
Effect of Tecoma stans (L.) Juss. ex Kunth in a Murine Model of Metabolic Syndrome

 , , ,
, , , 

Abstract
:1. Introduction
2. Results
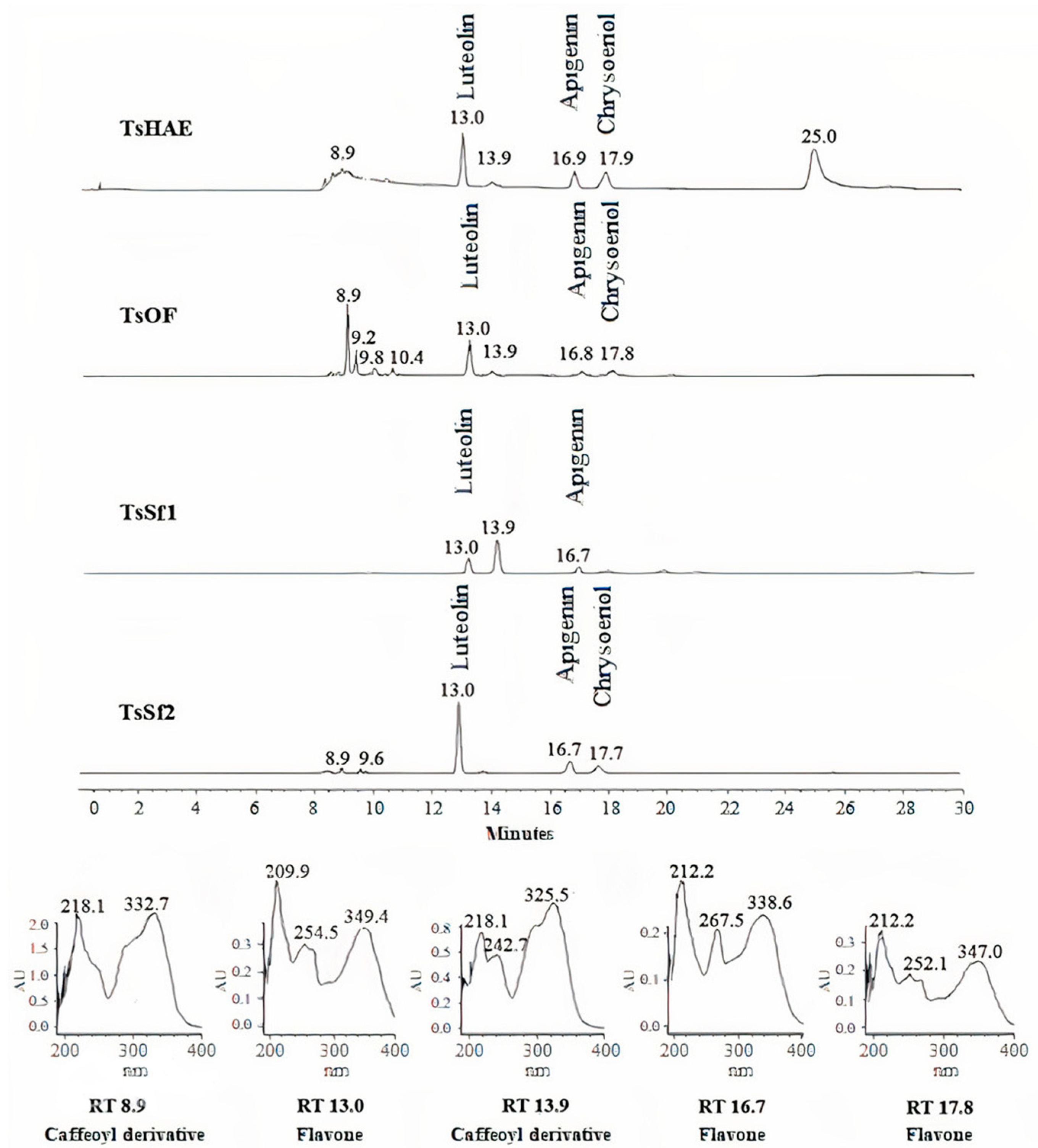
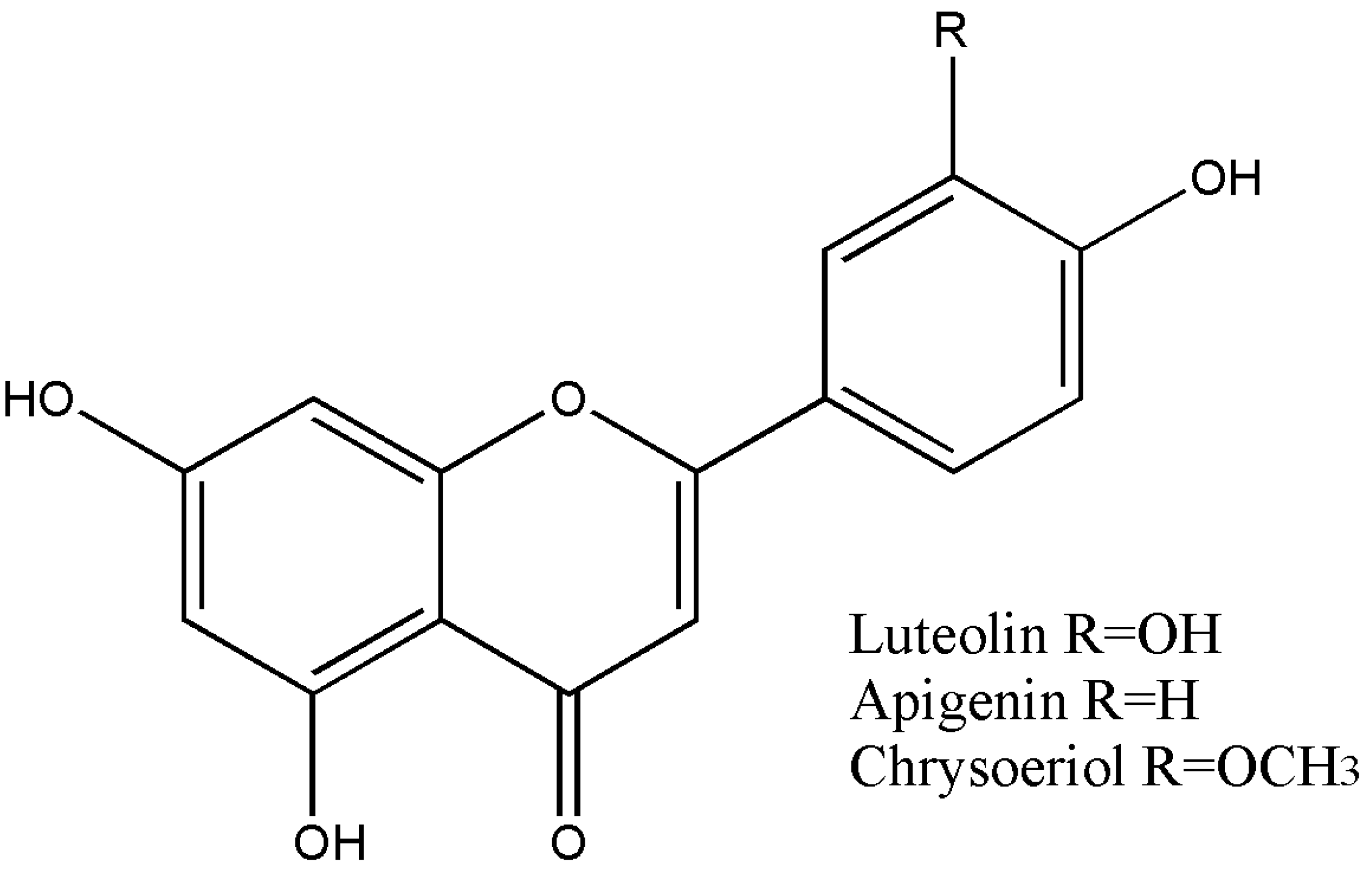
2.1. Chemical Characterization
2.2. Hypercaloric Diet Model
2.2.1. Insulin Resistance Parameters
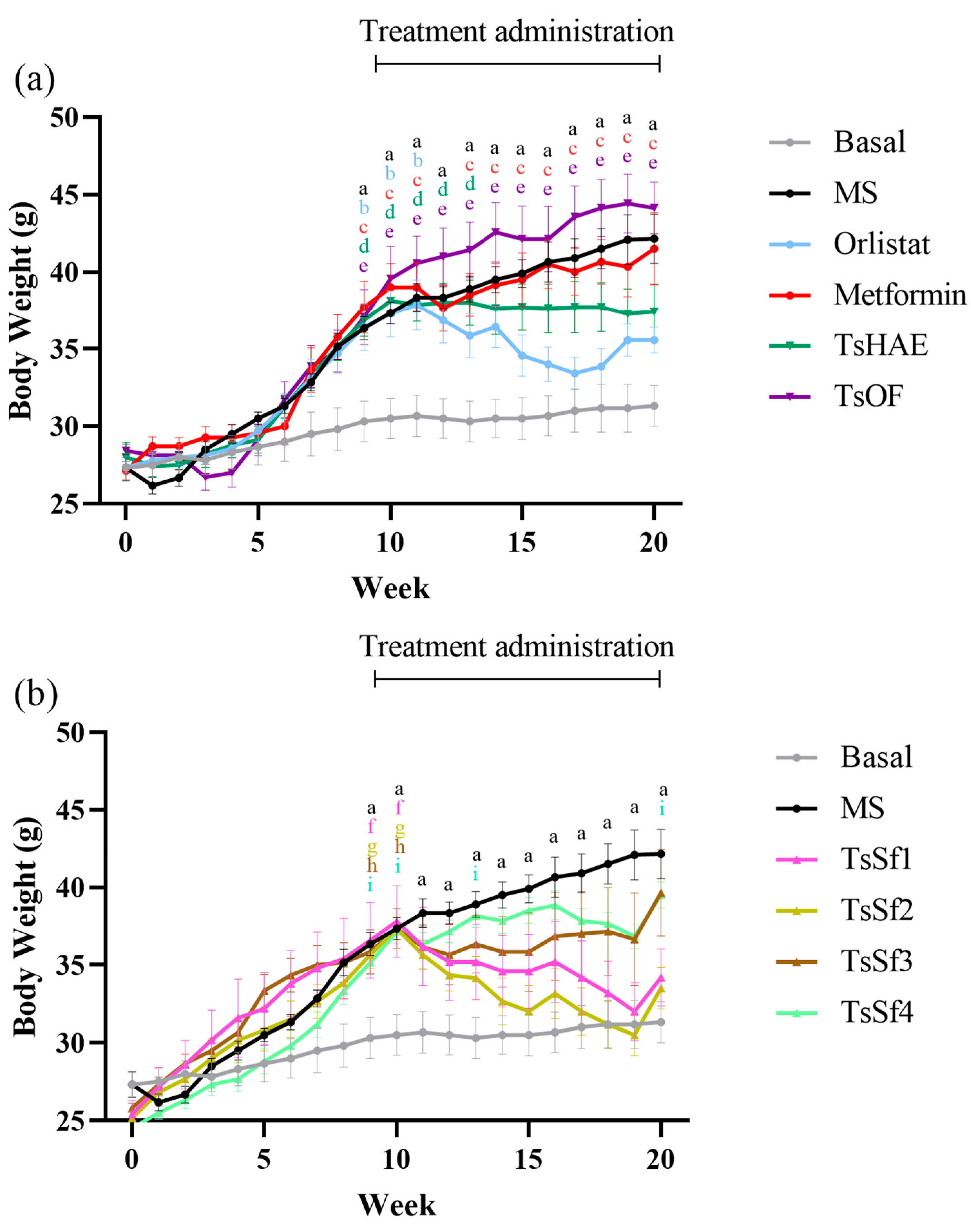
2.2.2. Body Weight Parameters
2.2.3. Atherogenic Dyslipidaemia
2.2.4. Hypertension
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Phytochemical Procedures
4.3. Experimental Animals and MS Induction Model
4.4. Evaluated Parameters
4.5. Data Processing and Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, S.; O’Driscoll, L. Metabolic Syndrome: A Closer Look at the Growing Epidemic and Its Associated Pathologies. Obes. Rev. 2015, 16, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z. The Metabolic Syndrome. Lancet 2005, 366, 1415–1428. [Google Scholar] [CrossRef]
- Esposito, K.; Chiodini, P.; Colao, A.; Lenzi, A.; Giugliano, D. Metabolic Syndrome and Risk of Cancer: A Systematic Review and Meta-Analysis. Diabetes Care 2012, 35, 2402–2411. [Google Scholar] [CrossRef] [Green Version]
- McCracken, E.; Monaghan, M.; Sreenivasan, S. Pathophysiology of the Metabolic Syndrome. Clin. Dermatol. 2018, 36, 14–20. [Google Scholar] [CrossRef]
- Dimitriadis, G.; Boutati, E.; Raptis, S.A. Pathogenesis: The Lipotoxicity and Glucotoxicity Hypothesis in the Metabolic Syndrome and Type 2 Diabetes. In The Metabolic Syndrome at the Beginning of the XXI Century: A Genetic and Molecular Approach; Serrano Ríos, M., Caro, J.F., Carraro, R., Gutiérrez Fuentes, J.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 271–282. ISBN 8481748927. [Google Scholar]
- Laclaustra Gimeno, M.; Bergua Martínez, C.; Pascual Calleja, I.; Casasnovas Lenguas, J.A. Síndrome Metabólico. Concepto y Fisiopatología [Metabolic Syndrome. Concept and Pathophysiology]. Rev. Esp. Cardiol. 2006, 5, 3–10. [Google Scholar] [CrossRef]
- UNAM Atlas de Las Plantas de La Medicina Tradicional Mexicana. Available online: http://www.medicinatradicionalmexicana.unam.mx/atlas.php (accessed on 13 June 2019).
- Giovannini, P.; Howes, M.J.R.; Edwards, S.E. Medicinal Plants Used in the Traditional Management of Diabetes and Its Sequelae in Central America: A Review. J. Ethnopharmacol. 2016, 184, 58–71. [Google Scholar] [CrossRef] [Green Version]
- Román-Ramos, R.; Flores-Sáenz, J.L.; Partida-Hernández, G.; Lara-Lemus, A.; Alarcón Aguilar, F. Experimental Study of the Hypoglycemic Effect of Some Antidiabetic Plants. Arch. Investig. Med. 1991, 22, 87–93. [Google Scholar]
- Aguilar-Santamaría, L.; Ramírez, G.; Nicasio, P.; Alegría-Reyes, C.; Herrera-Arellano, A. Antidiabetic Activities of Tecoma Stans (L.) Juss. Ex Kunth L. J. Ethnopharmacol. 2009, 124, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, G.; Zavala, M.; Pérez, J.; Zamilpa, A. In Vitro Screening of Medicinal Plants Used in Mexico as Antidiabetics with Glucosidase and Lipase Inhibitory Activities. Evid. Based Complement. Alternat. Med. 2012, 2012, 701261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oršolić, N.; Sirovina, D.; Odeh, D.; Gajski, G.; Balta, V.; Šver, L.; Jembrek, M.J. Efficacy of Caffeic Acid on Diabetes and Its Complications in the Mouse. Molecules 2021, 26, 3262. [Google Scholar] [CrossRef]
- Naowaboot, J.; Pannangpetch, P.; Kukongviriyapan, V.; Prawan, A.; Kukongviriyapan, U.; Itharat, A. Mulberry Leaf Extract Stimulates Glucose Uptake and GLUT4 Translocation in Rat Adipocytes. Am. J. Chin. Med. 2012, 40, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Ong, K.W.; Hsu, A.; Tan, B.K.H. Anti-Diabetic and Anti-Lipidemic Effects of Chlorogenic Acid Are Mediated by Ampk Activation. Biochem. Pharmacol. 2013, 85, 1341–1351. [Google Scholar] [CrossRef]
- Zuñiga, L.Y.; De Aceves-De La Mora, M.C.A.; González-Ortiz, M.; Ramos-Núñez, J.L.; Martínez-Abundis, E. Effect of Chlorogenic Acid Administration on Glycemic Control, Insulin Secretion, and Insulin Sensitivity in Patients with Impaired Glucose Tolerance. J. Med. Food 2018, 21, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; He, X.; Ma, Y.; Zhao, X.; Hou, X.; Hao, E.; Deng, J.; Bai, G. Chlorogenic Acid Targeting of the AKT PH Domain Activates AKT/GSK3β/FOXO1 Signaling and Improves Glucose Metabolism. Nutrients 2018, 10, 1366. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Castro, A.J.; Zapata-Bustos, R.; Romo-Yañez, J.; Camarillo-Ledesma, P.; Gómez-Sánchez, M.; Salazar-Olivo, L.A. The Antidiabetic Plants Tecoma Stans (L.) Juss. Ex Kunth (Bignoniaceae) and Teucrium Cubense Jacq (Lamiaceae) Induce the Incorporation of Glucose in Insulin-Sensitive and Insulin-Resistant Murine and Human Adipocytes. J. Ethnopharmacol. 2010, 127, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Weng, D.; Zhou, F.; Owen, Y.D.; Qin, H.; Zhao, J.; Yu, W.; Huang, Y.; Chen, J.; Fu, H.; et al. Activation of PPARγ by a Natural Flavonoid Modulator, Apigenin Ameliorates Obesity-Related Inflammation via Regulation of Macrophage Polarization. EBioMedicine 2016, 9, 61–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, B.; Qin, W.; Wu, F.; Wang, S.; Pan, C.; Wang, L.; Zeng, B.; Ma, S.; Liang, J. Apigenin and Naringenin Regulate Glucose and Lipid Metabolism, and Ameliorate Vascular Dysfunction in Type 2 Diabetic Rats. Eur. J. Pharmacol. 2016, 773, 13–23. [Google Scholar] [CrossRef]
- Xu, N.; Zhang, L.; Dong, J.; Zhang, X.; Chen, Y.G.; Bao, B.; Liu, J. Low-Dose Diet Supplement of a Natural Flavonoid, Luteolin, Ameliorates Diet-Induced Obesity and Insulin Resistance in Mice. Mol. Nutr Food Res. 2014, 58, 1258–1268. [Google Scholar] [CrossRef]
- Su, T.; Huang, C.; Yang, C.; Jiang, T.; Su, J.; Chen, M.; Fatima, S.; Gong, R.; Hu, X.; Bian, Z.; et al. Apigenin Inhibits STAT3/CD36 Signaling Axis and Reduces Visceral Obesity. Pharmacol. Res. 2020, 152, 104586. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.S.; Qu, W. Dietary Apigenin Promotes Lipid Catabolism, Thermogenesis, and Browning in Adipose Tissues of HFD-Fed Mice. Food Chem. Toxicol. 2019, 133, 110780. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Gao, M.; Liu, D. Chlorogenic Acid Improves High Fat Diet-Induced Hepatic Steatosis and Insulin Resistance in Mice. Pharm. Res. 2015, 32, 1200–1209. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, G.; Zamilpa, A.; Zavala, M.; Perez, J.; Morales, D.; Tortoriello, J. Chrysoeriol and Other Polyphenols from Tecoma Stans with Lipase Inhibitory Activity. J. Ethnopharmacol. 2016, 185, 1–8. [Google Scholar] [CrossRef]
- Foster, M.T.; Shi, H.; Seeley, R.J.; Woods, S.C. Removal of Intra-Abdominal Visceral Adipose Tissue Improves Glucose Tolerance in Rats: Role of Hepatic Triglyceride Storage. Physiol. Behav. 2011, 104, 845–854. [Google Scholar] [CrossRef] [Green Version]
- Franklin, S.S. Hypertension in the Metabolic Syndrome. Metab. Syndr. Relat. Disord. 2006, 4, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, A.; Sadeghi, M.; Barghikar, M. Which Components of Metabolic Syndrome Have a Greater Effect on Mortality, CVA and Myocardial Infarction, Hyperglycemia, High Blood Pressure or Both? Adv. Biomed. Res. 2017, 6, 121. [Google Scholar] [CrossRef]
- Botello-Amaro, A.C.; González-Cortazar, M.; Herrera-Ruiz, M.; Román-Ramos, R.; Aguilar-Santamaría, L.; Tortoriello, J.; Jiménez-Ferrer, E. Hypoglycemic and Hypotensive Activity of a Root Extract of Smilax Aristolochiifolia, Standardized on N-Trans-Feruloyl-Tyramine. Molecules 2014, 19, 11366–11384. [Google Scholar] [CrossRef] [Green Version]
- Guerrero-Romero, F.; Simental-Mendía, L.E.; González-Ortiz, M.; Martínez-Abundis, E.; Ramos-Zavala, M.G.; Hernández-González, S.O.; Jacques-Camarena, O.; Rodríguez-Morán, M. The Product of Triglycerides and Glucose, a Simple Measure of Insulin Sensitivity. Comparison with the Euglycemic-Hyperinsulinemic Clamp. J. Clin. Endocrinol. Metab. 2010, 95, 3347–3351. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Compound | % |
|---|---|---|
| TsHAE | Luteolin | 0.65 ± 0.02 |
| Apigenin | 0.49 ± 0.01 | |
| Chrysoeriol | 0.39 ± 0.01 | |
| TsOF | Luteolin | 3.70 ± 0.02 |
| Apigenin | 0.91 ± 0.05 | |
| Chrysoeriol | 0.77 ± 0.05 | |
| TsSf1 | Luteolin | 4.07 ± 0.06 |
| Apigenin | 1.52 ± 0.14 | |
| Chrysoeriol | 1.18 ± 0.05 | |
| TsSf2 | Luteolin | 86.12 ± 0.41 |
| Apigenin | 12.25 ± 0.24 | |
| Chrysoeriol | 1.08 ± 0.01 | |
| TsSf3 | Luteolin | 30.86 ± 0.23 |
| Apigenin | 3.74 ± 0.06 | |
| Chrysoeriol | 1.95 ± 0.05 | |
| TsSf4 | Luteolin | 1.03 ± 0.03 |
| Apigenin | 0.16 ± 0.01 | |
| Chrysoeriol | 0.25 ± 0.01 |
| Parameter/ Group | Glucose (mg/dL) | TyG Index | GTT AUC | BW (g) | AI (%) | MAT (mg/g) | TG (mg/dL) | Cholesterol (mg/dL) | SBP (mmHg) | DBP (mmHg) |
|---|---|---|---|---|---|---|---|---|---|---|
| Basal | 65.3 ± 1.8 # | 4.47 ± 0.05 # | 19,727 ± 848 # | 31.3 ± 1.3 # | 5.4 ± 0.77 # | 2.7 ± 0.9 # | 91.4 ± 15.1 # | 158.1 ± 1.6 # | 89.9 ± 2.4 # | 61.6 ± 1.7 # |
| MS | 221.5 ± 14.6 * | 5.16 ± 0.05 * | 28,140 ± 966 * | 42.2 ± 1.6 * | 20.7 ± 0.95 * | 22.8 ± 3.7 * | 177.7 ± 14.7 * | 165.3 ± 1.2 * | 110.3 ± 3.5 * | 73.7 ± 1.8 * |
| Orlistat | 177.2 ± 12.2 * | 4.81 ± 0.05 | 22,903 ± 545 # | 35.6 ± 0.8 | 11.1 ± 0.88 *# | 10.2 ± 1.3 # | 101.9 ± 5.8 | 160.1 ± 1.8 | 87.4 ± 1.5 # | 53.0 ± 1.0 *# |
| Metformin | 178.0 ± 9.8 * | 5.01 ± 0.05 * | 18,996 ± 482 # | 41.5 ± 2.3 * | 16.5 ± 0.87 * | 20.0 ± 3.2 * | 130.2 ± 8.7 | 153.4 ± 1.1 # | 86.8 ± 1.3 # | 60.2 ± 2.3 # |
| TsHAE | 166.5 ± 12.5 * | 4.93 ± 0.10 * | 20,739 ± 1202 # | 37.4 ± 1.8 | 13.1 ± 1.96 *# | 10.3 ± 2.2 # | 125.0 ± 21.4 | 163.7 ± 0.9 * | 91.8 ± 3.9 # | 56.4 ± 4.5 # |
| TsOF | 140.3 ± 13.4 *# | 4.78 ± 0.14 # | 22,730 ± 1173 # | 44.1 ± 1.7 * | 14.6 ± 0.75 *# | 15.7 ± 3.1 * | 124.2 ± 19.5 | 158.1 ± 0.3 # | 86.6 ± 1.0 # | 52.8 ± 1.2 *# |
| TsSf1 | 144.2 ± 5.7 *# | 4.72 ± 0.02 # | 23,478 ± 580 | 34.2 ± 1.8 | 7.1 ± 0.97 # | 4.4 ± 1.1 # | 87.8 ± 2.6 # | 161.0 ± 1.3 | 75.6 ± 0.7 # | 58.8 ± 1.7 # |
| TsSf2 | 165.6 ± 5.3 * | 4.80 ± 0.03 | 22,818 ± 392 # | 33.5 ± 1.3 # | 3.9 ± 1.14 # | 2.0 ± 0.9 # | 89.6 ± 4.8 # | 160.8 ± 0.6 | 91.4 ± 3.4 # | 74.9 ± 3.5 * |
| TsSf3 | 208.6 ± 14.5 * | 4.93 ± 0.05 * | 23,798 ± 389 | 39.7 ± 2.8 | 8.9 ± 1.43 # | 9.4 ± 2.1 # | 93.6 ± 4.3 # | 161.0 ± 3.7 | 81.1 ± 2.4 # | 65.0 ± 2.3 |
| TsSf4 | 200.0 ± 7.8 * | 4.86 ± 0.03 | 24,609 ± 785 | 39.5 ± 0.8 | 8.0 ± 0.82 # | 7.4 ± 1.0 # | 83.4 ± 2.2 # | 166.0 ± 2.0 * | 75.1 ± 1.5 # | 57.8 ± 2.1 # |
| Group | Diet | Treatment in Weeks 10 to 20 |
|---|---|---|
| Basal | Standard | Vehicle (5% tween 20) |
| MS | Hypercaloric | Vehicle (5% tween 20) |
| Orlistat | Hypercaloric | Orlistat 40 mg/kg |
| Metformin | Hypercaloric | Metformin 100 mg/kg |
| TsHAE | Hypercaloric | T. stans hydroalcoholic extract 25 mg/kg |
| TsOF | Hypercaloric | T. stans organic fraction 25 mg/kg |
| TsSf1 | Hypercaloric | T. stans subfraction 1 25 mg/kg |
| TsSf2 | Hypercaloric | T. stans subfraction 2 25 mg/kg |
| TsSf3 | Hypercaloric | T. stans subfraction 3 25 mg/kg |
| TsSf4 | Hypercaloric | T. stans subfraction 4 25 mg/kg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales-Ferra, D.L.; Zavala-Sánchez, M.Á.; Jiménez-Ferrer, E.; González-Cortazar, M.; Zamilpa, A. Effect of Tecoma stans (L.) Juss. ex Kunth in a Murine Model of Metabolic Syndrome. Plants 2022, 11, 1794. https://doi.org/10.3390/plants11141794
Morales-Ferra DL, Zavala-Sánchez MÁ, Jiménez-Ferrer E, González-Cortazar M, Zamilpa A. Effect of Tecoma stans (L.) Juss. ex Kunth in a Murine Model of Metabolic Syndrome. Plants. 2022; 11(14):1794. https://doi.org/10.3390/plants11141794
Chicago/Turabian StyleMorales-Ferra, Dulce Lourdes, Miguel Ángel Zavala-Sánchez, Enrique Jiménez-Ferrer, Manasés González-Cortazar, and Alejandro Zamilpa. 2022. "Effect of Tecoma stans (L.) Juss. ex Kunth in a Murine Model of Metabolic Syndrome" Plants 11, no. 14: 1794. https://doi.org/10.3390/plants11141794
APA StyleMorales-Ferra, D. L., Zavala-Sánchez, M. Á., Jiménez-Ferrer, E., González-Cortazar, M., & Zamilpa, A. (2022). Effect of Tecoma stans (L.) Juss. ex Kunth in a Murine Model of Metabolic Syndrome. Plants, 11(14), 1794. https://doi.org/10.3390/plants11141794