
Efficient In Vitro Sterilization and Propagation from Stem Segment Explants of Cnidoscolus aconitifolius (Mill.) I.M. Johnst, a Multipurpose Woody Plant

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material Preparation and Growth Conditions
2.2. Sterilization of Explants
2.2.1. Examining the Effect of Different Sterilization Times of 0.1% Mercuric Chloride
2.2.2. Examining the Effect of Different Concentrations of the Antibiotic PPM on Explant Sterilization
2.2.3. Examining the Effect of Different Pretreatment Methods on Explant Sterilization
2.2.4. Examining the Effect of Sampling Time on Explant Sterilization
2.3. Induction of Axillary Buds in Stem Segments
2.4. Induction of Adventitious Shoot
2.5. Induction of Adventitious Shoot Elongation
2.6. Induction of Adventitious Shoot Rooting
2.7. Plant Domestication and Transplanting
2.8. Statistical Analyses
3. Results
3.1. Effect of 0.1% Mercuric Chloride with Different Soaking Times on Explant Sterilization
3.2. Effect of Different Concentrations of the Antibiotic PPM on Explant Sterilization
3.3. Effect of Different Pretreatment Methods on Explant Sterilization
3.4. Effect of Sampling Season on Explant Sterilization
3.5. Effect of Medium Types and PGRs on Axillary Bud Induction
3.6. Effects of PGRs on Adventitious Shoot Induction and Proliferation
3.7. Effect of PGRs on Adventitious Shoot Elongation
3.8. Adventitious Shoot Rooting
3.9. Plant Domestication and Transplanting
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MS | Murashige and Skoog |
| 1/2 MS | Half-strength Murashige and Skoog |
| PPM | Plant Preservative Mixture |
| 6-BA | 6-benzyladenine |
| IBA | Indole-3-butyric acid |
| IAA | Indole-3-acetic acid |
| NAA | α-Naphthalene acetic acid |
| PGRs | Plant growth regulators |
References
- Orji, O.U.; Ibiam, U.A.; Aja, P.M.; Ugwu, P.; Uraku, A.J.; Aloke, C.; Nwali, B.U. Evaluation of the phytochemical and nutritional profiles of Cnidoscolus aconitifolius leaf collected in Abakaliki South East Nigeria. World J. Med. Sci. 2016, 13, 213–217. [Google Scholar]
- Amaya, N.; Padulosi, S.; Meldrum, G. Value chain analysis of chaya (Mayan spinach) in Guatemala. Econ. Bot. 2020, 74, 100–114. [Google Scholar] [CrossRef] [Green Version]
- Munguía-Rosas, M.A. Artificial selection optimizes clonality in chaya (Cnidoscolus aconitifolius). Sci. Rep. 2021, 11, 21017. [Google Scholar] [CrossRef]
- Kuti, J.O.; Konuru, H.B. Antioxidant capacity and phenolic content in leaf extracts of tree spinach (Cnidoscolus spp.). J. Agric. Food Chem. 2004, 52, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Aguilar, D.M.; Grusak, M.A. Evaluation of minerals, phytochemical compounds and antioxidant activity of Mexican, Central American, and African green leafy vegetables. Plant Foods Hum. Nutr. 2015, 70, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Lyons, G.; Dean, G.; Tongaiaba, R.; Halavatau, S.; Nakabuta, K.; Lonalona, M.; Susumu, G. Macro-and micronutrients from traditional food plants could improve nutrition and reduce non-communicable diseases of islanders on atolls in the South Pacific. Plants 2020, 9, 942. [Google Scholar] [CrossRef] [PubMed]
- Totakul, P.; Viennasay, B.; Sommai, S.; Matra, M.; Infascelli, F.; Wanapat, M. Supplemental effect of Chaya (Cnidoscolus aconitifolius) leaf pellet on rumen fermentation, nutrients digestibility and microbial protein synthesis in growing crossbred bulls. Ital. J. Anim. Sci. 2021, 20, 279–287. [Google Scholar] [CrossRef]
- Godinez-Santillan, R.I.; Chavez-Servin, J.L.; Garcia-Gasca, T.; Guzman-Maldonado, S.H. Phenolic characterization and antioxidant capacity of alcoholic extracts from raw and boiled leaves of Cnidoscolus aconitifolius (Euphorbiaceae). Acta. Bot. Mex. 2019, 126, e1493. [Google Scholar] [CrossRef] [Green Version]
- Guzman, E.L.; Castaneda, C.T.B.; Ocanas, L.G.; Pescador, M.G.N.; Cruz, F.J.M. Evaluation of antioxidant and protective effect against genotoxic damage of two extracts of chaya leaves (Cnidoscolus aconitifolius). J. Med. Plants Res. 2020, 14, 555–558. [Google Scholar]
- Adeniran, O.I.; Olajide, O.O.; Igwemmar, N.C.; Orishadipe, A.T. Phytochemical constituents, antimicrobial and antioxidant potentials of tree spinach [Cnidoscolus aconitifolius (Miller) IM Johnston]. J. Med. Plants Res. 2013, 7, 1310–1316. [Google Scholar]
- Iyke, W.I.; Nwafor, A.; Njoku, B.; Nwoke, K.U.; Deebii, N. Investigation of phytocomponents and hypoglycaemic effect of hydro-methanolic leaf extract of Cnidoscolus aconitifolius (Spinach Tree) in streptozotocin induced-diabetic wistar rats. J. Complement. Altern. Med. Res. 2018, 5, 1–9. [Google Scholar] [CrossRef]
- Oyagbemi, A.A.; Odetola, A.A.; Azeez, O.I. Ameliorative effects of Cnidoscolus aconitifolius on anaemia and osmotic fragility induced by proteinenergy malnutrition. Afr. J. Biotechnol. 2008, 7, 1721–1726. [Google Scholar] [CrossRef] [Green Version]
- Olivia, N.U.; Happiness, U.C.; Obinna, O.M. Protective effect of Cnidoscolus aconitifolius leaves against diclofenacinduced gastric mucosal damage. Pak. J. Pharm. Sci. 2020, 33, 651–657. [Google Scholar]
- Kuri-Garcia, A.; Godinez-Santillan, R.I.; Mejia, C.; Ferriz-Martínez, R.A.; Garcia-Solis, P.; Enríquez-Vázquez, A.; Chavez-Servin, J.L. Preventive effect of an infusion of the aqueous extract of Chaya leaves (Cnidoscolus aconitifolius) in an aberrant crypt foci rat model induced by azoxymethane and dextran sulfate sodium. J. Med. Food 2019, 22, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento-Franco, L.; McNab, J.M.; Pearson, R.A.; Belmar-Casso, R. Performance of broilers fed on diets containing different amounts of chaya (Cnidoscolus aconitifolius) leaf meal. Trop. Anim. Health Prod. 2002, 34, 257–269. [Google Scholar] [CrossRef]
- Sarmiento-Franco, L.; McNab, J.M.; Pearson, A.; Belmar-Casso, R. The effect of chaya (Cnidoscolus aconitifolius) leaf meal and of exogenous enzymes on amino acid digestibility in broilers. Br. Poult. Sci. 2003, 44, 458–463. [Google Scholar] [CrossRef]
- Herdian, H.; Sofyan, A.; Sakti, A.A.; Karimy, M.F.; Fitriana, E.L.; Laconi, E.B. Effects of spinach tree leaves and high concentrates diets supplemented with micro minerals on in vitro rumen fermentation profiles. IOP Conf. Ser. Earth Environ. Sci. 2020, 462, 012004. [Google Scholar] [CrossRef]
- Donkoh, A.; Atuahene, C.C.; Poku-Prempeh, Y.B.; Twum, I.G. The nutritive value of chaya leaf meal (Cnidoscolus aconitifolius (Mill.) Johnston): Studies with broiler chickens. Anim. Feed Sci. Technol. 1999, 77, 163–172. [Google Scholar] [CrossRef]
- Kuri-GarcÃa, A.; GuzmÃ, S.H. Phenolic profile and antioxidant capacity of Cnidoscolus chayamansa and Cnidoscolus aconitifolius: A review. J. Med. Plants Res. 2017, 11, 713–727. [Google Scholar]
- Munguia-Rosas, M.A.; Jacome-Flores, M.E.; Bello-Bedoy, R.; Solís-Montero, V.; Ochoa-Estrada, E. Morphological divergence between wild and cultivated chaya (Cnidoscolus aconitifolius) (Mill.) IM Johnst. Genet. Resour. Crop Evol. 2019, 66, 1389–1398. [Google Scholar] [CrossRef]
- Isah, T.; Umar, S.; Mujib, A.; Sharma, M.P.; Rajasekharan, P.E.; Zafar, N.; Frukh, A. Secondary metabolism of pharmaceuticals in the plant in vitro cultures: Strategies, approaches, and limitations to achieving higher yield. Plant Cell Tissue Organ Cult. 2017, 132, 239–265. [Google Scholar] [CrossRef]
- Perez-Gonzalez, M.; Nieto-Trujillo, A.; Gutierrez-Rebolledo, G.A.; Garcia-Martinez, I.; Estrada-Zuniga, M.E.; Bernabe-Antonio, A.; Cruz-Sosa, F. Lupeol acetate production and antioxidant activity of a cell suspension culture from Cnidoscolus chayamansa leaves. S. Afr. J. Bot. 2019, 125, 30–38. [Google Scholar] [CrossRef]
- Bednarek, P.T.; Orłowska, R. Plant tissue culture environment as a switch-key of (epi) genetic changes. Plant Cell Tissue Organ Cult. 2020, 140, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Gaur, A.; Kumar, P.; Thakur, A.K.; Srivastava, D.K. In vitro plant regeneration studies and their potential applications in Populus spp.: A review. Isr. J. Plant Sci. 2016, 63, 77–84. [Google Scholar] [CrossRef]
- Hou, J.; Mao, Y.; Su, P.; Wang, D.; Chen, X.; Huang, S.; Wu, L. A high throughput plant regeneration system from shoot stems of Sapium sebiferum Roxb., a potential multi-purpose bioenergy tree. Ind. Crops Prod. 2020, 154, 112653. [Google Scholar] [CrossRef]
- Loyola-Vargas, V.M.; Ochoa-Alejo, N. An introduction to plant tissue culture: Advances and perspectives. Plant Cell Cult. Protoc. 2018, 1815, 3–13. [Google Scholar]
- Taskin, H.; Baktemur, G.; Kurul, M.; Büyükalaca, S. Use of tissue culture techniques for producing virus-free plant in garlic and their identification through real-time PCR. Sci. World J. 2013, 2013, 781282. [Google Scholar] [CrossRef] [Green Version]
- Kamal, O.M.; Shah, S.H.A.; Li, Y.; Hou, X.; Li, Y. Production of ascorbic acid, total protein, callus and root in vitro of non-heading Chinese cabbage by tissue culture. Mol. Biol. Rep. 2020, 47, 6887–6897. [Google Scholar] [CrossRef]
- Ticha, M.; Illesova, P.; Hrbackova, M.; Basheer, J.; Novak, D.; Hlavackova, K.; Samaj, J. Tissue culture, genetic transformation, interaction with beneficial microbes, and modern bio-imaging techniques in alfalfa research. Crit. Rev. Biotechnol. 2020, 40, 1265–1280. [Google Scholar] [CrossRef]
- Kang, L.; Zheng, K.; Xie, Y.; Deng, Y.; Yu, Y.; Zhu, M.; Deng, X. Efficient tissue culture protocol for Magnolia lucida (Magnoliaceae) and confirmation of genetic stability of the regenerated plants. Plants 2020, 9, 997. [Google Scholar] [CrossRef]
- Toppo, R.; Beura, S. Effect of surface sterilization time on leaf explants for aseptic culture in Anthurium adraeanum (Hort.) cv. Fire. Int. J. Curr. Microbiol. App. Sci. 2018, 7, 2509–2515. [Google Scholar] [CrossRef]
- Bhadrawale, D.; Mishra, J.P.; Mishra, Y. An improvised in vitro vegetative propagation technique for Bambusa tulda: Influence of season, sterilization and hormones. J. For. Res. 2018, 29, 1069–1074. [Google Scholar] [CrossRef]
- Rafiq, S.; Rather, Z.A.; Bhat, R.A.; Nazki, I.T.; Al-Harbi, M.S.; Banday, N.; Andrabi, N. Standardization of in vitro micropropagation procedure of Oriental Lilium Hybrid Cv.‘Ravenna’. Saudi J. Biol. Sci. 2021, 28, 7581–7587. [Google Scholar] [CrossRef] [PubMed]
- Poobathy, R.; Zakaria, R.; Murugaiyah, V.; Subramaniam, S. Surface sterilization and micropropagation of Ludisia discolor. Biocatal. Agric. Biotechnol. 2019, 22, 101380. [Google Scholar] [CrossRef]
- Li, X.; Zhang, F.; He, N.; Zhang, B.; Liu, X.; Tan, X. In vitro influence of Fe3O4 nanoparticles on Rhizoma polygonati micropropagation. J. Nanosci. Nanotechnol. 2017, 17, 2047–2053. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, W.; Saha, M.C.; Udvardi, M.K.; Kang, Y. Improved node culture methods for rapid vegetative propagation of switchgrass (Panicum virgatum L.). BMC Plant Biol. 2021, 21, 128. [Google Scholar] [CrossRef]
- Niedz, R.P. Using isothiazolone biocides to control microbial and fungal contaminants in plant tissue cultures. Hort. Technol. 1998, 8, 598–601. [Google Scholar] [CrossRef]
- Torres, G.R.C.; Houllou, L.M.; de Souza, R.A. Control of contaminants during introduction and establishment of Bambusa vulgaris in vitro. Res. Biotechnol. 2016, 7, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Silveira, S.S.; Cordeiro-Silva, R.; Degenhardt-Goldbach, J.; Quoirin, M. Micropropagation of Calophyllum brasiliense (Cambess.) from nodal segments. Braz. J. Biol. 2016, 76, 656–663. [Google Scholar] [CrossRef] [Green Version]
- Rihan, H.Z.; Al-Issawi, M.; Al-Swedi, F.; Fuller, M.P. The effect of using PPM (plant preservative mixture) on the development of cauliflower microshoots and the quality of artificial seed produced. Sci. Hortic. 2012, 141, 47–52. [Google Scholar] [CrossRef]
- Orlikowska, T.; Zawadzka, M.; Zenkteler, E.; Sobiczewski, P. Influence of the biocides PPMTM and Vitrofural on bacteria isolated from contaminated plant tissue cultures and on plant microshoots grown on various media. J. Hortic. Sci. Biotechnol. 2012, 87, 223–230. [Google Scholar] [CrossRef]
- Miyazaki, J.; Tan, B.H.; Errington, S.G. Eradication of endophytic bacteria via treatment for axillary buds of Petunia hybrida using Plant Preservative Mixture (PPMTM). Plant Cell Tissue Organ Cult. 2010, 102, 365–372. [Google Scholar] [CrossRef]
- Huang, L.; Wang, H.; Shahid, M.; Zeng, B. Chlorothalonil: An effective bacterio-static agent for bud induction of Acacia auriculiformis under open condition (non-axenic). Plant Methods 2019, 15, 5. [Google Scholar] [CrossRef]
- da Silva, J.A.T.; Winarto, B.; Dobránszki, J.; Cardoso, J.C.; Zeng, S. Tissue disinfection for preparation of culture. Folia. Hortic. 2016, 28, 57–75. [Google Scholar] [CrossRef] [Green Version]
- Ho, W.J.; Huang, Y.K.; Huang, W.W.; Huang, Y.C.; Chung, J.P. Effective in vitro culture using dormant bud of nodal sections from a mature Acacia tree. Vitr. Cell. Dev. Biol.-Plant 2021, 58, 437–466. [Google Scholar] [CrossRef]
- Zahid, N.A.; Jaafar, H.Z.; Hakiman, M. Micropropagation of ginger (Zingiber officinale Roscoe)‘Bentong’and evaluation of its secondary metabolites and antioxidant activities compared with the conventionally propagated plant. Plants 2021, 10, 630. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Zou, J.; Zhang, B.; Que, Q.; Zhang, J.; Chen, X.; Zhou, W. An efficient in vitro propagation protocol for direct organogenesis from root explants of a multi-purpose plant, Broussonetia papyrifera (L.) L’Hér. ex Vent. Ind. Crops Prod. 2021, 170, 113686. [Google Scholar] [CrossRef]
- Lijalem, T.; Feyissa, T. In vitro propagation of Securidaca longipedunculata (Fresen) from shoot tip: An endangered medicinal plant. J. Genet. Eng. Biotechnol. 2020, 18, 3. [Google Scholar] [CrossRef] [Green Version]
- Mao, W.; Song, H.; Li, Y.; Wang, Y.; Lin, H.; Yao, C.; Li, P. Efficient plant regeneration and genetic transformation system of the precious fast-growing tree Toona ciliata. Ind. Crops Prod. 2021, 172, 114015. [Google Scholar] [CrossRef]
- Ioannidis, K.; Dadiotis, E.; Mitsis, V.; Melliou, E.; Magiatis, P. Biotechnological approaches on two high CBD and CBG Cannabis sativa L. (Cannabaceae) varieties: In vitro regeneration and phytochemical consistency evaluation of micropropagated plants using quantitative 1H-NMR. Molecules 2020, 25, 5928. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
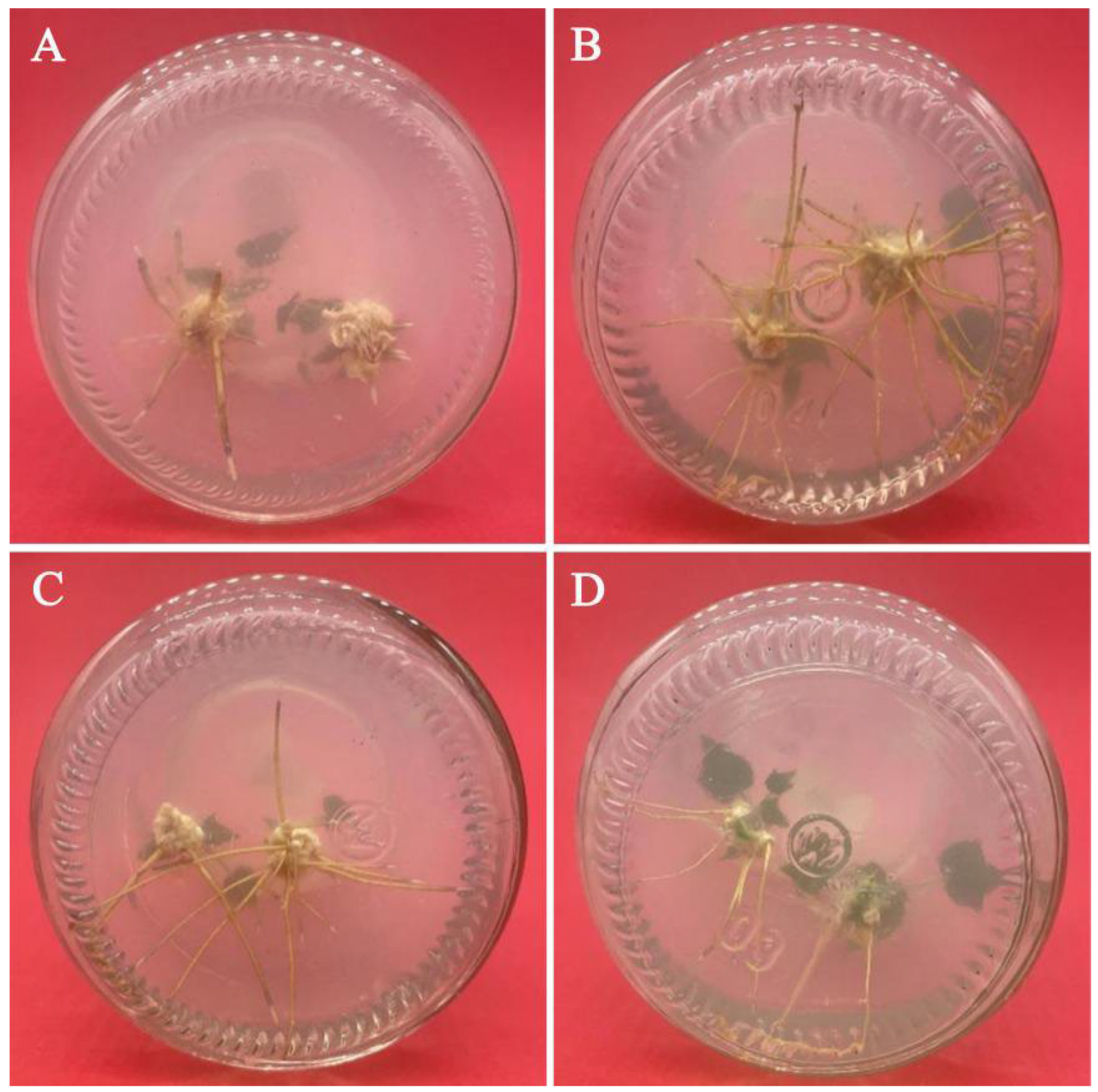{kind=link}
| 0.1% Mercuric Chloride (min) | Contamination (%) | Death (%) | Survival (%) |
|---|---|---|---|
| 5 | 74.44 ± 5.09 a | 0.00 ± 0.00 b | 25.56 ± 5.09 c |
| 8 | 65.56 ± 1.93 b | 0.00 ± 0.00 b | 34.44 ± 1.93 b |
| 10 | 54.44 ± 3.85 c | 0.00 ± 1.92 b | 44.44 ± 5.09 a |
| 12 | 52.22 ± 5.09 c | 6.67 ± 0.00 a | 41.11 ± 5.09 a |
| 15 | 48.89 ± 5.09 c | 8.89 ± 3.85 a | 42.22 ± 1.92 a |
| PPM (ml/L) | Contamination (%) | Death (%) | Survival (%) |
|---|---|---|---|
| 0 | 55.55 ± 3.85 a | 1.11 ± 1.92 a | 43.33 ± 3.34 d |
| 1 | 44.44 ± 1.93 b | 0.00 ± 0.00 a | 55.56 ± 1.93 c |
| 2 | 36.67 ± 3.34 c | 0.00 ± 0.00 a | 63.33 ± 3.34 b |
| 3 | 28.89 ± 3.85 d | 1.11 ± 1.92 a | 70.00 ± 3.34 a |
| 4 | 26.67 ± 0.00 d | 1.11 ± 1.92 a | 72.22 ± 1.93 a |
| Pretreatment Methods | Contamination (%) | Death (%) | Survival (%) |
|---|---|---|---|
| Running water rinsing for 0 min | 52.22 ± 1.92 a | 0.00 ± 0.00 b | 47.78 ± 1.92 e |
| Running water rinsing for 30 min | 45.56 ± 1.92 b | 0.00 ± 0.00 b | 54.44 ± 1.92 cd |
| Running water rinsing for 60 min | 38.89 ± 1.92 c | 0.00 ± 0.00 b | 61.11 ± 1.92 b |
| Running water rinsing for 120 min | 26.67 ± 0.00 d | 1.11 ± 1.92 b | 72.22 ± 1.92 a |
| Running water rinsing for 150 min | 24.44 ± 1.92 d | 2.22 ± 1.92 b | 73.33 ± 3.33 a |
| 0.1% Carbendazim soaking for 5 min | 41.11 ± 1.92 c | 0.00 ± 0.00 b | 58.89 ± 1.92 bc |
| 0.1% Carbendazim soaking for 10 min | 47.78 ± 1.92 b | 0.00 ± 0.00 b | 52.22 ± 1.92 de |
| 0.1% Carbendazim soaking for 20 min | 47.78 ± 1.92 b | 1.11 ± 1.92 b | 51.11 ± 1.92 de |
| 0.1% Carbendazim soaking for 30 min | 38.89 ± 1.92 c | 10.00 ± 5.77 a | 51.11 ± 6.94 de |
| Sampling Season | Contamination (%) | Death (%) | Survival (%) |
|---|---|---|---|
| March–May | 25.56 ± 1.92 c | 1.11 ± 1.92 a | 73.33 ± 3.33 a |
| June–August | 27.78 ± 1.92 bc | 2.22 ± 1.92 a | 70.00 ± 3.33 ab |
| September–November | 31.11 ± 1.92 ab | 1.11 ± 1.92 a | 67.78 ± 3.85 ab |
| December–February | 32.22 ± 1.92 a | 3.33 ± 0.00 a | 64.44 ± 1.92 b |
| Medium | 6-BA (mg/L) | Frequency of Axillary Bud Induction (%) | Mean Length of Axillary Buds (cm) |
|---|---|---|---|
| MS | 0 | 88.89 ± 1.92 c | 1.56 ± 0.45 b |
| MS | 0.5 | 92.22 ± 1.92 ab | 1.62 ± 0.71 b |
| MS | 1.0 | 88.89 ± 1.92 c | 1.34 ± 0.39 c |
| 1/2 MS | 0 | 90.00 ± 0.00 bc | 1.61 ± 0.38 b |
| 1/2 MS | 0.5 | 93.33 ± 0.00 a | 2.47 ± 0.67 a |
| 1/2 MS | 1.0 | 87.78 ± 1.92 cd | 1.35 ± 0.48 c |
| B5 | 0 | 88.89 ± 1.92 c | 1.48 ± 0.33 bc |
| B5 | 0.5 | 88.89 ± 1.92 c | 1.59 ± 0.39 b |
| B5 | 1.0 | 85.56 ± 1.92 d | 1.33 ± 0.31 c |
| F value | |||
| Medium | 4.20 * | 37.24 *** | |
| 6-BA | 21.60 *** | 82.17 *** | |
| Medium × 6-BA | 2.40 ns | 26.27 *** | |
| 6-BA (mg/L) | MS | 1/2 MS | B5 | |
|---|---|---|---|---|
| Medium | ||||
| 0 | 1.56 ± 0.45 a (AB) | 1.61 ± 0.38 b (A) | 1.48 ± 0.33 a (A) | |
| 0.5 | 1.62 ± 0.71 a (B) | 2.47 ± 0.67 a (A) | 1.59 ± 0.39 a (B) | |
| 1.0 | 1.34 ± 0.39 b (A) | 1.35 ±0.48 c (A) | 1.33 ± 0.31 b (A) | |
| F value | ||||
| Medium: 37.24 *** 6-BA: 82.17 *** Medium × 6-BA: 26.27 *** | ||||
| 6-BA (mg/L) | IBA (mg/L) | Frequency of Shoot Induction (%) | Net Proliferation Coefficient | Total Proliferation Coefficient |
|---|---|---|---|---|
| 1.0 | 0.1 | 22.22 ± 3.85 f | 2.70 ± 0.67 e | 0.60 ± 1.18 d |
| 1.0 | 0.2 | 42.22 ± 3.85 cd | 3.42 ± 0.96 cde | 1.44 ± 1.82 cd |
| 1.0 | 0.5 | 31.11 ± 3.85 e | 2.79 ± 0.80 e | 0.87 ± 1.38 cd |
| 2.0 | 0.1 | 28.89 ± 3.85 ef | 3.00 ± 0.91 de | 0.87 ± 1.45 cd |
| 2.0 | 0.2 | 44.44 ± 3.85 c | 3.90 ± 1.17 bc | 1.73 ± 2.10 bc |
| 2.0 | 0.5 | 35.56 ± 3.85 de | 3.63 ± 0.96 bcd | 1.29 ± 1.84 cd |
| 4.0 | 0.1 | 42.22 ± 3.85 cd | 4.00 ± 0.82 bc | 1.69 ± 2.07 bc |
| 4.0 | 0.2 | 80.00 ± 6.67 a | 5.69 ± 1.45 a | 4.56 ± 2.64 a |
| 4.0 | 0.5 | 57.78 ± 3.85 b | 4.35 ± 1.20 b | 2.69 ± 2.35 b |
| 5.0 | 0.1 | 28.89 ± 3.85 ef | 3.46 ± 0.52 cde | 0.87 ± 1.47 cd |
| 5.0 | 0.2 | 44.44 ± 3.85 c | 3.85 ± 0.75 bc | 1.73 ± 1.94 bc |
| 5.0 | 0.5 | 35.56 ± 3.85 de | 3.69 ± 0.79 bcd | 1.20 ± 1.78 cd |
| F value | ||||
| 6-BA | 84.86 *** | 29.55 *** | 28.62 *** | |
| IBA | 86.36 *** | 23.48 *** | 13.03 *** | |
| 6-BA × IBA | 4.93 ** | 3.61 ** | 2.58 * | |
| 6-BA (mg/L) | 1.0 | 2.0 | 4.0 | 5.0 | F Value | ||
|---|---|---|---|---|---|---|---|
| IBA (mg/L) | |||||||
| 0.1 | Frequency of shoot induction (%) | 22.22 ± 3.85 c (B) | 28.89 ± 3.85 b (B) | 42.22 ± 3.85 c (A) | 28.89 ± 3.85 b (B) | 6-BA: 84.86 *** | |
| 0.2 | 42.22 ± 3.85 a (B) | 44.44 ± 3.85 a (B) | 80.00 ± 6.67 a (A) | 44.44 ± 3.85 a (B) | IBA: 86.36 *** | ||
| 0.5 | 31.11 ± 3.85 b (B) | 35.56 ± 3.85 b (B) | 57.78 ± 3.85 b (A) | 35.56 ± 3.85 b (B) | 6-BA × IBA: 4.93 ** | ||
| 0.1 | Net proliferation coefficient | 2.70 ± 0.67 b (C) | 3.00 ± 0.91 b (BC) | 4.00 ± 0.82 b (A) | 3.46 ± 0.52 a (AB) | 6-BA: 29.55 *** | |
| 0.2 | 3.42 ± 0.96 a (B) | 3.90 ± 1.17 a (B) | 5.69 ± 1.45 a (A) | 3.85 ± 0.75 a (B) | IBA: 23.48 *** | ||
| 0.5 | 2.79 ± 0.80 b (C) | 3.63 ± 0.96 ab (B) | 4.35 ± 1.20 b (A) | 3.69 ± 0.79 a (B) | 6-BA × IBA: 3.61 ** | ||
| 0.1 | Total proliferation coefficient | 0.60 ± 1.18 b (B) | 0.87 ± 1.45 b (B) | 1.69 ± 2.07 b (A) | 0.87 ± 1.47 b (B) | 6-BA: 28.62 *** | |
| 0.2 | 1.44 ± 1.82 a (B) | 1.73 ± 2.10 a (B) | 4.56 ± 2.64 a (A) | 1.73 ± 1.94 a (B) | IBA: 13.03 *** | ||
| 0.5 | 0.87 ± 1.38 ab (B) | 1.29 ± 1.84 ab (B) | 2.69 ± 2.35 b (A) | 1.20 ±1.78 ab (B) | 6-BA × IBA: 2.58 * | ||
| 6-BA (mg/L) | IAA (mg/L) | Mean Length of Adventitious Shoots (cm) |
|---|---|---|
| 0 | 1.0 | 1.38 ± 0.25 d |
| 0 | 1.5 | 1.63 ± 0.23 bc |
| 0 | 2.0 | 1.57 ± 0.23 c |
| 0.1 | 1.0 | 1.47 ± 0.22 d |
| 0.1 | 1.5 | 1.93 ± 0.19 a |
| 0.1 | 2.0 | 1.68 ± 0.22 bc |
| 0.2 | 1.0 | 1.42 ± 0.25 d |
| 0.2 | 1.5 | 1.69 ± 0.24 b |
| 0.2 | 2.0 | 1.62 ± 0.22 bc |
| F value | ||
| 6-BA | 16.34 *** | |
| IAA | 62.42 *** | |
| 6-BA × IAA | 2.97 * | |
| IAA (mg/L) | 0 | 0.1 | 0.2 | |
|---|---|---|---|---|
| 6-BA (mg/L) | ||||
| 1.0 | 1.38 ± 0.25 b (A) | 1.47 ± 0.22 c (A) | 1.42 ± 0.25 b (A) | |
| 1.5 | 1.63 ± 0.23 a (B) | 1.93 ± 0.19 a (A) | 1.69 ± 0.24 a (B) | |
| 2.0 | 1.57 ± 0.23 a (B) | 1.68 ± 0.22 b (A) | 1.62 ± 0.22 a (AB) | |
| F value | ||||
| 6-BA: 16.34 *** IAA: 62.42 *** 6-BA × IAA: 2.97 * | ||||
| NAA (mg/L) | Rooting Percentage (%) | Mean number of Roots Per Adventitious Shoot | Mean Length of Roots (cm) |
|---|---|---|---|
| 0 | 42.22 ± 3.85 c | 2.47 ± 0.96 c | 1.72 ± 0.55 d |
| 0.1 | 82.00 ± 3.85 a | 4.84 ± 1.14 a | 3.91 ± 0.95 a |
| 0.2 | 73.33 ± 6.67 b | 4.45 ± 0.98 a | 3.52 ± 0.71 b |
| 0.5 | 48.89 ± 3.85 c | 3.63 ± 0.90 b | 3.11 ± 0.88 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, M.; Li, Y.; Jiang, H.; Zhang, S.; Que, Q.; Chen, X.; Zhou, W. Efficient In Vitro Sterilization and Propagation from Stem Segment Explants of Cnidoscolus aconitifolius (Mill.) I.M. Johnst, a Multipurpose Woody Plant. Plants 2022, 11, 1937. https://doi.org/10.3390/plants11151937
Gu M, Li Y, Jiang H, Zhang S, Que Q, Chen X, Zhou W. Efficient In Vitro Sterilization and Propagation from Stem Segment Explants of Cnidoscolus aconitifolius (Mill.) I.M. Johnst, a Multipurpose Woody Plant. Plants. 2022; 11(15):1937. https://doi.org/10.3390/plants11151937
Chicago/Turabian StyleGu, Min, Youli Li, Huier Jiang, Shihu Zhang, Qingmin Que, Xiaoyang Chen, and Wei Zhou. 2022. "Efficient In Vitro Sterilization and Propagation from Stem Segment Explants of Cnidoscolus aconitifolius (Mill.) I.M. Johnst, a Multipurpose Woody Plant" Plants 11, no. 15: 1937. https://doi.org/10.3390/plants11151937
APA StyleGu, M., Li, Y., Jiang, H., Zhang, S., Que, Q., Chen, X., & Zhou, W. (2022). Efficient In Vitro Sterilization and Propagation from Stem Segment Explants of Cnidoscolus aconitifolius (Mill.) I.M. Johnst, a Multipurpose Woody Plant. Plants, 11(15), 1937. https://doi.org/10.3390/plants11151937