
Polyphenolic Compound Variation in Globe Artichoke Cultivars as Affected by Fertilization and Biostimulants Application

 ,
, 
 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Caffeoylquinic Acids Content
2.2. Flavonoids: Luteolin and Apigenin Content
3. Materials and Methods
3.1. Analysis of Polyphenols
3.1.1. Reagents and Solvents
3.1.2. Extraction Procedure
3.1.3. HPLC-DAD Analysis
3.1.4. Qualitative and Quantitative Analysis
3.2. HPLC-DAD-ESI-MS/MS Analysis
3.3. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lattanzio, V.; Kroon, P.A.; Linsalata, V.; Cardinali, A. Globe artichoke: A functional food and source of nutraceutical ingredients. J. Funct. Foods 2009, 1, 131–144. [Google Scholar] [CrossRef]
- Sonnante, G.; Pignone, D.; Hammer, K. The Domestication of Artichoke and Cardoon: From Roman Times to the Genomic Age. Ann. Bot. 2007, 100, 1095–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. Food, Trade Statistics. Available online: http://www.fao.org/faostat (accessed on 29 May 2022).
- Wang, M.; Simon, J.E.; Aviles, I.F.; He, K.; Zheng, Q.Y.; Tadmor, Y. Analysis of antioxidative phenolic compounds in artichoke (Cynara scolymus L.). J. Agric. Food Chem. 2003, 51, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Sonnante, G.; D’Amore, R.; Blanco, E.; Pierri, C.L.; De Palma, M.; Luo, J.; Tucci, M.; Martin, C. Novel hydroxycinnamoyl-Coenzyme A quinate transferase genes from artichoke are involved in the synthesis of chlorogenic acid. Plant Physiol. 2010, 153, 1224–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnante, G.; Gatto, A.; Morgese, A.; Montemurro, F.; Sarli, G.; Blanco, E.; Pignone, D. Genetic map of artichoke × wild cardoon: Toward a consensus map for Cynara cardunculus. Theor. Appl. Genet. 2011, 123, 1215–1229. [Google Scholar] [CrossRef] [PubMed]
- Negro, D.; Montesano, V.; Grieco, S.; Crupi, P.; Sarli, G.; De Lisi, A.; Sonnante, G. Polyphenol Compounds in Artichoke Plant Tissues and Varieties. J. Food Sci. 2012, 77, C244–C252. [Google Scholar] [CrossRef] [PubMed]
- Fratianni, F.; Tucci, M.; De Palma, M.; Pepe, R.; Nazzaro, F. Polyphenolic composition in different parts of some cultivars of globe artichoke (Cynara cardunculus L. var. scolymus (L.) Fiori). Food Chem. 2007, 104, 1282–1286. [Google Scholar] [CrossRef]
- Romani, A.; Pinelli, P.; Cantini, C.; Cimato, A.; Heimler, D. Characterization of Violetto di Toscana, a typical Italian variety of artichoke (Cynara scolymus L.). Food Chem. 2006, 95, 221–225. [Google Scholar] [CrossRef]
- Pandino, G.; Lombardo, S.; Mauromicale, G. Mineral profile in globe artichoke as affected by genotype, head part and environment. J. Sci. Food Agric. 2011, 91, 302–308. [Google Scholar] [CrossRef] [PubMed]
- United Nations Educational Scientific and Cultural Organization. UNESCO Intangible Heritage Lists. 2010. Available online: http://www.unesco.org (accessed on 10 May 2022).
- De Falco, B.; Incerti, G.; Amato, M.; Lanzotti, V. Artichoke: Botanical, agronomical, phytochemical, and pharmacological overview. Phytochem. Rev. 2015, 14, 993–1018. [Google Scholar] [CrossRef]
- Huang, M.T.; Ferraro, T. Phenolic compounds in food and cancer prevention. In Phenolic Compounds in Food and Their Effects on Health. II. Antioxidants and Cancer Prevention; American Chemical Society: Washington, DC, USA, 1992; pp. 8–34. [Google Scholar]
- Halliwell, B. Free radicals, antioxidants and human disease: Curiosity, cause, or consequence. Lancet 1994, 344, 721–724. [Google Scholar] [CrossRef]
- Hung, H.C.; Joshipura, K.J.; Jiang, R.; Hu, F.B.; Hunter, D.; Smith-Warner, S.A. Fruit and vegetable intake and risk of major chronic diseases. J. Natl. Cancer Inst. 2004, 96, 1577–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adzet, T.; Camarasa, J.; Carlos Laguna, J. Hepatoprotective activity of polyphenolic compounds from Cynara scolymus against CCl4 toxicity in isolated rat hepatocytes. J. Nat. Prod. 1987, 50, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, R. Antioxidative and protective properties of extracts from leaves of the artichoke (Cynara scolymus L.) against hydroperoxide-induced oxidative stress in cultured rat hepatocytes. Toxicol. Appl. Pharmacol. 1997, 144, 279–286. [Google Scholar] [CrossRef]
- Perez-Garcia, F.; Adzet, T.; Canigueral, S. Activity of artichoke leaf extract on reactive oxygen species in human leukocytes. Free. Radic. Res. 2000, 33, 661–665. [Google Scholar] [CrossRef]
- Schütz, K.; Kammerer, D.; Carle, R.; Schieber, A. Identification and quantification of caffeoylquinic acids and flavonoids from artichoke (Cynara scolymus L.) heads, juice, and pomace by HPLC-DAD-ESI/MSn. J. Agric. Food Chem. 2004, 52, 4090–4096. [Google Scholar] [CrossRef]
- Lombardo, S.; Pandino, G.; Mauromicale, G.; Knödler, M.; Carle, R.; Schieber, A. Influence of genotype, harvest time and plant part on polyphenolic composition of globe artichoke [Cynara cardunculus L. var. scolymus (L.) Fiori]. Food Chem. 2010, 119, 1175–1181. [Google Scholar] [CrossRef]
- Lombardo, S.; Pandino, G.; Mauro, R.; Mauromicale, G. Variation of phenolic content in globe artichoke in relation to biological, technical and environmental factors. Ital. J. Agron. 2009, 4, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Sonnante, G.; De Paolis, A.; Pignone, D. Relationships among artichoke cultivars and some related wild taxa based on AFLP markers. Plant Genet. Resour. 2004, 1, 125–133. [Google Scholar] [CrossRef]
- Pavan, S.; Curci, P.L.; Zuluaga, D.L.; Blanco, E.; Sonnante, G. Genotyping-by-sequencing highlights patterns of genetic structure and domestication in artichoke and cardoon. PLoS ONE 2018, 13, e0205988. [Google Scholar] [CrossRef]
- De Martonne, E. Aridité et indices d’aridité. Académie Sci. Comptes Rendus 1923, 182, 1935–1938. [Google Scholar]
- Pandino, G.; Courts, F.L.; Lombardo, S.; Mauromicale, G.; Williamson, G. Caffeoylquinic acids and flavonoids in the immature inflorescence of globe artichoke, wild cardoon, and cultivated cardoon. J. Agric. Food Chem. 2010, 58, 1026–1031. [Google Scholar] [CrossRef] [PubMed]
- Pandino, G.; Lombardo, S.; Mauromicale, G.; Williamson, G. Profile of polyphenols and phenolic acids in bracts and receptacles of globe artichoke (Cynara cardunculus var. scolymus) germplasm. J. Food Compos. Anal. 2011, 24, 148–153. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures in Agricultural Research, 2nd ed.; John Wiley & Sons: New York, NY, USA; Chichester, UK, 1984. [Google Scholar]
- Pandino, G.; Lombardo, S.; Mauromicale, G. Globe artichoke leaves and floral stems as a source of bioactive compounds. Ind. Crop Prod. 2013, 44, 44–49. [Google Scholar] [CrossRef]
- Negro, D.; Montesano, V.; Sonnante, G.; Rubino, P.; de Lisi, A.; Sarli, G. Fertilization strategies on cultivars of globe artichoke: Effects on yield and quality performance. J. Plant Nutr. 2016, 39, 279–287. [Google Scholar] [CrossRef]
- D’Antuono, I.; Garbetta, A.; Linsalata, V.; Minervini, F.; Cardinali, A. Polyphenols from artichoke heads (Cynara cardunculus L.) subsp. scolymus Hayek): In vitro bio-accessibility, intestinal uptake and bioavailability. Food Funct. 2015, 6, 1268–1277. [Google Scholar] [CrossRef]
- Tulipani, S.; Mezzetti, B.; Capocasa, F.; Bompadre, S.; Beekwilder, J.; De Vos, C.H.R.; Capanoglu, E.; Bovy, A.; Battino, M. Antioxidants, phenolic compounds, and nutritional quality of different strawberry genotypes. J. Agric. Food Chem. 2008, 56, 696–704. [Google Scholar] [CrossRef]
- Veberic, R.; Colaric, M.; Stampar, F. Phenolic acids and flavonoids of fruits (Ficus carica L.) in the northern Mediterranean region. Food Chem. 2008, 106, 153–157. [Google Scholar] [CrossRef]
- Andre, C.M.; Ghislain, M.; Bertin, P.; Oufir, M.; Del Rosario Herrera, M.; Hoffmann, L.; Hausman, J.F.; Larondelle, Y.; Evers, D. Andean potato cultivars (Solanum tuberosum L.) as a source of antioxidant and mineral micronutrients. J. Agric. Food Chem. 2007, 55, 366–378. [Google Scholar] [CrossRef]
- Imeh, U.; Khokhar, S. Distribution of conjugated and free phenols in fruits: Antioxidant activity and cultivar variations. J. Agric. Food Chem. 2002, 50, 6301–6306. [Google Scholar] [CrossRef]
- Carle, R.; Gomaa, K. Chamomile: A pharmacological and clinical profile. Drugs Today 1992, 28, 559–565. [Google Scholar]
- Bendini, A.; Cerretani, L.; Carrasco-Pancorbo, A.; Gómez-Caravaca, A.M.; Segura-Carretero, A.; Fernández-Gutiérrez, A.; Lercker, G. Phenolic Molecules in Virgin Olive Oils: A Survey of Their Sensory Properties, Health Effects, Antioxidant Activity and Analytical Methods. An Overview of the Last Decade. Molecules 2007, 12, 1679–1719. [Google Scholar] [CrossRef] [PubMed]
- Heilmann, J.; Merfort, I.; Weiss, M. Radical scavenger activity of different 3,4-dihydroxyflavonols and 1,5-dicaffeoylquinic acid studied by inhibition of chemiluminescence. Planta Med. 1995, 61, 435–438. [Google Scholar] [CrossRef]
- Fukumoto, L.R.; Mazza, G. Assessing antioxidant and prooxidant activities of phenolic compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. Soc. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azzini, E.; Bugianesi, R.; Romano, F.; Di Venere, D.; Miccadei, S.; Durazzo, A.; Foddai, M.S.; Catasta, G.; Linsalata, V.; Maiani, G. Absorption and metabolism of bioactive molecules after oral consumption of cooked edible heads of Cynara scolymus L. (cultivar Violetto di Provenza) in human subjects: A pilot study. Br. J. Nutr. 2007, 97, 963–969. [Google Scholar] [CrossRef] [Green Version]
- Justesen, U.; Knuthsen, P.; Leth, T. Quantitative analysis of flavonols, flavones, and flavanones in fruits, vegetables and beverages by high-performance liquid chromatography with photo-diode array and mass spectrometric detection. J. Chromatogr. A 1998, 799, 101–110. [Google Scholar] [CrossRef]
- Lattanzio, V.; Cardinali, A.; Di Venere, D.; Linsalata, V.; Palmieri, S. Browning phenomena in stored artichoke (Cynara scolymus L.) heads: Enzymic or chemical reactions? Food Chem. 1994, 50, 1–7. [Google Scholar] [CrossRef]
- Lattanzio, V. Bioactive polyphenols: Their role in quality and storability of fruit and vegetables. J. Appl. Bot. 2003, 77, 128–146. [Google Scholar]
- Nichiforesco, E.V.; Coucou, V. Variation of caffeic acid type o-dihydroxyphenolic derivatives of the artichoke (Cynara scolymus L.) during its period of vegetation. Ann. Pharm. Fr. 1966, 24, 451–456. [Google Scholar]
- Lattanzio, V.; Van Sumere, C.F. Changes in phenolic compounds during the development and cold storage of artichoke (Cynara scolymus L.) heads. Food Chem. 1987, 24, 37–50. [Google Scholar] [CrossRef]
- Faulds, C.B.; Willimason, G. The role of hydroxycinnamates in the plant cell wall. J. Sci. Food Agric. 1999, 79, 393–395. [Google Scholar] [CrossRef]
- Peschel, W.; Sánchez-Rabaneda, F.; Diekmann, W.; Plescher, A.; Gartzía, I.; Jiménez, D.; Lamuela-Raventos, R.; Buxaderas, S.; Codina, C. An industrial approach in the search of natural antioxidants from vegetable and fruit wastes. Food Chem. 2006, 97, 137–150. [Google Scholar] [CrossRef]
- Blanco E, Musio B, Todisco S, Mastrorilli P, Gallo V, Sonnante G, Non-targeted NMR approach to unveil and promote the biodiversity of globe artichoke in the Mediterranean area. J. Food Compos. Anal. 2022, 110, 104539. [CrossRef]
- Alamanni, M.C.; Cossu, M. Antioxidant activity of the extracts of the edible part of artichoke (Cynara scolymus L.) var. Spinososardo. Ital. J. Food Sci. 2003, 15, 187–195. [Google Scholar]
- Curadi, M.; Picciarelli, P.; Lorenzi, R.; Graifenberg, A.; Ceccarelli, N. Antioxidant activity and phenolic compounds in the edible parts of early and late Italian artichoke (Cynara scolymus L.) varieties. Ital. J. Food Sci. 2005, 17, 33–44. [Google Scholar]
- Eich, J.; Baier, C.; Grun, M.; Wagenbreth, D.; Zimmermann, R. Artichoke leaves used for herbal drug production: Influence of nitrogen fertilization on yield and on pharmaceutical quality. Acta Hortic. 2005, 681, 545–554. [Google Scholar]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenyl-propanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Kirakosyan, A.; Kaufman, P.; Warber, S.; Zick, S.; Aaronson, K.; Bolling, S.; Chang, S.C. Applied environmental stresses to enhance the levels of polyphenolics in leaves of hawthorn plants. Physiol. Plant 2004, 121, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Gil-Izqierdo, A.; Gil, M.I.; Conesa, M.A.; Ferreres, F. The effect of storage temperature on vitamin C and phenolics content of artichoke (Cynara scólymus L.) heads. Innov. Food Sci. Emerg. Technol. 2001, 2, 199–202. [Google Scholar] [CrossRef]
- Llorach, R.; Espin, J.C.; Tomás-Barberán, F.A.; Ferreres, F. Artichoke (Cynara scolymus L.) byproducts as a potential source of health-promoting antioxidant phenolics. J. Agric. Food Chem. 2002, 50, 3458–3464. [Google Scholar] [CrossRef]
- Di Venere, D.; Sergio, L.; Cardinali, A.; Linsalata, V.; Pieralice, M.; Massignan, L.; Lovino, R. Biochemical changes during modified atmosphere storage of artichoke heads. Acta Hortic. 2004, 660, 587–592. [Google Scholar] [CrossRef]


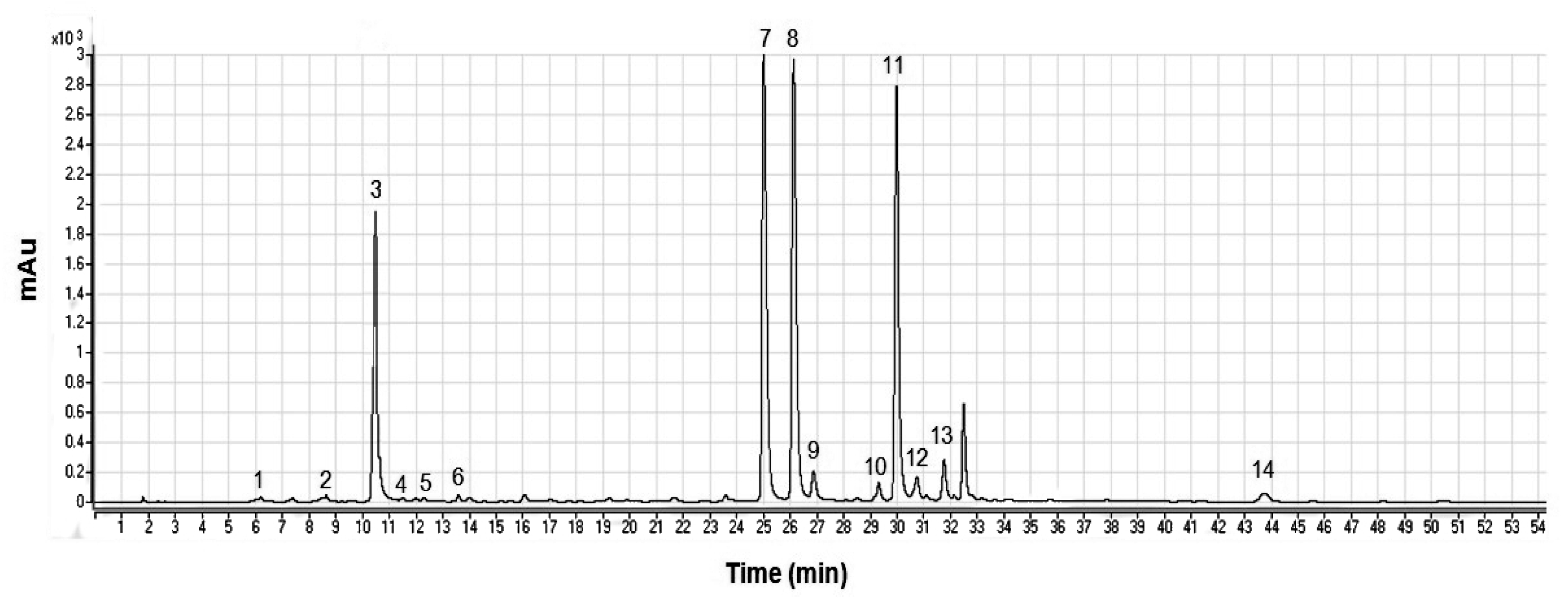{kind=link}
| No. | Compound | Retention Time (min) | HPLC-DAD λmax (nm) | [M-H]− a m/z | MS/MS b m/z | Comparison with Standard |
|---|---|---|---|---|---|---|
| 1 | 1-O-Caffeoylquinic acid | 6.21 | 233, 305sh, 328 | 353 | 191 | no |
| 2 | 3-O-Caffeoylquinic acid | 8.65 | 241, 303sh, 325 | 353 | 191 | no |
| 3 | 5-O-Caffeoylquinic acid | 10.48 | 241, 305sh, 326 | 353 | 191 | yes |
| 4 | 4-O-Caffeoylquinic acid | 11.51 | 236, 303sh, 326 | 353 | 191 | no |
| 5 | Caffeic acid | 12.30 | 237, 302sh, 323 | 179 | 135 | yes |
| 6 | 1,3-di-O-Caffeoylquinic acid | 13.59 | 242, 307sh, 322 | 515 | 353 | yes |
| 7 | Luteolin 7-O-rutinoside | 25.01 | 256, 266sh, 350 | 593 | 285 | no |
| 8 | Luteolin 7-O-glucoside | 26.13 | 253, 266sh, 347 | 447 | 285 | yes |
| 9 | Luteolin 7-O-glucuronide | 26.88 | 254, 268sh, 343 | 461 | 285 | no |
| 10 | Apigenin 7-O-rutinoside | 29.32 | 249, 304sh, 328 | 577 | 269 | no |
| 11 | 1,5-di-O-Caffeoylquinic acid | 29.97 | 243, 303sh, 329 | 515 | 353 | yes |
| 12 | Apigenin 7-O-glucoside | 30.74 | 229, 266, 339 | 431 | 269 | yes |
| 13 | Apigenin 7-O-glucuronide | 31.76 | 267, 335 | 445 | 269 | no |
| 14 | Luteolin | 43.74 | 254, 266sh, 347 | 285 | no |
| 1-Caffeoylquinic Acid | 3-Caffeoylquinic Acid | 4-Caffeoylquinic Acid | 5-Caffeoylquinic Acid | Caffeic Acid | 1,3-Dicaffeoylquinic Acid | 1,5-Dicaffeoylquinic Acid | Total | |
|---|---|---|---|---|---|---|---|---|
| Year | ** | ** | ** | ** | ** | ** | ** | ** |
| 2016 | 11.99 b | 7.69 b | 16.91 b | 255.09 b | 43.94 b | 13.01 a | 16,781.58 b | 17,130.22 b |
| 2017 | 18.71 a | 9.16 a | 35.54 a | 531.52 a | 80.51 a | 5.62 b | 26,051.68 a | 26,732.74 a |
| Cultivar | * | * | * | * | ||||
| Opal | 19.05 | 9.26 a | 32.08 | 497.25 a | 60.92 | 9.00 | 24,970.34 a | 25,597.90 a |
| Madrigal | 11.65 | 7.58 b | 20.38 | 289.36 b | 63.54 | 9.64 | 17,862.91 b | 18,265.06 b |
| Nitrogen Rate (kg/ha) | ||||||||
| 0 | 14.44 | 8.39 | 31.05 | 418.12 | 66.72 | 9.62 | 24,528.32 | 25,076.66 |
| 50 | 13.02 | 8.29 | 22.109 | 302.93 | 56.44 | 9.06 | 17,974.98 | 18,386.80 |
| 100 | 16.81 | 8.94 | 29.27 | 503.20 | 65.44 | 10.09 | 24,573.34 | 25,207.09 |
| 150 | 17.14 | 8.09 | 22.49 | 348.97 | 60.31 | 8.51 | 18,589.87 | 19,055.38 |
| Biostimulating Rate (L/ha) | ||||||||
| 0 | 15.98 a | 8.02 b | 24.68 b | 356.27 b | 56.79 b | 10.36 a | 18,720.40 b | 19,192.51 b |
| 3 | 14.72 b | 8.82 a | 27.78 a | 430.34 a | 67.66 a | 8.28 b | 24,112.85 a | 24,670.46 a |
| Head Parts | ** | ** | ** | ** | ** | ** | ** | ** |
| External Bracts | 11.08 d | 7.70 c | 13.71 d | 174.93 d | 33.30 d | 1.07 d | 8504.33 d | 8746.12 b |
| Intermediate Bracts | 17.76 b | 8.95 b | 31.30 b | 437.54 b | 78.79 b | 7.77 c | 21,017.42 c | 21,581.54 c |
| Inner Bracts | 14.04 c | 7.70 c | 32.90 a | 533.54 a | 56.21 c | 15.03 a | 27,092.25 b | 27,753.79 b |
| Receptacle | 18.52 a | 7.22 d | 27.00 c | 427.22 c | 98.61 a | 13.40 b | 29,052.51 a | 29,644.49 a |
| Luteolin Rutinoside | Luteolin Glucoside | Luteolin Glucuron | Luteolin | Total | |
|---|---|---|---|---|---|
| Year | ** | ** | ** | ** | ** |
| 2016 | 22,009.10 b | 7761.27 b | 8.94 a | 371.82 a | 30,151.10 b |
| 2017 | 24,667.50 a | 14,279.03 a | 8.50 b | 223.30 b | 39,178.40 a |
| Cultivar | ** | * | |||
| Opal | 28,728.70 a | 9139.41 | 10.17 | 195.18 b | 38,073.40 |
| Madrigal | 17,947.90 b | 12,900.88 | 7.28 | 399.96 a | 31,256.00 |
| Nitrogen Rate (kg/ha) | |||||
| 0 | 27,686.20 | 13,480.50 | 12.66 | 384.49 | 41,563.90 |
| 50 | 21,349.30 | 6383.00 | 6.70 | 327.38 | 28,066.30 |
| 100 | 27,377.80 | 18,994.10 | 8.70 | 196.08 | 46,576.70 |
| 150 | 16,940.00 | 5222.90 | 6.84 | 282.32 | 22,452.10 |
| Biostimulating Rate (L/ha) | * | ||||
| 0 | 21,001.90 b | 13,786.66 a | 8.16 b | 289.21 b | 35,085.90 |
| 3 | 25,674.70 a | 8253.64 b | 9.29 a | 305.92 a | 34,243.60 |
| Head Parts | ** | ** | ** | ||
| External Bracts | 5529.50 d | 2514.60 c | 6.12 | 215.37 | 8265.60 d |
| Intermediate Bracts | 22,932.80 c | 2200.90 d | 7.52 | 367.53 | 25,508.80 c |
| Inner Bracts | 34,658.60 a | 9968.40 b | 8.07 | 348.33 | 44,983.40 b |
| Receptacle | 30,232.30 b | 29,396.70 a | 13.18 | 259.03 | 59,901.20 a |
| Apigenin Rutinoside | Apigenin 7-O Glucos | Apigenin 7-O Glucur | Total | |
|---|---|---|---|---|
| Year | ** | ** | ** | ** |
| 2016 | 17.11 a | 29.92 a | 42.74 a | 89.78 a |
| 2017 | 11.53 b | 24.58 b | 35.12 b | 71.23 b |
| Cultivar | ||||
| Opal | 17.72 | 22.69 | 32.41 | 72.82 |
| Madrigal | 10.92 | 31.82 | 45.45 | 88.19 |
| Nitrogen Rate (kg/ha) | ||||
| 0 | 24.01 | 41.55 | 59.36 | 124.91 |
| 50 | 10.98 | 20.88 | 29.83 | 61.69 |
| 100 | 13.99 | 22.39 | 31.98 | 68.36 |
| 150 | 8.29 | 24.20 | 34.57 | 67.07 |
| Biostimulating Rate (L/ha) | ||||
| 0 | 12.65 | 24.83 | 35.47 | 72.96 |
| 3 | 15.99 | 29.67 | 42.39 | 88.06 |
| Head Parts | ** | * | * | * |
| External Bracts | 7.26 d | 58.25 a | 83.21 a | 148.72 a |
| Intermediate Bracts | 9.05 c | 13.19 d | 18.84 d | 41.07 d |
| Inner Bracts | 15.84 b | 16.15 c | 23.07 c | 55.06 c |
| Receptacle | 25.13 a | 21.43 b | 30.62 b | 77.18 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montesano, V.; Negro, D.; Sonnante, G.; Laghetti, G.; Urbano, M. Polyphenolic Compound Variation in Globe Artichoke Cultivars as Affected by Fertilization and Biostimulants Application. Plants 2022, 11, 2067. https://doi.org/10.3390/plants11152067
Montesano V, Negro D, Sonnante G, Laghetti G, Urbano M. Polyphenolic Compound Variation in Globe Artichoke Cultivars as Affected by Fertilization and Biostimulants Application. Plants. 2022; 11(15):2067. https://doi.org/10.3390/plants11152067
Chicago/Turabian StyleMontesano, Vincenzo, Donatella Negro, Gabriella Sonnante, Gaetano Laghetti, and Marcella Urbano. 2022. "Polyphenolic Compound Variation in Globe Artichoke Cultivars as Affected by Fertilization and Biostimulants Application" Plants 11, no. 15: 2067. https://doi.org/10.3390/plants11152067
APA StyleMontesano, V., Negro, D., Sonnante, G., Laghetti, G., & Urbano, M. (2022). Polyphenolic Compound Variation in Globe Artichoke Cultivars as Affected by Fertilization and Biostimulants Application. Plants, 11(15), 2067. https://doi.org/10.3390/plants11152067