. Plant Ecol. Evol. Syst. 2020, 47, 125576. [Google Scholar] [CrossRef]
- Jump, A.S.; Hunt, J.M.; Peñuelas, J. Rapid Climate Change-Related Growth Decline at the Southern Range Edge of Fagus Sylvatica. Glob. Chang. Biol. 2006, 12, 2163–2174. [Google Scholar] [CrossRef] [Green Version]
- Martinez del Castillo, E.; Zang, C.S.; Buras, A.; Hacket-Pain, A.; Esper, J.; Serrano-Notivoli, R.; Hartl, C.; Weigel, R.; Klesse, S.; Resco de Dios, V.; et al. Climate-Change-Driven Growth Decline of European Beech Forests. Commun. Biol. 2022, 5, 1–9. [Google Scholar] [CrossRef]
- Kramer, K.; Degen, B.; Buschbom, J.; Hickler, T.; Thuiller, W.; Sykes, M.T.; de Winter, W. Modelling Exploration of the Future of European Beech (Fagus sylvatica L.) under Climate Change—Range, Abundance, Genetic Diversity and Adaptive Response. For. Ecol. Manag. 2010, 259, 2213–2222. [Google Scholar] [CrossRef]
- Spanos, K.A.; Skouteri, A.; Gaitanis, D.; Petrakis, P.V.; Meliadis, I.; Michopoulos, P.; Solomou, A.; Koulelis, P.; Avramidou, E.V. Forests of Greece, Their Multiple Functions and Uses, Sustainable Management and Biodiversity Conservation in the Face of Climate Change. Open J. Ecol. 2021, 11, 374–406. [Google Scholar] [CrossRef]
- Chatziphilippidis, G.; Spyroglou, G. Modelling the Growth of Quercus frainetto in Greece. In Sustainable Forest Management; Hasenauer, H., Ed.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 373–395. [Google Scholar]
- Dimopoulos, P.; Bergmeier, E.; Eleftheriadou, E.; Theodoropoulos, K.; Gerasimidis, A.; Tsiafouli, M. Identification and Interpretation Guide for the Forest Habitats of Greece; University of Western Greece Publication: Agrinio, Greece, 2012. [Google Scholar]
- Frankis, M. 367. Pinus brutia: Pinaceae. Curtis’s Bot. Mag. 1999, 16, 173–184. [Google Scholar] [CrossRef]
- Chambel, M.R.; Climent, J.; Pichot, C.; Ducci, F. Mediterranean Pines (Pinus halepensis Mill. and brutia Ten.). In Forest Tree Breeding in Europe: Current State-of-the-Art and Perspectives; Pâques, L.E., Ed.; Managing Forest Ecosystems; Springer: Dordrecht, The Netherlands, 2013; pp. 229–265. ISBN 978-94-007-6146-9. [Google Scholar]
- Bergmeier, E. Plant Communities and Habitat Differentiation in the Mediterranean Coniferous Woodlands of Mt. Parnon (Greece). Folia Geobot. 2002, 37, 309–331. [Google Scholar] [CrossRef]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Climatologies at High Resolution for the Earth’s Land Surface Areas. Sci. Data 2017, 4, 170122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitrakos, K. A Theory for Mediterranean Plant Life [Evergreen Sclerophyllous Shrubs, Climatic Stresses, Mediterranean Climate]. Acta Oecologica-Oecologia Plant. (Fr.) 1980, 1, 245–252. [Google Scholar]
- Terradas, J.; Savé, R. The Influence of Summer and Winter Stress and Water Relationships on the Distribution of Quercus ilex L. Vegetatio 1992, 99/100, 137–145. [Google Scholar] [CrossRef]
- Paulsen, J.; Körner, C. A Climate-Based Model to Predict Potential Treeline Position around the Globe. Alp Bot. 2014, 124, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Eyring, V.; Bony, S.; Meehl, G.A.; Senior, C.A.; Stevens, B.; Stouffer, R.J.; Taylor, K.E. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) Experimental Design and Organization. Geosci. Model Dev. 2016, 9, 1937–1958. [Google Scholar] [CrossRef] [Green Version]
- Dunne, J.P.; Horowitz, L.W.; Adcroft, A.J.; Ginoux, P.; Held, I.M.; John, J.G.; Krasting, J.P.; Malyshev, S.; Naik, V.; Paulot, F.; et al. The GFDL Earth System Model Version 4.1 (GFDL-ESM 4.1): Overall Coupled Model Description and Simulation Characteristics. J. Adv. Model. Earth Syst. 2020, 12, e2019MS002015. [Google Scholar] [CrossRef]
- Panagos, P.; Van Liedekerke, M.; Jones, A.; Montanarella, L. European Soil Data Centre: Response to European Policy Support and Public Data Requirements. Land Use Policy 2012, 29, 329–338. [Google Scholar] [CrossRef]
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. SpThin: An R Package for Spatial Thinning of Species Occurrence Records for Use in Ecological Niche Models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- Anderson, R.P.; Raza, A. The Effect of the Extent of the Study Region on GIS Models of Species Geographic Distributions and Estimates of Niche Evolution: Preliminary Tests with Montane Rodents (Genus Nephelomys) in Venezuela. J. Biogeogr. 2010, 37, 1378–1393. [Google Scholar] [CrossRef]
- Hof, A.R.; Allen, A.M. An Uncertain Future for the Endemic Galliformes of the Caucasus. Sci. Total Environ. 2019, 651, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt Modeling to Predict the Potential Distribution of the Endemic Plant Rosa arabica Crép in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Atzeni, L.; Cushman, S.A.; Bai, D.; Wang, J.; Chen, P.; Shi, K.; Riordan, P. Meta-Replication, Sampling Bias, and Multi-Scale Model Selection: A Case Study on Snow Leopard (Panthera uncia) in Western China. Ecol. Evol. 2020, 10, 7686–7712. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel Methods Improve Prediction of Species’ Distributions from Occurrence Data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Grimmett, L.; Whitsed, R.; Horta, A. Presence-Only Species Distribution Models Are Sensitive to Sample Prevalence: Evaluating Models Using Spatial Prediction Stability and Accuracy Metrics. Ecol. Model. 2020, 431, 109194. [Google Scholar] [CrossRef]
- Tognelli, M.F.; Roig-Juñent, S.A.; Marvaldi, A.E.; Flores, G.E.; Lobo, J.M. An Evaluation of Methods for Modelling Distribution of Patagonian Insects. Rev. Chil. Hist. Nat. 2009, 82. [Google Scholar] [CrossRef] [Green Version]
- Antúnez, P.; Suárez-Mota, M.E.; Valenzuela-Encinas, C.; Ruiz-Aquino, F. The Potential Distribution of Tree Species in Three Periods of Time under a Climate Change Scenario. Forests 2018, 9, 628. [Google Scholar] [CrossRef] [Green Version]
- Vignali, S.; Barras, A.G.; Arlettaz, R.; Braunisch, V. SDMtune: An R Package to Tune and Evaluate Species Distribution Models. Ecol. Evol. 2020, 10, 11488–11506. [Google Scholar] [CrossRef]
- Engler, R.; Guisan, A.; Rechsteiner, L. An Improved Approach for Predicting the Distribution of Rare and Endangered Species from Occurrence and Pseudo-Absence Data. J. Appl. Ecol. 2004, 41, 263–274. [Google Scholar] [CrossRef]
- Baddeley, A.; Berman, M.; Fisher, N.I.; Hardegen, A.; Milne, R.K.; Schuhmacher, D.; Shah, R.; Turner, R. Spatial Logistic Regression and Change-of-Support in Poisson Point Processes. Electron. J. Stat. 2010, 4, 1151–1201. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting Thresholds of Occurrence in the Prediction of Species Distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Variables | Pinus halepensis | Pinus brutia | Pinus nigra | Abies spp. | Quercus ilex | Quercus pubescens | Quercus frainetto | Fagus sylvatica |
|---|---|---|---|---|---|---|---|---|
| Tmax | 47.4 | 41.9 | 49.2 | 29.9 | ||||
| Tmin | 12.2 | |||||||
| Pa | 7.6 | 12.4 | ||||||
| Pdm | 15.0 | 26.5 | 8.7 | 19.8 | 24.3 | 28.4 | ||
| GDD5 | 37.2 | 35.8 | 37.9 | |||||
| GSL | 8.7 | 13.6 | 16.3 | 36.2 | ||||
| GST | 6.5 | 7.3 | 8.5 | 13.4 | ||||
| GSP | 18.8 | 9.0 | 10.2 | |||||
| parmat | 37.6 | 9.1 | 30.3 | 12.8 | 12.8 | 26.9 | 15.0 | 8.1 |
| texture | 18.5 | 46.2 | ||||||
| # occurrences (thinned) | 1181 | 38 | 2041 | 67 | 2455 | 1896 | 209 | 6634 |
| TSS | 0.80 | 0.91 | 0.58 | 0.81 | 0.78 | 0.66 | 0.77 | 0.61 |
| AUC | 0.95 | 0.98 | 0.86 | 0.94 | 0.94 | 0.90 | 0.96 | 0.86 |
| Change of Habitat Availability (%) | Pinus halepensis | Pinus brutia | Pinus nigra | Abies spp. | Quercus ilex | Quercus pubescens | Quercus frainetto | Fagus sylvatica |
|---|---|---|---|---|---|---|---|---|
| SSP1-2.6_2070 | −8 | −17 | −38 | +17 | −14 | −16 | −28 | −56 |
| SSP1-2.6_2100 | −6 | −14 | −36 | +25 | −1 | −24 | −28 | −60 |
| SSP5-8.5_2070 | −21 | −32 | −53 | 0 | −18 | −42 | −44 | −75 |
| SSP5-8.5_2100 | −45 | −54 | −77 | −27 | −47 | −64 | −72 | −93 |
| Shift in Mean Elevation (m) | Pinus halepensis | Pinus brutia | Pinus nigra | Abies spp. | Quercus ilex | Quercus pubescens | Quercus frainetto | Fagus sylvatica |
| SSP1-2.6_2070 | +139 | +175 | +257 | -3 | +95 | +143 | +233 | +285 |
| SSP1-2.6_2100 | +159 | +164 | +233 | −13 | +71 | +159 | +253 | +293 |
| SSP5-8.5_2070 | +209 | +236 | +359 | +63 | +195 | +262 | +375 | +434 |
| SSP5-8.5_2100 | +330 | +333 | +599 | +185 | +387 | +307 | +650 | +655 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fyllas, N.M.; Koufaki, T.; Sazeides, C.I.; Spyroglou, G.; Theodorou, K. Potential Impacts of Climate Change on the Habitat Suitability of the Dominant Tree Species in Greece. Plants 2022, 11, 1616. https://doi.org/10.3390/plants11121616
Fyllas NM, Koufaki T, Sazeides CI, Spyroglou G, Theodorou K. Potential Impacts of Climate Change on the Habitat Suitability of the Dominant Tree Species in Greece. Plants. 2022; 11(12):1616. https://doi.org/10.3390/plants11121616
Chicago/Turabian StyleFyllas, Nikolaos M., Theano Koufaki, Christodoulos I. Sazeides, Gavriil Spyroglou, and Konstantinos Theodorou. 2022. "Potential Impacts of Climate Change on the Habitat Suitability of the Dominant Tree Species in Greece" Plants 11, no. 12: 1616. https://doi.org/10.3390/plants11121616
APA StyleFyllas, N. M., Koufaki, T., Sazeides, C. I., Spyroglou, G., & Theodorou, K. (2022). Potential Impacts of Climate Change on the Habitat Suitability of the Dominant Tree Species in Greece. Plants, 11(12), 1616. https://doi.org/10.3390/plants11121616