
Functional Ecology of External Secretory Structures in Rivea ornata (Roxb.) Choisy (Convolvulaceae)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species and Sample Collection
2.2. Micromorphology and Anatomy
2.3. Histochemistry
2.4. Preliminary Observation of Rivea ornata Visitors
3. Results
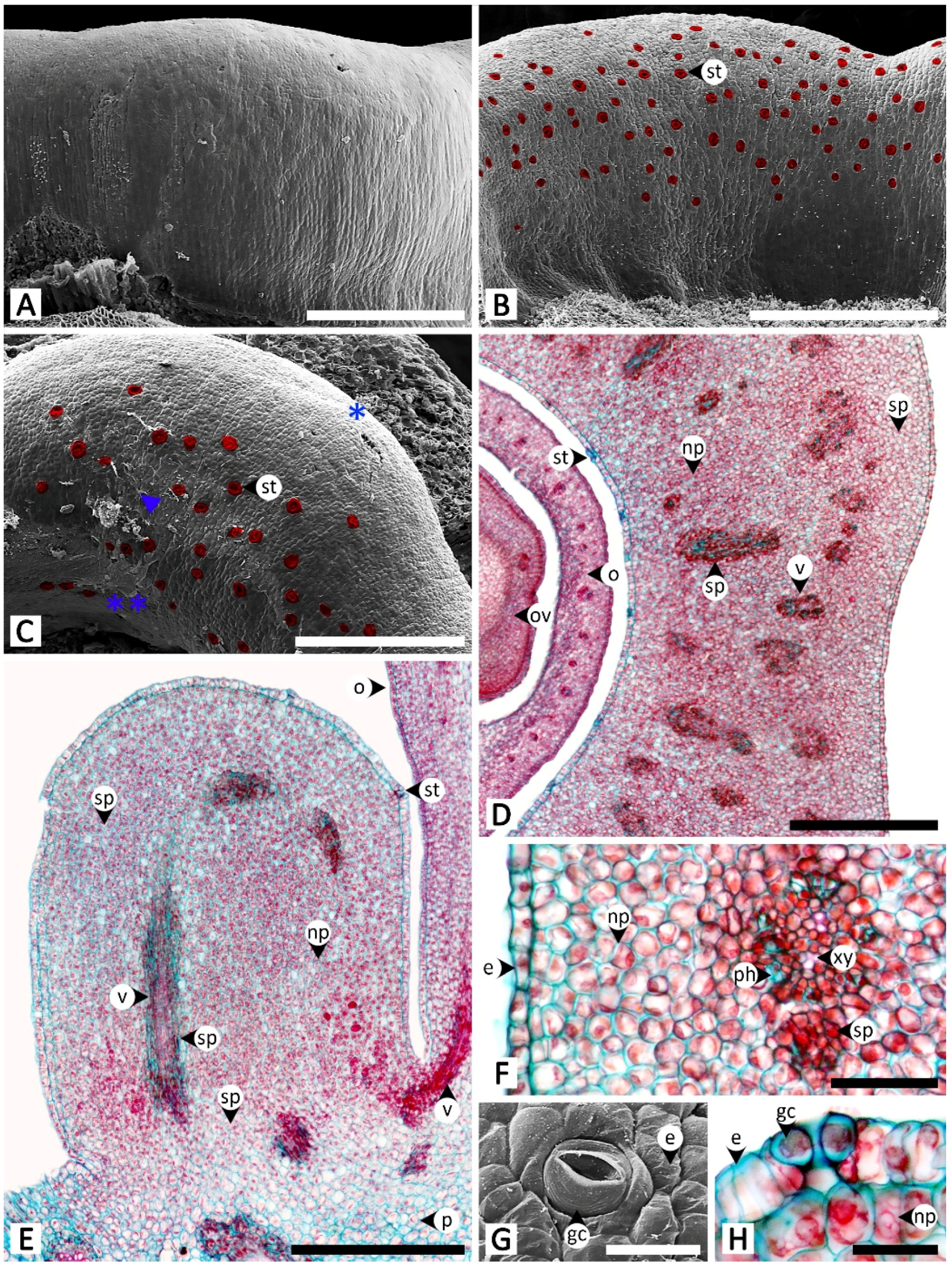
3.1. Micromorphology and Anatomy
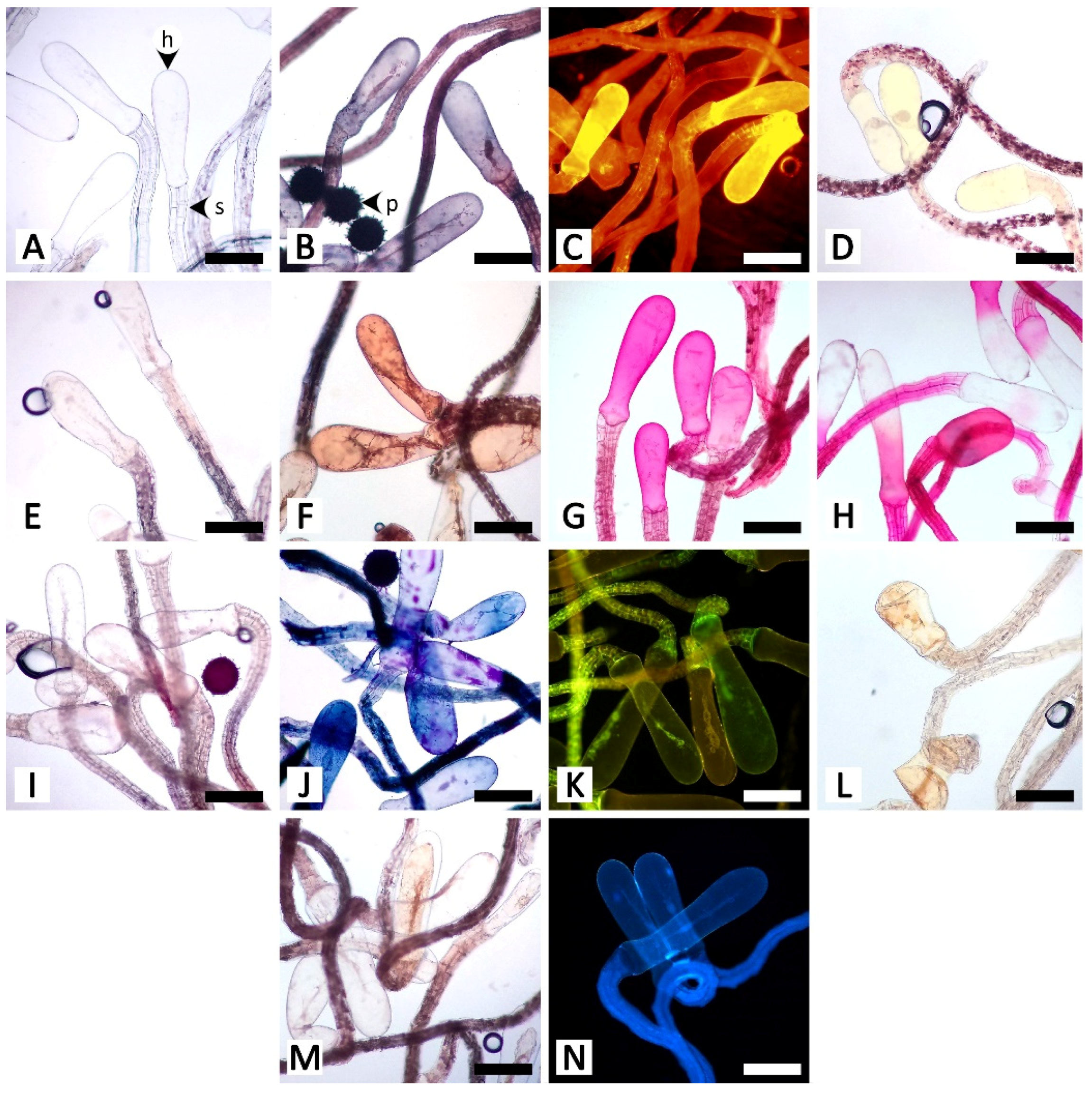
3.2. Histochemistry
3.3. Preliminary Observations of Rivea ornata Visitors
4. Discussion
4.1. Floral Secretory Structures Potentially Important to Pollination
4.2. Secretory Structures Potentially Important to Defense
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Approval
References
- Zahedi, S.M.; Karimi, M.; Venditti, A. Plants adapted to arid areas: Specialized metabolites. Nat. Prod. Res. 2021, 35, 3314–3331. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Jiang, J.-G. Osmotic adjustment and plant adaptation to environmental changes related to drought and salinity. Environ. Rev. 2010, 18, 309–319. [Google Scholar] [CrossRef]
- Halitschke, R.; Stenberg, J.A.; Kessler, D.; Kessler, A.; Baldwin, I.T. Shared signals–‘alarm calls’ from plants increase apparency to herbivores and their enemies in nature. Ecol. Lett. 2008, 11, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Willmer, P. Pollination and Floral Ecology; Princeton University Press: New Jersey, NY, USA, 2011. [Google Scholar]
- Opedal, Ø.H. The evolvability of animal-pollinated flowers: Towards predicting adaptation to novel pollinator communities. New Phytol. 2019, 221, 1128–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, R.D.; Peakall, R.; van der Niet, T.; Johnson, S.D. Niche perspectives on plant–pollinator interactions. Trends Plant Sci. 2020, 25, 779–793. [Google Scholar] [CrossRef] [PubMed]
- Hanley, M.E.; Lamont, B.B.; Fairbanks, M.M.; Rafferty, C.M. Plant structural traits and their role in anti-herbivore defence. Perspect. Plant Ecol. Evol. Syst. 2007, 8, 157–178. [Google Scholar] [CrossRef]
- War, A.R.; Taggar, G.K.; Hussain, B.; Taggar, M.S.; Nair, R.M.; Sharma, H.C. Plant defence against herbivory and insect adaptations. AoB Plants 2018, 10, ply037. [Google Scholar] [CrossRef]
- Nicolson, S.W. Nectar consumers. In Nectaries and Nectar; Nicolson, S.W., Nepi, M., Pacini, E., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 289–342. [Google Scholar]
- Ensikat, H.-J.; Wessely, H.; Engeser, M.; Weigend, M. Distribution, ecology, chemistry and toxicology of plant stinging hairs. Toxins 2021, 13, 141. [Google Scholar] [CrossRef]
- Ketjarun, K.; Traiperm, P.; Suddee, S.; Watthana, S.; Gale, S.W. Labellar anatomy of the Nervilia plicata complex (Orchidaceae: Epidendroideae) in tropical Asia. Kew Bull. 2019, 74, 1. [Google Scholar] [CrossRef]
- Kowalkowska, A.K.; Pawłowicz, M.; Guzanek, P.; Krawczyńska, A.T. Floral nectary and osmophore of Epipactis helleborine (L.) Crantz (Orchidaceae). Protoplasma 2018, 255, 1811–1825. [Google Scholar] [CrossRef] [Green Version]
- Evert, R.F. Esau’s Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body: Their Structure, Function, and Development; John Wiley & Sons: New Jersey, NY, USA, 2006. [Google Scholar]
- Beck, C.B. An Introduction to Plant Structure and Development: Plant Anatomy for the Twenty-First Century; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Fahn, A. Secretory tissues in vascular plants. New Phytol. 1988, 108, 229–257. [Google Scholar] [CrossRef] [PubMed]
- Crang, R.; Lyons-Sobaski, S.; Wise, R. Plant Anatomy: A Concept-Based Approach to the Structure of Seed Plants; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Staples, G.W.; Brummitt, R.K. Convolvulaceae. In Flowering Plant Families of the World; Heywood, V.H., Brummit, R.K., Culham, A., Seberg, O., Eds.; Royal Botanic Gardens, Kew: London, UK, 2007. [Google Scholar]
- Metcalfe, C.R.; Chalk, L. Convolvulaceae. In Anatomy of the Dicotyledons; Clarendon Press: Oxford, UK, 1950; Volume 2, pp. 954–964. [Google Scholar]
- Kuster, V.C.; da Silva, L.C.; Meira, R.M.S.A.; Azevedo, A.A. Glandular trichomes and laticifers in leaves of Ipomoea pes-caprae and I. imperati (Convolvulaceae) from coastal Restinga formation: Structure and histochemistry. Rev. Bras. Bot. 2016, 39, 1117–1125. [Google Scholar] [CrossRef]
- Traiperm, P.; Chow, J.; Nopun, P.; Staples, G.W.; Swangpol, S.C. Identification among morphologically similar Argyreia (Convolvulaceae) based on leaf anatomy and phenetic analyses. Bot. Stud. 2017, 58, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, D.; de Arruda, E.C.P.; Buril, M.T. Hidden in the rocks: A new species of Evolvulus L.(Convolvulaceae) revealed by anatomy. Brittonia 2020, 72, 282–289. [Google Scholar] [CrossRef]
- Tayade, S.K.; Patil, D.A. Foliar epidermal features and their taxonomic significance in the genus Argyreia Lour. (Convolvulaceae). J. Swamy Bot. Club 2003, 20, 15–18. [Google Scholar]
- Chitchak, N.; Traiperm, P.; Staples, G.; Rattanakrajang, P.; Sumanon, P. Species delimitation of some Argyreia (Convolvulaceae) using phenetic analyses: Insights from leaf anatomical data reveal a new species. Botany 2018, 96, 217–233. [Google Scholar] [CrossRef]
- Ashfaq, S.; Ahmad, M.; Zafar, M.; Sultana, S.; Bahadur, S.; Ullah, F.; Zaman, W.; Ahmed, S.N.; Nazish, M. Foliar micromorphology of Convolvulaceous species with special emphasis on trichome diversity from the arid zone of Pakistan. Flora 2019, 255, 110–124. [Google Scholar] [CrossRef]
- Olaranont, Y.; Stauffer, F.W.; Traiperm, P.; Staples, G.W. Investigation of the black dots on leaves of Stictocardia species (Convolvulaceae) using anatomical and histochemical analyses. Flora 2018, 249, 133–142. [Google Scholar] [CrossRef]
- Paiva, E.A.S.; Martins, L.C. Calycinal trichomes in Ipomoea cairica (Convolvulaceae): Ontogenesis, structure and functional aspects. Aust. J. Bot. 2011, 59, 91–98. [Google Scholar] [CrossRef]
- van Ooststroom, S.J. The Convolvulaceae of Malaysia, IV. Blumea 1943, 5, 339–411. [Google Scholar]
- Staples, G.W. Convolvulaceae. In Flora of Thailand; Santisuk, T., Larsen, K., Eds.; Prachachon: Bangkok, Thailand, 2010; Volume 10, pp. 330–468. [Google Scholar]
- Jirabanjongjit, A.; Traiperm, P.; Sando, T.; Stewart, A.B. Pollination and floral biology of a rare morning glory species endemic to Thailand, Argyreia siamensis. Plants 2021, 10, 2402. [Google Scholar] [CrossRef] [PubMed]
- Galetto, L.; Bernardello, G. Floral nectaries, nectar production dynamics and chemical composition in six Ipomoea species (Convolvulaceae) in relation to pollinators. Ann. Bot. 2004, 94, 269–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Santiago-Hernández, M.H.; Martén-Rodríguez, S.; Lopezaraiza-Mikel, M.; Oyama, K.; González-Rodríguez, A.; Quesada, M. The role of pollination effectiveness on the attributes of interaction networks: From floral visitation to plant fitness. Ecology 2019, 100, e02803. [Google Scholar] [CrossRef] [PubMed]
- Hassa, P.; Traiperm, P.; Stewart, A.B. Pollinator visitation and female reproductive success in two floral color morphs of Ipomoea aquatica (Convolvulaceae). Plant Syst. Evol. 2020, 306, 88. [Google Scholar] [CrossRef]
- Keeler, K.H. The extrafloral nectaries of Ipomoea carnea (Convolvulaceae). Am. J. Bot. 1977, 64, 1182–1188. [Google Scholar] [CrossRef] [Green Version]
- Beckmann Jr, R.L.; Stucky, J.M. Extrafloral nectaries and plant guarding in Ipomoea pandurata (L.) GFW Mey.(Convolvulaceae). Am. J. Bot. 1981, 68, 72–79. [Google Scholar] [CrossRef]
- Keeler, K.H.; Kaul, R.B. Distribution of defense nectaries in Ipomoea (Convolvulaceae). Am. J. Bot. 1984, 71, 1364–1372. [Google Scholar] [CrossRef]
- Martins, F.M.; Lima, J.F.; Mascarenhas, A.A.S.; Macedo, T.P. Secretory structures of Ipomoea asarifolia: Anatomy and histochemistry. Rev. Bras. Farm. 2012, 22, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Mondal, A.K.; Chakraborty, T. Ant foraging on extrafloral nectaries [EFNs] of Ipomoea pes-caprae (Convolvulaceae) in the dune vegetation: Ants as potential antiherbivore agents. Indian J. Mar. Sci. 2013, 42, 67–74. [Google Scholar]
- Paiva, E.A.S.; Martins, L.C. Structure of the receptacular nectary and circadian metabolism of starch in the ant-guarded plant Ipomoea cairica (C onvolvulaceae). Plant Biol. 2014, 16, 244–251. [Google Scholar] [CrossRef]
- Staples, G.W. A synopsis of Rivea (Convolvulaceae). Edinb. J. Bot. 2007, 64, 213–223. [Google Scholar] [CrossRef]
- Clarke, C.B. India Convolvulaceae. In Flora of British India; Hooker, J.D., Ed.; L. Reeve & Co.: London, UK, 1883; Volume 4, pp. 179–228. [Google Scholar]
- Mill, R.R. Convolvulaceae. In Flora of Bhutan; Grierson, A.J.C., Long, D.G., Eds.; Royal Botanic Gardens: Edinburgh, UK, 1999; Volume 2, pp. 834–862. [Google Scholar]
- Keeler, K.H. The extrafloral nectaries of Ipomoea leptophylla (Convolvulaceae). Am. J. Bot. 1980, 67, 216–222. [Google Scholar] [CrossRef] [Green Version]
- Keeler, K.H.; Kaul, R.B. Morphology and distribution of petiolar nectaries in Ipomoea (Convolvulaceae). Am. J. Bot. 1979, 66, 946–952. [Google Scholar] [CrossRef] [Green Version]
- Bridson, D.M.; Forman, L. Herbarium Handbook; Royal Botanic Gardens, Kew: London, UK, 1998. [Google Scholar]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill Book Company: London, UK, 1940. [Google Scholar]
- Werker, E. Trichome diversity and development. In Advances in Botanical Research; Academic Press: New York, NY, USA, 2000; Volume 31, pp. 1–35. [Google Scholar]
- Brundrett, M.C.; Kendrick, B.; Peterson, C.A. Efficient lipid staining in plant material with Sudan Red 7B or Fluoral Yellow 088 in polyethylene glycol-glycerol. Biotech. Histochem. 1991, 66, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.W. Neutral red as a lipid fluorochrome. Stain Technol. 1970, 45, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Gabe, M. Techniques Histologiques; Masson et Cie: Paris, France, 1968. [Google Scholar]
- Mace, M.E.; Howell, C.R. Histochemistry and identification of condensed tannin precursors in roots of cotton seedlings. Can. J. Bot. 1974, 52, 2423–2426. [Google Scholar] [CrossRef]
- McManus, J.F.A. Histological and histochemical uses of periodic acid. Stain Technol. 1948, 23, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Mazia, D.; Brewer, P.A.; Alfert, M. The cytochemical staining and measurement of protein with mercuric bromphenol blue. Biol. Bull. 1953, 104, 57–67. [Google Scholar] [CrossRef]
- David, R.; Carde, J.P. Coloration différentielle des inclusions lipidiques et terpéniques des pseudophylles du Pin maritime au moyen du réactif nadi. C. R. Acad. Sci. 1964, 258, 1338–1340. [Google Scholar]
- Svendsen, A.B.; Verpoorte, R. Chromatography of Alkaloids; Elsevier Scientific Publishing Company: New York, NY, USA, 1983. [Google Scholar]
- Furr, M.; Mahlberg, P.G. Histochemical analyses of laticifers and glandular trichomes in Cannabis sativa. J. Nat. Prod. 1981, 44, 153–159. [Google Scholar] [CrossRef]
- Jaitrong, W.; Suwannaphak, K.; Samung, Y.; Jeenthong, T. Ants of Thailand; National Science Museum: Pathum Thani, Thailand, 2020. [Google Scholar]
- Suwanphak, K. Thailand Butterfly Guide; Sarakadee: Nonthaburi, Thailand, 2013. [Google Scholar]
- Svedelius, N. Über das postflorale Wachstum der Kelchblätter einiger Convolvulaceen. Flora Oder Allg. Bot. Ztg. 1906, 96, 231–259. [Google Scholar] [CrossRef]
- Leistner, E.; Steiner, U. Fungal origin of ergoline alkaloids present in dicotyledonous plants (Convolvulaceae). In Physiology and Genetics; Schuffler, A., Anke, T., Eds.; Springer: Berlin, Germany, 2009; pp. 197–208. [Google Scholar]
- Leistner, E.; Steiner, U. The genus Periglandula and its symbiotum with morning glory plants (Convolvulaceae). In Physiology and Genetics, 2nd ed.; Anke, T., Schuffler, A., Eds.; Springer: Cham, Switzerland, 2018; pp. 131–147. [Google Scholar]
- Steiner, U.; Leistner, E. Ergoline alkaloids in convolvulaceous host plants originate from epibiotic clavicipitaceous fungi of the genus Periglandula. Fungal Ecol. 2012, 5, 316–321. [Google Scholar] [CrossRef]
- Steiner, U.; Leistner, E. Ergot alkaloids and their hallucinogenic potential in morning glories. Planta Med. 2018, 84, 751–758. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, W.T.; Panaccione, D.G.; Ryan, K.L.; Kaonongbua, W.; Clay, K. Phylogenetic and chemotypic diversity of Periglandula species in eight new morning glory hosts (Convolvulaceae). Mycologia 2015, 107, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Steiner, U.; Hellwig, S.; Ahimsa-Müller, M.A.; Grundmann, N.; Li, S.-M.; Drewke, C.; Leistner, E. The key role of peltate glandular trichomes in symbiota comprising clavicipitaceous fungi of the genus Periglandula and their host plants. Toxins 2015, 7, 1355–1373. [Google Scholar] [CrossRef] [Green Version]
- Gui, M.-Y.; Liu, W.-Z. Programmed cell death during floral nectary senescence in Ipomoea purpurea. Protoplasma 2014, 251, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Rattanakrajang, P.; Traiperm, P.; Staples, G.W. Re-evaluation of generic characters for Blinkworthia (Convolvulaceae) based on morphology and reproductive organ development. Plant Syst. Evol. 2018, 304, 415–429. [Google Scholar] [CrossRef]
- Bernardello, G. A systematic survey of floral nectaries. In Nectaries and Nectar; Nicolson, S.W., Nepi, M., Pacini, E., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 19–128. [Google Scholar]
- Nepi, M. Nectary structure and ultrastructure. In Nectaries and Nectar; Nicolson, S.W., Nepi, M., Pacini, E., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 129–166. [Google Scholar]
- Nepi, M.; Grasso, D.A.; Mancuso, S. Nectar in plant–insect mutualistic relationships: From food reward to partner manipulation. Front. Plant Sci. 2018, 9, 1063. [Google Scholar] [CrossRef] [PubMed]
- Potascheff, C.d.M.; de Brito, V.L.G.; Galetto, L.; Sebbenn, A.M. Nectar features, diurnal and nocturnal pollinators, and male fitness in Qualea grandiflora (Vochysiaceae). Plant Syst. Evol. 2020, 306, 3. [Google Scholar] [CrossRef]
- Tölke, E.D.; Bachelier, J.B.; Lima, E.A.; Galetto, L.; Demarco, D.; Carmello-Guerreiro, S.M. Diversity of floral nectary secretions and structure, and implications for their evolution in Anacardiaceae. Bot. J. Linn. Soc. 2018, 187, 209–231. [Google Scholar] [CrossRef]
- Phukela, B.; Adit, A.; Tandon, R. Evolutionary trends and diversity of major floral nectary types across Solanaceae. Planta 2021, 254, 55. [Google Scholar] [CrossRef] [PubMed]
- Machado, S.R.; Souza, C.V.d. A reduced, yet functional, nectary disk integrates a complex system of floral nectar secretion in the genus Zeyheria (Bignoniaceae). Acta Bot. Bras. 2016, 31, 344–357. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.N.; Li, Y.; Yang, F.S.; Wang, X.Q. Floral nectary, nectar production dynamics, and floral reproductive isolation among closely related species of Pedicularis. J. Integr. Plant Biol. 2016, 58, 178–187. [Google Scholar] [CrossRef]
- Ren, G.; Healy, R.A.; Klyne, A.M.; Horner, H.T.; James, M.G.; Thornburg, R.W. Transient starch metabolism in ornamental tobacco floral nectaries regulates nectar composition and release. Plant Sci. 2007, 173, 277–290. [Google Scholar] [CrossRef]
- Paiva, E.A.S.; Machado, S.R. The floral nectary of Hymenaea stigonocarpa (Fabaceae, Caesalpinioideae): Structural aspects during floral development. Ann. Bot. 2008, 101, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Gudino, W.; Marquez-Guzman, J.; de la Barrera, E. Structural implications for nectar secretion by nectaries of three columnar cacti. Maejo Int. J. Sci. 2015, 9, 156. [Google Scholar] [CrossRef]
- Machado, S.R.; Rodrigues, T.M. Apoplasmic barrier in the extrafloral nectary of Citharexylum myrianthum (Verbenaceae). Planta 2021, 254, 19. [Google Scholar] [CrossRef]
- Rhoades, D.F.; Bergdahl, J.C. Adaptive significance of toxic nectar. Am. Nat. 1981, 117, 798–803. [Google Scholar] [CrossRef]
- Adler, L.S. The ecological significance of toxic nectar. Oikos 2000, 91, 409–420. [Google Scholar] [CrossRef]
- Stevenson, P.C.; Nicolson, S.W.; Wright, G.A. Plant secondary metabolites in nectar: Impacts on pollinators and ecological functions. Funct. Ecol. 2017, 31, 65–75. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- War, A.R.; Buhroo, A.A.; Hussain, B.; Ahmad, T.; Nair, R.M.; Sharma, H.C. Plant defense and insect adaptation with reference to secondary metabolites. In Co-Evolution of Secondary Metabolites; Springer: Cham, Switzerland, 2020; pp. 795–822. [Google Scholar]
- Knudsen, J.T.; Gershenzon, J. The chemical diversity of floral scent. In Biology of Floral Scent; Dudareva, N., Pichersky, E., Eds.; CRC press: Boca Raton, FL, USA, 2006; pp. 27–52. [Google Scholar]
- Raguso, R.A. Why are some floral nectars scented? Ecology 2004, 85, 1486–1494. [Google Scholar] [CrossRef]
- Kelber, A.; Balkenius, A.; Warrant, E.J. Scotopic colour vision in nocturnal hawkmoths. Nature 2002, 419, 922–925. [Google Scholar] [CrossRef]
- Raguso, R.A.; Willis, M.A. Synergy between visual and olfactory cues in nectar feeding by naıve hawkmoths, Manduca sexta. Anim. Behav. 2002, 64, 685–695. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.D.; Balducci, M.G.; Shuttleworth, A. Hawkmoth pollination of the orchid Habenaria clavata: Mechanical wing guides, floral scent and electroantennography. Biol. J. Linn. Soc. 2020, 129, 213–226. [Google Scholar] [CrossRef]
- Dobson, H.E.M. Relationship between floral fragrance composition and type of pollinator. In Biology of Floral Scent; Dudareva, N., Pichersky, E., Eds.; CRC press: Boca Raton, FL, USA, 2006; pp. 147–198. [Google Scholar]
- Liu, C.Q.; Sun, H. Pollination in Lilium sargentiae (Liliaceae) and the first confirmation of long-tongued hawkmoths as a pollinator niche in Asia. J. Syst. Evol. 2019, 57, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Steen, R.; Norli, H.R.; Thöming, G. Volatiles composition and timing of emissions in a moth-pollinated orchid in relation to hawkmoth (Lepidoptera: Sphingidae) activity. Arthropod Plant Interact. 2019, 13, 581–592. [Google Scholar] [CrossRef]
- Gibb, T.J.; Oseto, C. Insect Collection and Identification: Techniques for the Field and Laboratory; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Vlasáková, B.; Pinc, J.; Jůna, F.; Kotyková Varadínová, Z. Pollination efficiency of cockroaches and other floral visitors of Clusia blattophila. Plant Biol. 2019, 21, 753–761. [Google Scholar] [CrossRef]
- Xiong, W.; Ollerton, J.; Liede-Schumann, S.; Zhao, W.; Jiang, Q.; Sun, H.; Liao, W.; You, W. Specialized cockroach pollination in the rare and endangered plant Vincetoxicum hainanense in China. Am. J. Bot. 2020, 107, 1355–1365. [Google Scholar] [CrossRef] [PubMed]
- Schaffner, G. Extraflorale Nektarien bei Cuscuta. Ber. Deutsch. Bot. Ges. 1979, 92, 721–729. [Google Scholar] [CrossRef]
- Clayson, C.; García-Ruiz, I.; Costea, M. Diversity, evolution, and function of stomata bearing structures in Cuscuta (dodders, Convolvulaceae): From extrafloral nectar secretion to transpiration in arid conditions. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 310–321. [Google Scholar] [CrossRef]
- Blüthgen, N.; Reifenrath, K. Extrafloral nectaries in an Australian rainforest: Structure and distribution. Aust. J. Bot. 2003, 51, 515–527. [Google Scholar] [CrossRef]
- Coutinho, Í.A.C.; Valente, V.M.M.; Meira, R.M.S.A. Ontogenetic, anatomical and histochemical study of the extrafloral nectaries of Sapium biglandulosum (Euphorbiaceae). Aust. J. Bot. 2010, 58, 224–232. [Google Scholar] [CrossRef]
- Pires, M.S.; Calixto, E.S.; Oliveira, D.C.; Del-Claro, K. A new extrafloral nectary-bearing plant species in the Brazilian savanna and its associated ant community: Nectary structure, nectar production and ecological interactions. Sociobiology 2017, 64, 228–236. [Google Scholar] [CrossRef]
- Machado, S.R.; Morellato, L.P.C.; Sajo, M.G.; Oliveira, P.S. Morphological patterns of extrafloral nectaries in woody plant species of the Brazilian cerrado. Plant Biol. 2008, 10, 660–673. [Google Scholar] [CrossRef] [PubMed]
- Craven, L.A.; Miller, C.; White, R.G. A new name, and notes on extra-floral nectaries, in Lagunaria (Malvaceae, Malvoideae). Blumea 2006, 51, 345–353. [Google Scholar] [CrossRef] [Green Version]
- Silva, E.O.; Feio, A.C.; Cardoso-Gustavson, P.; Milward-De-Azevedo, M.A.; Dos Santos, J.U.M.; De Aguiar Dias, A.C.A. Extrafloral nectaries and plant–insect interactions in Passiflora L.(Passifloraceae). Rev. Bras. Bot. 2017, 40, 331–340. [Google Scholar] [CrossRef]
- Twyford, A.D.; Caola, A.M.; Choudhary, P.; Raina, R.; Friedman, J. Loss of color pigmentation is maintained at high frequency in a monkey flower population. Am. Nat. 2018, 191, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Del Valle, J.C.; Alcalde-Eon, C.; Escribano-Bailón, M.; Buide, M.; Whittall, J.B.; Narbona, E. Stability of petal color polymorphism: The significance of anthocyanin accumulation in photosynthetic tissues. BMC Plant Biol. 2019, 19, 496. [Google Scholar] [CrossRef]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.; Visser, R.G.; Bovy, A. Anthocyanin biosynthesis and degradation mechanisms in Solanaceous vegetables: A review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef]
- Vezza, M.; Nepi, M.; Guarnieri, M.; Artese, D.; Rascio, N.; Pacini, E. Ivy (Hedera helix L.) flower nectar and nectary ecophysiology. Int. J. Plant Sci. 2006, 167, 519–527. [Google Scholar] [CrossRef]
- Ballego-Campos, I.; Forzza, R.C.; Paiva, E.A. More than scales: Evidence for the production and exudation of mucilage by the peltate trichomes of Tillandsia cyanea (Bromeliaceae: Tillandsioideae). Plants 2020, 9, 763. [Google Scholar] [CrossRef] [PubMed]
- Rico-Gray, V.; Oliveira, P.S. The Ecology and Evolution of Ant-Plant Interactions; University of Chicago Press: Chicago, IL, USA, 2008. [Google Scholar]
- Mesquita-Neto, J.N.; Paiva, E.A.S.; Galetto, L.; Schlindwein, C. Nectar secretion of floral buds of Tococa guianensis mediates interactions with generalist ants that reduce florivory. Front. Plant Sci. 2020, 11, 627. [Google Scholar] [CrossRef]
- Anjos, D.V.; Caserio, B.; Rezende, F.T.; Ribeiro, S.P.; Del-Claro, K.; Fagundes, R. Extrafloral-nectaries and interspecific aggressiveness regulate day/night turnover of ant species foraging for nectar on Bionia coriacea. Austral Ecol. 2017, 42, 317–328. [Google Scholar] [CrossRef]
- Fagundes, R.; Dáttilo, W.; Ribeiro, S.; Rico-Gray, V.; Jordano, P.; Del-Claro, K. Differences among ant species in plant protection are related to production of extrafloral nectar and degree of leaf herbivory. Biol. J. Linn. Soc. 2017, 122, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, T.V.; Izzo, T.J.; Teixido, A.L. Safe sex: Ant defense does not interfere with pollination in passion flowers. Acta Bot. Bras. 2021, 35, 290–297. [Google Scholar] [CrossRef]
- Macêdo, L.P.M.; Silva, E.O.; de Aguiar-Dias, A.C.A. Morphoanatomy and ecology of the extrafloral nectaries in two species of Passiflora L.(Passifloraceae). S. Afr. J. Bot. 2021, 143, 248–255. [Google Scholar] [CrossRef]
- Santos, A.T.F.; Leal, L.C. My plant, my rules: Bodyguard ants of plants with extrafloral nectaries affect patterns of pollinator visits but not pollination success. Biol. J. Linn. Soc. 2019, 126, 158–167. [Google Scholar] [CrossRef]
- Yamawo, A.; Suzuki, N.; Tagawa, J. Extrafloral nectary-bearing plant Mallotus japonicus uses different types of extrafloral nectaries to establish effective defense by ants. J. Plant Res. 2019, 132, 499–507. [Google Scholar] [CrossRef]
- Martins, J.K.S.S.; Carneiro, A.; Souza, L.; Almeida-Cortez, J. How pollinator visits are affected by flower damage and ants presence in Ipomoea carnea subs. fistulosa (Martius and Choise) (Convolvulaceae)? Braz. J. Biol. 2019, 80, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Villamil, N.; Boege, K.; Stone, G.N. Ant-pollinator conflict results in pollinator deterrence but no nectar trade-offs. Front. Plant Sci. 2018, 9, 1093. [Google Scholar] [CrossRef] [PubMed]
- Olaranont, Y.; Stewart, A.B.; Songnuan, W.; Traiperm, P. How and where Periglandula fungus interacts with different parts of Ipomoea asarifolia. J. Fungi 2022, 8, 823. [Google Scholar] [CrossRef]
- Fürstenberg-Hägg, J.; Zagrobelny, M.; Bak, S. Plant defense against insect herbivores. Int. J. Mol. Sci. 2013, 14, 10242–10297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; van Doan, C.; Arce, C.C.M.; Hu, L.; Gruenig, S.; Parisod, C.; Hibbard, B.E.; Hervé, M.R.; Nielson, C.; Robert, C.A.M. Plant defense resistance in natural enemies of a specialist insect herbivore. Proc. Natl. Acad. Sci. USA 2019, 116, 23174–23181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heron, H.D.C. Tortoise beetles (Chrysomelidae: Cassidinae) and their feeding patterns from the North Park Nature Reserve, Durban, KwaZulu-Natal, South Africa. Durb. Mus. Novit. 2003, 28, 31–44. [Google Scholar]
- Dube, Z.P.; Visser, D.; Grobbelaar, E. Aspidimorpha (Megaspidomorpha) angolensis Weise (Coleoptera: Chrysomelidae: Cassidinae): A potential new pest of sweet potato (Ipomoea batatas) in South Africa. J. Plant Dis. Prot. 2020, 127, 119–122. [Google Scholar] [CrossRef]
- Heron, H.D.C. The life history of Aspidimorpha areata (KLUG, 1835) (Coleoptera: Chrysomelidae: Cassidinae). Afr. Entomol. 2007, 15, 75–87. [Google Scholar] [CrossRef]
- Leong, T.M.; D’Rozario, V. Mimicry of the weaver ant Oecophylla smaragdina by the moth caterpillar, Homodes bracteigutta, the crab spider, Amyciaea lineatipes, and the jumping spider, Myrmarachne plataleoides. NiS 2012, 5, 39–56. [Google Scholar]
- Shelford, R. Observations on some mimetic insects and spiders from Borneo and Singapore, with appendices containing descriptions of new species. In Proceedings of the General Meetings for Scientific Business of the Zoological Society of London; Messrs. Longmans, Green, & Co.: London, UK, 1902; Volume 2, pp. 230–284. [Google Scholar]
- Shelford, R.W.C. A Naturalist in Borneo; T. Fisher Unwin Ltd.: London, UK, 1916. [Google Scholar]
- Kalshoven, L.G.E. Larvae of Homodes mimicking the aggressive Oecophylla ant in Southeast Asia (Lepidoptera, Noctuidae). Tijdschr. Entomol. 1961, 104, 43–50. [Google Scholar]
- Common, I.F.B. Moths of Australia; Melbourne University Press: Carlton, Victoria, 1990. [Google Scholar]
- Holloway, J.D. The moths of Borneo: Family Noctuidae, subfamily Catocalinae. Malay. Nat. J. 2005, 58, 1–529. [Google Scholar]
- Gurevitch, J.; Scheiner, S.M.; Fox, G.A. The Ecology of Plants; Sinauer Associates: Sunderland, UK, 2002. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite Group | Test | Positive Chromatic Reaction | Nectary Disc | Petiolar Nectaries | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Glandular Trichome | Epidermis | Nectariferous Parenchyma | |||||||||
| Head | Stalk | Base | |||||||||
| Total lipids | Sudan black B | Dark blue to black | +, s | +, s | +, s | - | +, s | - | |||
| Neutral red | Yellow | + | +, s | +, s | - | +, s | - | ||||
| General phenolic compounds | Ferric chloride | Brown or black | +, c | +, c | - | - | +, c | +, c | |||
| Potassium dichromate | Brown | +, c | +, c | - | - | - | - | ||||
| Starch | Lugol’s iodine | Dark blue to black | +, c | - | - | - | - | - | |||
| Neutral polysaccharides | Periodic acid–Schiff’s reagent (PAS) | Pink | +, c, s | +, c, s | +, s | +, s | +, c, s | +, c, s | |||
| Acidic polysaccharides | Ruthenium red | Pink to red | +, c, s | +, c, s | +, s | +, s | +, c, s | +, c, s | |||
| Proteins | Mercuric bromophenol blue | Blue | - | - | - | - | - | - | |||
| Terpenoids | Nadi reagent | Dark blue or violet | +, c | +, c | - | - | +, c | +, c | |||
| Flavonoids | Naturstoff reagent A | Yellow | +, c | +, s | - | - | +, s | - | |||
| Alkaloids | Dragendorff reagent | Reddish brown | +, c | +, c | - | - | - | - | |||
| Wagner’s reagent | Reddish brown | +, c | +, c | - | - | +, c | +, c | ||||
| Metabolite Group | Test | Positive Chromatic Reaction | Calycinal Glands | Foliar Glands | Staminal Hairs | ||||||
| Head | Stalk | Base | Head | Stalk | Base | Head | Stalk | ||||
| Total lipids | Sudan black B | Dark blue to black | +, s | +, s | - | + | + | - | + | + | |
| Neutral red | Yellow | - | +, s | - | - | +, s | - | + | + | ||
| General phenolic compounds | Ferric chloride | Brown or black | +, c | +, c | - | +, c | + | - | - | +, c | |
| Potassium dichromate | Brown | +, c | +, c | - | +, c | - | - | - | +, c | ||
| Starch | Lugol’s iodine | Dark blue to black | - | - | - | - | - | - | - | - | |
| Neutral polysaccharides | Periodic acid–Schiff’s reagent (PAS) | Pink | + | + | + | +, c, s | +, s | +, s | +, s | +, s | |
| Acidic polysaccharides | Ruthenium red | Pink to red | +, c, s | +, c, s | +, s | +, s | +, s | +, s | +, s | +, s | |
| Proteins | Mercuric bromophenol blue | Blue | - | - | - | - | - | - | - | - | |
| Terpenoids | Nadi reagent | Dark blue or violet | + | + | +, c | + | + | - | +, c | +, c | |
| Flavonoids | Naturstoff reagent A | Yellow | +, c, s | +, c, s | - | + | + | - | +, c | + | |
| Alkaloids | Dragendorff reagent | Reddish brown | +, c | +, c | - | +, c | +, s | - | +, c | + | |
| Wagner’s reagent | Reddish brown | +, c | +, c | - | +, c | - | - | +, c | +, c | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chitchak, N.; Stewart, A.B.; Traiperm, P. Functional Ecology of External Secretory Structures in Rivea ornata (Roxb.) Choisy (Convolvulaceae). Plants 2022, 11, 2068. https://doi.org/10.3390/plants11152068
Chitchak N, Stewart AB, Traiperm P. Functional Ecology of External Secretory Structures in Rivea ornata (Roxb.) Choisy (Convolvulaceae). Plants. 2022; 11(15):2068. https://doi.org/10.3390/plants11152068
Chicago/Turabian StyleChitchak, Natthaphong, Alyssa B. Stewart, and Paweena Traiperm. 2022. "Functional Ecology of External Secretory Structures in Rivea ornata (Roxb.) Choisy (Convolvulaceae)" Plants 11, no. 15: 2068. https://doi.org/10.3390/plants11152068
APA StyleChitchak, N., Stewart, A. B., & Traiperm, P. (2022). Functional Ecology of External Secretory Structures in Rivea ornata (Roxb.) Choisy (Convolvulaceae). Plants, 11(15), 2068. https://doi.org/10.3390/plants11152068