
Analysis of the RNA-Dependent RNA Polymerase 1 (RDR1) Gene Family in Melon


 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants, Pathogens and Inoculations
2.2. Phylogenetic Analysis
2.3. Genome Mapping and Expression Analysis by PCR and QPCR
2.4. CRISPR/Cas9 Binary Construct Design
2.5. Agrobacterium-Mediated Transformation
2.6. Genotyping and Mutant Verification
3. Results
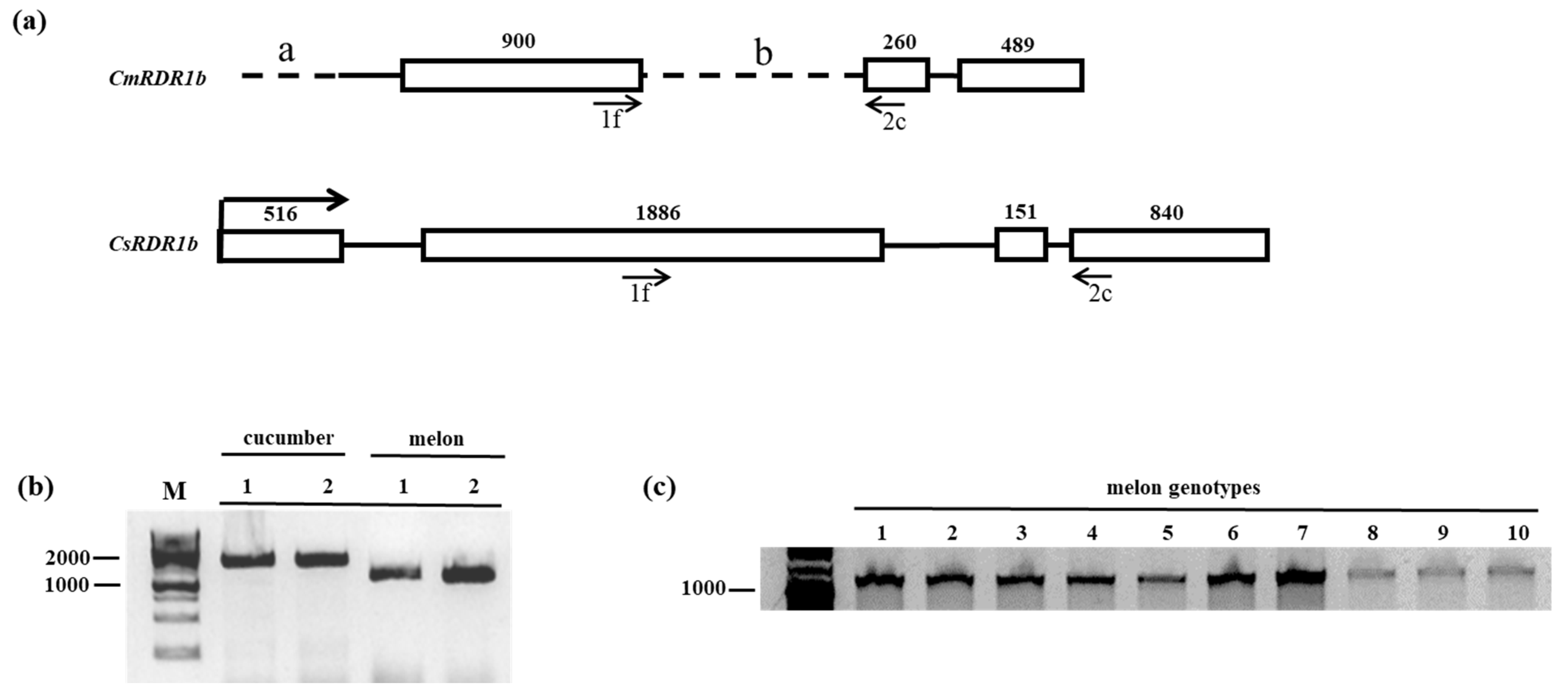
3.1. CmRDR1 Gene Family Mapping in Melon
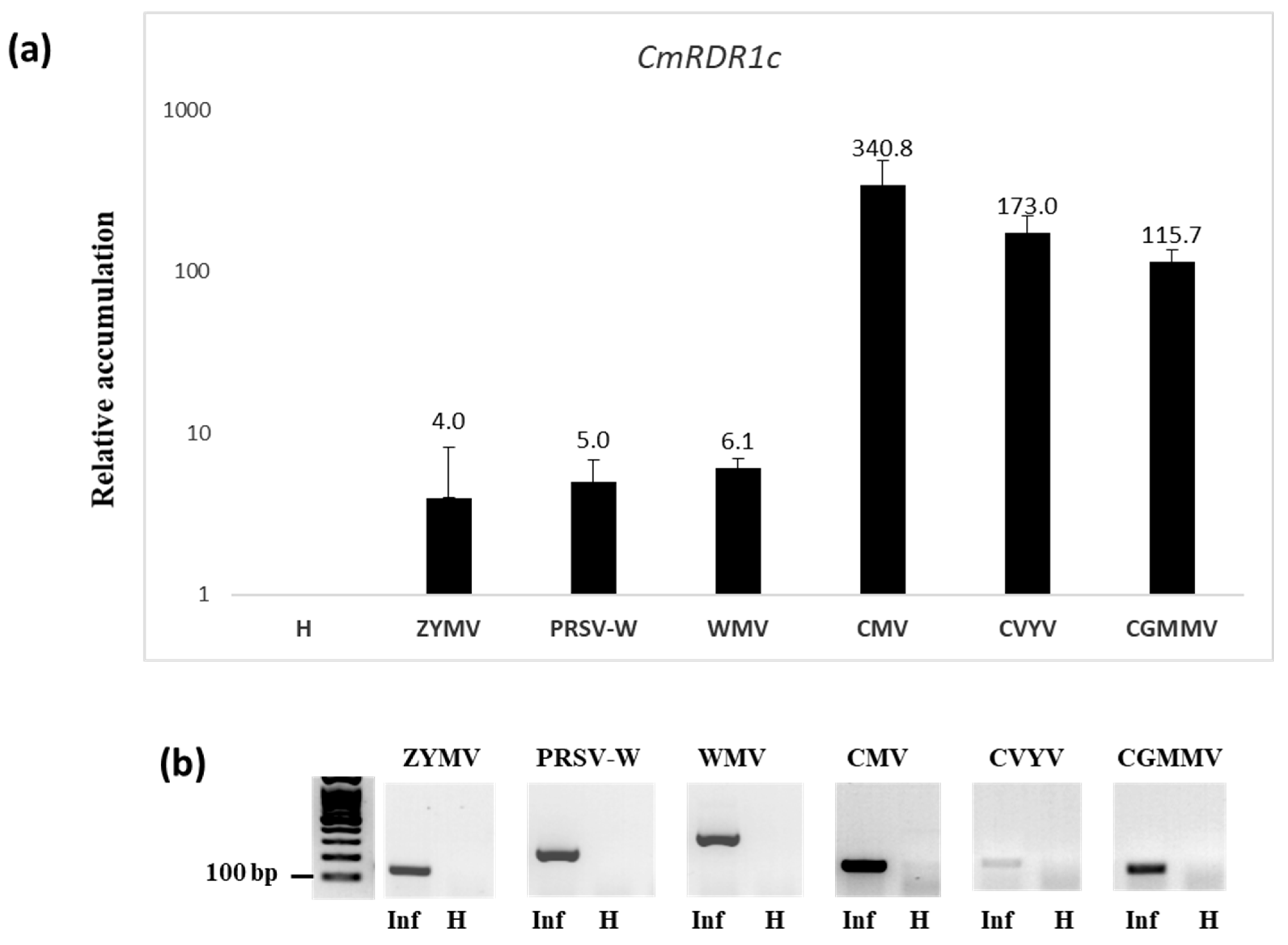
3.2. Expression Level of RDR1 Gene Family in Melon Infected with Different Viruses
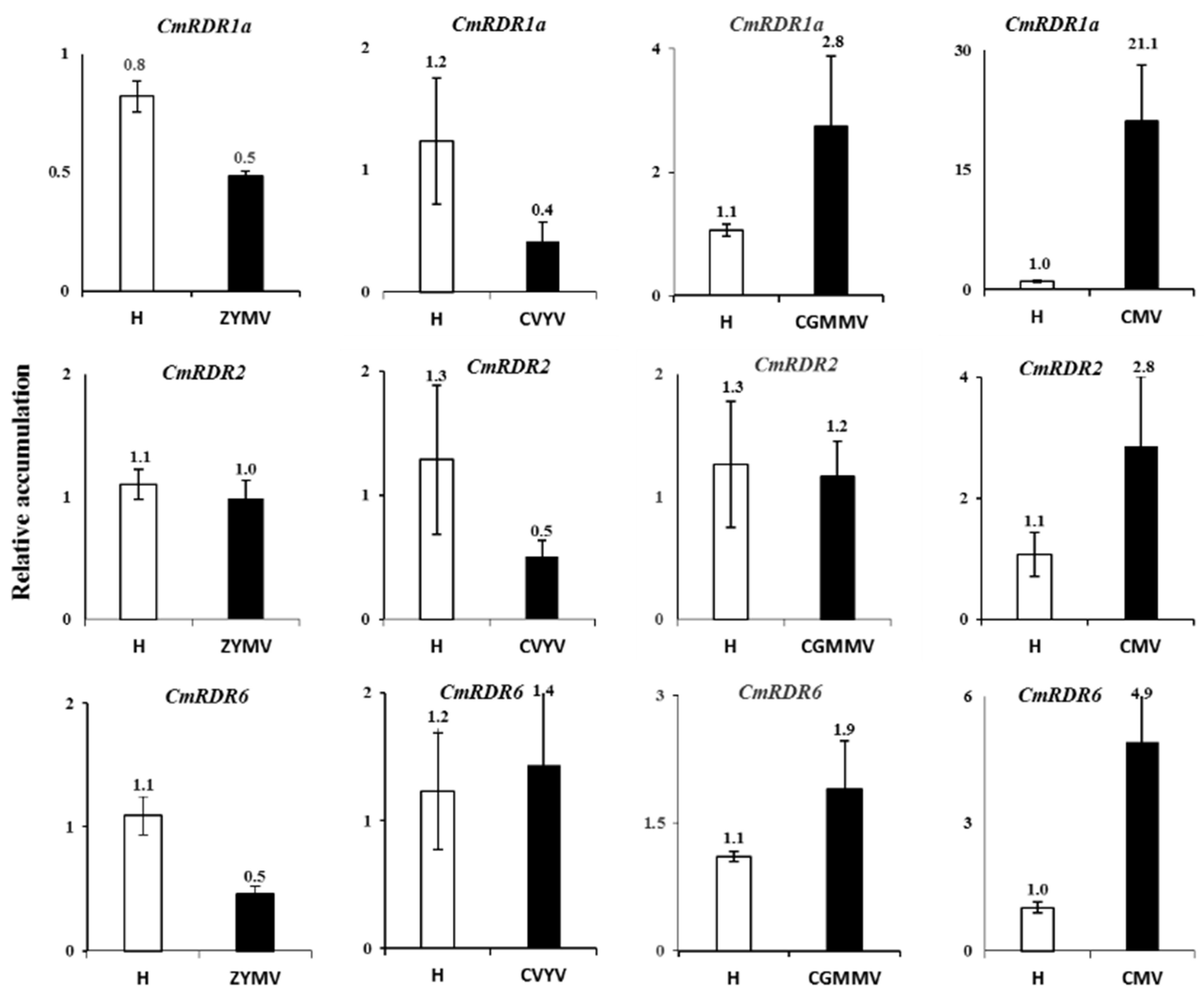
3.3. Expression of CmRDR2, CmRDR6 and CmRDR1a Genes upon Virus Infection
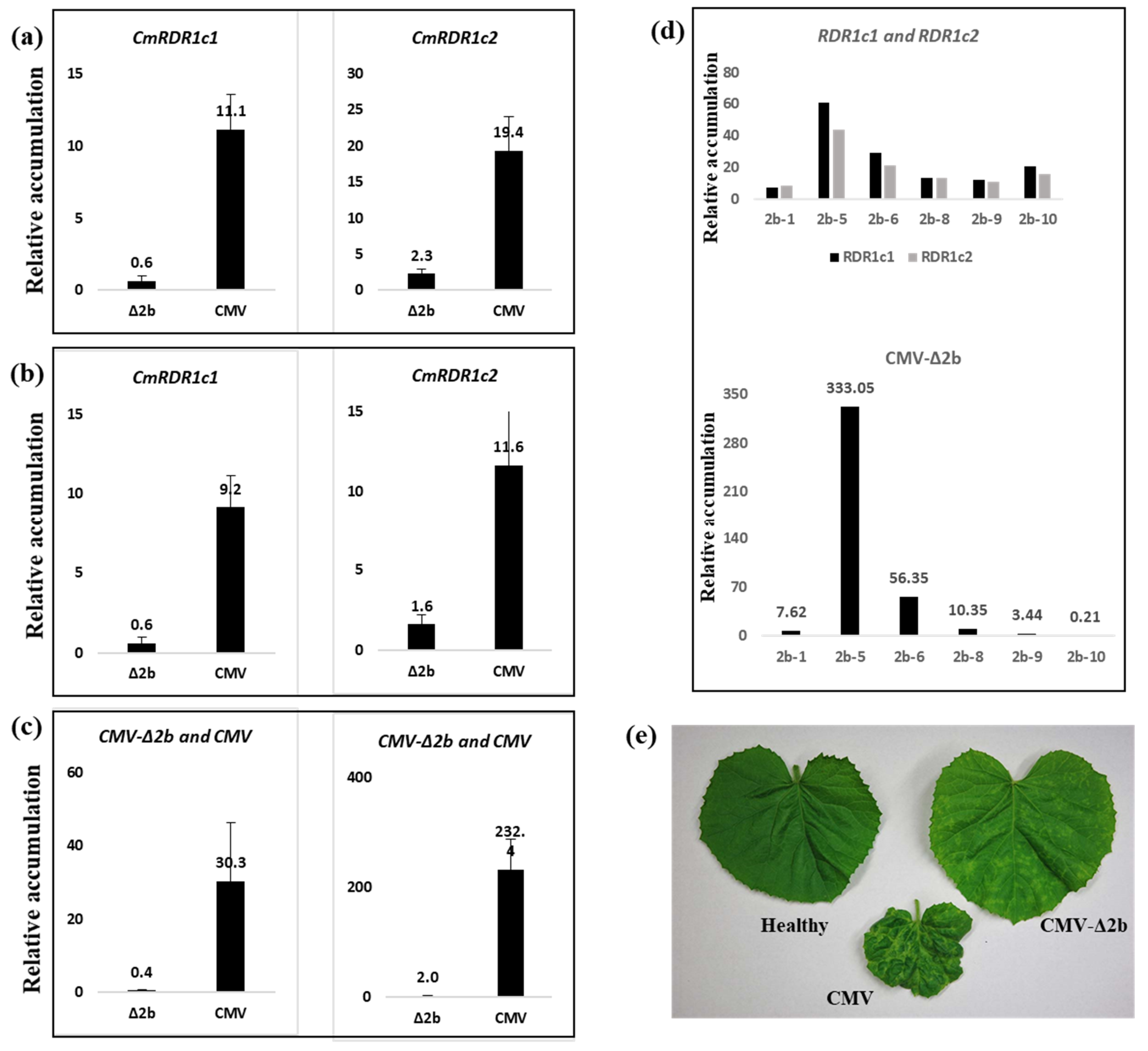
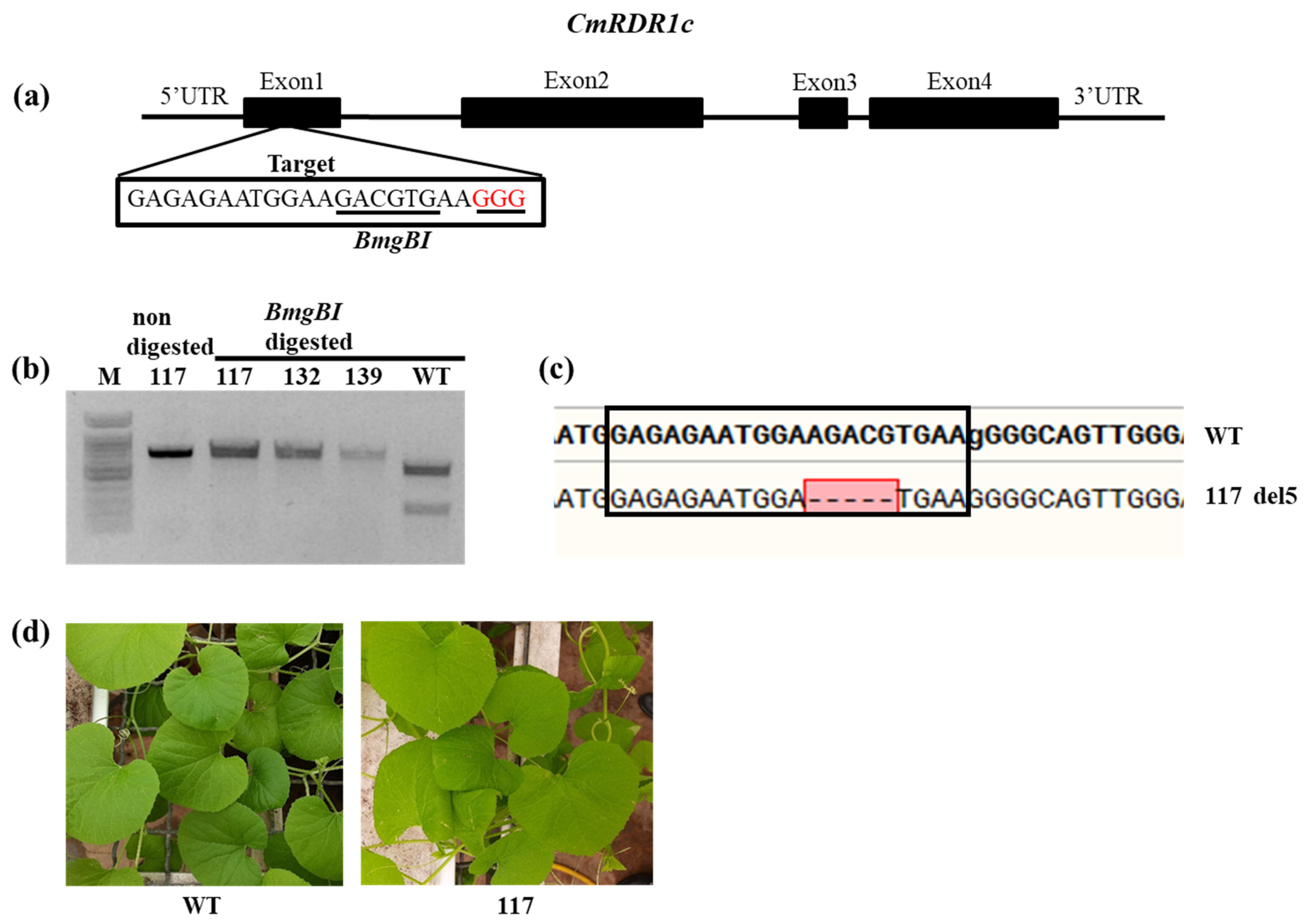
3.4. Evaluation of CmRDR1c (1+2) Contribution to Virus Severity
4. Discussion
4.1. CmRDR1 Gene Organization
4.2. Characterization of Truncated CmRDR1b Gene in Melon Genome
4.3. Kinetics of RDR1a, b, c1+c2 Gene Expression Response to Virus Infection
4.4. The Sensitivity of cmrdr1c1+2 Mutant Plants to Virus Infection
4.5. Virus Susceptibility in Melon
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#home (accessed on 22 June 2022).
- Lecoq, H.; Desbiez, C. Viruses of cucurbit crops in the Mediterranean region: An ever-changing picture. Adv. Virus Res. 2012, 84, 67–126. [Google Scholar] [CrossRef] [PubMed]
- Calil, I.P.; Fontes, E.P.B. Plant immunity against viruses: Antiviral immune receptors in focus. Ann. Bot. 2017, 119, 711–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassenegger, M.; Krczal, G. Nomenclature and functions of RNA-directed RNA polymerases. Trends Plant Sci. 2006, 11, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.-W.; Voinnet, O. Antiviral Immunity Directed by Small RNAs. Cell 2007, 130, 413–426. [Google Scholar] [CrossRef] [Green Version]
- Meister, G.; Tuschl, T. Mechanisms of gene silencing by double-stranded RNA. Nature 2004, 431, 343–349. [Google Scholar] [CrossRef]
- Lopez-Gomollon, S.; Baulcombe, D.C. Roles of RNA silencing in viral and non-viral plant immunity and in the crosstalk between disease resistance systems. Nat. Rev. Mol. Cell Biol. 2022, 1–18. [Google Scholar] [CrossRef]
- Donaire, L.; Barajas, D.; Martínez-García, B.; Martínez-Priego, L.; Pagán, I.; Llave, C. Structural and Genetic Requirements for the Biogenesis of Tobacco Rattle Virus-Derived Small Interfering RNAs. J. Virol. 2008, 82, 5167–5177. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Ruiz, H.; Takeda, A.; Chapman, E.J.; Sullivan, C.M.; Fahlgren, N.; Brempelis, K.J.; Carrington, J.C. Arabidopsis RNA-dependent RNA polymerases and dicer-like proteins in antiviral defense and small interfering RNA biogenesis during Turnip Mosaic Virus infection. Plant Cell 2010, 22, 481–496. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.; Bao, F.S.; Xie, Z. Small RNA Deep Sequencing Reveals Role for Arabidopsis thaliana RNA-Dependent RNA Polymerases in Viral siRNA Biogenesis. PLoS ONE 2009, 4, e4971. [Google Scholar] [CrossRef]
- Wang, X.-B.; Wu, Q.; Ito, T.; Cillo, F.; Li, W.-X.; Chen, X.; Yu, J.-L.; Ding, S.-W. RNAi-mediated viral immunity requires amplification of virus-derived siRNAs in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 484–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, M.; Du, P.; Wang, X.; Yu, Y.-Q.; Qiu, Y.-H.; Li, W.; Gal-On, A.; Zhou, C.; Li, Y.; Ding, S.-W. Virus infection triggers widespread silencing of host genes by a distinct class of endogenous siRNAs in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 14613–14618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leibman, D.; Kravchik, M.; Wolf, D.; Haviv, S.; Weissberg, M.; Ophir, R.; Paris, H.S.; Palukaitis, P.; Ding, S.-W.; Gaba, V.; et al. Differential expression of cucumber RNA-dependent RNA polymerase 1 genes during antiviral defence and resistance. Mol. Plant Pathol. 2018, 19, 300–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, L.J.R.; Brockington, S.F.; Murphy, A.M.; Pate, A.E.; Gruden, K.; MacFarlane, S.A.; Palukaitis, P.; Carr, J.P. RNA-dependent RNA polymerase 1 in potato (Solanum tuberosum) and its relationship to other plant RNA-dependent RNA polymerases. Sci. Rep. 2016, 6, 23082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen, C.T.; Stephens, J.; Hornyik, C.; Shaw, J.; Collinge, D.B.; Lacomme, C.; Albrechtsen, M. Identification and characterization of barley RNA-directed RNA polymerases. Biochim. Biophys. Acta BBA-Gene Regul. Mech. 2009, 1789, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Fan, B.; MacFarlane, S.A.; Chen, Z. Analysis of the Involvement of an Inducible Arabidopsis RNA-Dependent RNA Polymerase in Antiviral Defense. Mol. Plant-Microbe Interact. 2003, 16, 206–216. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Dong, Z.; Jia, Z.; Wang, J.; Wang, G. Isolation, expression and functional analysis of a putative RNA-dependent RNA polymerase gene from maize (Zea mays L.). Mol. Biol. Rep. 2010, 37, 865–874. [Google Scholar] [CrossRef]
- Gao, Q.; Liu, Y.; Wang, M.; Zhang, J.; Gai, Y.; Zhu, C.; Guo, X. Molecular cloning and characterization of an inducible RNA-dependent RNA polymerase gene, GhRdRP, from cotton (Gossypium hirsutum L.). Mol. Biol. Rep. 2009, 36, 47–56. [Google Scholar] [CrossRef]
- Pandey, S.P.; Baldwin, I.T. RNA-directed RNA polymerase 1 (RdR1) mediates the resistance of Nicotiana attenuata to herbivore attack in nature. Plant J. 2007, 50, 40–53. [Google Scholar] [CrossRef]
- Liao, Y.-W.-K.; Liu, Y.-R.; Liang, J.-Y.; Wang, W.-P.; Zhou, J.; Xia, X.-J.; Zhou, Y.-H.; Yu, J.-Q.; Shi, K. The relationship between the plant-encoded RNA-dependent RNA polymerase 1 and alternative oxidase in tomato basal defense against Tobacco mosaic virus. Planta 2015, 241, 641–650. [Google Scholar] [CrossRef]
- Wang, H.; Jiao, X.; Kong, X.; Hamera, S.; Wu, Y.; Chen, X.; Fang, R.; Yan, Y. A Signaling Cascade from miR444 to RDR1 in Rice Antiviral RNA Silencing Pathway. Plant Physiol. 2016, 170, 2365–2377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, A.; Iida, K.; Tanaka, M.; Yamaguchi, K.; Mizuhashi, K.; Kim, J.-M.; Takahashi, S.; Kobayashi, N.; Shigenobu, S.; Shinozaki, K.; et al. Novel Stress-Inducible Antisense RNAs of Protein-Coding Loci Are Synthesized by RNA-Dependent RNA Polymerase. Plant Physiol. 2017, 175, 457–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polydore, S.; Axtell, M.J. Analysis of RDR1/RDR2/RDR6-independent small RNAs in Arabidopsis thaliana improves MIRNA annotations and reveals unexplained types of short interfering RNA loci. Plant J. 2018, 94, 1051–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, S.P.; Shahi, P.; Gase, K.; Baldwin, I.T. Herbivory-induced changes in the small-RNA transcriptome and phytohormone signaling in Nicotiana attenuata. Proc. Natl. Acad. Sci. USA 2008, 105, 4559–4564. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.-J.; Carter, S.A.; Cole, A.B.; Cheng, N.-H.; Nelson, R.S. A natural variant of a host RNA-dependent RNA polymerase is associated with increased susceptibility to viruses by Nicotiana benthamiana. Proc. Natl. Acad. Sci. USA 2004, 101, 6297–6302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, L.; Mo, N.; Zhang, Y.; Muhammad, T.; Zhao, G.; Zhang, Y.; Liang, Y. CaRDR1, an RNA-Dependent RNA Polymerase Plays a Positive Role in Pepper Resistance against TMV. Front. Plant Sci. 2017, 8, 1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakhshandehroo, F.; Takeshita, M.; Squires, J.; Palukaitis, P. The influence of RNA-dependent RNA polymerase 1 on potato virus Y infection and on other antiviral response genes. Mol. Plant-Microbe Interact. MPMI 2009, 22, 1312–1318. [Google Scholar] [CrossRef] [Green Version]
- Ying, X.-B.; Dong, L.; Zhu, H.; Duan, C.-G.; Du, Q.-S.; Lv, D.-Q.; Fang, Y.-Y.; Garcia, J.A.; Fang, R.-X.; Guo, H.-S. RNA-dependent RNA polymerase 1 from Nicotiana tabacum suppresses RNA silencing and enhances viral infection in Nicotiana benthamiana. Plant Cell 2010, 22, 1358–1372. [Google Scholar] [CrossRef] [Green Version]
- Qi, G.; Chen, J.; Chang, M.; Chen, H.; Hall, K.; Korin, J.; Liu, F.; Wang, D.; Fu, Z.Q. Pandemonium Breaks Out: Disruption of Salicylic Acid-Mediated Defense by Plant Pathogens. Mol. Plant 2018, 11, 1427–1439. [Google Scholar] [CrossRef] [Green Version]
- Alamillo, J.M.; Saénz, P.; García, J.A. Salicylic acid-mediated and RNA-silencing defense mechanisms cooperate in the restriction of systemic spread of plum pox virus in tobacco. Plant J. Cell Mol. Biol. 2006, 48, 217–227. [Google Scholar] [CrossRef]
- Hunter, L.J.R.; Westwood, J.H.; Heath, G.; Macaulay, K.; Smith, A.G.; MacFarlane, S.A.; Palukaitis, P.; Carr, J.P. Regulation of RNA-Dependent RNA Polymerase 1 and Isochorismate Synthase Gene Expression in Arabidopsis. PLoS ONE 2013, 8, e66530. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-S.; Fu, S.-F.; Li, Z.; Murphy, A.M.; Dobson, E.A.; Garland, L.; Chaluvadi, S.R.; Lewsey, M.G.; Nelson, R.S.; Carr, J.P. Salicylic acid treatment and expression of an RNA-dependent RNA polymerase 1 transgene inhibit lethal symptoms and meristem invasion during tobacco mosaic virus infection in Nicotiana benthamiana. BMC Plant Biol. 2016, 16, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Zhang, D.; Wang, Z.; Xun, H.; Ma, J.; Wang, H.; Huang, W.; Liu, Y.; Lin, X.; Li, N.; et al. Mutation of the RDR1 gene caused genome-wide changes in gene expression, regional variation in small RNA clusters and localized alteration in DNA methylation in rice. BMC Plant Biol. 2014, 14, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, P.; Zhao, L.; McFarlane, H.E.; Aiga, M.; Lam, V.; Hooker, T.S.; Kunst, L. RDR1 and SGS3, Components of RNA-Mediated Gene Silencing, Are Required for the Regulation of Cuticular Wax Biosynthesis in Developing Inflorescence Stems of Arabidopsis. Plant Physiol. 2012, 159, 1385–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, R.; Kumar, S.; Leibman, D.; Abebie, B.; Shnaider, Y.; Ding, S.-W.; Gal-On, A. Cucumber RDR1s and cucumber mosaic virus suppressor protein 2b association directs host defence in cucumber plants. Mol. Plant Pathol. 2021, 22, 1317–1331. [Google Scholar] [CrossRef]
- Leibman, D.; Wolf, D.; Saharan, V.; Zelcer, A.; Arazi, T.; Yoel, S.; Gaba, V.; Gal-On, A. A High Level of Transgenic Viral Small RNA Is Associated with Broad Potyvirus Resistance in Cucurbits. Mol. Plant. Microbe Interact. 2011, 24, 1220–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Sela, I.; Ashkenazy, H.; Katoh, K.; Pupko, T. GUIDANCE2: Accurate detection of unreliable alignment regions accounting for the uncertainty of multiple parameters. Nucleic Acids Res. 2015, 43, W7–W14. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version II. Plant Mol. Biol. Report. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Chandrasekaran, J.; Brumin, M.; Wolf, D.; Leibman, D.; Klap, C.; Pearlsman, M.; Sherman, A.; Arazi, T.; Gal-On, A. Development of broad virus resistance in non-transgenic cucumber using CRISPR/Cas9 technology. Mol. Plant Pathol. 2016, 17, 1140–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peccarelli, M.; Kebaara, B.W. Regulation of natural mRNAs by the nonsense-mediated mRNA decay pathway. Eukaryot. Cell 2014, 13, 1126–1135. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Cuevas, H.E.; Yang, L.; Li, Y.; Garcia-Mas, J.; Zalapa, J.; Staub, J.E.; Luan, F.; Reddy, U.; He, X.; et al. Syntenic relationships between cucumber (Cucumis sativus L.) and melon (C. melo L.) chromosomes as revealed by comparative genetic mapping. BMC Genom. 2011, 12, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, V.; Chakraborty, S. Identification of transcription factor binding sites on promoter of RNA dependent RNA polymerases (RDRs) and interacting partners of RDR proteins through in silico analysis. Physiol. Mol. Biol. Plants Int. J. Funct. Plant Biol. 2019, 25, 1055–1071. [Google Scholar] [CrossRef]
- Xu, T.; Zhang, L.; Zhen, J.; Fan, Y.; Zhang, C.; Wang, L. Expressional and regulatory characterization of Arabidopsis RNA-dependent RNA polymerase 1. Planta 2013, 237, 1561–1569. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mas, J.; Benjak, A.; Sanseverino, W.; Bourgeois, M.; Mir, G.; González, V.M.; Hénaff, E.; Câmara, F.; Cozzuto, L.; Lowy, E.; et al. The genome of melon (Cucumis melo L.). Proc. Natl. Acad. Sci. USA 2012, 109, 11872–11877. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Murphy, A.M.; Lewsey, M.G.; Westwood, J.H.; Zhang, H.-M.; González, I.; Canto, T.; Carr, J.P.Y. 2014 Domains of the cucumber mosaic virus 2b silencing suppressor protein affecting inhibition of salicylic acid-induced resistance and priming of salicylic acid accumulation during infection. J. Gen. Virol. 2014, 95, 1408–1413. [Google Scholar] [CrossRef] [Green Version]
- Allan, A.C.; Lapidot, M.; Culver, J.N.; Fluhr, R. An early tobacco mosaic virus-induced oxidative burst in tobacco indicates extracellular perception of the virus coat protein. Plant Physiol. 2001, 126, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, T.; Zhang, F.; Zhang, Y.; Liang, Y. RNA Interference: A Natural Immune System of Plants to Counteract Biotic Stressors. Cells 2019, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Valli, A.; Oliveros, J.C.; Molnar, A.; Baulcombe, D.; García, J.A. The specific binding to 21-nt double-stranded RNAs is crucial for the anti-silencing activity of Cucumber vein yellowing virus P1b and perturbs endogenous small RNA populations. RNA 2011, 17, 1148–1158. [Google Scholar] [CrossRef] [Green Version]
- Wechter, W.P.; Whitehead, M.P.; Thomas, C.E.; Dean, R.A. Identification of a randomly amplified polymorphic DNA marker linked to the Fom 2 Fusarium wilt resistance gene in muskmelon MR-1. Phytopathology 1995, 85, 1245–1249. [Google Scholar] [CrossRef]
- Gal-On, A. A Point Mutation in the FRNK Motif of the Potyvirus Helper Component-Protease Gene Alters Symptom Expression in Cucurbits and Elicits Protection against the Severe Homologous Virus. Phytopathology 2000, 90, 467–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, T.M.; Palukaitis, P. Construction of full-length cDNA clones of cucumber mosaic virus RNAs 1, 2 and 3: Generation of infectious RNA transcripts. Mol. Gen. Genet. MGG 1990, 222, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Reingold, V.; Lachman, O.; Blaosov, E.; Dombrovsky, A. Seed disinfection treatments do not sufficiently eliminate the infectivity of Cucumber green mottle mosaic virus (CGMMV) on cucurbit seeds. Plant Pathol. 2015, 64, 245–255. [Google Scholar] [CrossRef]
- Martínez-garcía, B.; Marco, C.F.; Goytia, E.; López-abella, D.; Serra, M.T.; Aranda, M.A.; López-moya, J.J. Development and use of detection methods specific for Cucumber vein yellowing virus (CVYV). Eur. J. Plant Pathol. 2004, 110, 811–821. [Google Scholar] [CrossRef]
- Desbiez, C.; Lecoq, H. The nucleotide sequence of Watermelon mosaic virus (WMV, Potyvirus) reveals interspecific recombination between two related potyviruses in the 5’ part of the genome. Arch. Virol. 2004, 149, 1619–1632. [Google Scholar] [CrossRef]
- Ali, A. First Complete Genome Sequence of Papaya ringspot virus-W Isolated from a Gourd in the United States. Genome Announc. 2017, 5, 1434–1516. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leibman, D.; Pashkovsky, E.; Shnaider, Y.; Shtarkman, M.; Gaba, V.; Gal-On, A. Analysis of the RNA-Dependent RNA Polymerase 1 (RDR1) Gene Family in Melon. Plants 2022, 11, 1795. https://doi.org/10.3390/plants11141795
Leibman D, Pashkovsky E, Shnaider Y, Shtarkman M, Gaba V, Gal-On A. Analysis of the RNA-Dependent RNA Polymerase 1 (RDR1) Gene Family in Melon. Plants. 2022; 11(14):1795. https://doi.org/10.3390/plants11141795
Chicago/Turabian StyleLeibman, Diana, Ekaterina Pashkovsky, Yulia Shnaider, Meital Shtarkman, Victor Gaba, and Amit Gal-On. 2022. "Analysis of the RNA-Dependent RNA Polymerase 1 (RDR1) Gene Family in Melon" Plants 11, no. 14: 1795. https://doi.org/10.3390/plants11141795
APA StyleLeibman, D., Pashkovsky, E., Shnaider, Y., Shtarkman, M., Gaba, V., & Gal-On, A. (2022). Analysis of the RNA-Dependent RNA Polymerase 1 (RDR1) Gene Family in Melon. Plants, 11(14), 1795. https://doi.org/10.3390/plants11141795