
Leaf Extracts from Resistant Wild Tomato Can Be Used to Control Late Blight (Phytophthora infestans) in the Cultivated Tomato

 ,
, 
 and
and
Abstract
:1. Introduction
2. Results
2.1. Late Blight Severity on Resistant and Susceptible Tomato Genotypes
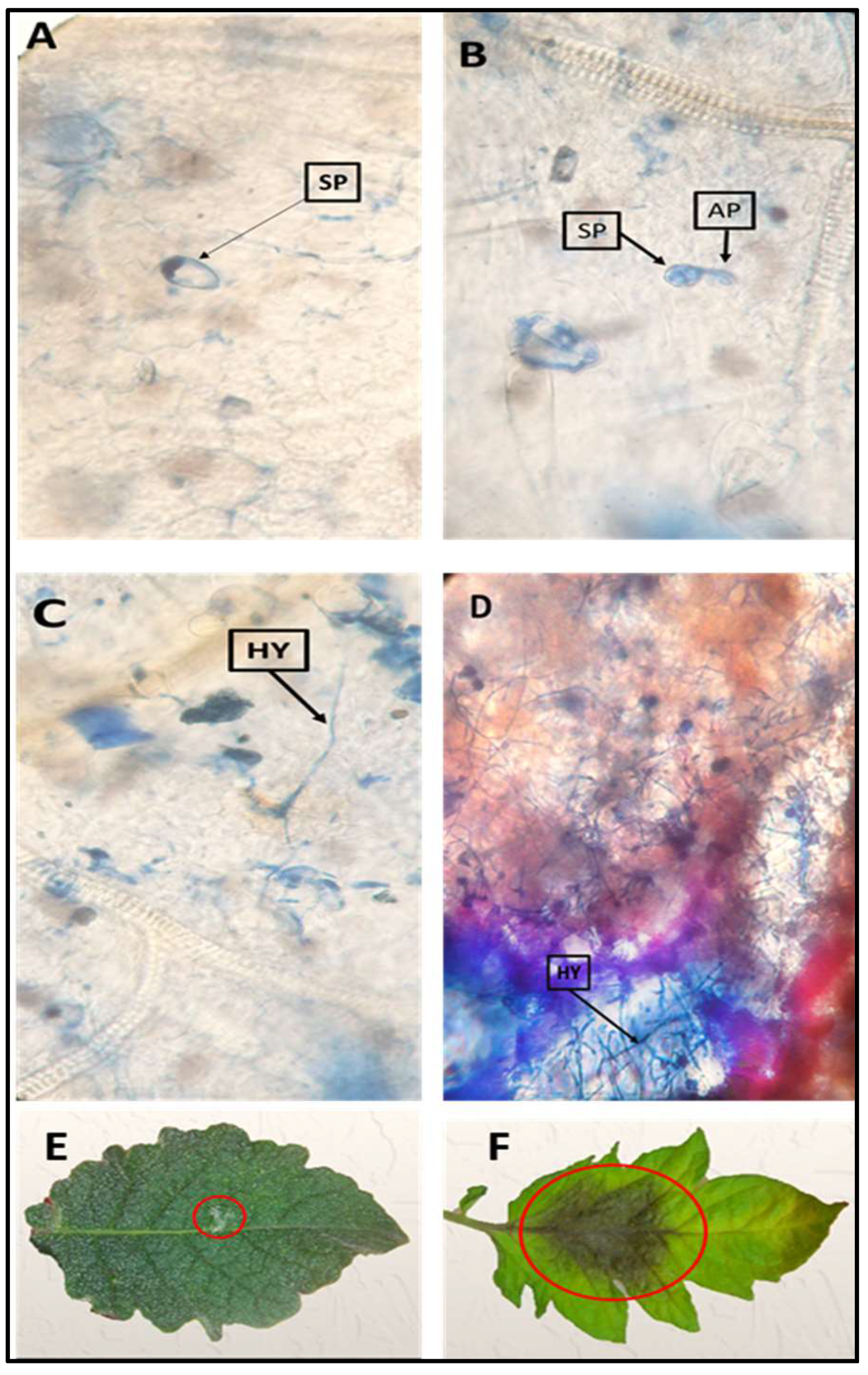
2.2. Microscopic Investigation of the Infection
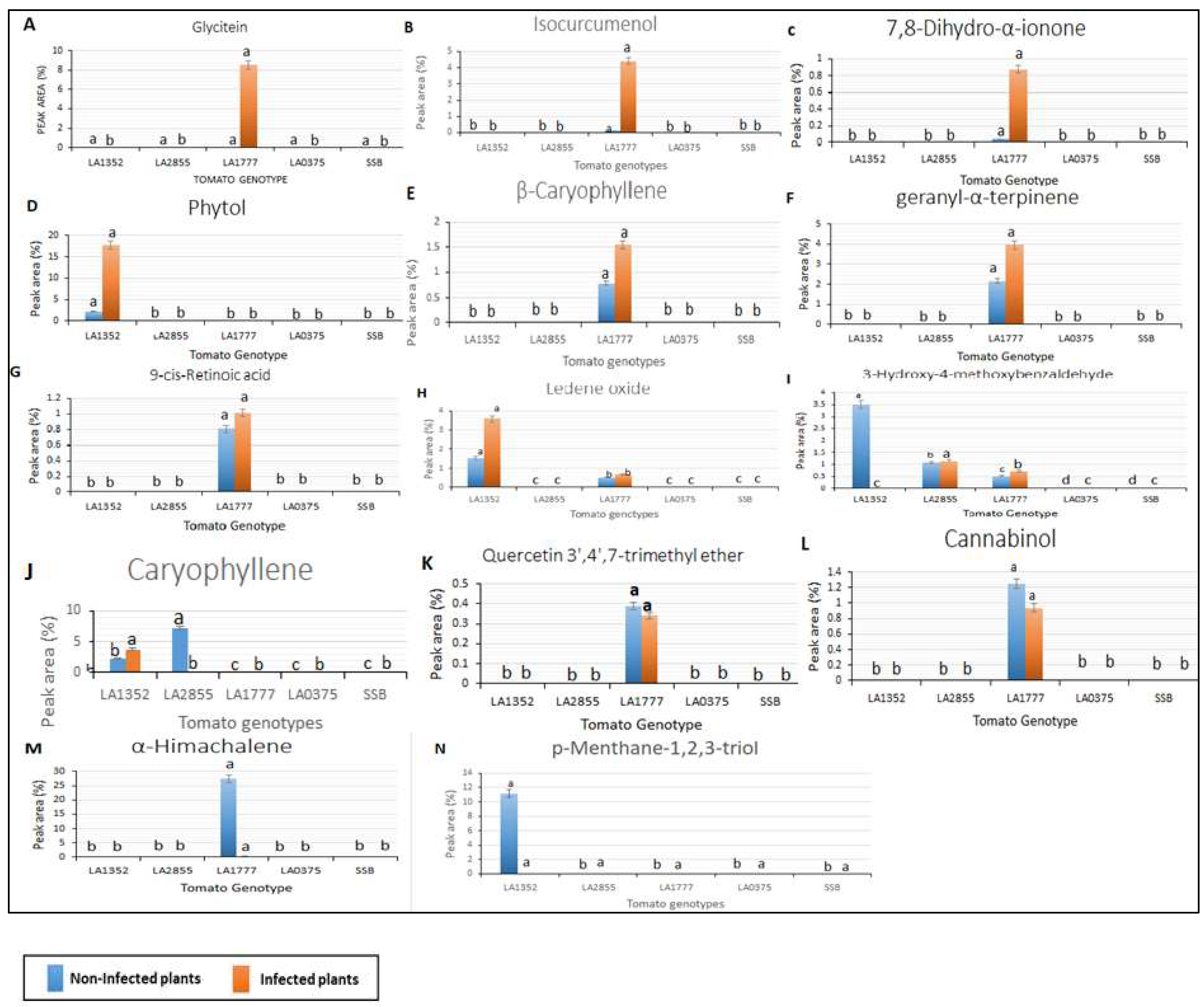
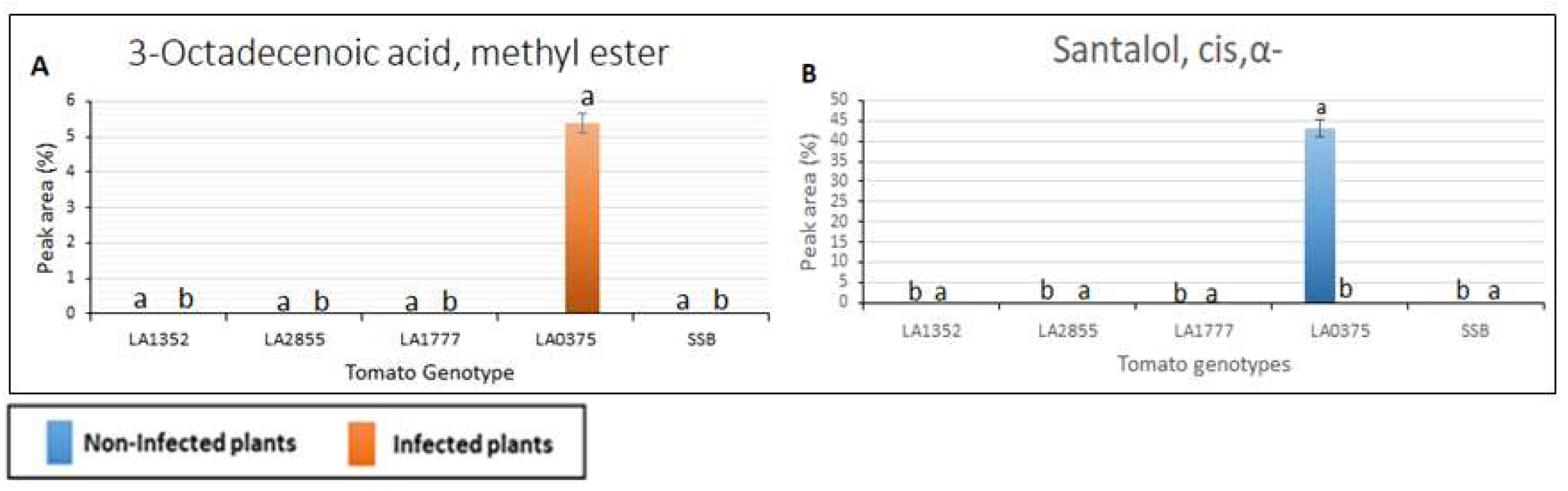
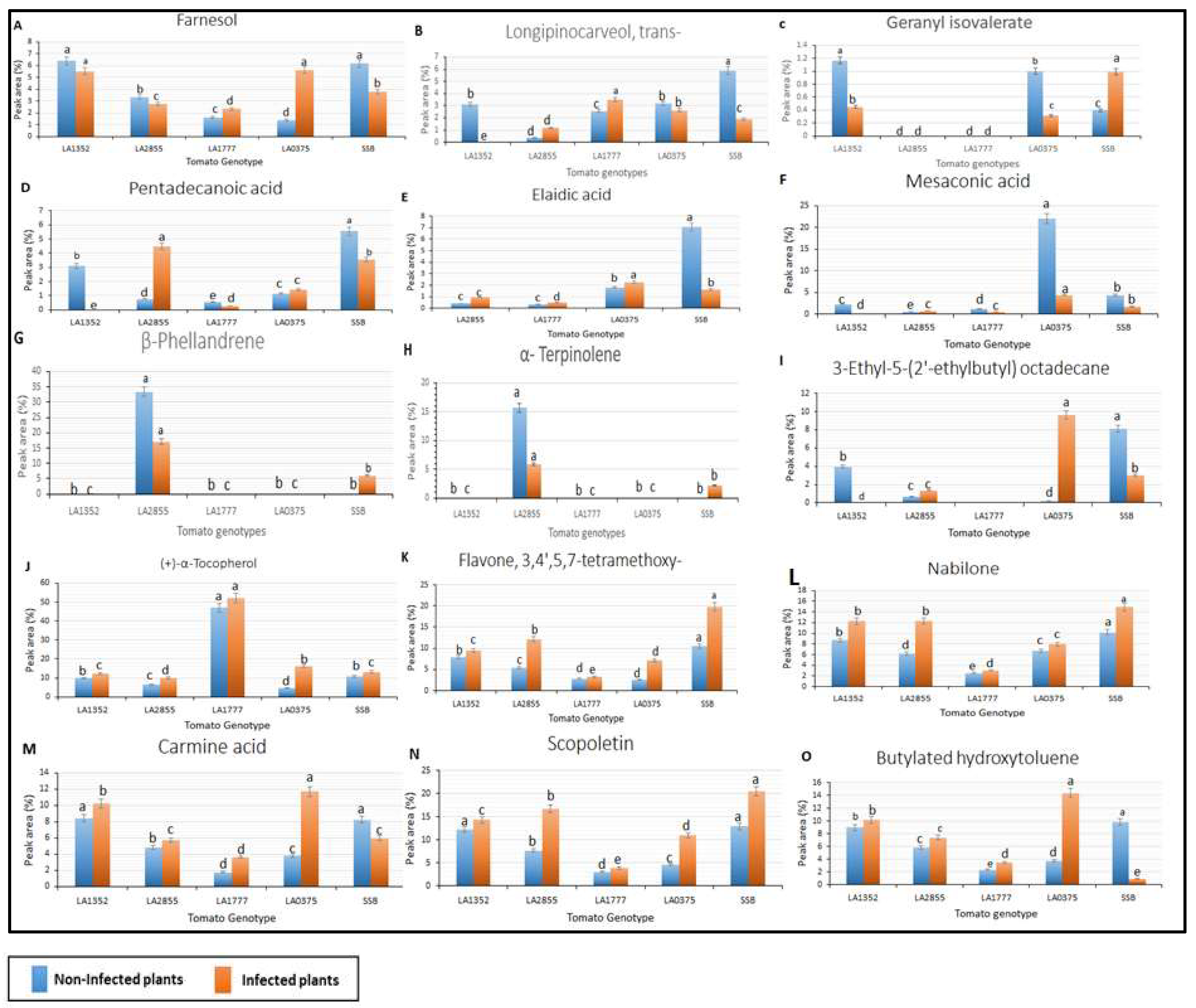
2.3. Metabolic Profiling in Late Blight Resistant and Susceptible Tomato Genotypes
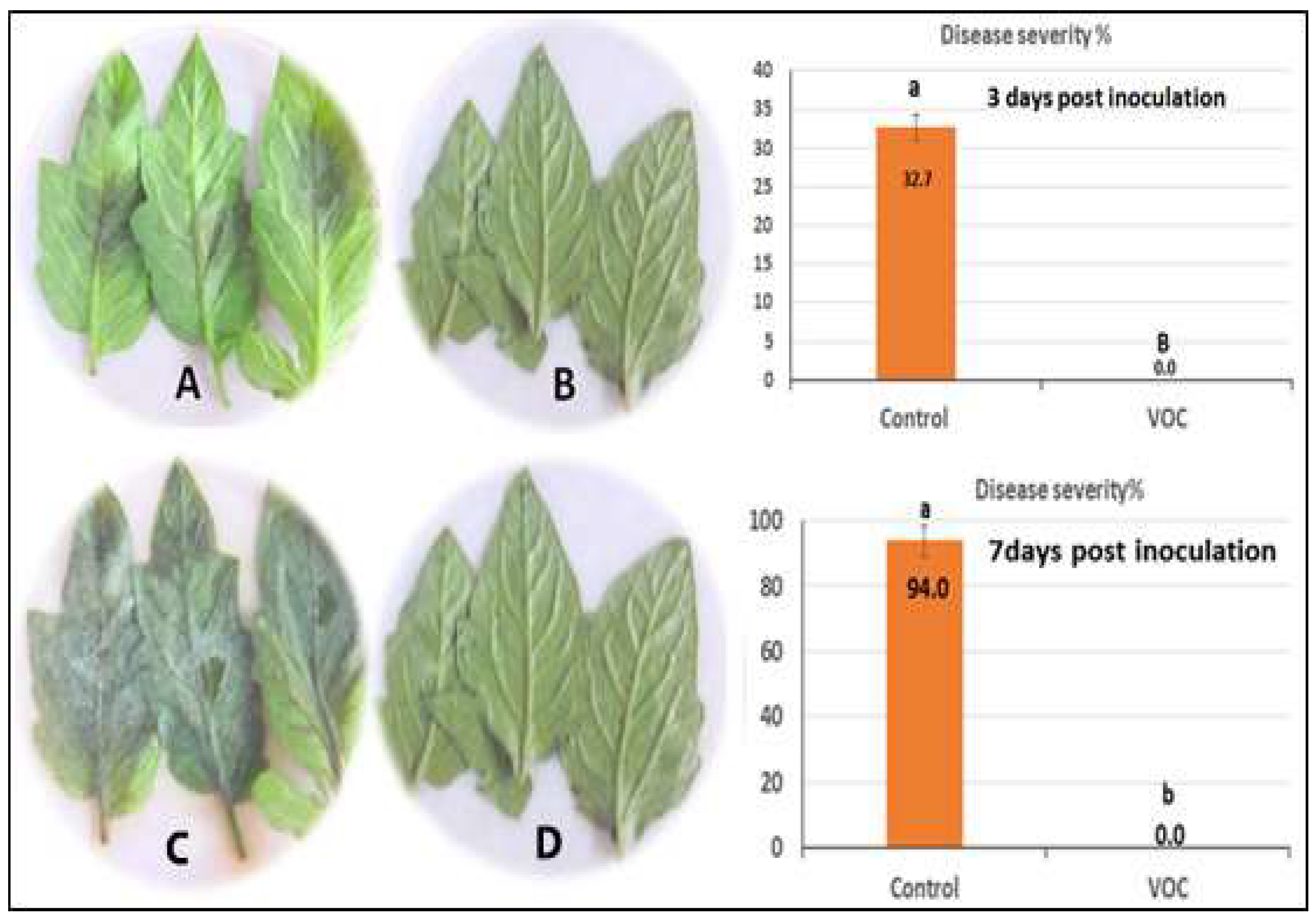
2.4. Inhibition of VOCs on Mycelial Growth of P. infestans
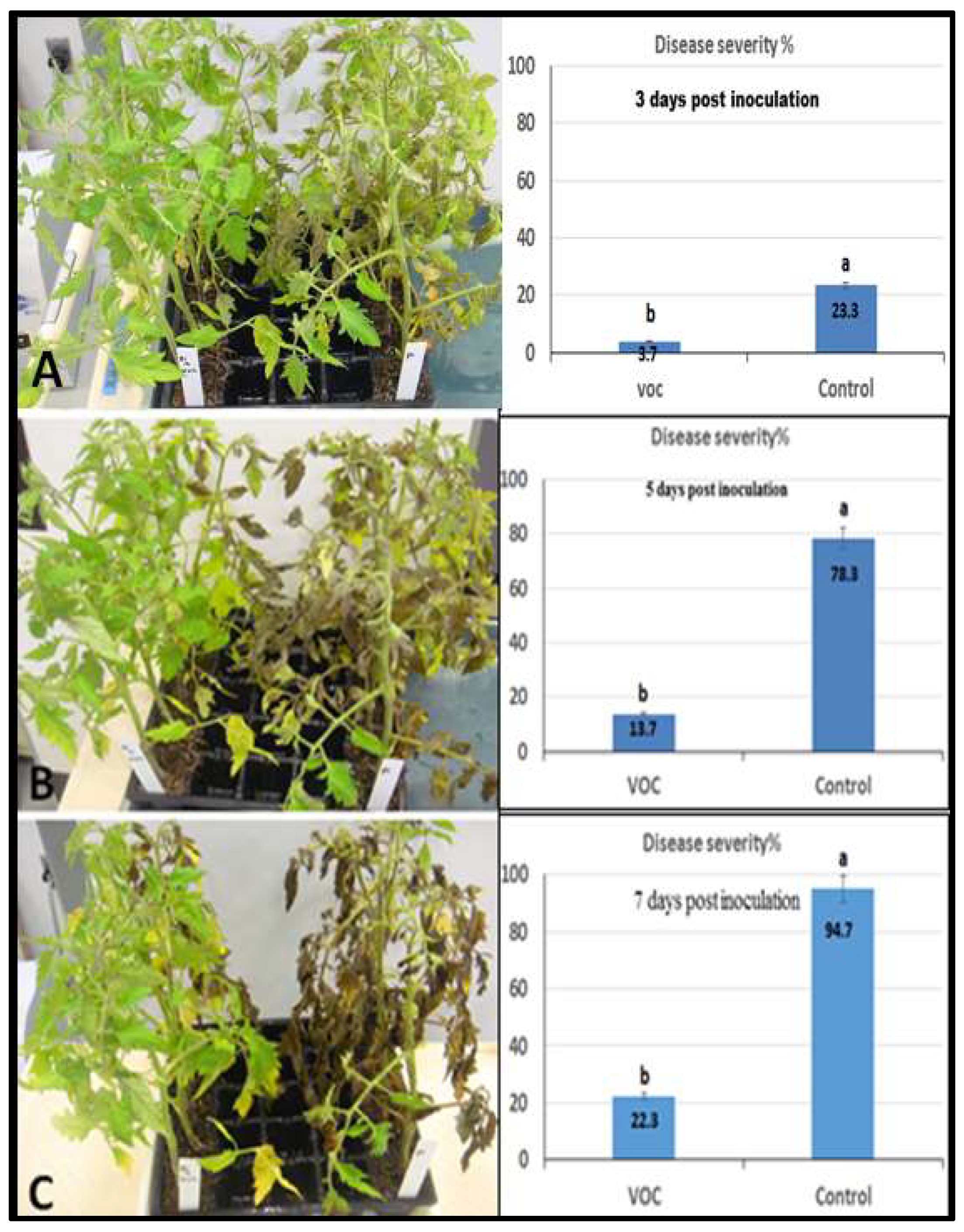
2.5. Efficacy of VOCs on Tomato Late Blight Management
3. Discussion
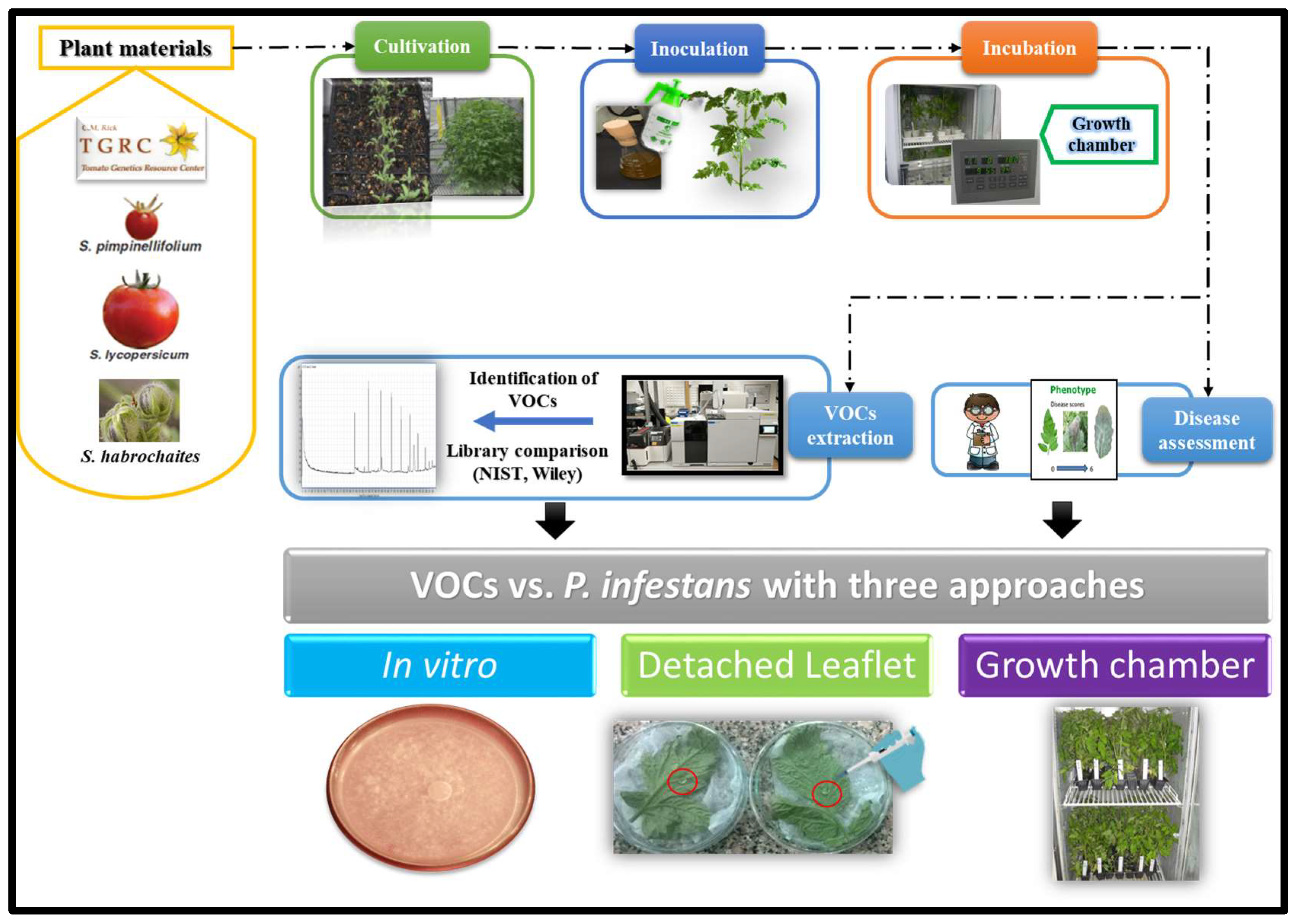
4. Materials and Methods
4.1. Plant Material and Condition
4.2. Phytophthora infestans Inoculum
4.3. Inoculation and Assessment of Late Blight Disease
4.4. Microscopic Investigation of the Infection
4.5. Gas Chromatography/Mass Spectrometry (GC/MS) Analysis of Volatiles
4.6. Effect of VOCs on P. infestans under In Vitro Conditions
4.7. Effect of VOCs on P. infestans in Leaflet and Whole Plant Assays
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quinet, M.; Angosto, T.; Yuste-Lisbona, F.J.; Blanchard-Gros, R.; Bigot, S.; Martinez, J.-P.; Lutt, S. Tomato fruit development and metabolism. Front. Plant Sci. 2019, 10, 1554. [Google Scholar] [CrossRef] [Green Version]
- FAO. FAOSTAT Statistics; Food and Agriculture Organization of United Nations: Rome, Italy, 2019. [Google Scholar]
- Sunera, A.; Saqib, S.; Uddin, S.; Zaman, W.; Ullah, F.; Ayaz, A.; Asghar, M.; Rehman, S.; Munis, M.F.H.; Chaudhary, H.J. Characterization and phytostimulatory activity of bacteria isolated from tomato (Lycopersicon esculentum L.) rhizosphere. Microb Pathog. 2020, 140, 103966. [Google Scholar] [CrossRef]
- Naeem, M.; Shahzad, K.; Saqib, S.; Shahzad, A.; Nasrullah; Younas, M.; Afridi, M.I. The Solanum melongena COP1LIKE manipulates fruit ripening and flowering time in tomato (Solanum lycopersicum). Plant Growth Regul. 2022, 96, 369–382. [Google Scholar] [CrossRef]
- Akhtar, K.P.; Saleem, M.Y.; Asghar, M.; Ahmad, M.; Sarwar, N. Resistance of Solanum species to Cucumber mosaic virus subgroup IA and its vector Myzus persicae. Eur. J. Plant Pathol. 2010, 128, 435–450. [Google Scholar] [CrossRef]
- El-Ganainy, S.M.; Iqbal, Z.; Awad, H.M.; Sattar, M.N.; Tohamy, A.M.; Abbas, A.O.; Squires, J.; Cooke, D.E.L. Genotypic and phenotypic structure of the population of Phytophthora infestans in Egypt revealed the presence of European genotypes. J. Fungi 2022, 8, 468. [Google Scholar] [CrossRef]
- Foolad, M.R.; Merk, H.L.; Ashrafi, H. Genetics, genomics and breeding of late blight and early blight resistance in tomato. Crit. Rev. Plant Sci. 2008, 27, 75–107. [Google Scholar] [CrossRef]
- Nowicki, M.; Foolad, M.R.; Nowakowska, M.; Kozik, E.U. Potato and tomato late blight caused by Phytophthora infestans: An overview of pathology and resistance breeding. Plant Dis. 2012, 96, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.L.; Liu, C.Y.; Chen, C.H.; Wang, J.F.; Liao, Y.C.; Chang, C.H.; Tsai, M.H.; Hwu, K.K.; Chen, K.Y. Reassessment of QTLs for late blight resistance in the tomato accession L3708 using a restriction site associated DNA (RAD) linkage map and highly aggressive isolates of Phytophthora infestans. PLoS ONE 2014, 9, e96417. [Google Scholar] [CrossRef] [Green Version]
- Merk, H.L.; Ashrafi, H.; Foolad, M.R. Selective genotyping to identify late blight resistance genes in an accession of the tomato wild species Solanum pimpine lifolium. Euphytica 2012, 187, 63–75. [Google Scholar] [CrossRef]
- Li, J.; Liu, L.; Bai, Y.; Finkers, R.; Wang, F.; Du, Y.; van Heusden, A.W. Identification and mapping of quantitative resistance to late blight (Phytophthora infestans) in Solanum habrochaites LA1777. Euphytica 2011, 179, 427–438. [Google Scholar] [CrossRef] [Green Version]
- Arafa, R.A.; Moussa, O.M.; Soliman, N.E.; Shirasawa, K.; Kamel, S.M.; Rakha, M.T. Resistance to Phytophthora infestans in tomato wild relatives. Afr. J. Agric. Res. 2017, 12, 2188–2196. [Google Scholar]
- Brouwer, D.J.; St. Clair, D.A. Fine mapping of three quantitative trait loci for late blight resistance in tomato using near isogenic lines (NILs) and sub-NILs. Theor. Appl. Genet. 2004, 108, 628–638. [Google Scholar] [CrossRef]
- Johnson, E.B.; Haggard, J.E.; St. Clair, D.A. Fractionation, stability, and isolate-specificity of QTL for resistance to Phytophthora infestans in cultivated tomato (Solanum lycopersicum). Genes Genomes Genet. 2012, 2, 1145–1159. [Google Scholar]
- Arafa, R.A.; Rakha, M.T.; Soliman, N.E.K.; Moussa, O.M.; Kamel, S.M.; Shirasawa, K. Rapid identification of candidate genes for resistance to tomato late blight disease using next-generation sequencing technologies. PLoS ONE 2017, 12, e0189951. [Google Scholar] [CrossRef] [Green Version]
- Arafa, R.A.; Rakha, M.T.; Kamel, S.M.; Soliman, N.E.K.; Moussa, O.M.; Shirasawa, K. Identification of resistance locus and characterization of effector genes toward breeding for resistance to Phytophthora infestans. Acta Hortic. 2021, 1316, 9–14. [Google Scholar] [CrossRef]
- Bennewitz, S.; Bergau, N.; Tissier, A. QTL Mapping of the Shape of Type VI Glandular Trichomes in Tomato. Front. Plant Sci. 2018, 9, 1421. [Google Scholar] [CrossRef] [Green Version]
- Bergau, N.; Bennewitz, S.; Syrowatka, F.; Hause, G.; Tissier, A. The development of type VI glandular trichomes in the cultivated tomato Solanum lycopersicum and a related wild species S. habrochaites. BMC Plant Biol. 2015, 15, 289. [Google Scholar] [CrossRef] [Green Version]
- Fiehn, O. Metabolomics-the link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Panthee, D.R.; Chen, F. Genomics of Fungal Disease Resistance in Tomato. Curr. Genom. 2010, 11, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Kliebenstein, D.J. Secondary metabolites and plant/environment interactions: A view through Arabidopsis thaliana tinged glasses. Plant Cell Environ. 2004, 27, 675–684. [Google Scholar] [CrossRef]
- Yuan, J.S.; Kollner, T.G.; Wiggins, G.; Grant, J.; Degenhardt, J.; Chen, F. Molecular and genomic basis of volatile-mediated indirect defense against insects in rice. Plant J. 2008, 55, 491–503. [Google Scholar] [CrossRef]
- Abu-Nada, Y.; Kushalappa, A.C.; Prasher, S.O.; Al-Mughrabi, K.I.; Murphy, A.M. Metabolic profiling to phenotype potato genotypes varying in horizontal resistance to leaf infection by Phytophthora infestans. Am. J. Plant Sci. Biotechnol. 2010, 4, 55–64. [Google Scholar]
- Karlsson, M.; Atanasova, L.; Jensen, D.F.; Zeilinger, S. Necrotrophic mycoparasites and their genomes. Microbiol. Spectr. 2017, 5, 1005–1026. [Google Scholar] [CrossRef]
- Abu-Nada, Y.; Kushalappa, A.C.; Marshall, W.D.; Al-Mughrabi, K.; Murphy, A.M. Temporal dynamics of pathogenesis-related metabolites and their plausible pathways of induction in potato leaves following inoculation with Phytophthora infestans. Eur. J. Plant Pathol. 2007, 118, 375–391. [Google Scholar] [CrossRef]
- Laothawornkitkul, J.; Jansen, R.M.C.; Smid, H.M.; Bouwmeester, H.J.; Muller, J.; van Bruggen, H.C. Volatile organic compounds as a diagnostic marker of late blight infected potato plants: A pilot study. Crop Prot. 2010, 29, 872–878. [Google Scholar] [CrossRef]
- Haverkort, A.; Struik, P.; Visser, R.G.F.; Jacobsen, E. Applied biotechnology to combat late blight in potato caused by Phytophthora infestans. Potato Res. 2009, 52, 249–264. [Google Scholar] [CrossRef]
- Lima, M.A.; Maffia, L.A.; Barreto, R.W.; Mizubuti, E.S.G. Phytophthora infestans in a subtropical region: Survival on tomato debris, temporal dynamics of airborne sporangia and alternative hosts. Plant Pathol. 2009, 58, 87–99. [Google Scholar] [CrossRef]
- Elsayed, A.Y.; Silva, D.J.H.D.; Carneiro, P.C.S.; Mizubuti, E.S.G. The inheritance of late blight resistance derived from Solanum habrochaites. Crop Breed. Appl. Biotechnol. 2012, 12, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Kushalappa, A.C.; Gunnaiah, R. Metabolo-proteomics to discover plant biotic stress resistance genes. Trends Plant Sci. 2013, 18, 522–531. [Google Scholar] [CrossRef]
- McDowell, E.T.; Kapteyn, J.; Schmidt, A.; Li, C.; Kang, J.H.; Descour, A.; Shi, F.; Larson, M.; Schilmiller, A.; An, L.; et al. Comparative functional genomic analysis of Solanum glandular trichome types. Plant Physiol. 2011, 155, 524–539. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, B.; Westbrook, T.C.; Jones, A.D. Comparative structural profiling of trichome specialized metabolites in tomato (Solanum lycopersicum) and S. habrochaites: Acylsugar profiles revealed by UHPLC/MS and NMR. Metabolomics 2014, 10, 496–507. [Google Scholar] [CrossRef] [Green Version]
- Eigenbrode, S.D.; Trumble, J.T. Antibiosis to beet armyworm (Spodoptera exigua) in Lycopersicon accessions. HortScience 1993, 28, 932–934. [Google Scholar] [CrossRef] [Green Version]
- Al-Abdallat, A.M.; Al-Debei, H.S.; Asmar, H.; Misbeh, S.; Quraan, A.; Kvarnheden, A. An efficient in vitro-inoculation method for Tomato yellow leaf curl virus. Virol. J. 2010, 7, 84. [Google Scholar] [CrossRef] [Green Version]
- Momotaz, A.; Scott, J.W.; Schuster, D.J. Identification of quantitative trait loci conferring resistance to Bemisia tabaci in an F2 population of Solanum lycopersicum × Solanum habrochaites accession LA1777. J. Am. Soc. Hortic. Sci. 2010, 135, 134–142. [Google Scholar] [CrossRef]
- Lai, A.; Cianciolo, V.; Chiavarini, S.; Sonnino, A. Effects of glandular trichomes on the development of Phytophthora infestans infection in potato (S. tuberosum). Euphytica 2000, 114, 165–174. [Google Scholar] [CrossRef]
- Yamagiwa, Y.; Inagaki, Y.; Ichinose, Y.; Toyoda, K.; Hyakumachi, M.; Shiraishi, T. Talaromyces wortmannii FS2 emits β-Caryophyllene, which promotes plant growth and induces resistance. J. Gen. Plant Pathol. 2011, 77, 336–341. [Google Scholar] [CrossRef]
- Bettarini, F.; Borgonovi, G.E.; Fiorani, T.; Gagliardi, I.; Caprioli, V.; Massardo, P.; Chapya, A. Antiparasitic compounds from East African plants: Isolation and biological activity of anonaine, matricarianol, canthin-6-one and caryophyllene oxide. Insect Set. Applic. 1993, 14, 93–99. [Google Scholar] [CrossRef]
- Hsouna, A.B.; Hamdi, N. Phytochemical composition and antimicrobial activities of the essential oils and organic extracts from Pelargonium graveolens growing in Tunisia. Lipids Health Dis. 2012, 11, 167. [Google Scholar] [CrossRef] [Green Version]
- Smith, W.E.C.; Shivaji, R.; Williams, W.P.; Luthe, D.S.; Sandoya, G.V.; Smith, C.L.; Sparks, D.L.; Brown, A.E. A maize line resistant to herbivory constitutively releases (E)-β-Caryophyllene. J. Econ. Entomol. 2012, 105, 120–128. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sultana, F.; Kubota, M.; Koyama, H.; Hyakumachi, M. The plant growth-promoting fungus Penicillium simplicissimum GP17-2 induces resistance in Arabidopsis thaliana by activation of multiple defense signals. Plant Cell Physiol. 2007, 48, 1724–1736. [Google Scholar] [CrossRef] [Green Version]
- Naka, K.; Yokozaki, H.; Domen, T.; Hayashi, K.; Kuniyasu, H.; Yasui, W.; Lotan, R.; Tahara, E. Growth inhibition of cultured human gastric cancer cells by 9-cis-retinoic acid with induction of cdk inhibitor Wafl/Cip1/Sdil/p21 protein. Differentiation 1997, 61, 313–320. [Google Scholar] [CrossRef]
- Al-Askar, A.A.; Rashad, Y.M. Efficacy of some plant extracts against Rhizoctonia solani on pea. J. Plant Prot. Res. 2010, 50, 239–243. [Google Scholar] [CrossRef]
- Derwich, E.; Benziane, Z.; Boukir, A. Chemical composition and In vitro antibacterial activity of the essential oil of Cedrus atlantica. Int. J. Agric. Biol. 2010, 12, 381–385. [Google Scholar]
- Saini, R.K.; Devi, M.K.A.; Giridhar, P.; Ravishankar, G.A. Augmentation of major isoflavones in Glycine max L. through the elicitor-mediated approach. Acta Bot. Croat. 2013, 72, 311–322. [Google Scholar] [CrossRef] [Green Version]
- Aoki, T.; Akashi, T.; Ayabe, S. Flavonoids of leguminous plants: Structure, biological activity, and biosynthesis. J. Plant Res. 2000, 113, 475–488. [Google Scholar] [CrossRef]
- Dixon, R.A.; Ferreria, D. Molecules of interest. Genestein. Phytochemistry 2002, 60, 205–211. [Google Scholar] [CrossRef]
- Steinkellner, S.; Mammerler, R. Effect of flavonoids on the development of Fusarium oxysporum f. sp. lycopersici. J. Plant Interact. 2007, 2, 17–23. [Google Scholar] [CrossRef]
- Wegulo, S.N.; Yang, X.B.; Martinson, C.A.; Murphy, P.A. Effects of wounding and inoculation with Sclerotinia sclerotiorum on isoflavone concentrations in soybean. Can. J. Plant Sci. 2005, 85, 749–760. [Google Scholar] [CrossRef]
- Holopainen, K.; Heijari, J.; Nerg, A.M.; Vuorinen, M.; Kainulainen, P. Potential for the use of exogenous chemical elicitors in disease and insect pest management of conifer seedling production. Open For. Sci. J. 2009, 2, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Ryu, C.M.; Farag, M.A.; Hu, C.H.; Reddy, M.S.; Kloepper, J.W.; Pare, P.W. Bacterial Volatiles induce systemic resistance in Arabidopsis. Plant Physiol. 2004, 134, 1017–1026. [Google Scholar] [CrossRef] [Green Version]
- Farag, M.A.; Ryu, C.M.; Sumner, L.W.; Pare, P.W. GC-MS SPME profiling of rhizobacterial volatiles reveals prospective inducers of growth promotion and induced systemic resistance in plants. Phytochemistry 2006, 67, 2262–2268. [Google Scholar] [CrossRef] [PubMed]
- Wan, M.; Li, G.; Zhang, J.; Jiang, D.; Huang, H.C. Effect of volatile substances of Streptomyces platensis F-1 on control of plant fungal diseases. Biol. Control 2008, 46, 552–559. [Google Scholar] [CrossRef]
- Kai, M.; Haustei, M.; Molina, F.; Petri, A.; Scholz, B.; Piechulla, B. Bacterial volatiles and their action potential. Appl. Microbiol. Biotechnol. 2009, 81, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ning, P.; Zheng, L.; Huang, J.; Li, G.; Hsiang, T. Fumigant activity of volatiles of Streptomyces globisporus JK-1 against Penicillium italicum on Citrus microcarpa. Postharvest Biol. Technol. 2010, 58, 157–165. [Google Scholar] [CrossRef]
- Kong, W.-L.; Rui, L.; Ni, H.; Wu, X.-Q. Antifungal effects of volatile organic compounds produced by Rahnella aquatilis JZ-GX1 against Colletotrichum gloeosporioides in Liriodendron chinense X tulipifera. Front Microbiol. 2020, 11, 1114. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.B.; Lei, H.H.; Ma, X.Y.; Lai, T.F.; Song, H.M.; Shi, X.Q.; Li, J. Antifungal activity of 1-methylcyclopropene (1-MCP) against anthracnose (Colletotrichum gloeosporioides) in postharvest mango fruit and its possible mechanisms of action. Int. J. Food Microbiol. 2017, 241, 1–6. [Google Scholar] [CrossRef]
- Li, Y.; Cooke, D.E.L.; Jacobsen, E.; van der Lee, T. Efficient multiplex simple sequence repeat genotyping of the oomycete plant pathogen Phytophthora Infestans. J. Microbiol. Methods 2013, 92, 316–322. [Google Scholar] [CrossRef]
- Arafa, R.A.; Soliman, N.E.; Moussa, O.M.; Kamel, S.M.; Shirasawa, K. Characterization of Egyptian Phytophthora infestans population using simple sequence repeat markers. J. Gen. Plant Pathol. 2018, 84, 104–107. [Google Scholar] [CrossRef]
- Caten, C.E.; Jinks, J.L. Spontaneous variability of single isolates of Phytophthora infestans. I. Cultural variation. Canad. J. Bot. 1968, 46, 329–348. [Google Scholar] [CrossRef]
- Ivanovic, M.; Niepold, F.; Mijatovic, M.; Zecevic, B. Occurrence of new populations and mating types of Phytophthora infestans (Mont.) de Bary in Serbia. Acta Hortic. 2004, 729, 499–502. [Google Scholar] [CrossRef]
- Descalzo, R.C.; Rahe, J.E.; Mauza, B. Comparative efficacy of induced resistance for selected diseases of greenhouse cucumber. Can. J. Plant Pathol. 1990, 12, 16–24. [Google Scholar] [CrossRef]
- Pandey, H.N.; Menon, T.C.M.; Rao, M.V. A simple formula for calculating area under disease progress curve. Rachis 1989, 8, 38–39. [Google Scholar]
- Saqib, S.; Zaman, W.; Ayaz, A.; Habib, S.; Bahadur, S.; Hussain, S.; Shabbir, M.; Ullah, F. Postharvest disease inhibition in fruit by synthesis and characterization of chitosan iron oxide nanoparticles. Biocatal. Agric. Biotechnol. 2020, 28, 101729. [Google Scholar] [CrossRef]
- Wilson, U.E.; Coffey, M.D. Cytological evaluation of general resistance to Phytophthora infestans in potato foliage. Ann. Bot. 1980, 45, 81–90. [Google Scholar]
- Yeom, S.I.; Seo, E.; Oh, S.K.; Kim, K.W.; Choi, D. A common plant cell-wall protein HyPRP1 has dual roles as a positive regulator of cell death and a negative regulator of basal defense against pathogens. Plant J. 2012, 69, 755–768. [Google Scholar] [CrossRef] [Green Version]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; Wiley: New York, NY, USA, 1984; p. 704. [Google Scholar]
- Hoshmand, R. Design of Experiments for Agriculture and the Natural Sciences, 2nd ed.; Chapman and Hall: New York, NY, USA, 2006; p. 456. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S No. | Taxon/Accession | Disease Severity (%) | Efficacy (%) | AUDPC | Disease Response | ||||
|---|---|---|---|---|---|---|---|---|---|
| Zero Time | 5 DPI | 10 DPI | 15 DPI | 20 DPI | |||||
| 1 | S. habrochaites (LA1777) | 0.0 | 0.7 | 1.4 | 1.9 | 2.3 d | 97.7 | 25.8 e | HR |
| 2 | S. habrochaites (LA2855) | 0.0 | 0.8 | 1.9 | 3.4 | 8.9 c | 91.0 | 52.8 d | R |
| 3 | S. habrochaites (LA1352) | 0.0 | 1.3 | 2.4 | 4.1 | 9.3 c | 90.6 | 62.3 c | R |
| 4 | S. pimpinellifolium (LA0375) | 0.0 | 24.3 | 63.7 | 77.3 | 93.7 b | 5.63 | 1060.8 b | HS |
| 5 | ‘Super Strain B’ (Cultivar) | 0.0 | 27.1 | 62.3 | 87.4 | 99.3 a | 0.0 | 1132.3 a | HS |
| L.S.D at 0.05 | 2.38 | 9.03 | |||||||
| S. No. | Taxon/Accession | Disease Severity (%) | Efficacy (%) | AUDPC | Disease Response | ||||
|---|---|---|---|---|---|---|---|---|---|
| Zero Time | 5 DPI | 10 DPI | 15 DPI | 20 DPI | |||||
| 1 | S. habrochaites (LA1777) | 0.0 | 1.1 | 1.9 | 2.3 | 3.3 e | 96.7 | 34.75 d | HR |
| 2 | S. habrochaites (LA2855) | 0.0 | 1.8 | 2.7 | 3.9 | 9.9 d | 90.1 | 66.75 c | R |
| 3 | S. habrochaites (LA1352) | 0.0 | 2.0 | 2.9 | 4.8 | 10.3 c | 89.7 | 74.25 c | R |
| 4 | S. pimpinellifolium (LA0375) | 0.0 | 28.3 | 64.8 | 79.7 | 95.3 b | 4.7 | 1102.3 b | HS |
| 5 | ‘Super Strain B’ (Commercial cultivar) | 0.0 | 37.1 | 67.9 | 88.4 | 100 a | 0.0 | 1217.0 a | HS |
| L.S.D at 0.05 | 2.973 | 10.418 | |||||||
| Name | RT | MF/MW |
|---|---|---|
| α-Terpinolene | 6.63 | C10H16/136.238 |
| β-Phellandrene | 7.01 | C10H16/136.238 |
| p-Menthane-1,2,3-triol | 10.80 | C10H20/188.264 |
| Flavone, 3,4′,5,7-tetramethoxy- | 11.18 | C19H18O6/342.347 |
| Quercetin 3′,4′,7-trimethyl ether | 11.70 | C18H16O7/344.319 |
| 7,8-Dihydro-α-ionone | 12.60 | C13H22O/194.318 |
| Caryophyllene | 12.70 | C15H24/204.357 |
| β-caryophyllene | 13.04 | C15H24/204.357 |
| Scopoletin | 13.38 | C10H8O4/192.17 |
| geranyl-α-terpinene | 14.16 | C20H32/272.47 |
| Santalol, cis, α- | 15.03 | C15H24O/220.356 |
| 9-cis-Retinoic acid | 15.22 | C20H28O2/300.44 |
| Nabilone | 15.33 | C24H36O3/372.549 |
| Ledene oxide | 15.46 | C15H24O/220.356 |
| Mesaconic acid | 15.87 | C5H6O4/130.099 |
| Longipinocarveol, trans- | 16.32 | C15H24O/220.356 |
| Geranyl isovalerate | 16.88 | C15H26O2/238.37 |
| (+)-α-Tocopherol | 17.01 | C29H50O2/430.71 |
| Isocurcumenol | 17.50 | C15H22O2/234.339 |
| α-Himachalene | 17.58 | C15H24/204.357 |
| 3-Hydroxy-4-methoxybenzaldehyde | 18.02 | C8H8O3/152.149 |
| 3-Octadecenoic acid, methyl ester | 18.03 | C19H36O2/296.495 |
| 3-Ethyl-5-(2′-ethylbutyl) octadecane | 18.45 | C26H54/366.718 |
| Butylated hydroxytoluene | 18.52 | C15H24O/220.36 |
| Cannabinol | 19.38 | C21H26O2/310.437 |
| Carmine acid | 19.90 | C22H20O13/492.389 |
| Elaidic acid | 20.01 | C18H34O2/282.468 |
| Phytol | 20.50 | C20H40O/296.539 |
| Farnesol | 21.17 | C15H26O/222.372 |
| Pentadecanoic acid | 22.15 | C15H30O2/242.403 |
| Glycitein | 22.77 | C16H12O5/284.267 |
| α-Terpinolene | 6.63 | C10H16/136.238 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arafa, R.A.; Kamel, S.M.; Taher, D.I.; Solberg, S.Ø.; Rakha, M.T. Leaf Extracts from Resistant Wild Tomato Can Be Used to Control Late Blight (Phytophthora infestans) in the Cultivated Tomato. Plants 2022, 11, 1824. https://doi.org/10.3390/plants11141824
Arafa RA, Kamel SM, Taher DI, Solberg SØ, Rakha MT. Leaf Extracts from Resistant Wild Tomato Can Be Used to Control Late Blight (Phytophthora infestans) in the Cultivated Tomato. Plants. 2022; 11(14):1824. https://doi.org/10.3390/plants11141824
Chicago/Turabian StyleArafa, Ramadan A., Said M. Kamel, Dalia I. Taher, Svein Ø. Solberg, and Mohamed T. Rakha. 2022. "Leaf Extracts from Resistant Wild Tomato Can Be Used to Control Late Blight (Phytophthora infestans) in the Cultivated Tomato" Plants 11, no. 14: 1824. https://doi.org/10.3390/plants11141824
APA StyleArafa, R. A., Kamel, S. M., Taher, D. I., Solberg, S. Ø., & Rakha, M. T. (2022). Leaf Extracts from Resistant Wild Tomato Can Be Used to Control Late Blight (Phytophthora infestans) in the Cultivated Tomato. Plants, 11(14), 1824. https://doi.org/10.3390/plants11141824