
The Anti-Candida Activity of Tephrosia apollinea Is More Superiorly Attributed to a Novel Steroidal Compound with Selective Targeting

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Discovery of Novel Phytochemicals from T. apollinea
2.2. TNS Compound Showed Promising Anti-Candida Activity
2.2.1. In Silico Target Analysis of TNS Compound
2.2.2. Anti-Candida Activity of TNS Compound
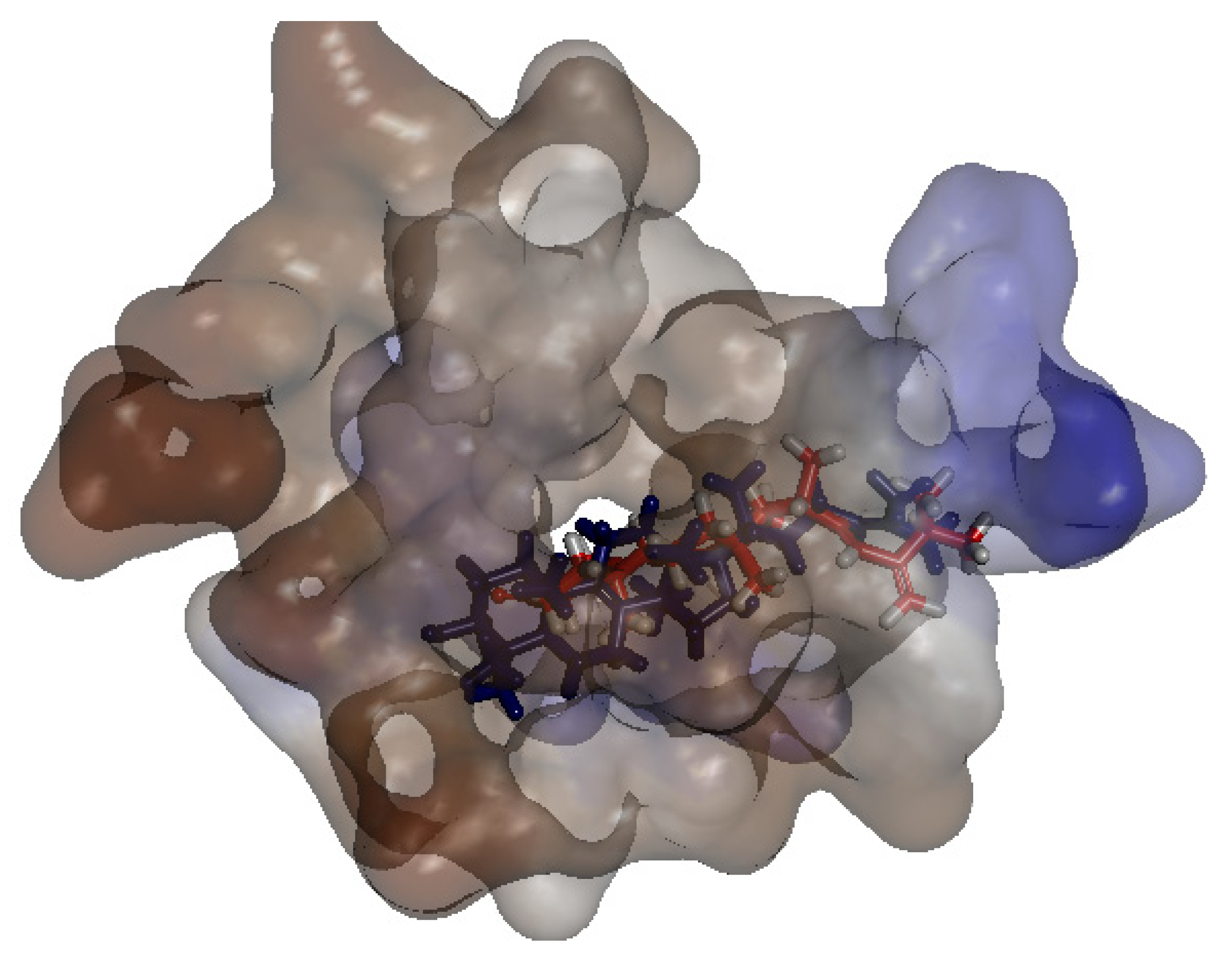
2.2.3. Binding Simulation of TNS with C. Auris 14-α-Demethylase Validate the Anti-Candida Activity of TNS
3. Discussion
4. Conclusions
5. Material and Methods
5.1. Preparation of Plant Material
5.2. Extraction and Compound Isolation
5.3. Purification and Identification of T. appolinea Compounds
5.4. In Silico Target Identification of the Newly Discovered Compound
5.4.1. Generation of Common Feature Pharmacophore
5.4.2. Mapping of the TNS Compound-Pharmacophore Feature
5.5. Homology Modeling and Molecular Docking Study of the TNS Compound against Candida 14-α-Demethylase Enzyme
5.6. Anti-Candida Activity of the TNS Compound
5.7. Cell Toxicity Assay Using MTT Staining
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Al-Ghamdi, F. Morphological Diversity of Some Tephrosia Species (Fabaceae) in Saudi Arabia. Am. J. Plant Sci. 2013, 4, 543–548. [Google Scholar] [CrossRef]
- Cheruth, A.J.; Al Baloushi, S.A.; Karthishwaran, K.; Maqsood, S.; Kurup, S.S.; Sakkir, S. Medicinally active principles analysis of Tephrosia apollinea (Delile) DC. growing in the United Arab Emirates. BMC Res. Notes 2017, 10, 61. [Google Scholar] [CrossRef]
- Ali, M.A.; Mosa, K.A.; El-Keblawy, A.; Alawadhi, H. Exogenous Production of Silver Nanoparticles by Tephrosia apollinea Living Plants under Drought Stress and Their Antimicrobial Activities. Nanomaterials 2019, 9, 1716. [Google Scholar] [CrossRef] [PubMed]
- Lakshmi, P.; Khan, P.A.; Reddy, P.N.; Lakshminarayana, K.; Ganapaty, S. Genetic relationship among Tephrosia species as revealed by RAPD analysis. Asian J. Biol. Sci. 2008, 1, 1–10. [Google Scholar] [CrossRef]
- Chen, Y.; Yan, T.; Gao, C.; Cao, W.; Huang, R. Natural Products from the Genus Tephrosia. Molecules 2014, 19, 1432–1458. [Google Scholar] [CrossRef]
- Rao, A.; Yadav, S.; Singh, P.; Nandal, A.; Singh, N.; Ganaie, S.; Yadav, N.; Kumar, R.; Bhandoria, M.; Bansal, P. A comprehensive review on ethnomedicine, phytochemistry, pharmacology, and toxicity of Tephrosia purpurea (L.) Pers. Phytother. Res. 2020, 34, 1902–1925. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, C.; Prameela Rani, A. In vivo and in vitro evaluation of tephrosia calophylla for anti-diabetic properties. Int. J. Pharm. Pharm. 2018, 10, 138–144. [Google Scholar]
- Divya, S.; Haritha, V.; Prasad, K. Evaluation of Tephrosia calophylla for antiulcer activity in experimental rats. Pharmacologyonline 2011, 3, 573–585. [Google Scholar]
- Lodhi, S.; Pawar, R.S.; Jain, A.P.; Singhai, A.K. Wound healing potential of Tephrosia purpurea (Linn.) Pers. in rats. J. Ethnopharmacol. 2006, 108, 204–210. [Google Scholar] [CrossRef]
- Praveena, R.; Amarnath, S.; Jegadeesan, M. Anti-Inflammatory activity of Tephrosia purpurea. Root. Int. J. Pharmacogn. Phytochem. Res. 2011, 3, 93–94. [Google Scholar]
- Gulecha, V.; Sivakuma, T. Anticancer activity of Tephrosia purpurea and Ficus religiosa using MCF 7 cell lines. Asian Pac. J. Trop. Med. 2011, 4, 526–529. [Google Scholar] [CrossRef]
- Arriaga, A.M.C.; Malcher, G.T.; Lima, J.Q.; Magalhães, F.E.A.; Gomes, T.M.B.M.; Da Conceição, M.; Oliveira, F.; Andrade-Neto, M.; Mafezolli, J.; Santiago, G.M.P. Composition and Larvicidal Activity of the Essential Oil from Tephrosia cinerea Pers. J. Essent. Oil Res. 2008, 20, 450–451. [Google Scholar] [CrossRef]
- Devi, B.P.; Swarnalatha, D.; Gopinath, C.; Adinarayana, K. Anthelmintic activity of Tephrosia calophylla. J. Pharm. Res. 2017, 11, 35–38. [Google Scholar]
- Lwande, W.; Hassanali, A.; Njoroge, P.W.; Bentley, M.D.; Monache, F.D.; Jondiko, J.I. A new 6a-hydroxypterocarpan with insect antifeedant and antifungal properties from the roots of Tephrosia hildebrandtii vatke. Int. J. Trop. Insect Sci. 1985, 6, 537–541. [Google Scholar] [CrossRef]
- Lakshmi, P.; Bobbarala, V.; Ganapaty, S. Chemical and biological evaluation of the leaves of Tephrosia tinctoria (L.) Pers. Int. J. Chem. Anal. Sci. 2010, 1, 14–17. [Google Scholar]
- Upadhyay, B.; Dhaker, A.K.; Kumar, A. Ethnomedicinal and ethnopharmaco-statistical studies of Eastern Rajasthan, India. J. Ethnopharmacol. 2010, 129, 64–86. [Google Scholar] [CrossRef]
- Ashokkumar, D.; Narayana, T.V.; Vidyasagar; Mazumder, U.K.; Gupta, M. Exploration of diuretic potential and electrolyte excretion of Tephrosia purpurea (Fabaceae) in rats. J. Diet. Suppl. 2012, 9, 9–18. [Google Scholar] [CrossRef]
- Akanksha, B.; Avijit, M.; Chakraborthy, G.; Seema, G. Phytopharmacological uses of tephrosia purpurea-a review. Pharmacophore 2014, 5, 658–665. [Google Scholar]
- Samuel, V.J.; Mahesh, A.R.; Murugan, V. Phytochemical and pharmacological aspects of Tephrosia genus: A brief review. J. Appl. Pharm. 2019, 9, 117–125. [Google Scholar]
- Ammar, M.I.; Nenaah, G.E.; Mohamed, A.H.H. Antifungal activity of prenylated flavonoids isolated from Tephrosia apollinea L. against four phytopathogenic fungi. Crop Prot. 2013, 49, 21–25. [Google Scholar] [CrossRef]
- Abou-Douh, A.M.; Ito, C.; Toscano, R.A.; El-Baga, N.Y.; El-Khrisy, E.E.-D.A.; Furukawa, H. Prenylated flavonoids from the root of Egyptian Tephrosia apollinea–crystal structure analysis. Z. Naturforschung B 2005, 60, 458–470. [Google Scholar] [CrossRef]
- Soliman, S.; Hamoda, A.M.; El-Shorbagi, A.-N.A.; El-Keblawy, A.A. Novel betulin derivative is responsible for the anticancer folk use of Ziziphus spina-christi from the hot environmental habitat of UAE. J. Ethnopharmacol. 2019, 231, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Mlozi, S.H.; Mmongoyo, J.A.; Chacha, M. Antimicrobial activities of Tephrosia vogelii against selected pathogenic fungi and bacteria strains. Mycology 2019, 11, 49–55. [Google Scholar] [CrossRef]
- Vyas, V.K.; Ukawala, R.D.; Ghate, M.; Chintha, C. Homology modeling a fast tool for drug discovery: Current perspectives. Indian J. Pharm. Sci. 2012, 74, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Monk, B.C.; Tomasiak, T.M.; Keniya, M.V.; Huschmann, F.U.; Tyndall, J.D.; O’Connell, J.D., 3rd; Cannon, R.D.; McDonald, J.G.; Rodriguez, A.; Finer-Moore, J.S.; et al. Architecture of a single membrane spanning cytochrome P450 suggests constraints that orient the catalytic domain relative to a bilayer. Proc. Natl. Acad. Sci. USA 2014, 111, 3865–3870. [Google Scholar] [CrossRef]
- Xiang, Z. Advances in homology protein structure modeling. Curr. Protein Pept. Sci. 2006, 7, 217–227. [Google Scholar] [CrossRef]
- Webb, B.; Sali, A. Comparative Protein Structure Modeling Using MODELLER. Curr. Protoc. Bioinform. 2016, 54, 5.6.1–5.6.37. [Google Scholar] [CrossRef]
- Gomez-Garibay, F.; Téllez-Valdez, O.; Moreno-Torres, G.; Calderón, J.S. Flavonoids from Tephrosia major. A new prenyl-β-hydroxychalcone. Z. Naturforschung C 2002, 57, 579–583. [Google Scholar] [CrossRef]
- Chen, M.; Wang, K.L.; Liu, M.; She, Z.G.; Wang, C.Y. Bioactive steroid derivatives and butyrolactone derivatives from a Gorgonian-derived Aspergillus sp. fungus. Chem. Biodivers. 2015, 12, 1398–1406. [Google Scholar] [CrossRef]
- Soliman, S.S.M.; Abouleish, M.; Abou-Hashem, M.M.M.; Hamoda, A.M.; El-Keblawy, A.A. Lipophilic Metabolites and Anatomical Acclimatization of Cleome amblyocarpa in the Drought and Extra-Water Areas of the Arid Desert of UAE. Plants 2019, 8, 132. [Google Scholar] [CrossRef]
- Sánchez, J.A. Diversity and Evolution of Octocoral Animal Forests at Both Sides of Tropical America. In Marine Animal Forests; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Soliman, S.; Alsaadi, A.; Youssef, E.; Khitrov, G.; Noreddin, A.; Husseiny, M.; Ibrahim, A. Calli Essential Oils Synergize with Lawsone against Multidrug Resistant Pathogens. Molecules 2017, 22, 2223. [Google Scholar] [CrossRef]
- Halo, B.A.; Al-Yahyai, R.A.; Al-Sadi, A.M. Aspergillus terreus Inhibits Growth and Induces Morphological Abnormalities in Pythium aphanidermatum and Suppresses Pythium-Induced Damping-Off of Cucumber. Front. Microbiol. 2018, 9, 95. [Google Scholar] [CrossRef] [PubMed]
- Zhabinskii, V.N.; Drasar, P.; Khripach, V.A. Structure and Biological Activity of Ergostane-Type Steroids from Fungi. Molecules 2022, 27, 2103. [Google Scholar] [CrossRef]
- Duecker, F.L.; Reuß, F.; Heretsch, P. Rearranged ergostane-type natural products: Chemistry, biology, and medicinal aspects. Org. Biomol. Chem. 2019, 17, 1624–1633. [Google Scholar] [CrossRef]
- Lee, S.R.; Choi, J.H.; Ryoo, R.; Kim, J.-C.; Pang, C.; Kim, S.-H.; Kim, K.H. Ergostane-Type Steroids from Korean Wild Mushroom Xerula furfuracea that Control Adipocyte and Osteoblast Differentiation. J. Microbiol. Biotechnol. 2020, 30, 1769–1776. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhang, M.; Yu, M.; Wang, J.; Zhu, H.; Chen, C.; Zhang, Y. Four new ergostane-type steroids from Lasiodiplodia pseudotheobromae. Tetrahedron Lett. 2020, 61, 151737. [Google Scholar] [CrossRef]
- Yu, J.-H.; Yu, S.-J.; Liu, K.-L.; Wang, C.; Liu, C.; Sun, J.-y.; Zhang, H. Cytotoxic ergostane-type steroids from Ganoderma lingzhi. Steroids 2021, 165, 108767. [Google Scholar] [CrossRef]
- Lima, L.S.; Lima, M.V.; David, J.P.; Giulietti, A.M.; Queiroz, L.P.d.; David, J.M. Megastimanes and ergostane type steroid from leaves Cratylia mollis (Leguminosae). J. Braz. Chem. Soc. 2009, 20, 1921–1924. [Google Scholar] [CrossRef]
- Happi, G.M.; Wouamba, S.C.N.; Ismail, M.; Kouam, S.F.; Frese, M.; Lenta, B.N.; Sewald, N. Ergostane-type steroids from the Cameroonian ′white tiama′ Entandrophragma angolense. Steroids 2020, 156, 108584. [Google Scholar] [CrossRef]
- Frolow, F.; Ray, A.B.; Sahai, M.; Glotter, E.; Gottlieb, H.E.; Kirson, I. Withaperuvin and 4-deoxyphysalolactone, two new ergostane-type steroids from Physalis peruviana(Solanaceae). J. Chem. Soc. 1981, 1029–1032. [Google Scholar] [CrossRef]
- Sagatova, A.A.; Keniya, M.V.; Wilson, R.K.; Monk, B.C.; Tyndall, J.D. Structural Insights into Binding of the Antifungal Drug Fluconazole to Saccharomyces cerevisiae Lanosterol 14α-Demethylase. Antimicrob. Agents Chemother. 2015, 59, 4982–4989. [Google Scholar] [CrossRef] [PubMed]
- Shafiei, M.; Peyton, L.; Hashemzadeh, M.; Foroumadi, A. History of the development of antifungal azoles: A review on structures, SAR, and mechanism of action. Bioorganic Chem. 2020, 104, 104240. [Google Scholar] [CrossRef]
- Lepesheva, G.I.; Hargrove, T.Y.; Kleshchenko, Y.; Nes, W.D.; Villalta, F.; Waterman, M.R. CYP51: A major drug target in the cytochrome P450 superfamily. Lipids 2008, 43, 1117–1125. [Google Scholar] [CrossRef]
- Watson, P.F.; Rose, M.E.; Ellis, S.W.; England, H.; Kelly, S.L. Defective sterol C5-6 desaturation and azole resistance: A new hypothesis for the mode of action of azole antifungals. Biochem. Biophys. Res. Commun. 1989, 164, 1170–1175. [Google Scholar] [CrossRef]
- Keniya, M.V.; Sabherwal, M.; Wilson, R.K.; Woods, M.A.; Sagatova, A.A.; Tyndall, J.D.A.; Monk, B.C. Crystal Structures of Full-Length Lanosterol 14α-Demethylases of Prominent Fungal Pathogens Candida albicans and Candida glabrata Provide Tools for Antifungal Discovery. Antimicrob. Agents Chemother. 2018, 62, e01134-18. [Google Scholar] [CrossRef] [PubMed]
- Alcazar-Fuoli, L.; Mellado, E. Ergosterol biosynthesis in Aspergillus fumigatus: Its relevance as an antifungal target and role in antifungal drug resistance. Front. Microbiol. 2013, 3, 439. [Google Scholar] [CrossRef]
- Wu, G.; Robertson, D.H.; Brooks, C.L., 3rd; Vieth, M. Detailed analysis of grid-based molecular docking: A case study of CDOCKER-A CHARMm-based MD docking algorithm. J. Comput. Chem. 2003, 24, 1549–1562. [Google Scholar] [CrossRef]
- Khatibi, S.; Taban, Z.F.; Roushandeh, A.M. In vitro evaluation of cytotoxic and antiproliferative effects of Portulaca oleracea ethanolic extracton on hela cell line. Gene Cell Tissue 2016, 4, e13301. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δ (H) | δ (C) | Position | δ (H) | δ (C) |
|---|---|---|---|---|---|
| 1 | 2.04–1.99 (m),1.64–1.62 (m) | 34.7 | 16 | 1.69–1.67 (m), 1.24–1.20 (m) | 26.9 |
| 2 | 2.40–2.36 (m) | 34.6 | 17 | 1.59–1.53 | 32.5 |
| 3 | -------- | 207.2 | 18 | 0.86 (s) | 19.4 |
| 4 | 2.21–2.10 (m) | 40.8 | 19 | 1.11 (s) | 14.2 |
| 5 | 2.38–2.04 (m) | 45.5 | 20 | 1.99–1.97 (m) | 39.8 |
| 6 | 5.25 (dd, J = 15.8, 9.9) | 129.8 | 21 | 0.89 (d, J = 6.8) | 20.8 |
| 7 | 5.13 (d, J = 9.9) | 133.6 | 22 | 5.11 (dd, J = 15.7, 8) | 124.4 |
| 8 | -------- | 149.1 | 23 | 5.78 (d, J = 15.7) | 135.6 |
| 9 | -------- | 45.5 | 24 | -------- | 135.2 |
| 10 | -------- | 37.23 | 25 | 1.99–2.03 (m) | 53.0 |
| 11 | 1.69–1.67 (m), 1.58–1.55 (m) | 22.8 | 26 | 0.81 (d, J = 6.8) | 19.5 |
| 12 | 1.61–1.59 (m), 1.44–1.40 (m) | 32.1 | 27 | 0.81 (d, J = 6.8) | 19.4 |
| 13 | -------- | 39.8 | 28 | 4.79, 4.74 (d, J = 2.3) | 109.1 |
| 14 | -------- | 148.9 | |||
| 15 | 2.13–2.10 (m), 2.08–2.05 (m) | 26.6 |
| Ligand | Binding Energy | Total Binding Energy | Ligand Energy | Protein Energy | Complex Energy | Entropic Energy | Ligand Conformational Energy | Ligand Conformational Entropy |
|---|---|---|---|---|---|---|---|---|
| TNS | −49.22 | −45.13 | 87.32 | 41,302.00 | 41,339.74 | 20.20 | 4.09 | 0.62 |
| Lanosterol | −46.65 | −45.82 | 132.76 | 41,302.00 | 41,387.74 | 20.40 | 0.83 | 0.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashmawy, N.S.; El-labbad, E.M.; Hamoda, A.M.; El-Keblawy, A.A.; El-Shorbagi, A.-N.A.; Mosa, K.A.; Soliman, S.S.M. The Anti-Candida Activity of Tephrosia apollinea Is More Superiorly Attributed to a Novel Steroidal Compound with Selective Targeting. Plants 2022, 11, 2120. https://doi.org/10.3390/plants11162120
Ashmawy NS, El-labbad EM, Hamoda AM, El-Keblawy AA, El-Shorbagi A-NA, Mosa KA, Soliman SSM. The Anti-Candida Activity of Tephrosia apollinea Is More Superiorly Attributed to a Novel Steroidal Compound with Selective Targeting. Plants. 2022; 11(16):2120. https://doi.org/10.3390/plants11162120
Chicago/Turabian StyleAshmawy, Naglaa S., Eman M. El-labbad, Alshaimaa M. Hamoda, Ali A. El-Keblawy, Abdel-Nasser A. El-Shorbagi, Kareem A. Mosa, and Sameh S. M. Soliman. 2022. "The Anti-Candida Activity of Tephrosia apollinea Is More Superiorly Attributed to a Novel Steroidal Compound with Selective Targeting" Plants 11, no. 16: 2120. https://doi.org/10.3390/plants11162120
APA StyleAshmawy, N. S., El-labbad, E. M., Hamoda, A. M., El-Keblawy, A. A., El-Shorbagi, A. -N. A., Mosa, K. A., & Soliman, S. S. M. (2022). The Anti-Candida Activity of Tephrosia apollinea Is More Superiorly Attributed to a Novel Steroidal Compound with Selective Targeting. Plants, 11(16), 2120. https://doi.org/10.3390/plants11162120