
Enclosure in Combination with Mowing Simultaneously Promoted Grassland Biodiversity and Biomass Productivity




Abstract
:
1. Introduction
2. Results
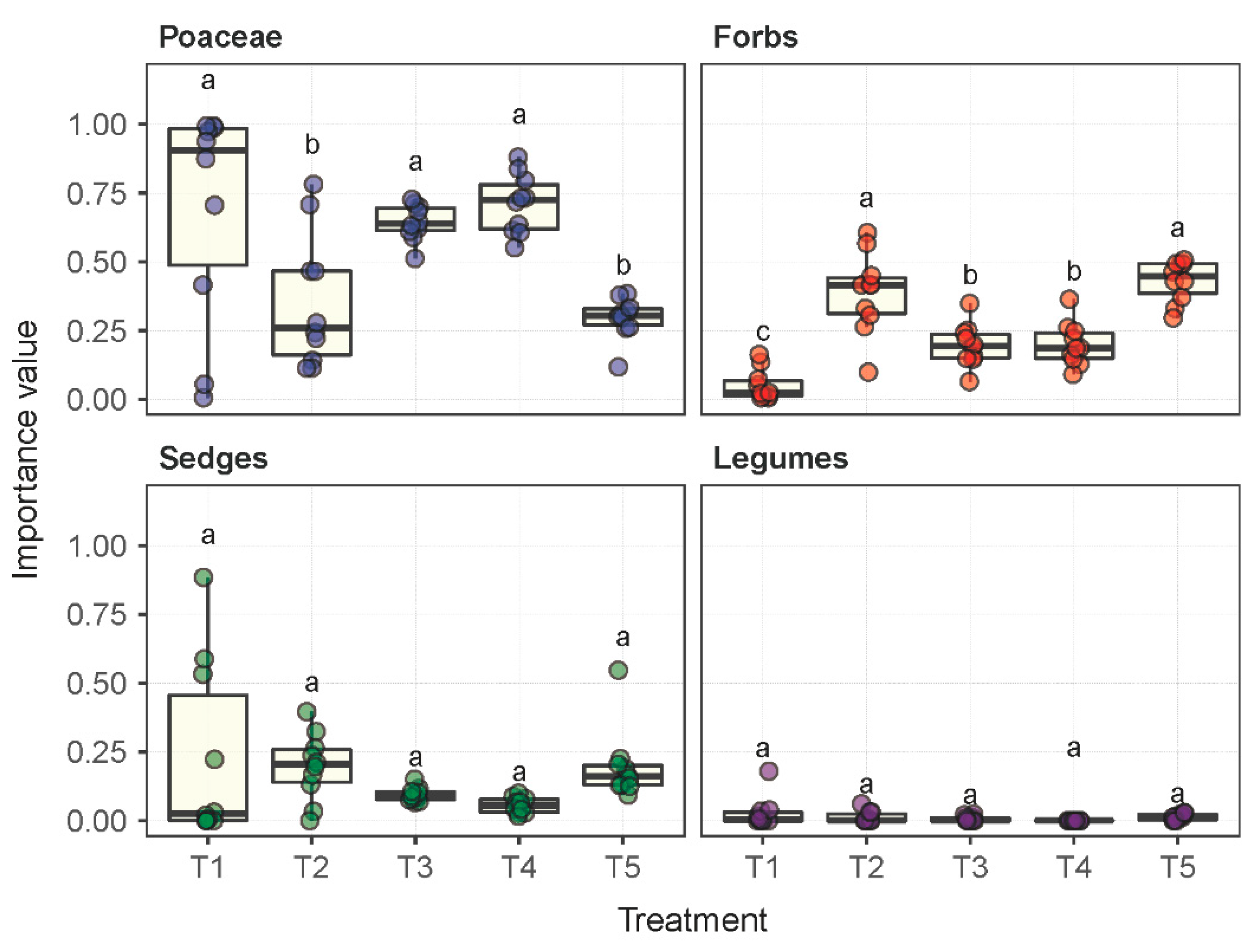
2.1. Plant Community Composition
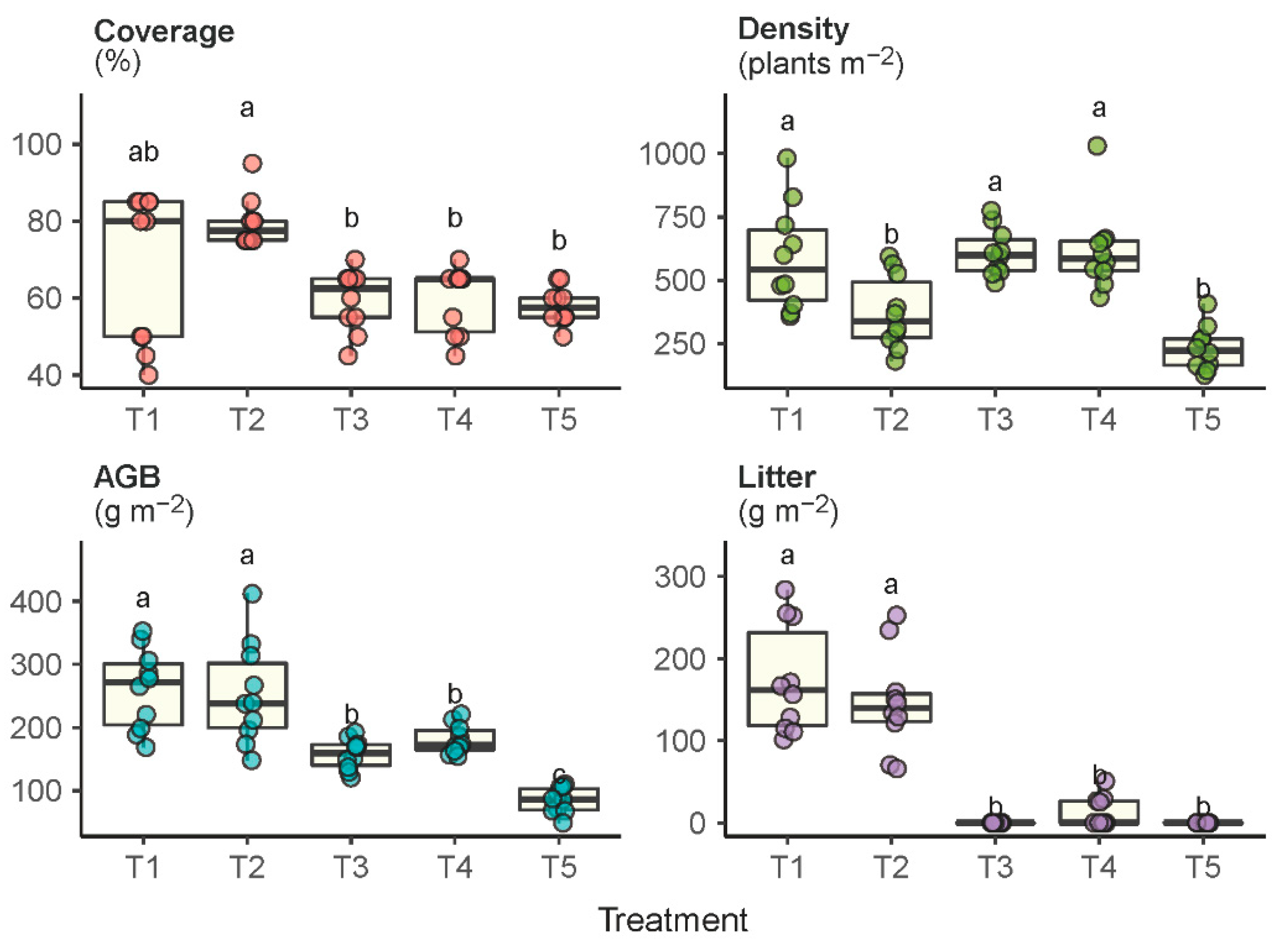
2.2. Vegetation Coverage, Density and Growth
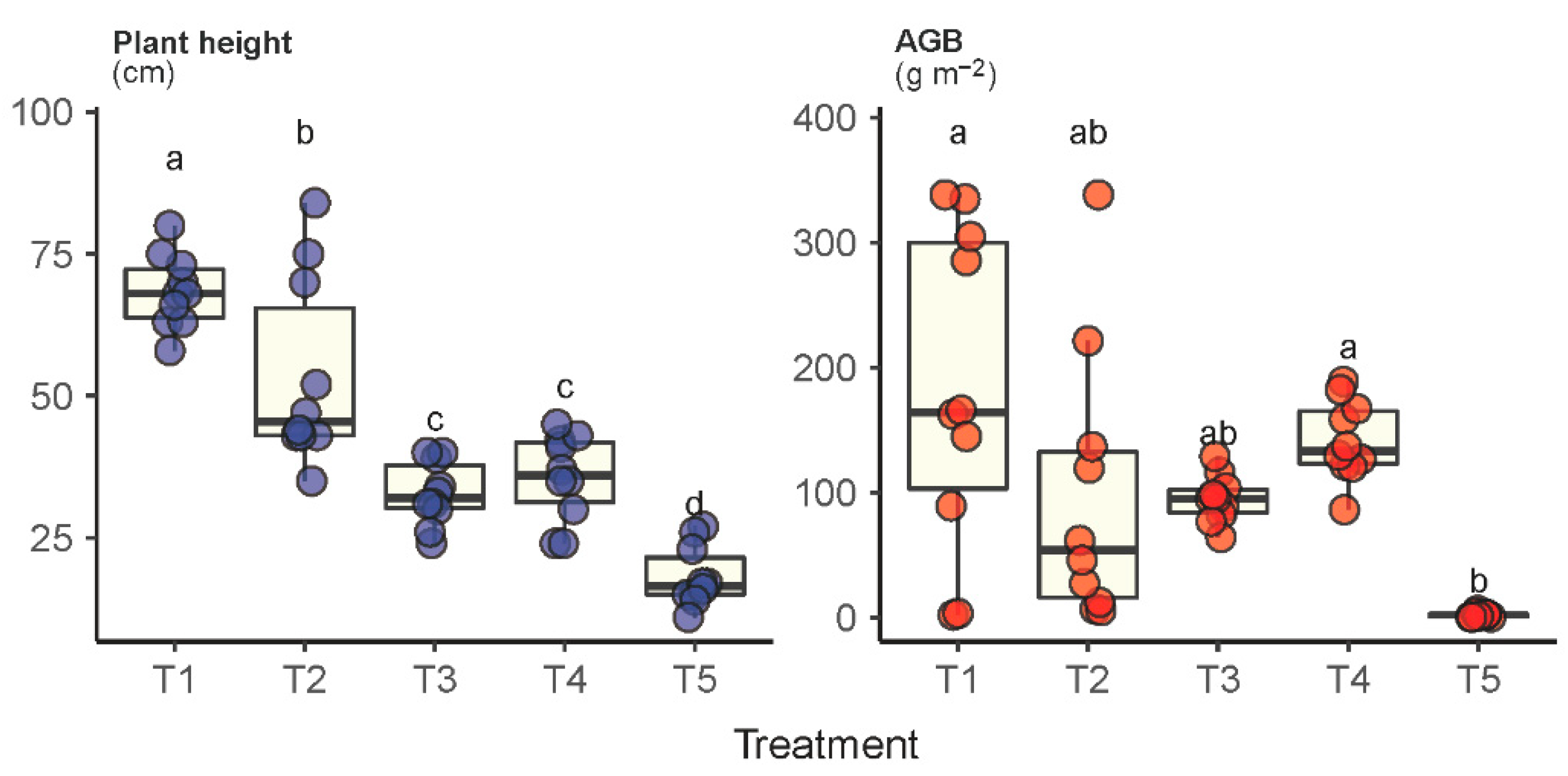
2.3. Plant Height and AGB of the Dominant Species
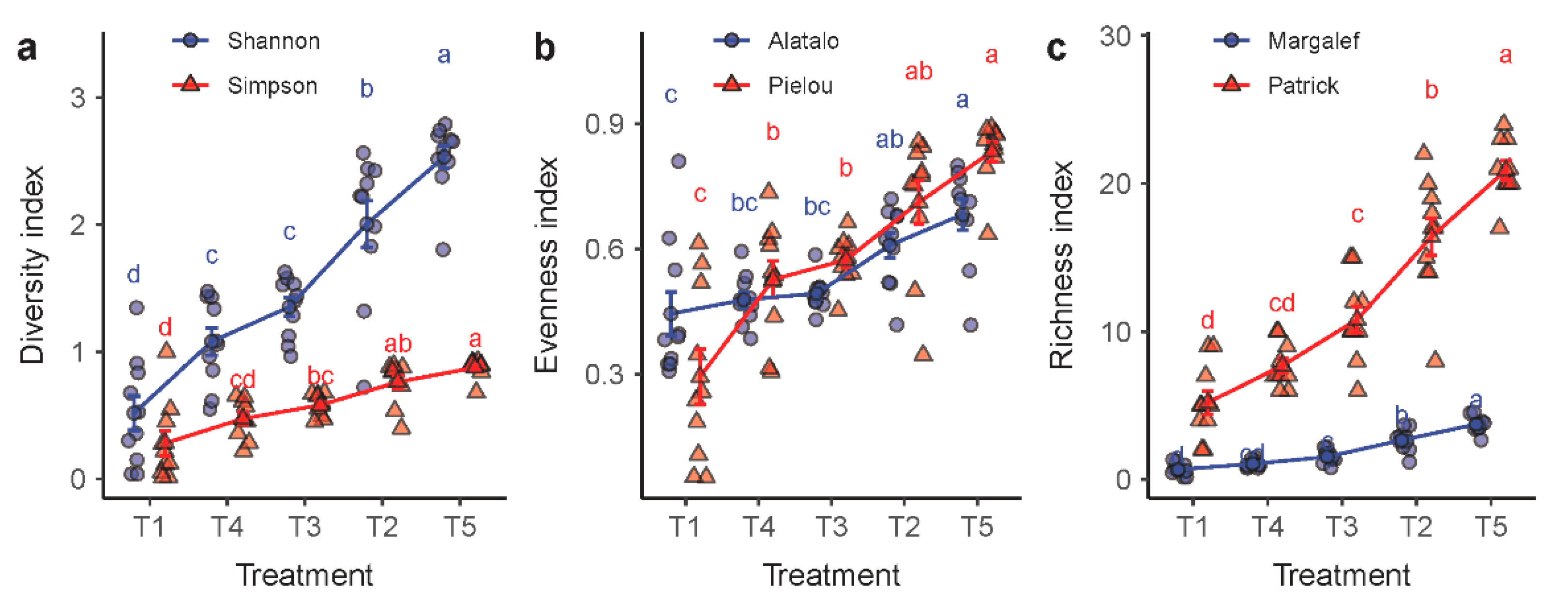
2.4. Species Diversity
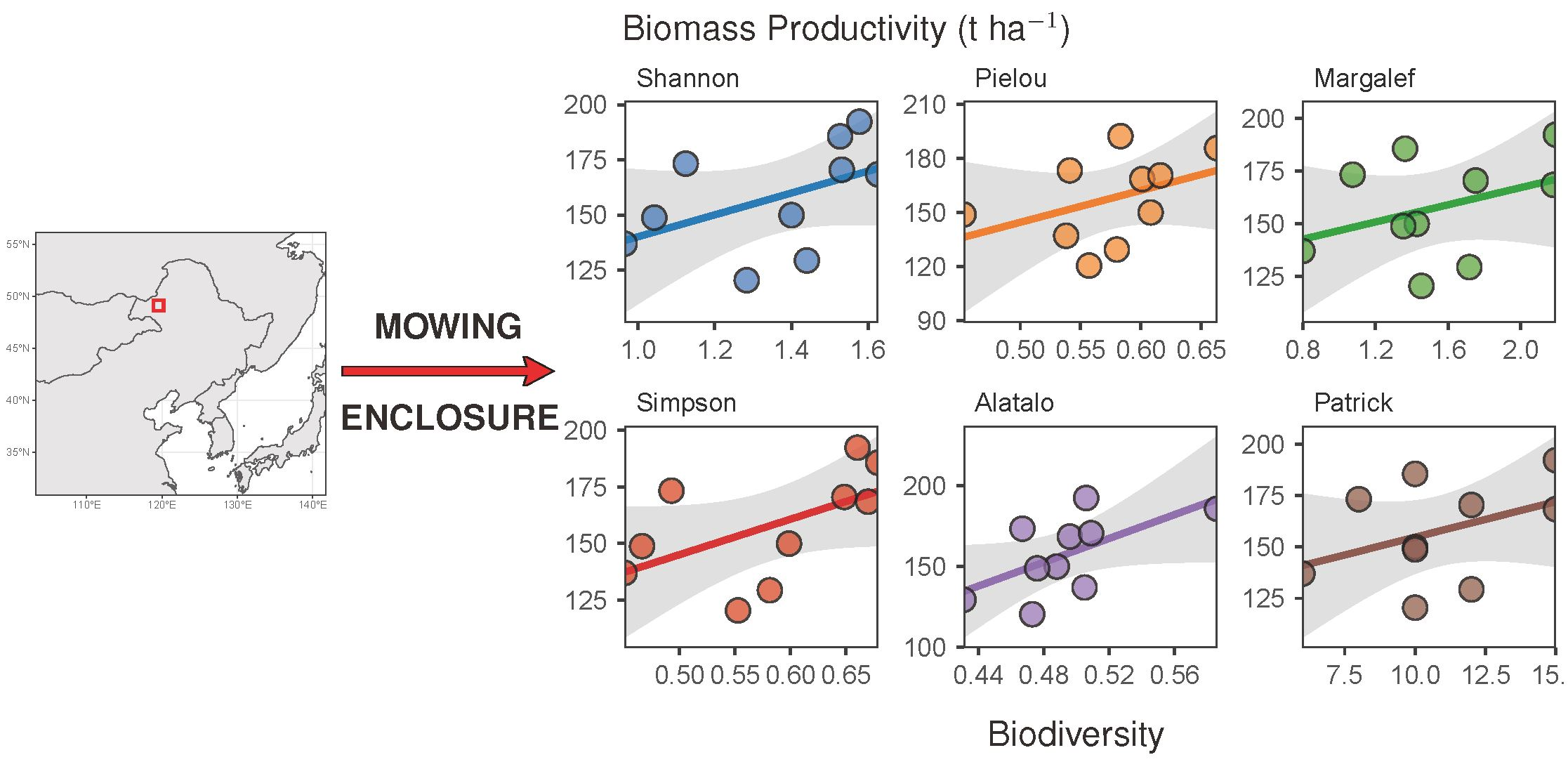
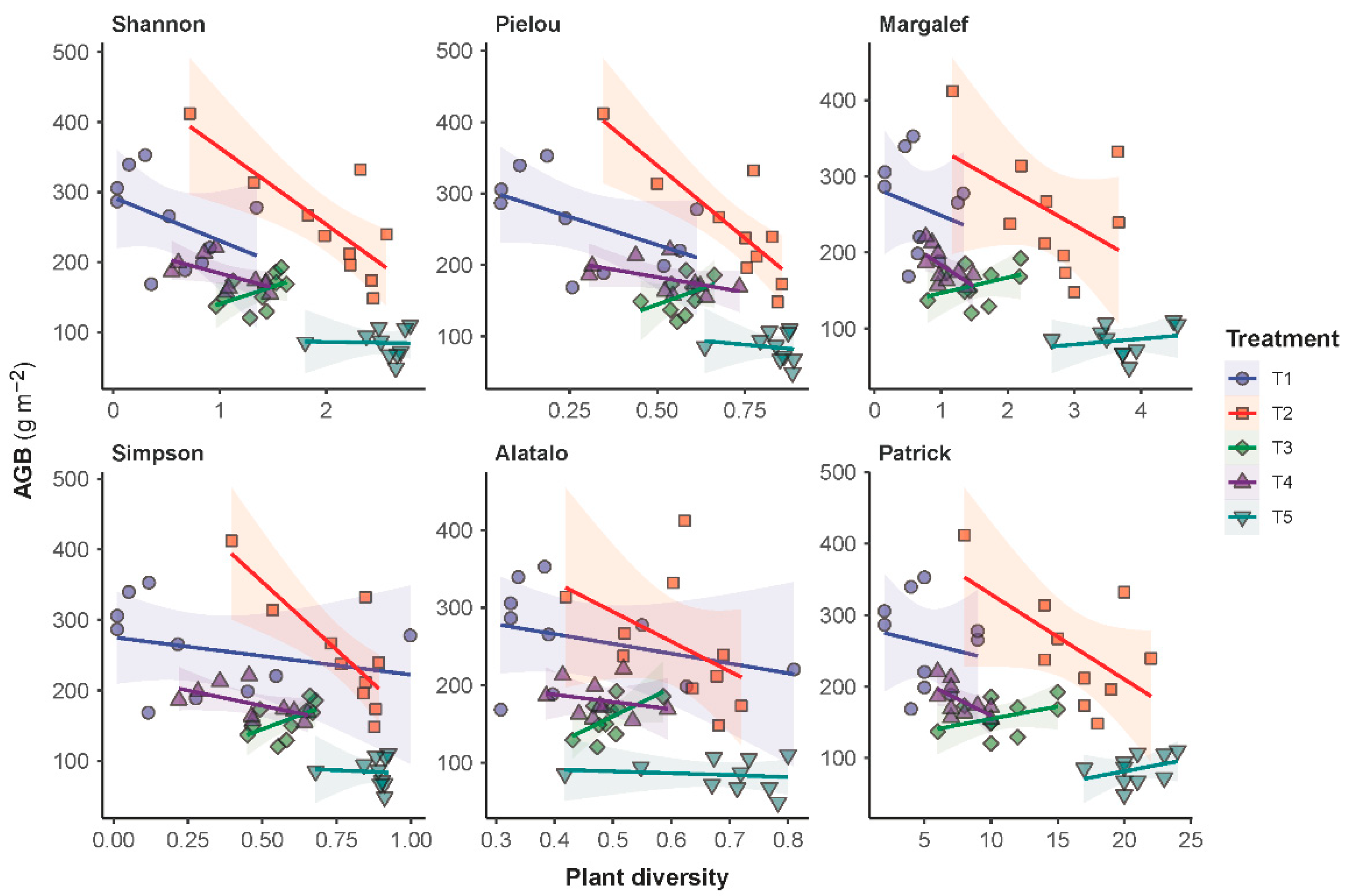
2.5. Relationship between AGB and Plant Diversity
3. Discussion
4. Materials and Methods
4.1. Study Area
4.2. Experimental Setting and Design
4.3. Vegetation Survey
4.4. Diversity Indices
4.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kang, L.; Han, X.; Zhang, Z.; Sun, O.J. Grassland ecosystems in China: Review of current knowledge and research advancement. Phil. Trans. R. Soc. B Biol. Sci. 2007, 362, 997–1008. [Google Scholar] [CrossRef]
- Li, L.; Chen, J.; Han, X.; Zhang, W.; Shao, C. Overview of Chinese grassland ecosystems. In Grassland Ecosystems of China: A Synthesis and Resume; Springer: Singapore, 2020; pp. 23–47. [Google Scholar] [CrossRef]
- Han, Z.; Han, C.; Yang, C. Spatial econometric analysis of environmental total factor productivity of ranimal husbandry and its influencing factors in China during 2001–2017. Sci. Total Environ. 2020, 723, 137726. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Shang, Z.; Gao, J.; Boone, R. Enhancing sustainability of grassland ecosystems through ecological restoration and grazing management in an era of climate change on Qinghai-Tibetan Plateau. Agric. Ecosyst. Environ. 2020, 287, 106684. [Google Scholar] [CrossRef]
- Wang, C.; Li, L.; Yan, Y.; Cai, Y.; Xu, D.; Wang, X.; Chen, J.; Xin, X. Effects of cultivation and agricultural abandonment on soil carbon, nitrogen and phosphorus in a meadow steppe in eastern Inner Mongolia. Agric. Ecosyst. Environ. 2021, 309, 107284. [Google Scholar] [CrossRef]
- Yao, Y.; Ye, L.; Tang, H.; Tang, P.; Wang, D.; Si, H.; Hu, W.; Van Ranst, E. Cropland soil organic matter content change in Northeast China, 1985–2005. Open Geosci. 2015, 7, 234–243. [Google Scholar] [CrossRef]
- Porensky, L.M.; Mueller, K.E.; Augustine, D.J.; Derner, J.D.; Moore, J. Thresholds and gradients in a semi-arid grassland: Long-term grazing treatments induce slow, continuous and reversible vegetation change. J. Appl. Ecol. 2016, 53, 1013–1022. [Google Scholar] [CrossRef] [Green Version]
- Godde, C.; Dizyee, K.; Ash, A.; Thornton, P.; Sloat, L.; Roura, E.; Henderson, B.; Herrero, M. Climate change and variability impacts on grazing herds: Insights from a system dynamics approach for semi-arid Australian rangelands. Glob. Chang. Biol. 2019, 25, 3091–3109. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Liu, Q.; Nie, Y.; Li, F.; Yang, G.; Tao, Y.; Lv, S.; Wu, X.; Ye, L. A multi-site evaluation of winter hardiness in indigenous alfalfa cultivars in northern China. Atmosphere 2021, 12, 1538. [Google Scholar] [CrossRef]
- Xu, L.; Nie, Y.; Chen, B.; Xin, X.; Yang, G.; Xu, D.; Ye, L. Effects of fence enclosure on vegetation community characteristics and productivity of a degraded temperate meadow steppe in northern China. Appl. Sci. 2020, 10, 2952. [Google Scholar] [CrossRef]
- Schönbach, P.; Wan, H.; Gierus, M.; Bai, Y.; Müller, K.; Lin, L.; Susenbeth, A.; Taube, F. Grassland responses to grazing: Effects of grazing intensity and management system in an Inner Mongolian steppe ecosystem. Plant Soil 2010, 340, 103–115. [Google Scholar] [CrossRef]
- Wang, Z.; Deng, X.; Song, W.; Li, Z.; Chen, J. What is the main cause of grassland degradation? A case study of grassland ecosystem service in the middle-south Inner Mongolia. Catena 2017, 150, 100–107. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Bullock, J.M.; Lavorel, S.; Manning, P.; Schaffner, U.; Ostle, N.; Chomel, M.; Durigan, G.; Fry, E.L.; Johnson, D.; et al. Combatting global grassland degradation. Nat. Rev. Earth Environ. 2021, 2, 720–735. [Google Scholar] [CrossRef]
- Ye, L.; Yang, J.; Verdoodt, A.; Moussadek, R.; Van Ranst, E. China’s food security threatened by soil degradation and biofuels production. In Proceedings of the 19th World Congress of Soil Science: Soil Solutions for a Changing World, Brisbane, Australia, 1–6 August 2010; pp. 5–8. [Google Scholar]
- Han, Z.; Song, W.; Deng, X.; Xu, X. Grassland ecosystem responses to climate change and human activities within the Three-River Headwaters region of China. Sci. Rep. 2018, 8, 9079. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Zhu, L.; Meng, J. Fuzzy evaluation of the ecological security of land resources in mainland China based on the Pressure-State-Response framework. Sci. Total Environ. 2022, 804, 150053. [Google Scholar] [CrossRef]
- Fulbright, T. Disturbance effects on species richness of herbaceous plants in a semi-arid habitat. J. Arid Environ. 2004, 58, 119–133. [Google Scholar] [CrossRef]
- Ye, L.-M.; Malingreau, J.-P.; Tang, H.-J.; Van Ranst, E. The breakfast imperative: The changing context of global food security. J. Integr. Agric. 2016, 15, 1179–1185. [Google Scholar] [CrossRef]
- Yang, G.; Peng, C.; Chen, H.; Dong, F.; Wu, N.; Yang, Y.; Zhang, Y.; Zhu, D.; He, Y.; Shi, S. Qinghai–Tibetan Plateau peatland sustainable utilization under anthropogenic disturbances and climate change. Ecosyst. Health Sustain. 2017, 3, e01263. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.A.; Holden, E.M.; Brown, C.; Cahill, J.F.J. Disturbance has lasting effects on functional traits and diversity of grassland plant communities. PeerJ 2022, 10, e13179. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.M. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 2006, 441, 629–632. [Google Scholar] [CrossRef]
- Sasaki, T.; Furukawa, T.; Iwasaki, Y.; Seto, M.; Mori, A. Perspectives for ecosystem management based on ecosystem resilience and ecological thresholds against multiple and stochastic disturbances. Ecol. Indic. 2015, 57, 395–408. [Google Scholar] [CrossRef]
- Guo, D.; Song, X.; Hu, R.; Cai, S.; Zhu, X.; Hao, Y. Grassland type-dependent spatiotemporal characteristics of productivity in Inner Mongolia and its response to climate factors. Sci. Total Environ. 2021, 775, 145644. [Google Scholar] [CrossRef]
- Bucharova, A.; Michalski, S.; Hermann, J.-M.; Heveling, K.; Durka, W.; Hölzel, N.; Kollmann, J.; Bossdorf, O.; Wan, S. Genetic differentiation and regional adaptation among seed origins used for grassland restoration: Lessons from a multispecies transplant experiment. J. Appl. Ecol. 2017, 54, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Xu, X.; Tang, X.; Xin, X.; Ye, L.; Yang, G.; Tang, H.; Lv, S.; Xu, D.; Zhang, Z. Managed grassland alters soil N dynamics and N2O emissions in temperate steppe. J. Environ. Sci. 2018, 66, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Yang, X.; Zhang, S.; An, S. Newly assimilated carbon allocation in grassland communities under different grazing enclosure times. Biol. Fertil. Soils 2021, 57, 563–574. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, Q.; Dong, S.; Liu, S.; Wang, X.; Su, X.; Li, Y.; Tang, L.; Wu, X.; Zhao, H. Effects of grazing and climate warming on plant diversity, productivity and living state in the alpine rangelands and cultivated grasslands of the Qinghai-Tibetan Plateau. Rangel. J. 2015, 37, 57–65. [Google Scholar] [CrossRef]
- Poeplau, C.; Marstorp, H.; Thored, K.; Kätterer, T. Effect of grassland cutting frequency on soil carbon storage—A case study on public lawns in three Swedish cities. Soil 2016, 2, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.; Wu, W.; Zhou, Q.; Tan, W.; Verburg, P.H.; Yang, P.; Ye, L. Modeling the spatio-temporal changes in land uses and its impacts on ecosystem services in Northeast China over 2000–2050. J. Geogr. Sci. 2018, 28, 1611–1625. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-J.; Xu, Y.; Shan, Y.-X.; Burgess, K.S.; Ge, X.-J.; Wang, S. Biotic and abiotic factors determine species diversity–productivity relationships in mountain meadows. J. Plant Ecol. 2021, 14, 1175–1188. [Google Scholar] [CrossRef]
- Niu, K.; Luo, Y.; Choler, P.; Du, G. The role of biomass allocation strategy in diversity loss due to fertilization. Basic Appl. Ecol. 2008, 9, 485–493. [Google Scholar] [CrossRef]
- Kang, M.; Dai, C.; Ji, W.; Jiang, Y.; Yuan, Z.; Chen, H. Biomass and its allocation in relation to temperature, precipitation, and soil nutrients in Inner Mongolia grasslands, China. PLoS ONE 2013, 8, e69561. [Google Scholar] [CrossRef] [PubMed]
- Ledger, M.; Edwards, F.; Brown, L.; Milner, A.; Woodward, G. Impact of simulated drought on ecosystem biomass production: An experimental test in stream mesocosms. Glob. Chang. Biol. 2011, 17, 2288–2297. [Google Scholar] [CrossRef]
- Ye, L.; Tang, H.; Yang, G.; Van Ranst, E. Adopting higher-yielding varieties to ensure Chinese food security under climate change in 2050. Proc. Environ. Sci. 2015, 29, 281. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Dong, S.; Xu, Y.; Wu, S.; Wu, X.; Zhang, X.; Zhi, Y.; Li, S.; Liu, S.; Li, Y.; et al. Resilience of revegetated grassland for restoring severely degraded alpine meadows is driven by plant and soil quality along recovery time: A case study from the Three-river Headwater Area of Qinghai-Tibetan Plateau. Agric. Ecosyst. Environ. 2019, 279, 169–177. [Google Scholar] [CrossRef]
- Tang, H.; Ye, L. Comparative Study on Methodology of Land Production Potential; China Agricultural Science and Technology Press: Beijing, China, 1997; p. 301. [Google Scholar]
- Wang, S.; Fan, J.; Li, Y.; Huang, L. Effects of grazing exclusion on biomass growth and species diversity among various grassland types of the Tibetan Plateau. Sustainability 2019, 11, 1705. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.-X.; Liu, G.-H.; Fu, B.-J.; Jin, T.-T.; Liu, Z.-F. Effects of biodiversity and plant community composition on productivity in semiarid grasslands of Hulunbeir, Inner Mongolia, China. Ann. N. Y. Acad. Sci. 2010, 1195, E52–E64. [Google Scholar] [CrossRef]
- Lamb, E.; Cahill, J. When competition does not matter: Grassland diversity and community composition. Am. Nat. 2008, 171, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Endara, M.-J.; Coley, P.D. The resource availability hypothesis revisited: A meta-analysis. Funct. Ecol. 2011, 25, 389–398. [Google Scholar] [CrossRef]
- Yue, K.; Jarvie, S.; Senior, A.; Van Meerbeek, K.; Peng, Y.; Ni, X.; Wu, F.; Svenning, J.C. Changes in plant diversity and its relationship with productivity in response to nitrogen addition, warming and increased rainfall. Oikos 2020, 129, 939–952. [Google Scholar] [CrossRef]
- Li, Y.; Dong, S.; Liu, S.; Su, X.; Wang, X.; Zhang, Y.; Zhao, Z.; Gao, X.; Li, S.; Tang, L. Relationships between plant diversity and biomass production of alpine grasslands are dependent on the spatial scale and the dimension of biodiversity. Ecol. Eng. 2019, 127, 375–382. [Google Scholar] [CrossRef]
- Wang, Z. Process strengths determine the forms of the relationship between plant species richness and primary productivity. PLoS ONE 2017, 12, e0185884. [Google Scholar] [CrossRef] [Green Version]
- Bonin, C.L.; Tracy, B.F. Diversity influences forage yield and stability in perennial prairie plant mixtures. Agric. Ecosyst. Environ. 2012, 162, 1–7. [Google Scholar] [CrossRef]
- Hagan, J.G.; Vanschoenwinkel, B.; Gamfeldt, L.; Courchamp, F. We should not necessarily expect positive relationships between biodiversity and ecosystem functioning in observational field data. Ecol. Lett. 2021, 24, 2537–2548. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.A.; Eviner, V.T.; Gaudin, A.C.M. Ways forward for resilience research in agroecosystems. Agric. Syst. 2018, 162, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Bakker, E.; Ritchie, M.; Olff, H.; Milchunas, D.; Knops, J. Herbivore impact on grassland plant diversity depends on habitat productivity and herbivore size. Ecol. Lett. 2006, 9, 780–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemp, D.R.; Guodong, H.; Xiangyang, H.; Michalk, D.L.; Fujiang, H.; Jianping, W.; Yingjun, Z. Innovative grassland management systems for environmental and livelihood benefits. Proc. Natl. Acad. Sci. USA 2013, 110, 8369–8374. [Google Scholar] [CrossRef] [Green Version]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015: International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; Food and Agriculture Organization of the United Nations: Rome, Italy, 2015; p. 192. [Google Scholar]
- Kent, M. Vegetation Description and Data Analysis: A Practical Approach, 2nd ed.; Wiley-Blackwell: Chichester, UK, 2012; p. 414. [Google Scholar]
- Spellerberg, I.F.; Fedor, P.J. A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon-Wiener’ Index. Glob. Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef] [Green Version]
- Morris, E.K.; Caruso, T.; Buscot, F.; Fischer, M.; Hancock, C.; Maier, T.S.; Meiners, T.; Müller, C.; Obermaier, E.; Prati, D.; et al. Choosing and using diversity indices: Insights for ecological applications from the German Biodiversity Exploratories. Ecol. Evol. 2014, 4, 3514–3524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dyke, F. Conservation Biology: Foundations, Concepts, Applications, 2nd ed.; Springer: Dordrecht, The Netherlands, 2008; p. 478. [Google Scholar] [CrossRef]
- Kvålseth, T.O. Evenness indices once again: Critical analysis of properties. SpringerPlus 2015, 4, 232. [Google Scholar] [CrossRef] [Green Version]
- Yeom, D.-J.; Kim, J.H. Comparative evaluation of species diversity indices in the natural deciduous forest of Mt. Jeombong. For. Sci. Technol. 2011, 7, 68–74. [Google Scholar] [CrossRef]
- Token, S.; Jiang, L.; Zhang, L.; Lv, G. Effects of plant diversity on primary productivity and community stability along soil water and salinity gradients. Glob. Ecol. Conserv. 2022, 36, e02095. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, Version 4.1.3; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 1 May 2022).
- Welham, S.J.; Gezan, S.A.; Clark, S.J.; Mead, A. Statistical Methods in Biology: Design and Analysis of Experiments and Regression; CRC Press: Boca Raton, FL, USA, 2015; p. 568. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot | Treatment | Fence-Enclosed | Size (Hectares) | Description |
|---|---|---|---|---|
| T1 | Fallow | Yes | 13 | Degraded forage grassland supplementally seeded with smooth bromegrass at 30 kg ha−1. In fallow since 2008. |
| T2 | Fallow | Yes | 46 | Degraded forage grassland under natural restoration. Fence-enclosed since 2005. |
| T3 | Mowing once per year | Yes | 12 | Degraded forage grassland. Fence-enclosed since 2005. Forage harvested by mowing in August every year at 7 cm above soil surface. |
| T4 | Mowingonce per three years | Yes | 12 | Degraded forage grassland. Fence-enclosed since 2005. Forage harvested by mowing once every three years in August at 7 cm above soil. |
| T5 | Livestock grazing | No | 60 | Degraded grazing grassland. Grazing intensity at 0.69 standard sheep units per hectare. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Nie, Y.; Xu, L.; Ye, L. Enclosure in Combination with Mowing Simultaneously Promoted Grassland Biodiversity and Biomass Productivity. Plants 2022, 11, 2037. https://doi.org/10.3390/plants11152037
Li D, Nie Y, Xu L, Ye L. Enclosure in Combination with Mowing Simultaneously Promoted Grassland Biodiversity and Biomass Productivity. Plants. 2022; 11(15):2037. https://doi.org/10.3390/plants11152037
Chicago/Turabian StyleLi, Da, Yingying Nie, Lijun Xu, and Liming Ye. 2022. "Enclosure in Combination with Mowing Simultaneously Promoted Grassland Biodiversity and Biomass Productivity" Plants 11, no. 15: 2037. https://doi.org/10.3390/plants11152037
APA StyleLi, D., Nie, Y., Xu, L., & Ye, L. (2022). Enclosure in Combination with Mowing Simultaneously Promoted Grassland Biodiversity and Biomass Productivity. Plants, 11(15), 2037. https://doi.org/10.3390/plants11152037