
Management and Utilization of Plant Genetic Resources for a Sustainable Agriculture



Abstract
:1. Introduction
2. Origin of Cultivated Species and Geographic Distribution of Crop Diversity
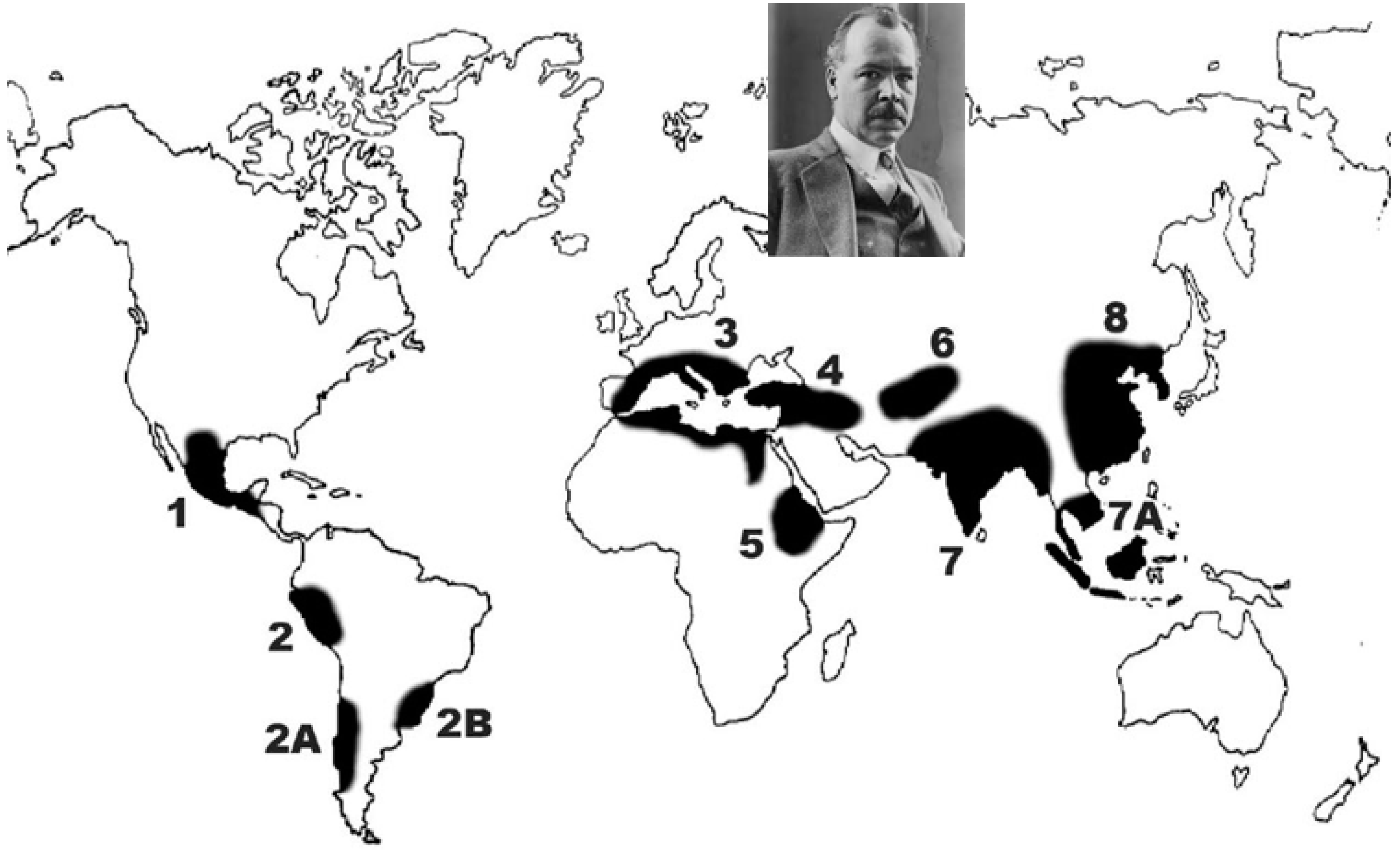
2.1. Vavilov’s Centres of Origin of Cultivated Plants and the Theory of Homologous Series of Variation
2.2. Landrace and Modern Cultivars; Their Genetic Structure in Relation to Diversity Management
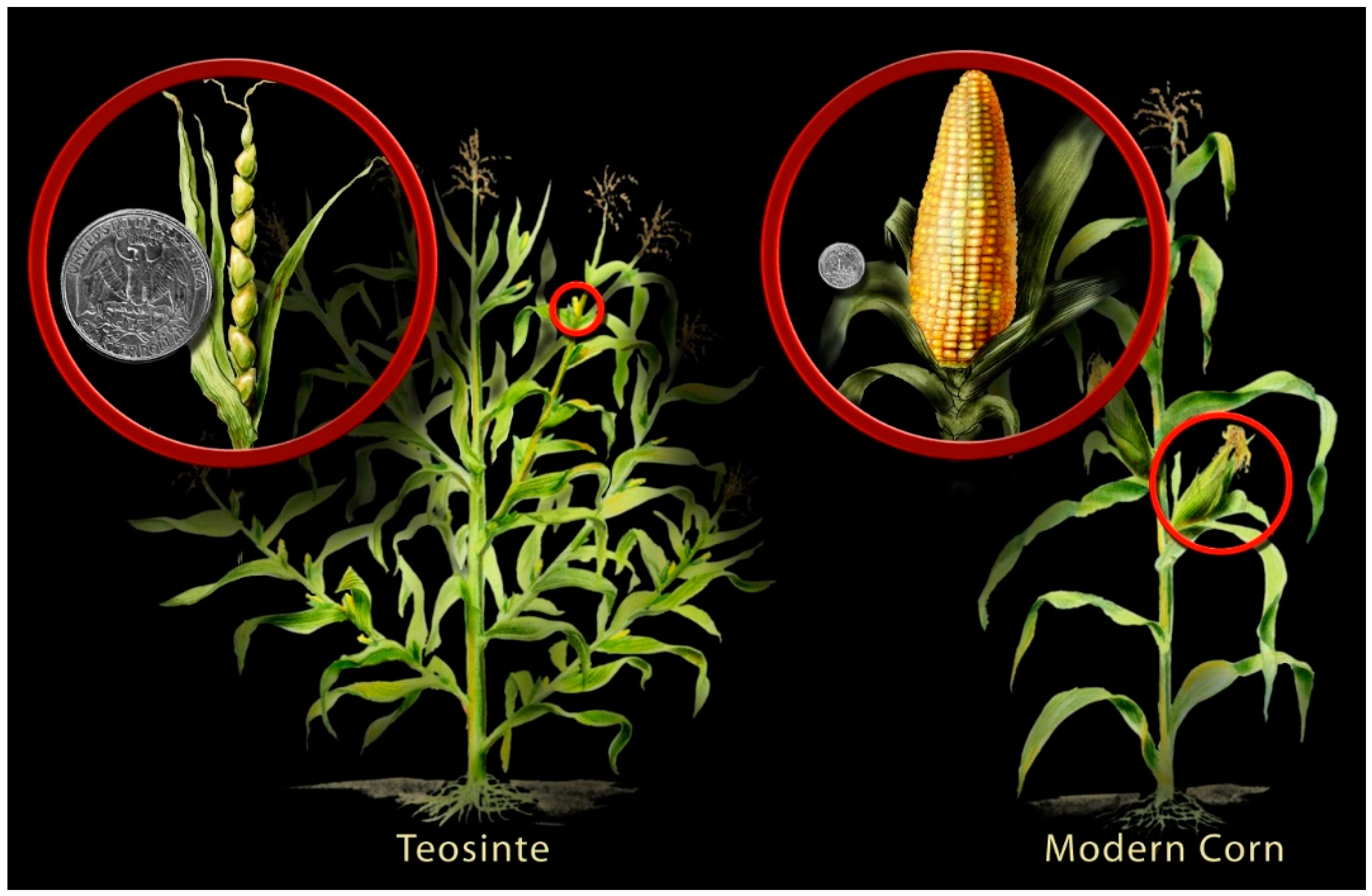
3. Crop Domestication and Domestication Traits
3.1. Primary Domestication Traits
3.2. Diversification Events
3.3. Physiological and Biochemical Changes
3.4. Genetic and Cytogenetic Changes
4. Current Status of Plant Genetic Resources in the Centres of Diversity
5. Role of Perennial Species in Sustainable Agriculture
Herbaceous Perennial Crops
6. Approaches to Germplasm Conservation
6.1. In Situ Conservation
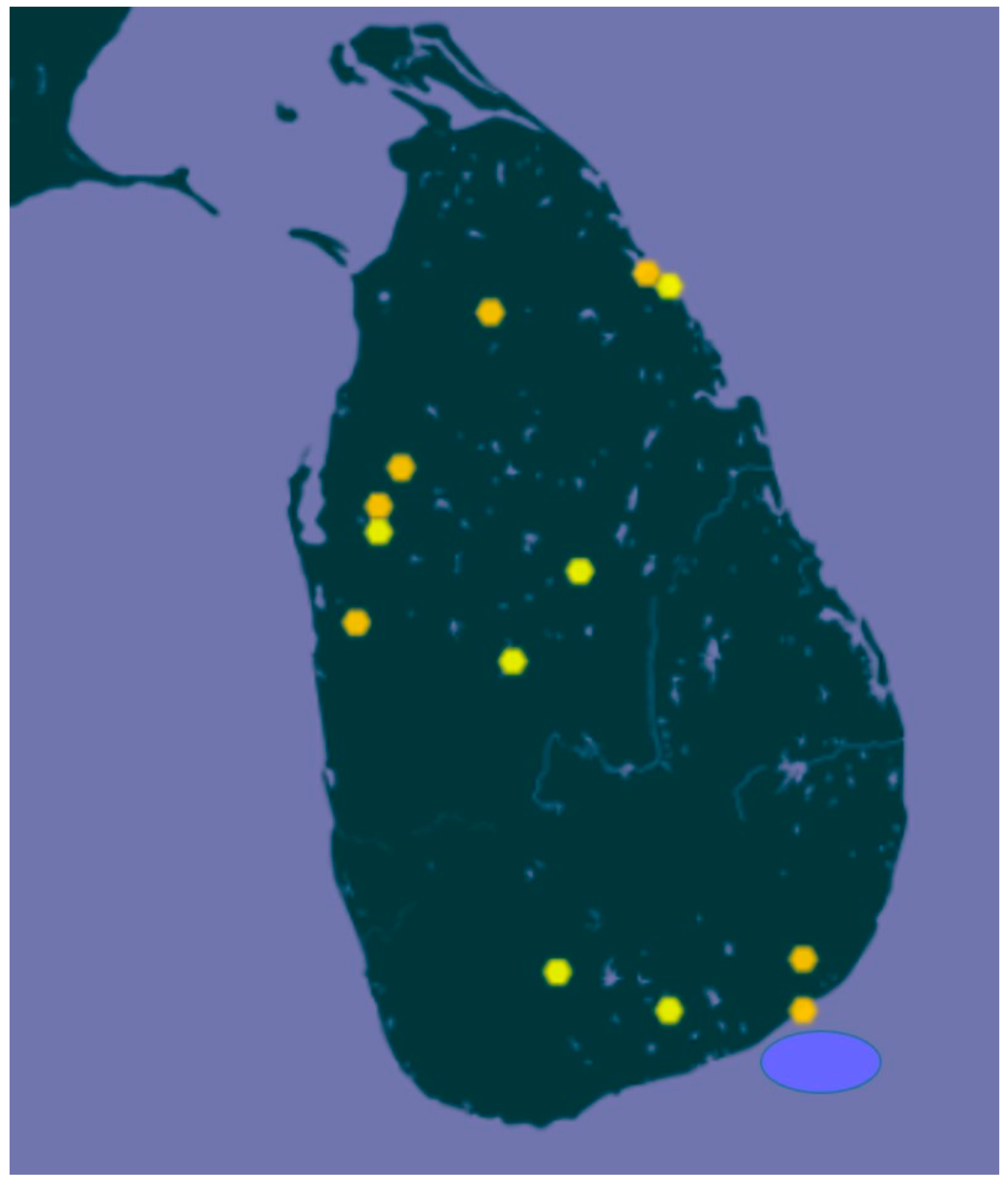
6.1.1. Identity of Crop Wild Relatives
6.1.2. Why In Situ Conservation?
6.1.3. Implementation of In Situ Conservation
6.2. Ex Situ Conservation
6.2.1. Origin of Genebanks and Their Spread
6.2.2. International and National Genebanks
6.3. Management of Ex Situ Collections
6.3.1. Management in Time
6.3.2. Identification of Duplicates
6.3.3. What to Conserve and Use—The Concept of Core Collections
6.4. Types of Collections in Genebanks, Their Management and Utilization
6.4.1. DNA Banks
6.4.2. Orthodox Seeds
6.4.3. Genebanking of Clonal Crops and Recalcitrant Species
Field Collections
In Vitro Collections
Cryopreserved Collections—Stopping the Biological Clock
7. Challenges to Plant Breeding in Search of Right Germplasm
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The Food and Agriculture Organisation. The State of Food Security and Nutrition in the World 2019. Safeguarding against Economic Slowdowns and Downturns. Available online: http://www.fao.org/3/ca5162en/ca5162en.pdf (accessed on 2 February 2022).
- UN. United Nations 2030 Agenda for Sustainable Development—Sustainable Development Goals. Goal 2: Zero Hunger. Available online: https://www.un.org/sustainabledevelopment/hunger/ (accessed on 7 June 2022).
- FAO. The State of Food Security and Nutrition in the World 2021: Transforming Food Systems for Food Security, Improved Nutrition and Affordable Healthy Diets for All. Available online: https://www.fao.org/3/cb4474en/online/cb4474en.html (accessed on 25 May 2022).
- IPBES. Global Assessment Report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; The Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES)—IPBES Secretariat: Bonn, Germany, 2019; p. 1144. [Google Scholar]
- IPCC. An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems. Available online: https://www.ipcc.ch/site/assets/uploads/2019/11/Headline-statements_Final.pdf (accessed on 5 May 2022).
- Song, X.-P. Global estimates of ecosystem service value and change: Taking into account uncertainties in satellite-based land cover data. Ecol. Econ. 2018, 143, 227–235. [Google Scholar] [CrossRef]
- Xu, H.; Cao, Y.; Yu, D.; Cao, M.; He, Y.; Gill, M.; Pereira, H.M. Ensuring effective implementation of the post-2020 global biodiversity targets. Nat. Ecol. Evol. 2021, 5, 411–418. [Google Scholar] [CrossRef] [PubMed]
- CBD. A New Global Framework for Managing Nature Through 2030: First Detailed Draft Agreement Debuts. Available online: https://www.cbd.int/article/draft-1-global-biodiversity-framework (accessed on 7 June 2022).
- Stevenson, J.R.; Villoria, N.; Byerlee, D.; Kelley, T.; Maredia, M. Green Revolution research saved an estimated 18 to 27 million hectares from being brought into agricultural production. Proc. Natl. Acad. Sci. USA 2013, 110, 8363–8368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pingali, P.L. Green Revolution: Impacts, limits, and the path ahead. Proc. Natl. Acad. Sci. USA 2012, 109, 12302–12308. [Google Scholar] [CrossRef] [Green Version]
- Karesh, W.B.; Dobson, A.; Lloyd-Smith, J.O.; Lubroth, J.; Dixon, M.A.; Bennett, M.; Aldrich, S.; Harrington, T.; Formenty, P.; Loh, E.H.; et al. Ecology of zoonoses: Natural and unnatural histories. Lancet 2012, 380, 1936–1945. [Google Scholar] [CrossRef]
- Jones, B.A.; Grace, D.; Kock, R.; Alonso, S.; Rushton, J.; Said, M.Y.; McKeever, D.; Mutua, F.; Young, J.; McDermott, J.; et al. Zoonosis emergence linked to agricultural intensification and environmental change. Proc. Natl. Acad. Sci. USA 2013, 110, 8399–8404. [Google Scholar] [CrossRef] [Green Version]
- Olivero, J.; Fa, J.E.; Real, R.; Márquez, A.L.; Farfán, M.A.; Vargas, J.M.; Gaveau, D.; Salim, M.A.; Park, D.; Suter, J.; et al. Recent loss of closed forests is associated with Ebola virus disease outbreaks. Sci. Rep. 2017, 7, 14291. [Google Scholar] [CrossRef] [Green Version]
- Rulli, M.C.; Santini, M.; Hayman, D.T.S.; D’Odorico, P. The nexus between forest fragmentation in Africa and Ebola virus disease outbreaks. Sci. Rep. 2017, 7, 41613. [Google Scholar] [CrossRef] [Green Version]
- Everard, M.; Johnston, P.; Santillo, D.; Staddon, C. The role of ecosystems in mitigation and management of COVID-19 and other zoonoses. Environ. Sci. Policy 2020, 111, 7–17. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [Green Version]
- Muehe, E.M.; Wang, T.; Kerl, C.F.; Planer-Friedrich, B.; Fendorf, S. Rice production threatened by coupled stresses of climate and soil arsenic. Nat. Commun. 2019, 10, 4985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meharg, A.A. Arsenic in rice—understanding a new disaster for South-East Asia. Trends Plant Sci. 2004, 9, 415–417. [Google Scholar] [CrossRef] [PubMed]
- Hassan, F.I.; Niaz, K.; Khan, F.; Maqbool, F.; Abdollahi, M. The relation between rice consumption, arsenic contamination, and prevalence of diabetes in South Asia. EXCLI J. 2017, 16, 1132–1143. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.L.; Puttick, M.N.; Clark, J.W.; Edwards, D.; Kenrick, P.; Pressel, S.; Wellman, C.H.; Yang, Z.; Schneider, H.; Donoghue, P.C.J. The timescale of early land plant evolution. Proc. Natl. Acad. Sci. USA 2018, 115, E2274–E2283. [Google Scholar] [CrossRef] [Green Version]
- Petraglia, M.D.; Haslam, M.; Fuller, D.Q.; Boivin, N.; Clarkson, C. Out of Africa: New hypotheses and evidence for the dispersal of Homo sapiens along the Indian Ocean rim. Ann. Hum. Biol. 2010, 37, 288–311. [Google Scholar] [CrossRef]
- Vavilov, N.I. Origin and Geography of Cultivated Plants; Nauka: Leningrad, Russia, 1987. (In Russian) [Google Scholar]
- Cohen, B.M. Nikolai Ivanovich Vavilov: The explorer and plant collector. Econ. Bot. 1991, 45, 38–46. [Google Scholar] [CrossRef]
- Zakharov, I.A. Nikolai I Vavilov (1887–1943). J. Biosci. 2005, 30, 299–301. [Google Scholar] [CrossRef]
- Dzyubenko, N.I. Vavilov’s collection of worldwide crop genetic resources in the 21st Century. Biopreservation Biobanking 2018, 16, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Vavilov, N.I. The law of homologous series in variation. J. Genet. 1922, 12, 47–89. [Google Scholar] [CrossRef]
- Voigt, W. Einige Gedanken zuVavilovs Gesetz der homologen Reihen und zu den Ursachen homologer genetischer Variationen aus wissenschaftstheoretischer Sicht. Die Kulturpflanze 1988, 36, 163–168. [Google Scholar] [CrossRef]
- Cheng, F.; Sun, R.; Hou, X.; Zheng, H.; Zhang, F.; Zhang, Y.; Liu, B.; Liang, J.; Zhuang, M.; Liu, Y.; et al. Subgenome parallel selection is associated with morphotype diversification and convergent crop domestication in Brassica rapa and Brassica oleracea. Nat. Genet. 2016, 48, 1218–1224. [Google Scholar] [CrossRef]
- Maluszynski, M.; Ahloowalia, B.S.; Sigurbjörnsson, B. Application of in vivo and in vitro mutation techniques for crop improvement. Euphytica 1995, 81, 303–315. [Google Scholar] [CrossRef]
- Pathirana, R. Plant mutation breeding in agriculture. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2011, 6, 1–20. [Google Scholar] [CrossRef]
- Foster, K.W.; Rutger, J.N. Inheritance of semidwarfism in rice Oryza sativa L. Genetics 1978, 88, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Rutger, J.N. Plant Mutation Breeding for Crop Improvement, Volume 1, Proceedings of the FAO/IAEA Symposium on Plant Mutation Breeding for Crop Improvement, Vienna, Austria, 18–22 June 1990; International Atomic Energy Agency: Vienna, Austria, 1991; pp. 155–165. [Google Scholar]
- Pathirana, R. Gamma-ray-induced field tolerance to Phytophthora blight in Sesame. Plant Breed. 1992, 108, 314–319. [Google Scholar] [CrossRef]
- Pathirana, R.; Weerasena, L.A.; Priyanthi, B. Development and release of gamma ray induced sesame mutant ANK-S2 in Sri Lanka. Trop. Agric. Res. Ext. 2000, 3, 19–24. [Google Scholar]
- Zhukovsky, P.M. Cultivated Plants and Their Progenitors; Kolos: Leningrad, Russia, 1971; p. 750. (In Russian) [Google Scholar]
- Harlan, J.R. Crops and Man; Crop Science Society of America: Madison, WI, USA, 1992. [Google Scholar]
- Meyer, R.S.; DuVal, A.E.; Jensen, H.R. Patterns and processes in crop domestication: An historical review and quantitative analysis of 203 global food crops. New Phytol. 2012, 196, 29–48. [Google Scholar] [CrossRef]
- Wang, M.; Yu, Y.; Haberer, G.; Marri, P.R.; Fan, C.; Goicoechea, J.L.; Zuccolo, A.; Song, X.; Kudrna, D.; Ammiraju, J.S.S.; et al. The genome sequence of African rice (Oryza glaberrima) and evidence for independent domestication. Nat. Genet. 2014, 46, 982–988. [Google Scholar] [CrossRef] [Green Version]
- Scarcelli, N.; Cubry, P.; Akakpo, R.; Thuillet, A.C.; Obidiegwu, J.; Baco, M.N.; Otoo, E.; Sonke, B.; Dansi, A.; Djedatin, G.; et al. Yam genomics supports West Africa as a major cradle of crop domestication. Sci. Adv. 2019, 5, eaaw1947. [Google Scholar] [CrossRef] [Green Version]
- Oumar, I.; Mariac, C.; Pham, J.-L.; Vigouroux, Y. Phylogeny and origin of pearl millet (Pennisetum glaucum [L.] R. Br) as revealed by microsatellite loci. Theor. Appl. Genet. 2008, 117, 489–497. [Google Scholar] [CrossRef]
- Li, H.-L. The origin of cultivated plants in Southeast Asia. Econ. Bot. 1970, 24, 3–19. [Google Scholar]
- Purugganan, M.D.; Fuller, D.Q. The nature of selection during plant domestication. Nature 2009, 457, 843–848. [Google Scholar] [CrossRef]
- Hyten, D.L.; Song, Q.; Zhu, Y.; Choi, I.-Y.; Nelson, R.L.; Costa, J.M.; Specht, J.E.; Shoemaker, R.C.; Cregan, P.B. Impacts of genetic bottlenecks on soybean genome diversity. Proc. Natl. Acad. Sci. USA 2006, 103, 16666–16671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.-H.; Zhou, G.; Ma, J.; Jiang, W.; Jin, L.-G.; Zhang, Z.; Guo, Y.; Zhang, J.; Sui, Y.; Zheng, L.; et al. De novo assembly of soybean wild relatives for pan-genome analysis of diversity and agronomic traits. Nat. Biotechnol. 2014, 32, 1045–1052. [Google Scholar] [CrossRef] [Green Version]
- Khush, G.S. Green revolution: The way forward. Nat. Rev. Genet. 2001, 2, 815–822. [Google Scholar] [CrossRef]
- Zakharova, E.S.; Epishin, S.M.; Vinetski, Y.P. An attempt to elucidate the origin of cultivated soybean via comparison of nucleotide sequences encoding glycinin B4 polypeptide of cultivated soybean, Glycine max, and its presumed wild progenitor, Glycine soja. Theor. Appl. Genet. 1989, 78, 852–856. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.-J.; Takahata, Y. A preliminary comparative evaluation of genetic diversity between Chinese and Japanese wild soybean (Glycine soja) germplasm pools using SSR markers. Genet. Resour. Crop Evol. 2007, 54, 157–165. [Google Scholar] [CrossRef]
- Doyle, J.J. 5S ribosomal gene variation in the soybean and its progenitor. Theor. Appl. Genet. 1988, 75, 621–624. [Google Scholar] [CrossRef]
- Mikel, M.A.; Diers, B.W.; Nelson, R.L.; Smith, H.H. Genetic diversity and agronomic improvement of North American soybean germplasm. Crop Sci. 2010, 50, 1219–1229. [Google Scholar] [CrossRef] [Green Version]
- Specht, J.E.; Williams, J.H. Contribution of Genetic Technology to Soybean Productivity—Retrospect and Prospect. In Genetic Contributions to Yield Gains of Five Major Crop Plants; Fehr, W.R., Ed.; Crop Science Society of America: Madison, WI, USA, 1984; pp. 49–74. [Google Scholar]
- Bisaliah, S. Soybean Development in India; UN/ESCAP CGPRT Centre: Bogor, Indonesia, 1986; p. 111. [Google Scholar]
- Agarwal, D.K.; Billore, S.D.; Sharma, A.N.; Dupare, B.U.; Srivastava, S.K. Soybean: Introduction, improvement, and utilization in India—Problems and prospects. Agric. Res. 2013, 2, 293–300. [Google Scholar] [CrossRef]
- Herath, E. Cultivation and uses of soybeans in Sri Lanka. In INTSOY Series—International Soybean Program (USA); College of Agricultural, University of Illinois at Urbana-Champaign: Champaign, IL, USA, 1976; Volume 10, pp. 250–252. [Google Scholar]
- Chithrapala, N.H.M.S.; Aberathne, M.S.; Samarakoon Manike, S.M.; Pushpakumara, U. MISB 01—A new soybean [Glycine max (L.) Merill] variety for the dry and intermediate zones of Sri Lanka. Ann. Sri Lanka Dep. Agric. 2015, 17, 180–186. [Google Scholar]
- Škorić, D. Achievements and future directions of sunflower breeding. Field Crops Res. 1992, 30, 231–270. [Google Scholar] [CrossRef]
- Goryunova, S.V.; Goryunov, D.V.; Chernova, A.I.; Martynova, E.U.; Dmitriev, A.E.; Boldyrev, S.V.; Ayupova, A.F.; Mazin, P.V.; Gurchenko, E.A.; Pavlova, A.S.; et al. Genetic and phenotypic diversity of the sunflower collection of the Pustovoit All-Russia Research Institute of Oil Crops (VNIIMK). Helia 2019, 42, 45–60. [Google Scholar] [CrossRef]
- Gavrilova, V.A.; Anisimova, I.N. Genealogy of the sunflower lines created on the basis of Russian varieties. Helia 2017, 40, 133–146. [Google Scholar] [CrossRef]
- Kumamoto, J.; Scora, R.W.; Lawton, H.W.; Clerx, W.A. Mystery of the forbidden fruit: Historical epilogue on the origin of the grapefruit, Citrus paradisi (Rutaceae). Econ. Bot. 1987, 41, 97–107. [Google Scholar] [CrossRef]
- Ferguson, A.R. Kiwifruit: The wild and the cultivated plants. Adv. Food Nutr. Res. 2013, 68, 15–32. [Google Scholar] [CrossRef]
- Ferguson, A.R.; Huang, H. Genetic resources of kiwifruit: Domestication and breeding. Hortic. Rev. 2007, 33, 1–121. [Google Scholar]
- Huang, H.; Wang, Y.; Zhang, Z.; Jiang, Z.; Wang, S. Actinidia germplasm resources and kiwifruit industry in China. Hortscience 2004, 39, 1165–1172. [Google Scholar] [CrossRef] [Green Version]
- Debenham, M.; Pathirana, R. Establishment and management of an in vitro repository of kiwifruit (Actinidia spp.) germplasm. In Meta-Topolin: A Growth Regulator for Plant Biotechnology and Agriculture; Ahmad, N., Strnad, M., Eds.; Springer Nature: Cham, Switzerland, 2020; pp. 279–291. [Google Scholar]
- Abraham, K.; Gopinathan Nair, P. Polyploidy and sterility in relation to sex in Dioscorea alata L. (Dioscoreaceae). Genetica 1991, 83, 93–97. [Google Scholar] [CrossRef]
- Meyer, R.S.; Purugganan, M.D. Evolution of crop species: Genetics of domestication and diversification. Nat. Rev. Genet. 2013, 14, 840–852. [Google Scholar] [CrossRef]
- Fuller, D.Q.; Harvey, E.L. The archaeobotany of Indian pulses: Identification, processing and evidence for cultivation. Environ. Archaeol. 2006, 11, 219–246. [Google Scholar] [CrossRef]
- Fuller, D.Q. Contrasting patterns in crop domestication and domestication rates: Recent archaeobotanical insights from the Old World. Ann. Bot. 2007, 100, 903–924. [Google Scholar] [CrossRef] [PubMed]
- Fuller, D.Q.; Denham, T.; Arroyo-Kalin, M.; Lucas, L.; Stevens, C.J.; Qin, L.; Allaby, R.G.; Purugganan, M.D. Convergent evolution and parallelism in plant domestication revealed by an expanding archaeological record. Proc. Natl. Acad. Sci. USA 2014, 111, 6147–6152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Zhou, A.; Sang, T. Rice domestication by reducing shattering. Science 2006, 311, 1936–1939. [Google Scholar] [CrossRef] [Green Version]
- Doebley, J.; Stec, A.; Hubbard, L. The evolution of apical dominance in maize. Nature 1997, 386, 485–488. [Google Scholar] [CrossRef]
- Wang, H.; Nussbaum-Wagler, T.; Li, B.; Zhao, Q.; Vigouroux, Y.; Faller, M.; Bomblies, K.; Lukens, L.; Doebley, J.F. The origin of the naked grains of maize. Nature 2005, 436, 714–719. [Google Scholar] [CrossRef]
- Harlan, J.R. Lettuce and the Sycomore: Sex and romance in ancient Egypt. Econ. Bot. 1986, 40, 4–15. [Google Scholar] [CrossRef]
- de Vries, I.M. Origin and domestication of Lactuca sativa L. Genet. Resour. Crop Evol. 1997, 44, 165–174. [Google Scholar] [CrossRef]
- Vaughan, D.A.; Balázs, E.; Heslop-Harrison, J.S. From crop domestication to super-domestication. Ann. Bot. 2007, 100, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Heslop-Harrison, J.S.; Schwarzacher, T. Domestication, genomics and the future for banana. Ann. Bot. 2007, 100, 1073–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trucchi, E.; Benazzo, A.; Lari, M.; Iob, A.; Vai, S.; Nanni, L.; Bellucci, E.; Bitocchi, E.; Raffini, F.; Xu, C.; et al. Ancient genomes reveal early Andean farmers selected common beans while preserving diversity. Nat. Plants 2021, 7, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Hammer, K.; Laghetti, G. Genetic erosion—Examples from Italy. Genet. Resour. Crop Evol. 2005, 52, 629–634. [Google Scholar] [CrossRef]
- Hammer, K.; Teklu, Y. Plant genetic resources: Selected issues from genetic erosion to genetic engineering. J. Agric. Rural. Dev. Trop. Subtrop. (JARTS) 2008, 109, 15–50. [Google Scholar]
- WCD. Dams and Development: A New Framework for Decision-Making. In The Report of the World Commission on Dams; Earthscan Publications Ltd.: London, UK, 2000; p. 356. [Google Scholar]
- Wu, J.; Huang, J.; Han, X.; Xie, Z.; Gao, X. Three-Gorges Dam--Experiment in Habitat Fragmentation? Science 2003, 300, 1239–1240. [Google Scholar] [CrossRef] [PubMed]
- Kleijn, D.; Bommarco, R.; Fijen, T.P.M.; Garibaldi, L.A.; Potts, S.G.; van der Putten, W.H. Ecological intensification: Bridging the gap between science and practice. Trends Ecol. Evol. 2019, 34, 154–166. [Google Scholar] [CrossRef] [Green Version]
- Ciotir, C.; Applequist, W.; Crews, T.E.; Cristea, N.; DeHaan, L.R.; Frawley, E.; Herron, S.; Magill, R.; Miller, J.; Roskov, Y.; et al. Building a botanical foundation for perennial agriculture: Global inventory of wild, perennial herbaceous Fabaceae species. Plants People Planet 2019, 1, 375–386. [Google Scholar] [CrossRef] [Green Version]
- McClure, K.A.; Sawler, J.; Gardner, K.M.; Money, D.; Myles, S. Genomics: A potential panacea for the perennial problem. Am. J. Bot. 2014, 101, 1780–1790. [Google Scholar] [CrossRef]
- Audu, V.; Rasche, F.; Martensson, L.M.D.; Emmerling, C. Perennial cereal grain cultivation: Implication on soil organic matter and related soil microbial parameters. Appl. Soil Ecol. 2022, 174, 104414. [Google Scholar] [CrossRef]
- Ledo, A.; Smith, P.; Zerihun, A.; Whitaker, J.; Vicente-Vicente, J.L.; Qin, Z.; McNamara, N.P.; Zinn, Y.L.; Llorente, M.; Liebig, M.; et al. Changes in soil organic carbon under perennial crops. Glob. Chang. Biol. 2020, 26, 4158–4168. [Google Scholar] [CrossRef]
- Means, M.; Crews, T.; Souza, L. Annual and perennial crop composition impacts on soil carbon and nitrogen dynamics at two different depths. Renew. Agric. Food Syst. 2022, 1–8. [Google Scholar] [CrossRef]
- Kreitzman, M.; Eyster, H.; Mitchell, M.; Czajewska, A.; Keeley, K.; Smukler, S.; Sullivan, N.; Verster, A.; Chan, K.M.A. Woody perennial polycultures in the U.S. Midwest enhance biodiversity and ecosystem functions. Ecosphere 2022, 13, e03890. [Google Scholar] [CrossRef]
- Félix, G.F.; Scholberg, J.M.S.; Clermont-Dauphin, C.; Cournac, L.; Tittonell, P. Enhancing agroecosystem productivity with woody perennials in semi-arid West Africa. A meta-analysis. Agron. Sustain. Dev. 2018, 38, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeHaan, L.; Larson, S.; López-Marqués, R.L.; Wenkel, S.; Gao, C.; Palmgren, M. Roadmap for accelerated domestication of an emerging perennial grain crop. Trends Plant Sci. 2020, 25, 525–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Tassel, D.L.; DeHaan, L.R.; Cox, T.S. Missing domesticated plant forms: Can artificial selection fill the gap? Evol. Appl. 2010, 3, 434–452. [Google Scholar] [CrossRef] [PubMed]
- Lundgren, M.R.; Des Marais, D.L. Life history variation as a model for understanding trade-offs in plant–environment interactions. Curr. Biol. 2020, 30, R180–R189. [Google Scholar] [CrossRef]
- Friedman, J. The evolution of annual and perennial plant life histories: Ecological correlates and genetic mechanisms. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 461–481. [Google Scholar] [CrossRef]
- DeHaan, L.R.; Van Tassel, D.L. Useful insights from evolutionary biology for developing perennial grain crops. Am. J. Bot. 2014, 101, 1801–1819. [Google Scholar] [CrossRef]
- Vico, G.; Manzoni, S.; Nkurunziza, L.; Murphy, K.; Weih, M. Trade-offs between seed output and life span—A quantitative comparison of traits between annual and perennial congeneric species. New Phytol. 2016, 209, 104–114. [Google Scholar] [CrossRef] [Green Version]
- Smaje, C. The Strong perennial vision: A critical review. Agroecol. Sustain. Food Syst. 2015, 39, 471–499. [Google Scholar] [CrossRef]
- Crews, T.E.; DeHaan, L.R. The strong perennial vision: A response. Agroecol. Sustain. Food Syst. 2015, 39, 500–515. [Google Scholar] [CrossRef]
- Crews, T.E.; Carton, W.; Olsson, L. Is the future of agriculture perennial? Imperatives and opportunities to reinvent agriculture by shifting from annual monocultures to perennial polycultures. Glob. Sustain. 2018, 1, e11. [Google Scholar] [CrossRef] [Green Version]
- Crews, T.E.; Blesh, J.; Culman, S.W.; Hayes, R.C.; Jensen, E.S.; Mack, M.C.; Peoples, M.B.; Schipanski, M.E. Going where no grains have gone before: From early to mid-succession. Agric. Ecosyst. Environ. 2016, 223, 223–238. [Google Scholar] [CrossRef]
- Jackson, W. New Roots for Agriculture; Friends of the Earth: San Francisco, CA, USA, 1980. [Google Scholar]
- Yu, H.; Li, J. Breeding future crops to feed the world through de novo domestication. Nat. Commun. 2022, 13, 1171. [Google Scholar] [CrossRef]
- Duhamel, M.; Vandenkoornhuyse, P. Sustainable agriculture: Possible trajectories from mutualistic symbiosis and plant neodomestication. Trends Plant Sci. 2013, 18, 597–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaut, B.S.; Seymour, D.K.; Liu, Q.; Zhou, Y. Demography and its effects on genomic variation in crop domestication. Nat. Plants 2018, 4, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhong, L.; Zhou, K.; Song, Z.; Chen, J.; Rong, J. Seed characteristic variations and genetic structure of wild Zizania latifolia along a latitudinal gradient in China: Implications for neo-domestication as a grain crop. AoB Plants 2018, 10, ply072. [Google Scholar] [CrossRef]
- Nooryazdan, H.; Serieys, H.; David, J.; Bacilieri, R.; Bervillé, A.J. Construction of a crop—Wild hybrid population for broadening genetic diversity in cultivated sunflower and first evaluation of its combining ability: The concept of neodomestication. Euphytica 2010, 178, 159–175. [Google Scholar] [CrossRef]
- Kantar, M.B.; Hüber, S.; Herman, A.; Bock, D.G.; Baute, G.; Betts, K.; Ott, M.; Brandvain, Y.; Wyse, D.; Stupar, R.M.; et al. Neo-domestication of an interspecific tetraploid Helianthus annuus × Helianthus tuberosus population that segregates for perennial habit. Genes 2018, 9, 422. [Google Scholar] [CrossRef] [Green Version]
- Tomooka, N.; Naito, K.; Kaga, A.; Sakai, H.; Isemura, T.; Ogiso-Tanaka, E.; Iseki, K.; Takahashi, Y. Evolution, domestication and neo-domestication of the genus Vigna. Plant Genet. Resour. 2014, 12, S168–S171. [Google Scholar] [CrossRef]
- Ciotir, C.; Townesmith, A.; Applequist, W.; Herron, S.; Van Tassel, D.; DeHaan, L.; Crews, T.; Schlautman, B.; Jackson, W.; Miller, A. Global Inventory and Systematic Evaluation of Perennial Grain, Legume, and Oilseed Species for Pre-Breeding and Domestication; Missouri Botanical Garden: St. Louis, MO, USA, 2016; Available online: http://www.tropicos.org/Project/PAPGI (accessed on 3 July 2022).
- Maxted, N.; Guarino, L.; Myer, L.; Chiwona, E.A. Towards a methodology for on-farm conservation of plant genetic resources. Genet. Resour. Crop Evol. 2002, 49, 31–46. [Google Scholar] [CrossRef]
- Kell, S.P.; Knüpffer, H.; Jury, S.L.; Ford-Lloyd, B.V.; Maxted, N. Crops and wild relatives of the EuroMediterranean region: Making and using a conservation catalogue. In Crop Wild Relative Conservation and Use; Maxted, N., Ford-Lloyd, B.V., Kell, S.P., Iriondo, J., Dulloo, E., Turok, J., Eds.; CAB International: Wallingford, UK, 2008; pp. 69–109. [Google Scholar]
- Harlan, J.R.; de Wet, J.M.J. Toward a rational classification of cultivated plants. Taxon 1971, 20, 509–517. [Google Scholar] [CrossRef]
- Maxted, N.; Ford-Lloyd, B.V.; Jury, S.; Kell, S.; Scholten, M. Towards a definition of a crop wild relative. Biodivers. Conserv. 2006, 15, 2673–2685. [Google Scholar] [CrossRef]
- Vaughan, D.A. A new rhizomatous Oryza species (Poaceae) from Sri Lanka. Bot. J. Linn. Soc. 1990, 103, 159–163. [Google Scholar] [CrossRef]
- Phillips, J.; Yang, L. Oryza Rhizomatis. The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/species/112680830/113899786 (accessed on 12 July 2020).
- GBIF. Oryza Rhizomatis D.A.Vaughan. Available online: https://www.gbif.org/species/4135750 (accessed on 17 August 2020).
- Huang, X.; Kurata, N.; Wei, X.; Wang, Z.-X.; Wang, A.; Zhao, Q.; Zhao, Y.; Liu, K.; Lu, H.; Li, W.; et al. A map of rice genome variation reveals the origin of cultivated rice. Nature 2012, 490, 497–501. [Google Scholar] [CrossRef] [Green Version]
- Nowicki, M.; Foolad, M.R.; Nowakowska, M.; Kozik, E.U. Potato and tomato late blight caused by Phytophthora infestans: An overview of pathology and resistance breeding. Plant Dis. 2012, 96, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Brar, D.S.; Khush, G.S. Alien introgression in rice. In Oryza: From Molecule to Plant; Sasaki, T., Moore, G., Eds.; Springer: Dordrecht, The Netherlands, 1997; pp. 35–47. [Google Scholar]
- Ram, T.; Deen, R.; Gautam, S.K.; Ramesh, K.; Rao, Y.K.; Brar, D.S. Identification of new genes for Brown Planthopper resistance in rice introgressed from O. glaberrima and O. minuta. Rice Genet. Newsl. 2010, 25, 67–68. [Google Scholar]
- Mondal, T.; Henry, R. The Wild Oryza Genomes; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Dey, S.; Badri, J.; Prakasam, V.; Bhadana, V.P.; Eswari, K.B.; Laha, G.S.; Priyanka, C.; Rajkumar, A.; Ram, T. Identification and agro-morphological characterization of rice genotypes resistant to sheath blight. Australas. Plant Pathol. 2016, 45, 145–153. [Google Scholar] [CrossRef]
- Ganeshan, P.; Jain, A.; Parmar, B.; Rao, A.R.; Sreenu, K.; Mishra, P.; Mesapogu, S.; Subrahmanyam, D.; Ram, T.; Sarla, N.; et al. Identification of salt tolerant rice lines among interspecific BILs developed by crossing Oryza sativa × O. rufipogon and O. sativa x O. nivara. Aust. J. Crop Sci. 2016, 10, 220–228. [Google Scholar]
- Prasanth, V.V.; Chakravarthi, D.V.; Kiran, T.V.; Rao, Y.V.; Panigrahy, M.; Mangrauthia, S.K.; Viraktamath, B.C.; Subrahmanyam, D.; Voleti, S.R.; Sarla, N. Evaluation of rice germplasm and introgression lines for heat tolerance. Ann. Biol. Res. 2012, 3, 5060–5068. [Google Scholar]
- Prasanth, V.V.; Basava, K.R.; Babu, M.S.; V. G.N., V.T.; Devi, S.J.S.R.; Mangrauthia, S.K.; Voleti, S.R.; Sarla, N. Field level evaluation of rice introgression lines for heat tolerance and validation of markers linked to spikelet fertility. Physiol. Mol. Biol. Plants 2016, 22, 179–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surapaneni, M.; Balakrishnan, D.; Mesapogu, S.; Addanki, K.R.; Yadavalli, V.R.; Tripura Venkata, V.G.N.; Neelamraju, S. Identification of major effect QTLs for agronomic traits and CSSLs in rice from Swarna/Oryza nivara derived backcross inbred lines. Front. Plant Sci. 2017, 8, 1027. [Google Scholar] [CrossRef] [PubMed]
- Furuta, T.; Uehara, K.; Angeles-Shim, R.B.; Shim, J.; Nagai, K.; Ashikari, M.; Takashi, T. Development of chromosome segment substitution lines harboring Oryza nivara genomic segments in Koshihikari and evaluation of yield-related traits. Breed. Sci. 2016, 66, 845–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, W.; Wang, W.; Zhou, Y.; Li, X.; Zheng, X.; Zhang, Q.; Wang, G.; Zhu, L. Breeding bacterial blight-resistant hybrid rice with the cloned bacterial blight resistance gene Xa21. Mol. Breed. 2002, 8, 285–293. [Google Scholar] [CrossRef]
- Guo, S.-B.; Wei, Y.; Li, X.-Q.; Liu, K.-Q.; Huang, F.-K.; Chen, C.-H.; Gao, G.-Q. Development and identification of introgression lines from cross of Oryza sativa and Oryza minuta. Rice Sci. 2013, 20, 95–102. [Google Scholar] [CrossRef]
- Yoon, D.B.; Kang, K.H.; Kim, K.J.; Ju, H.G.; Kwon, S.J.; Suh, J.P. Mapping quantitative trait loci for yield components and morphological traits in an advanced back cross population between Oryza grandiglumis and the O. sativa japonica cultivar Hwaseongbyeo. Theor. Appl. Genet. 2006, 112, 1052–1062. [Google Scholar] [CrossRef]
- Wairich, A.; de Oliveira, B.N.; Wu, L.B.; Murugaiyan, V.; Margis-Pinheiro, M.; Fett, J.P.; Ricachenevsky, F.K.; Frei, M. Chromosomal introgressions from Oryza meridionalis into domesticated rice Oryza sativa result in iron tolerance. J. Exp. Bot. 2021, 72, 2242–2259. [Google Scholar] [CrossRef]
- Luo, Y.; Lao, L.Y.; Ai, B.; Zhang, M.; Xie, J.K.; Zhang, F.T. Development of a drought stress-resistant rice restorer line through Oryza sativa-rufipogon hybridization. J. Genet. 2019, 98, 55. [Google Scholar] [CrossRef]
- Frankel, O.H.; Frankel, O.; Soulé, M.E. Conservation and Evolution; Cambridge University Press: Cambridge, UK, 1981. [Google Scholar]
- Jarvis, D.I.; Hodgkin, T.; Sthapit, B.R.; Fadda, C.; Lopez-Noriega, I. An heuristic framework for identifying multiple ways of supporting the conservation and use of traditional crop varieties within the agricultural production system. Crit. Rev. Plant Sci. 2011, 30, 125–176. [Google Scholar] [CrossRef] [Green Version]
- Khoury, C.K.; Brush, S.; Costich, D.E.; Curry, H.A.; Haan, S.; Engels, J.M.M.; Guarino, L.; Hoban, S.; Mercer, K.L.; Miller, A.J.; et al. Crop genetic erosion: Understanding and responding to loss of crop diversity. New Phytol. 2022, 233, 84–118. [Google Scholar] [CrossRef]
- Jarvis, D.J.; Hodgkin, T.; Brown, A.H.D.; Tuxill, J.; Noriega, I.L.; Smale, M.; Sthapit, B. Crop Genetic Diversity in the Field and on the Farm: Principles and Applications in Research Practices; Yale University Press: New Haven, CT, USA; London, UK, 2016. [Google Scholar]
- Porcuna-Ferrer, A.; Fiala, V.; Freyer, B.; van Etten, J.; Vernooy, R.; Probst, L. Do community seed banks contribute to the social-ecological resilience of communities? A case-study from western Guatemala. Int. J. Agric. Sustain. 2020, 18, 232–249. [Google Scholar] [CrossRef]
- Diez, M.J.; De la Rosa, L.; Martin, I.; Guasch, L.; Cartea, M.E.; Mallor, C.; Casals, J.; Simo, J.; Rivera, A.; Anastasio, G.; et al. Plant Genebanks: Present Situation and Proposals for Their Improvement. the Case of the Spanish Network. Front. Plant Sci. 2018, 9, 1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pautasso, M.; Aistara, G.; Barnaud, A.; Caillon, S.; Clouvel, P.; Coomes, O.T.; Delêtre, M.; Demeulenaere, E.; De Santis, P.; Döring, T.; et al. Seed exchange networks for agrobiodiversity conservation. A review. Agron. Sustain. Dev. 2012, 33, 151–175. [Google Scholar] [CrossRef] [Green Version]
- Sievers-Glotzbach, S.; Euler, J.; Frison, C.; Gmeiner, N.; Kliem, L.; Mazé, A.; Tschersich, J. Beyond the material: Knowledge aspects in seed commoning. Agric. Hum. Values 2021, 38, 509–524. [Google Scholar] [CrossRef]
- Sievers-Glotzbach, S.; Tschersich, J.; Gmeiner, N.; Kliem, L.; Ficiciyan, A. Diverse seeds—Shared practices: Conceptualizing seed commons. Int. J. Commons 2020, 14, 418–438. [Google Scholar] [CrossRef]
- Hammer, K.; Gladis, T.; Diederichsen, A. In situ and on-farm management of plant genetic resources. Eur. J. Agron. 2003, 19, 509–517. [Google Scholar] [CrossRef]
- Bush, D.; Thomson, L.; Bolatolu, W.; Dutt, S.; Hamani, S.; Likiafu, H.; Mateboto, J.; Tauraga, J.; Young, E. Domestication provides the key to conservation of Santalum yasi—A threatened Pacific sandalwood. Aust. For. 2020, 83, 186–194. [Google Scholar] [CrossRef]
- Curry, H.A. From working collections to the World Germplasm Project: Agricultural modernization and genetic conservation at the Rockefeller Foundation. Hist. Philos. Life Sci. 2017, 39, 5. [Google Scholar] [CrossRef] [Green Version]
- FAO. Commission on Plant Genetic Resources: FAO Activities and Future Programme on Plant Genetic Resources. Available online: http://www.fao.org/3/a-aj427e.pdf (accessed on 22 March 2022).
- Noriega, I.L.; Halewood, M.; Abberton, M.; Amri, A.; Angarawai, I.I.; Anglin, N.; Blümmel, M.; Bouman, B.; Campos, H.; Costich, D.; et al. CGIAR Operations under the Plant Treaty Framework. Crop Sci. 2019, 59, 819–832. [Google Scholar] [CrossRef] [Green Version]
- Westengen, O.T.; Lusty, C.; Yazbek, M.; Amri, A.; Asdal, Å. Safeguarding a global seed heritage from Syria to Svalbard. Nature Plants 2020, 6, 1311–1317. [Google Scholar] [CrossRef]
- FAO. The Second Report on the State of the World’s Plant Genetic Resources for Food and Agriculture. Available online: http://www.fao.org/3/i1500e/i1500e.pdf (accessed on 22 March 2022).
- Singh, A.K.; Varaprasad, K.S.; Venkateswaran, K. Conservation costs of plant genetic resources for food and agriculture: Seed genebanks. Agric. Res. 2012, 1, 223–239. [Google Scholar] [CrossRef] [Green Version]
- SGSV. Svalbard Global Seed Vault. Available online: https://www.croptrust.org/work/svalbard-global-seed-vault/ (accessed on 18 July 2022).
- Engels, J.M.M.; Ebert, A.W. A critical review of the current global ex situ conservation system for plant agrobiodiversity. I. History of the development of the global system in the context of the political/legal framework and its major conservation components. Plants 2021, 10, 1557. [Google Scholar] [CrossRef] [PubMed]
- Bellon, M.R.; Dulloo, E.; Sardos, J.; Thormann, I.; Burdon, J.J. In situ conservation-harnessing natural and human-derived evolutionary forces to ensure future crop adaptation. Evol. Appl. 2017, 10, 965–977. [Google Scholar] [CrossRef] [PubMed]
- de Haan, S.; Núñez, J.; Bonierbale, M.; Ghislain, M.; van der Maesen, J. A Simple Sequence Repeat (SSR) marker comparison of a large in- and ex-situ potato landrace cultivar collection from Peru reaffirms the complementary nature of both conservation strategies. Diversity 2013, 5, 505–521. [Google Scholar] [CrossRef]
- Casals, J.; Casanas, F.; Simo, J. Is it still necessary to continue to collect crop genetic resources in the Mediterranean Area? A case study in Catalonia. Econ. Bot. 2017, 71, 330–341. [Google Scholar] [CrossRef]
- Smykal, P.; Hradilova, I.; Trneny, O.; Brus, J.; Rathore, A.; Bariotakis, M.; Das, R.R.; Bhattacharyya, D.; Richards, C.; Coyne, C.J.; et al. Genomic diversity and macroecology of the crop wild relatives of domesticated pea. Sci. Rep. 2017, 7, 17384. [Google Scholar] [CrossRef] [Green Version]
- Abe, A.; Kosugi, S.; Yoshida, K.; Natsume, S.; Takagi, H.; Kanzaki, H. Genome sequencing reveals agronomically important loci in rice using MutMap. Nat. Biotechnol. 2012, 30, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Kirk, C.; Deng, C.; Wiedow, C.; Knaebel, M.; Brewer, L. Genotyping-by-sequencing of pear (Pyrus spp.) accessions unravels novel patterns of genetic diversity and selection footprints. Hortic. Res. 2017, 4, 17015. [Google Scholar] [CrossRef] [Green Version]
- Norelli, J.L.; Wisniewski, M.; Fazio, G.; Burchard, E.; Gutierrez, B.; Levin, E.; Droby, S. Genotyping-by-sequencing markers facilitate the identification of quantitative trait loci controlling resistance to Penicillium expansum in Malus sieversii. PLoS ONE 2017, 12, e0172949. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Wu, S.Y.; Raupp, W.J.; Sehgal, S.; Arora, S.; Tiwari, V.; Vikram, P.; Singh, S.; Chhuneja, P.; Gill, B.S.; et al. Efficient curation of genebanks using next generation sequencing reveals substantial duplication of germplasm accessions. Sci. Rep. 2019, 9, 650. [Google Scholar] [CrossRef] [Green Version]
- Melo, A.T.O.; Bartaula, R.; Hale, I. GBS-SNP-CROP: A reference-optional pipeline for SNP discovery and plant germplasm characterization using variable length, paired-end genotyping-by-sequencing data. BMC Bioinform. 2016, 17, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Schlotterer, C.; Tobler, R.; Kofler, R.; Nolte, V. Sequencing pools of individuals—mining genome-wide polymorphism data without big funding. Nat. Rev. Genet. 2014, 15, 749–763. [Google Scholar] [CrossRef] [PubMed]
- Wells, R.; Trick, M.; Fraser, F.; Soumpourou, E.; Clissold, L.; Morgan, C.; Pauquet, J.; Bancroft, I. Sequencing-based variant detection in the polyploid crop oilseed rape. BMC Plant Biol. 2013, 13, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, N.B.; Bai, Y.; Sun, H.H.; Wang, N.; Ma, Y.M.; Li, M.J.; Wang, X.; Jiao, C.; Legall, N.; Mao, L.Y.; et al. Genome re-sequencing reveals the history of apple and supports a two-stage model for fruit enlargement. Nat. Commun. 2017, 8, 249. [Google Scholar] [CrossRef]
- Bouquet, A. Sequencing of the grapevine genome: A brilliant demonstration of a 55-year-old hypothesis. Prog. Agric. Vitic. 2007, 124, 337–341. [Google Scholar]
- Brenchley, R.; Spannagl, M.; Pfeifer, M.; Barker, G.L.A.; D’Amore, R.; Allen, A.M.; McKenzie, N.; Kramer, M.; Kerhornou, A.; Bolser, D.; et al. Analysis of the bread wheat genome using whole-genome shotgun sequencing. Nature 2012, 491, 705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graner, A.; Kilian, B. NGS technologies for analyzing germplasm diversity in genebanks. Brief. Funct. Genom. 2012, 11, 38–50. [Google Scholar] [CrossRef] [Green Version]
- Frankel, O.H.; Brown, A.H.D. Current plant genetic resources: A critical appraisal. In Genetics: New Frontiers; Oxford & IBH Publishing Company: New Delhi, India, 1984; pp. 3–13. [Google Scholar]
- Franco, J.; Crossa, J.; Díaz, J.; Taba, S.; Villaseñor, J.; Eberhart, S.A. A sequential clustering strategy for classifying gene bank accessions. Crop Sci. 1997, 37, 1656–1662. [Google Scholar] [CrossRef]
- Gouesnard, B.; Bataillon, T.M.; Decoux, G.; Rozale, C.; Schoen, D.J.; David, J.L. MSTRAT: An algorithm for building germplasm core collections by maximizing allelic or phenotypic richness. J. Hered. 2001, 92, 93–94. [Google Scholar] [CrossRef] [Green Version]
- Agrama, H.A.; Yan, W.; Lee, F.; Fjellstrom, R.; Chen, M.-H.; Jia, M.; McClung, A. Genetic assessment of a mini-core subset developed from the USDA Rice Genebank. Crop Sci. 2009, 49, 1336–1346. [Google Scholar] [CrossRef] [Green Version]
- Pathirana, R.; Molloy, C.; Erridge, Z.; McLachlan, A.; Seelye, J.; Kumar, S. Towards a cryopreserved germplasm collection of apple—Results of dormant bud cryopreservation in the mild maritime winter climate of Hawkes Bay, New Zealand. Acta Hortic. 2018, 1205, 769–778. [Google Scholar] [CrossRef]
- Pathirana, R.; Mathew, L.; Jibran, R.; Hunter, D.A.; Morgan, E.R. Cryopreservation and cryotherapy research on horticultural crops in New Zealand. Acta Hortic. 2019, 1234, 29–36. [Google Scholar] [CrossRef]
- Rana, J.C.; Sharma, T.R.; Tyagi, R.K.; Chahota, R.K.; Gautam, N.K.; Singh, M.; Sharma, P.N.; Ojha, S.N. Characterisation of 4274 accessions of common bean (Phaseolus vulgaris L.) germplasm conserved in the Indian gene bank for phenological, morphological and agricultural traits. Euphytica 2015, 205, 441–457. [Google Scholar] [CrossRef]
- ICRISAT. ICRISAT Sorghum Core. Available online: http://genebank.icrisat.org/IND/Core?Crop=Sorghum (accessed on 20 May 2022).
- Mattick, J.S.; Ablett, E.M.; Edmonson, D.L. The gene library-preservation and analysis of genetic diversity in Australasia. In Conservation of Plant Genes; Academic Press: Cambridge, MA, USA, 1992; pp. 15–35. [Google Scholar]
- Droege, G.; Barker, K.; Astrin, J.J.; Bartels, P.; Butler, C.; Cantrill, D.; Coddington, J.; Forest, F.; Gemeinholzer, B.; Hobern, D.; et al. The Global Genome Biodiversity Network (GGBN) Data Portal. Nucleic Acids Res. 2014, 42, D607–D612. [Google Scholar] [CrossRef] [Green Version]
- Gemeinholzer, B.; Dröge, G.; Zetzsche, H.; Haszprunar, G.; Klenk, H.; Güntsch, A.; Berendsohn, W.G.; Wägele, J. The DNA Bank Network: The start from a German initiative. Biopreservation Biobanking 2011, 9, 51–55. [Google Scholar] [CrossRef]
- GGBN. Newsletter. Available online: https://wiki.ggbn.org/ggbn/Newsletters (accessed on 20 June 2022).
- GGBN. The GGBN Data Portal. Available online: http://www.ggbn.org/ggbn_portal/ (accessed on 27 June 2022).
- Engels, J.M.M.; Ebert, A.W. A critical review of the current global ex situ conservation system for plant agrobiodiversity. II. Strengths and weaknesses of the current system and recommendations for its improvement. Plants 2021, 10, 1904. [Google Scholar] [CrossRef] [PubMed]
- FAO. Genebank Standards for Plant Genetic Resources for Food and Agriculture, Revised Edition. Available online: http://www.fao.org/3/a-i3704e.pdf (accessed on 20 May 2022).
- Thormann, I.; Engels, J.M.M.; Halewood, M. Are the old International Board for Plant Genetic Resources (IBPGR) base collections available through the Plant Treaty’s multilateral system of access and benefit sharing? A review. Genet. Resour. Crop Evol. 2019, 66, 291–310. [Google Scholar] [CrossRef]
- Migicovsky, Z.; Warschefsky, E.; Klein, L.L.; Miller, A.J. Using living germplasm collections to characterize, improve, and conserve woody perennials. Crop Sci. 2019, 59, 2365–2380. [Google Scholar] [CrossRef] [Green Version]
- Forsline, P.L.; Aldwinckle, H.S. Natural occurrence of fire blight in USDA apple germplasm collection after 10 years of observation. Acta Hortic. 2002, 590, 351–357. [Google Scholar] [CrossRef]
- Datson, P.; Nardozza, S.; Manako, K.; Herrick, J.; Martinez-Sanchez, M.; Curtis, C.; Montefiori, M. Monitoring the Actinidia germplasm for resistance to Pseudomonas syringae pv. actinidiae. Acta Hortic. 2015, 1095, 181–184. [Google Scholar] [CrossRef]
- IPGRI-APO. Establishment and Management of Field Genebank, a Training Manual. Available online: https://www.bioversityinternational.org/fileadmin/_migrated/uploads/tx_news/Establishment_and_management_of_field_genebank_786.pdf (accessed on 19 April 2022).
- IPGRI. Technical Guidelines for the Management of Field and In Vitro Genebanks: IPGRI Handbooks for Genebanks No. 7. Available online: https://cropgenebank.sgrp.cgiar.org/images/file/learning_space/genebankmanual7.pdf (accessed on 19 March 2022).
- Carimi, F.; Pathirana, R.; Carra, A. Biotechnologies for germplasm management and improvement. In Grapevines—Varieties, Cultivation and Management; Szabo, P.V., Shojania, J., Eds.; Nova Science Publishers: New York, NY, USA, 2012; pp. 199–249. [Google Scholar]
- Aranzales, E.R. Use of Tissue Culture Techniques in the Conservation and Exchange of Cassava Germplasm Material: CIAT Case. International Course on Modern Technologies for Production, Processing and Utilization of the Cassava Crop. “Support to sustainable development of the cassava sector in the Caribbean Region.” CIAT, Palmira, Colombia, 28 July 2014. Available online: https://cgspace.cgiar.org/handle/10568/82943 (accessed on 15 February 2022).
- CGIAR-SGRP. Refinement and Standardization of Storage Procedures for Clonal Crops—Global Public Goods Phase 2: Part III. Multi-Crop Guidelines for Developing In Vitro Conservation Best Practices for Clonal Crops. Available online: https://cropgenebank.sgrp.cgiar.org/images/file/learning_space/gpg2%20part%20iii_final%20for%20web.pdf (accessed on 5 April 2022).
- CGIAR-SGRP. Refinement and Standardization of Storage Procedures for Clonal Crops—Global Public Goods Phase 2: Part II. Status of In Vitro Conservation Technologies for: Andean Root and Tuber Crops, Cassava, Musa, Potato, Sweetpotato and Yam. Available online: https://cropgenebank.sgrp.cgiar.org/images/file/learning_space/gpg2%20part%20ii%20_07022012.pdf (accessed on 5 April 2022).
- Pence, V.C.; Engelmann, F.; Guarino, L.; RamanathaRao, V.; Goldberg, E. Collecting in vitro for genetic resources conservation. In Collecting Plant Genetic Diversity: Technical Guidelines; Bioversity International: Rome, Italy, 2011; Volume 47, pp. 5–16. [Google Scholar]
- CIP-Genebank. In Vitro Conservation. Available online: https://cipotato.org/genebankcip/process/in-vitro/ (accessed on 23 June 2022).
- IITA-GRC. Genetic Resources Center. Available online: https://www.genebanks.org/genebanks/iita/; https://www.genebanks.org/wp-content/uploads/2021/04/IITA-Closure-Report-IN-VITRO-CONS-2018-Round-2.xlsx (accessed on 22 May 2022).
- Cunha Alves, C.; Azevedo, A.A.; Rennó, V.C. EMBRAPA Network for Brazilian Plant Genetic Resources Conservation. Biopreservation Biobanking 2018, 16, 350–360. [Google Scholar] [CrossRef] [Green Version]
- Myeza, P.N.; Visser, A. Agricultural Research Council’s potato in vitro genebank. Acta Hortic. 2013, 1007, 727–731. [Google Scholar] [CrossRef]
- Tyagi, R.K.; Agrawal, A. Genetic Resources Conservation: Ex Situ Conservation Strategies. Available online: http://www.nbpgr.ernet.in/genomic_resources/03_rktyagi_ex-situ.pdf (accessed on 20 February 2022).
- ITC. International Musa Germplasm Transit Center (ITC). Available online: https://www.bioversityinternational.org/banana-genebank/ (accessed on 22 August 2020).
- Bamberg, J.B.; Martin, M.W.; Abad, J.; Jenderek, M.M.; Tanner, J.; Donnelly, D.J.; Nassar, A.M.K.; Veilleux, R.E.; Novy, R.G. In vitro technology at the US Potato Genebank. Vitr. Cell. Dev. Biol.-Plant 2016, 52, 213–225. [Google Scholar] [CrossRef]
- Sakai, A. Survival of the twig of woody plants at −196 °C. Nature 1960, 185, 392–394. [Google Scholar] [CrossRef]
- Sakai, A.; Nishiyama, Y. Cryopreservation of winter vegetative buds of hardy fruit-trees in liquid-nitrogen. Hortscience 1978, 13, 225–227. [Google Scholar]
- Latta, R. Preservation of suspensions cultures of plant cells by freezing. Can. J. Bot. 1971, 49, 1253–1254. [Google Scholar] [CrossRef]
- Wang, M.-R.; Lambardi, M.; Engelmann, F.; Pathirana, R.; Panis, B.; Volk, G.M.; Wang, Q.-C. Advances in cryopreservation of in vitro-derived propagules: Technologies and explant sources. Plant Cell Tiss. Organ Cult. 2020, 144, 7–20. [Google Scholar] [CrossRef]
- Carra, A.; Carimi, F.; Bettoni, J.C.; Pathirana, R. Progress and challenges in the application of synthetic seed technology for ex situ germplasm conservation in grapevine (Vitis spp.). In Synthetic Seeds: Germplasm Regeneration, Preservation and Prospects; Faisal, M., Alatar, A.A., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 439–467. [Google Scholar]
- Sakai, A.; Kobayashi, S.; Oiyama, I. Cryopreservation of nucellar cells of navel orange (Citrus sinensis Osb. var. brasiliensis Tanaka) by vitrification. Plant Cell Rep. 1990, 9, 30–33. [Google Scholar] [CrossRef]
- Niino, T.; Wunna; Watanabe, K.; Nohara, N.; Rafique, T.; Yamamoto, S.; Fukui, K.; Arizaga, M.V.; Martinez, C.R.C.; Matsumoto, T.; et al. Cryopreservation of mat rush lateral buds by air dehydration using aluminum cryo-plate. Plant Biotechnol. 2014, 31, 281–287. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, S.; Rafique, T.; Fukui, K.; Sekizawa, K.; Niinois, T. V-cryo-plate procedure as an effective protocol for cryobanks: Case study of mint cryopreservation. Cryoletters 2012, 33, 12–23. [Google Scholar]
- Yamamoto, S.; Rafique, T.; Priyantha, W.S.; Fukui, K.; Matsumoto, T.; Niino, T. Development of a cryopreservation procedure using aluminium cryo-plates. Cryoletters 2011, 32, 256–265. [Google Scholar]
- Bettoni, J.C.; Pathirana, R.; Bonnart, R.; Shepherd, A.; Volk, G. Cryopreservation of grapevine shoot tips from in vitro plants using droplet vitrification and V cryo-plate techniques. In Synthetic Seeds: Germplasm Regeneration, Preservation and Prospects; Faisal, M., Alatar, A.A., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 469–482. [Google Scholar]
- Bettoni, J.C.; Mathew, L.; Pathirana, R.; Wiedow, C.; Hunter, D.A.; McLachlan, A.; Khan, S.; Tang, J.; Nadarajan, J. Eradication of Potato Virus S, Potato Virus A, and Potato Virus M from infected in vitro-grown potato shoots using in vitro therapies. Front. Plant Sci. 2022, 13, 878733. [Google Scholar] [CrossRef]
- Bettoni, J.C.; Souza, J.A.; Volk, G.M.; Dalla Costa, M.; da Silva, F.N.; Kretzschmar, A.A. Eradication of latent viruses from apple cultivar ‘Monalisa’ shoot tips using droplet-vitrification cryotherapy. Sci. Hortic. 2019, 250, 12–18. [Google Scholar] [CrossRef]
- Farhadi-Tooli, S.; Ghanbari, A.; Kermani, M.J.; Zeinalabedini, M.; Bettoni, J.C.; Naji, A.M.; Kazemi, N. Droplet-vitrification cryotherapy and thermotherapy as efficient tools for the eradication of apple chlorotic leaf spot virus and apple stem grooving virus from virus-infected quince in vitro cultures. Eur. J. Plant Pathol. 2022, 162, 31–43. [Google Scholar] [CrossRef]
- Mathew, L.; Tiffin, H.; Erridge, Z.; McLachlan, A.; Hunter, D.; Pathirana, R. Efficiency of eradication of Raspberry bushy dwarf virus from infected raspberry (Rubus idaeus) by in vitro chemotherapy, thermotherapy and cryotherapy and their combinations. Plant Cell Tiss. Organ Cult. 2021, 144, 133–141. [Google Scholar] [CrossRef]
- Pathirana, R.; McLachlan, A.; Hedderley, D.; Carra, A.; Carimi, F.; Panis, B. Removal of leafroll viruses from infected grapevine plants by droplet vitrification. Acta Hortic. 2015, 1083, 491–498. [Google Scholar] [CrossRef]
- Wang, Q.; Valkonen, J.P.T. Elimination of viruses and phytoplasma by cryotherapy of in vitro-grown shoot tips: Analysis of all cases. Adv. Hortic. Sci. 2007, 21, 265–269. [Google Scholar]
- Wang, Q.C.; Valkonen, J.P.T. Efficient elimination of sweetpotato little leaf phytoplasma from sweetpotato by cryotherapy of shoot tips. Plant Pathol. 2008, 57, 338–347. [Google Scholar] [CrossRef]
- Keller, E.R.J.; Zanke, C.D.; Senula, A.; Breuing, A.; Hardeweg, B.; Winkelmann, T. Comparing costs for different conservation strategies of garlic (Allium sativum L.) germplasm in genebanks. Genet. Resour. Crop Evol. 2013, 60, 913–926. [Google Scholar] [CrossRef] [Green Version]
- Dulloo, M.E.; Ebert, A.W.; Dussert, S.; Gotor, E.; Astorga, C.; Vasquez, N.; Rakotomalala, J.J.; Rabemiafara, A.; Eira, M.; Bellachew, B.; et al. Cost efficiency of cryopreservation as a long-term conservation method for coffee genetic resources. Crop Sci. 2009, 49, 2123–2138. [Google Scholar] [CrossRef]
- Jenderek, M.M.; Reed, B.M. Cryopreserved storage of clonal germplasm in the USDA National Plant Germplasm System. Vitr. Cell. Dev. Biol.-Plant 2017, 53, 299–308. [Google Scholar] [CrossRef]
- Panis, B. Sixty years of plant cryopreservation: From freezing hardy mulberry twigs to establishing reference crop collections for future generations. Acta Hortic. 2019, 1234, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Wilms, H.; Rhee, J.H.; Rivera, R.L.; Longin, K.; Panis, B. Developing coconut cryopreservation protocols and establishing cryo-genebank at RDA; a collaborative project between RDA and Bioversity International. Acta Hortic. 2019, 1234, 343–348. [Google Scholar] [CrossRef]
- Rajasekharan, P.E.; Ravish, B.S.; Kumar, T.V.; Ganeshan, S. Pollen Cryobanking for Tropical Plant Species. In Conservation of Tropical Plant Species; Normah, M.N., Chin, H.F., Reed, B.M., Eds.; Springer: New York, NY, USA, 2013; pp. 65–75. [Google Scholar]
- Carimi, F.; Carra, A.; Panis, B.; Pathirana, R. Strategies for conservation of endangered wild grapevine (Vitis vinifera L. subsp. sylvestris (C.C. Gmel.) Hegi). Acta Hortic. 2016, 1115, 81–86. [Google Scholar] [CrossRef]
- O’Brien, C.; Hiti-Bandaralage, J.; Folgado, R.; Hayward, A.; Lahmeyer, S.; Folsom, J.; Mitter, N. Cryopreservation of woody crops: The avocado case. Plants 2021, 10, 934. [Google Scholar] [CrossRef]
- Pathirana, R.; Mathew, L.; McLachlan, A. A simplified method for high recovery of kiwifruit (Actinidia spp.) shoot tips after droplet vitrification cryopreservation suitable for long-term conservation. Plant Cell Tiss. Organ Cult. 2021, 144, 97–102. [Google Scholar] [CrossRef]
- Mathew, L.; Burritt, D.J.; McLachlan, A.; Pathirana, R. Combined pre-treatments enhance antioxidant metabolism and improve survival of cryopreserved kiwifruit shoot tips. Plant Cell Tiss. Organ Cult. 2019, 138, 193–205. [Google Scholar] [CrossRef]
- Carra, A.; Sajeva, M.; Abbate, L.; Siragusa, M.; Pathirana, R.; Carimi, F. Factors affecting somatic embryogenesis in eight Italian grapevine cultivars and the genetic stability of embryo-derived regenerants as assessed by molecular markers. Sci. Hortic. 2016, 204, 123–127. [Google Scholar] [CrossRef]
- Niskanen, A.-M.; Lu, J.; Seitz, S.; Keinonen, K.; Von Weissenberg, K.; Pappinen, A. Effect of parent genotype on somatic embryogenesis in Scots pine (Pinus sylvestris). Tree Physiol. 2004, 24, 1259–1265. [Google Scholar] [CrossRef] [Green Version]
- Panis, B.; Nagel, M.; Van den houwe, I. Challenges and prospects for the conservation of crop genetic resources in field genebanks, in in vitro collections and/or in liquid nitrogen. Plants 2020, 9, 1634. [Google Scholar] [CrossRef]
- Nadarajan, J.; van der Walt, K.; Lehnebach, C.A.; Saeiahagh, H.; Pathirana, R. Integrated ex situ conservation strategies for endangered New Zealand Myrtaceae species. N. Z. J. Bot. 2020, 59, 72–89. [Google Scholar] [CrossRef]
- Byrne, P.F.; Volk, G.M.; Gardner, C.; Gore, M.A.; Simon, P.W.; Smith, S. Sustaining the future of plant breeding: The critical role of the USDA-ARS National Plant Germplasm System. Crop Sci. 2018, 58, 451–468. [Google Scholar] [CrossRef] [Green Version]
- GRIN-Global. U.S. National Plant Germplasm System. Available online: https://npgsweb.ars-grin.gov/gringlobal/search.aspx (accessed on 25 March 2022).
- CGIAR-Genebank-Platform. CGIAR Centers’ Use of Digital Object Identifiers (DOIs): A Submission to the Advisory Committee on the Global Information System 10 p. Available online: https://cgspace.cgiar.org/handle/10568/102457 (accessed on 22 May 2022).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time in History | Event |
|---|---|
| 515–470 mya | First land plants |
| 350 mya | Emergence of angiosperms |
| 160 mya | Monocots separated from dicots |
| 6.5 mya | Hominids appear |
| 2 mya | Homo habilis |
| 1.75 mya | Homo erectus |
| 195,000–160,000 ya | Homo sapiens |
| 130,000–120,000 ya | Human migration out of Africa |
| 13,000 ya | Settled agriculture and beginning of crop domestication |
| Region | Crop |
|---|---|
| South Mexico–Central America | Avocado, Maize, Sweet Potato, Tomato, Capsicum spp., Tobacco, Cucurbita pepo, C. moschata, Phaseolus spp., Amarnthus cruentus, A. hypochondriacus, and Gossypium hirsutum |
| South American Andes (Peru, Bolivia, and Ecuador) | Potato, Quinoa, Lima Bean, Common Bean, Tomato, Capsicum spp., Cucurbita maxima, C. moschata, Grain amaranth (Amaranthus caudtus), Oca (Oxalis tuberosa), Ulluco (Ullucus tuberosus), Añu (Tropaeolum tuberosum), Achira (Canna edulis), Coca, Gossypium barbadense |
| Tropical lowland South America (Chile, Paraguay, and Southern Brazil) | Cassava, Arrowroot, Cocoyam, Peanut, Pineapple, and Capsicum chinense |
| Mediterranean | Grapevine, Carrot, Cabbage, Olive, Sugar Beet, European Pear, Vicia faba, V. sativa, Lathyrus ochrus, Cicer arietinum, and Almond |
| Asia Minor (Middle East) | Cicer arietinum (secondary centre), Lens culinaris, L. orientalis, Vicia ervilia, Pisum sativum, Medicago sativa, Trifolium resupinatum, Trigonella foenum-graecum, Onobrychis spp., Lathyrus cicera, Vicia spp., Date Palm, and Lettuce |
| Abyssinia (Ethiopian Centre) | Millets, Sorghum, Castor, Coffee (Coffea arabica L.), Peanut, Teff (Eragrostis abyssiniaca Link.), Finger Millet, Sesame, and Niger (Guizotia abyssiniaca Cass.) |
| Inner Asia | Wheat, Barley, Apple, and Onion |
| India | Mung Bean, Rice, Black Gram, Pigeon Pea, Horsegram, Mango, Little Millet (Panicum sumatrense), and Flax |
| Indo-Malaya | Rambuttan, Banana, Sugarcane, and Yam |
| China | Rice, Soybean, Peach, Foxtail Millet (Setaria italica), Proso Millet (Panicum miliaceum), Hemp, Tea, Chinese Cabbage, Mulberry, and Citrus spp. |
| Oryza Species | Genome | Trait of Interest | Line Number | Reference |
|---|---|---|---|---|
| O. glaberrima | AA | Brown planthopper (Nilaparvata lugens Stål.) resistance | IR 75870-5-8-5-B-1-B), IR 75870-5-8-5-B-2-B) | [117] |
| O. nivara | AA | Brown planthopper tolerance | IR28, IR29, IR30, IR34, IR36, IR38, IR40, IR48, IR50, IR56, IR58, | [118] |
| O. minuta | BBCC | Brown plant hopper resistance | IR 71033-62- 15, IR 71033-121-15 | [117] |
| O. nivara | AA | Sheath blight (Rhizoctonia solani) tolerance | RPBio4918-10-3 | [119] |
| O. nivara | AA | Salinity tolerance | 14S, 75S, 166S, IL 3-1K | [120] |
| O. rufipogon | AA | Salinity tolerance | Chinsurah Nona 2 | [118] |
| O. nivara | AA | Heat tolerance | 166-2, 175-2, 3-1K | [121] |
| O. rufipogon | AA | Heat tolerance | 377-13, 50 | [121] |
| O. nivara | AA | Heat tolerance | 24S, 70S, 14-3S | [122] |
| O. nivara | AA | High yield | 220S, 10-2S | [123] |
| O. nivara | AA | 100 grain weight, early flowering | NSL-15, NSL-22 | [124] |
| O. sativa f. spontanea | AA | Cytoplasmic male sterility | Mondal and Henry [118] | |
| O. rufipogon | AA | Rice tungro bacilliform virus resistance | Matatag 9 | [118] |
| O. longistaminata | AA | Bacterial blight (Xanthomonas oryzae pv. oryzae) | Shanyou63-Xa21 | [125] |
| O. rufipogon | AA | Acid sulphate tolerance | AS 996 | [118] |
| O. minuta | BBCC | Bacterial blight | 41 Lines | [126] |
| O. minuta | BBCC | Brown planthopper | 11 Lines | [126] |
| O. minuta | BBCC | Whitebacked planthopper (Sogatella furcifera) | 7 Lines | [126] |
| O.grandiglumis | CCDD | Grain weight and other yield traits | HG 101 | [127] |
| O. meridionalis | AA | Iron tolerance | CM 23, CM 24 | [128] |
| Oryza rufipogon ‘DXWR’ | AA | Drought tolerance | Restorer line BIL627 | [129] |
| Genebank | Country | Crop | Number of Accessions | Reference |
|---|---|---|---|---|
| International Potato Centre | Peru | Potato, Andean Root and Tubers | >11,000 | CIP-Genebank [190] |
| International Institute for Tropical Agriculture | Nigeria | Cassava | >2500 | IITA-GRC [191] |
| Yam | >2500 | |||
| Banana | >500 | |||
| EMBRAPA Genebank | Brazil | 24 genera, 63 species | 1250 | Cunha Alves, et al. [192] |
| Agricultural Research Council | South Africa | Potato | 1100 | Myeza and Visser [193] |
| National Bureau of Plant Genetic Resources | India | Fruit crops | 743 | Tyagi and Agrawal [194] |
| Tuber crops | 611 | |||
| Spices | 380 | |||
| Bulbous crops | 171 | |||
| Medicinal and Aromatic | 170 | |||
| Total 24 Genera, 63 spp. | 1250 | |||
| Bioversity International Transit Centre | Belgium | Musa spp. | >1500 | ITC [195] |
| International Centre for Tropical Agriculture (CIAT) | Columbia | Cassava | 6632 | Rondon [186] |
| The New Zealand Institute for Plant and Food Research Limited | New Zealand | Kiwifruit (Actinidia spp.) | 1012 | Debenham and Pathirana [62] |
| United States Department of Agriculture Agricultural Research Service | USA | Potato | ~1000 | Bamberg et al. [196] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pathirana, R.; Carimi, F. Management and Utilization of Plant Genetic Resources for a Sustainable Agriculture. Plants 2022, 11, 2038. https://doi.org/10.3390/plants11152038
Pathirana R, Carimi F. Management and Utilization of Plant Genetic Resources for a Sustainable Agriculture. Plants. 2022; 11(15):2038. https://doi.org/10.3390/plants11152038
Chicago/Turabian StylePathirana, Ranjith, and Francesco Carimi. 2022. "Management and Utilization of Plant Genetic Resources for a Sustainable Agriculture" Plants 11, no. 15: 2038. https://doi.org/10.3390/plants11152038
APA StylePathirana, R., & Carimi, F. (2022). Management and Utilization of Plant Genetic Resources for a Sustainable Agriculture. Plants, 11(15), 2038. https://doi.org/10.3390/plants11152038