
Preliminary Studies on the In Vitro Interactions Between the Secondary Metabolites Produced by Esca-Associated Fungi and Enological Saccharomyces cerevisiae Strains
 , and
, and
Abstract
:1. Introduction
2. Results
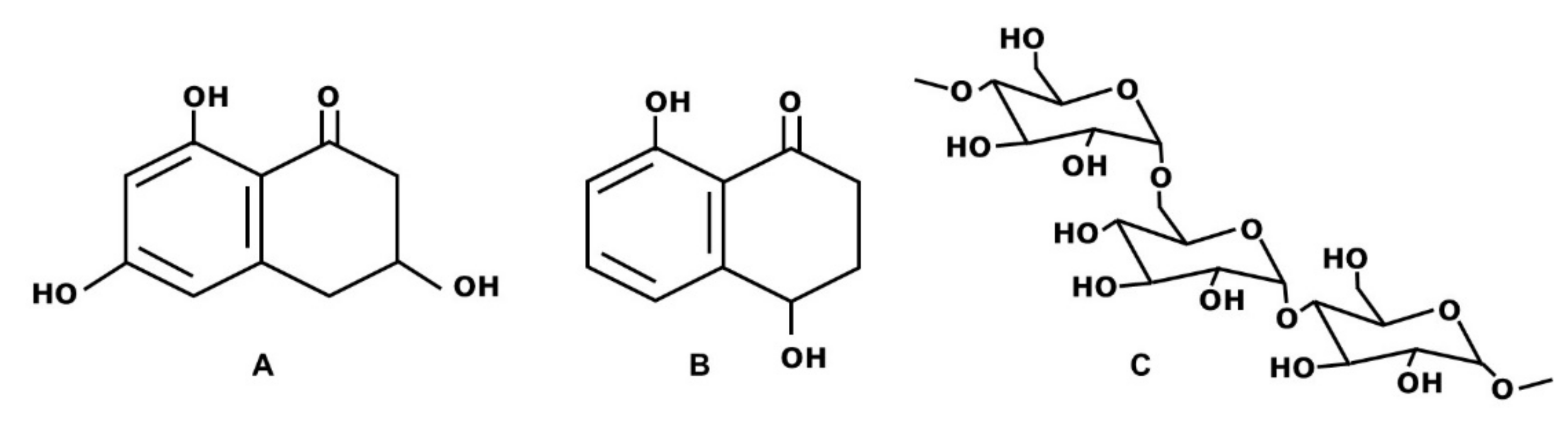
2.1. Pullulan and Pentaketides Production, Extraction and Purification
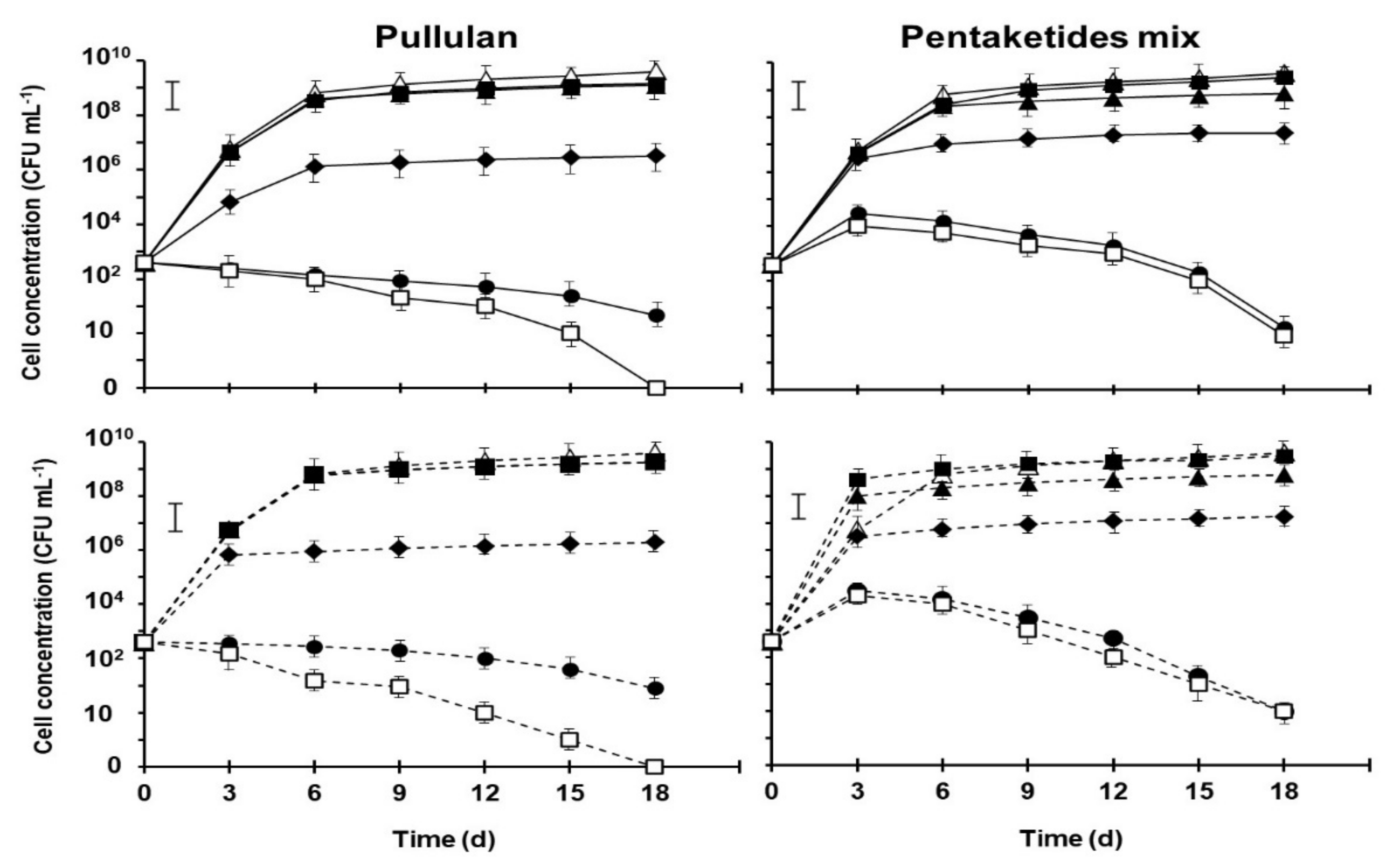
2.2. Pullulan and Pentaketides Mix Interaction with Saccharomyces Cerevisiae Strains
3. Discussion
4. Materials and Methods
4.1. Strains
4.2. Production, Extraction and Purification of Secondary Metabolites
4.3. Pullulan and Pentaketides Mix Interaction with S. Cerevisiae Strains
4.3.1. Effects on Growth
4.3.2. Effects on Enological Parameters
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kenfaoui, J.; Radouane, N.; Mennani, M.; Tahiri, A.; El Ghadraoui, L.; Belabess, Z.; Fontaine, F.; El Hamss, H.; Amiri, S.; Lahlali, R.; et al. A panoramic view on Grapevine Trunk Diseases threats: Case of Eutypa dieback, Botryosphaeria dieback, and Esca disease. J. Fungi 2022, 8, 595. [Google Scholar] [CrossRef] [PubMed]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clément, C.; Mugnai, L.; Fontaine, F. Grapevine Trunk Diseases: A review of fifteen years of trials for their control with chemicals and biocontrol agents. Plant Dis. 2018, 102, 1189–1217. [Google Scholar] [CrossRef] [PubMed]
- Hofstetter, V.; Buyck, B.; Croll, D.; Viret, O.; Couloux, A.; Gindro, K. What if esca disease of grapevine were not a fungal disease? Fungal Divers. 2012, 54, 51–67. [Google Scholar] [CrossRef]
- Bruno, G.L.; Ippolito, M.P.; Mannerucci, F.; Bragazzi, L.; Tommasi, F. Physiological responses of ‘Italia’ grapevines infected with Esca pathogens. Phytopathol. Mediterr. 2021, 60, 321–336. [Google Scholar] [CrossRef]
- Perez-Gonzalez, G.; Sebestyen, D.; Petit, E.; Jellison, J.; Mugnai, L.; Gelhaye, E.; Lee, N.; Farine, S.; Bertsch, C.; Goodell, B. Oxygen Radical-Generating Metabolites Secreted by Eutypa and Esca Fungal Consortia: Understanding the Mechanisms Behind Grapevine Wood Deterioration and Pathogenesis. Front. Plant Sci. 2022, 13, 921961. [Google Scholar] [CrossRef] [PubMed]
- Graniti, A.; Surico, G.; Mugnai, L. Esca of grapevine: A disease complex or a complex of diseases? Phytopathol. Mediterr. 2000, 39, 16–20. [Google Scholar]
- Surico, G. Towards a redefinition of the diseases within the esca complex of grapevine. Phytopathol. Mediterr. 2009, 48, 5–10. [Google Scholar] [CrossRef]
- Bertsch, C.; Ramírez-Suero, M.; Magnin-Robert, M.; Larignon, P.; Chong, J.; Abou-Mansour, E.; Spagnolo, A.; Clément, C.; Fontaine, F. Grapevine trunk diseases: Complex and still poorly understood. Plant Pathol. 2012, 62, 243–265. [Google Scholar] [CrossRef]
- Mugnai, L.; Graniti, A.; Surico, G. Esca (Black Measles) and brown wood-streaking: Two old and elusive diseases of grapevines. Plant Dis. 1999, 83, 404–418. [Google Scholar] [CrossRef]
- Chervin, J.; Romeo-Oliván, A.; Fournier, S.; Puech-Pages, V.; Dumas, B.; Jacques, A.; Marti, G. Modification of Early Response of Vitis vinifera to Pathogens Relating to Esca Disease and Biocontrol Agent Vintec® Revealed by Untargeted Metabolomics on Woody Tissues. Front. Microbiol. 2022, 13, 835463. [Google Scholar] [CrossRef]
- Fischer, M. A new wood-decaying basidomycete species associated with esca of grapevine: Fomitiporia mediterranea (Hymenochaetales). Mycol. Prog. 2002, 1, 315–324. [Google Scholar] [CrossRef]
- Fischer, M. Biodiversity and geographic distribution of basidiomycetes causing esca-associated white rot in grapevine: A worldwide perspective. Phytopathol. Mediterr. 2006, 45, 30–42. [Google Scholar] [CrossRef]
- Spasova, M.; Manolova, N.; Rashkov, I.; Naydenov, M. Eco-Friendly Hybrid PLLA/Chitosan/Trichoderma asperellum Nanomaterials as Biocontrol Dressings against Esca Disease in Grapevines. Polymers 2022, 14, 2356. [Google Scholar] [CrossRef]
- Bruno, G.; Sparapano, L.; Graniti, A. Effects of three esca-associated fungi on Vitis vinifera L.: IV. Diffusion through the xylem of metabolites produced by two tracheiphilous fungi in the woody tissue of grapevine leads to esca-like symptoms on leaves and berries. Physiol. Mol. Plant Pathol. 2007, 71, 106–124. [Google Scholar] [CrossRef]
- Andolfi, A.; Mugnai, L.; Luque, J.; Surico, G.; Cimmino, A.; Evidente, A. Phytotoxins produced by fungi associated with grapevine trunk diseases. Toxins 2011, 3, 1569–1605. [Google Scholar] [CrossRef]
- Lecomte, P.; Darrieutort, G.; Liminana, J.M.; Comont, G.; Muruamendiaraz, A.; Legorburu, F.-J.; Choueiri, E.; Jreijiri, F.; El Amil, R.; Fermaud, M. New insights into esca of grapevine: The development of foliar symptoms and their association with xylem discoloration. Plant Dis. 2012, 96, 924–934. [Google Scholar] [CrossRef]
- Amalfitano, C.; Evidente, A.; Surico, G.; Tegli, S.; Bertelli, E.; Mugnai, L. Phenols and stilbene polyphenols in the wood of esca-diseased grapevines. Phytopathol. Mediter. 2000, 39, 178–183. [Google Scholar]
- Amalfitano, C.; Agrelli, D.; Arrigo, A.; Mugnai, L.; Surico, G.; Evidente, A. Stilbene Polyphenols in the brown red wood of Vitis vinifera cv. sangiovese affected by “Esca Proper”. Phytopathol. Mediter. 2011, 50, S224–S235. [Google Scholar]
- Lima, M.R.M.; Felgueiras, M.L.; Cunha, A.; Chicau, G.; Ferreres, F.; Dias, A.C.P. Differential phenolic production in leaves of Vitis vinifera cv. alvarinho affected with esca disease. Plant Physiol. Biochem. 2017, 112, 45–52. [Google Scholar] [CrossRef]
- Lima, M.R.M.; Machado, A.F.; Gubler, W.D. Metabolomic study of chardonnay grapevines double stressed with esca-associated fungi and drought. Phytopathology 2017, 107, 669–680. [Google Scholar] [CrossRef]
- Lima, M.R.M.; Felgueiras, M.L.; Graca, G.; Rodrigues, J.E.A.; Barros, A.; Gil, A.M.; Dias, A.C. NMR metabolomics of Esca disease-affected Vitis vinifera cv. Alvarinho leaves. J. Exp. Bot. 2010, 61, 4033–4042. [Google Scholar] [CrossRef] [PubMed]
- Calzarano, F.; Amalfitano, C.; Seghetti, L.; Cozzolino, V. Nutritional status of vines affected with esca proper. Phytopathol. Mediterr. 2009, 48, 20–31. [Google Scholar] [CrossRef]
- Calzarano, F.; Di Marco, S.; D’agostino, V.; Schiff, S.; Mugnai, L. Grapevine leaf stripe disease symptoms (esca complex) are reduced by a nutrients and seaweed mixture. Phytopathol. Mediterr. 2014, 53, 543–558. [Google Scholar] [CrossRef]
- Calzarano, F.; Di Marco, S. Further evidence that calcium, magnesium and seaweed mixtures reduce grapevine leaf stripe symptoms and increase grape yield. Phytopathol. Mediterr. 2018, 57, 459–471. [Google Scholar] [CrossRef]
- Evidente, A.; Sparapano, L.; Andolfi, A.; Bruno, G. Two naphthalenone pentaketides isolated from liquid cultures of Phaeoacremonium aleophilum, a fungus associated with esca disease syndrome. Phytopathol. Mediterr. 2000, 39, 162–168. [Google Scholar] [CrossRef]
- Fkyerat, C.; Dubin, G.-M.; Tabacchi, R. Phytotoxins from fungi of esca of grapevine. Phytopathol. Mediterr. 2000, 39, 156–161. [Google Scholar] [CrossRef]
- Bruno, G.; Sparapano, L. Effects of three esca-associated fungi on Vitis vinifera L.: I. Characterization of secondary metabolites in culture media and host responses to the pathogens in calli. Physiol. Mol. Plant Pathol. 2006, 69, 209–223. [Google Scholar] [CrossRef]
- Bruno, G.; Sparapano, L. Effects of three esca-associated fungi on Vitis vinifera L.: II. Characterization of biomolecules in xylem sap and leaves of healthy and diseased vines. Physiol. Mol. Plant Pathol. 2006, 69, 195–208. [Google Scholar] [CrossRef]
- Luini, E.; Fleurat-Lessard, P.; Rousseau, L.; Roblin, G.; Berjeaud, J. Inhibitory effects of polypeptides secreted by the grapevine pathogens Phaeomoniella chlamydospora and Phaeoacremonium aleophilum on plant cell activities. Physiol. Mol. Plant Pathol. 2010, 74, 403–411. [Google Scholar] [CrossRef]
- Petit, A.N.; Vaillant, N.; Boulay, M.; Clement, C.; Fontaine, F. Alteration of photosynthesis in grapevines affected by esca. Phytopathology 2006, 96, 1060–1066. [Google Scholar] [CrossRef]
- Magnin-Robert, M.; Letousey, P.; Spagnolo, A.; Rabenoelina, F.; Jacquens, L.; Mercier, L.; Clément, C.; Fontaine, F. Leaf strip of esca induces alteration of photosynthesis and defence reactions in presymptomatic leaves. Funct. Plant Biol. 2011, 38, 856–866. [Google Scholar] [CrossRef]
- Valtaud, C.; Thibault, F.; Larignon, P.; Berstch, C.; Fleurat-Lessard, P.; Bourbouloux, A. Systemic damage in leaf metabolism caused by esca infection in grapevines. Aust. J. Grape Wine Res. 2011, 17, 101–110. [Google Scholar] [CrossRef]
- Fontaine, F.; Pinto, C.; Vallet, J.; Clément, C.; Gomes, A.C.; Spagnolo, A. The effects of grapevine trunk diseases (GTDs) on vine physiology. Eur. J. Plant Pathol. 2016, 144, 707–721. [Google Scholar] [CrossRef]
- Calzarano, F.; D’Agostino, V.; Pepe, A.; Osti, F.; Della Pelle, F.; De Rosso, M.; Flamini, R.; Di Marco, S. Patterns of phytoalexins in the grapevine leaf stripe disease (esca complex)/grapevine pathosystem. Phytopathol. Mediterr. 2016, 55, 410–426. [Google Scholar] [CrossRef]
- Calzarano, F.; Osti, F.; D’Agostino, V.; Pepe, A.; Di Marco, S. Mixture of calcium, magnesium and seaweed affects leaf phytoalexin contents and grape ripening on vines with grapevine leaf stripe disease. Phytopathol. Mediterr. 2017, 56, 445–457. [Google Scholar] [CrossRef]
- Calzarano, F.; Osti, F.; D’Agostino, V.; Pepe, A.; Della Pelle, F.; De Rosso, M.; Flamini, R.; Di Marco, S. Levels of phytoalexins in vine leaves with different degrees of grapevine leaf stripe disease symptoms (Esca complex of diseases). Phytopathol. Mediterr. 2017, 56, 494–501. [Google Scholar] [CrossRef]
- Fischer, M.; Peighami Ashnaei, S. Grapevine, esca complex, and environment: The disease triangle. Phytopathol. Mediterr. 2019, 58, 17–37. [Google Scholar] [CrossRef]
- Songy, A.; Fernandez, O.; Clément, C.; Larignon, P.; Fontaine, F. Grapevine trunk diseases under thermal and water stresses. Planta 2019, 249, 1655–1679. [Google Scholar] [CrossRef]
- Calzarano, F.; Seghetti, L.; Del Carlo, M.; Cichelli, A. Effect of esca on the quality of berries, musts and wines. Phytopathol. Mediterr. 2004, 43, 125–135. [Google Scholar] [CrossRef]
- Lorrain, B.; Ky, I.; Pasquier, G.; Jourdes, M.; Guerin Dubrana, L.; Gény, L.; Rey, P.; Donèche, B.; Teissedre, P.-L. Effect of Esca disease on the phenolic and sensory attributes of Cabernet Sauvignon grapes, musts and wines. Aust. J. Grape Wine Res. 2012, 18, 64–72. [Google Scholar] [CrossRef]
- Cheynier, V.; Dueñas-Paton, M.; Salas, E.; Maury, C.; Souquet, J.-M.; Sarni-Manchado, P.; Fulcrand, H. Structure and Properties of Wine Pigments and Tannins. Am. J. Enol. Vitic. 2006, 57, 298–305. [Google Scholar]
- Peleg, H.; Gacon, K.; Schlich, P.; Noble, A.C. Bitterness and astringency of flavan-3-ol monomers, dimers and trimers. J. Sci. Food Agric. 1999, 79, 1123–1128. [Google Scholar] [CrossRef]
- Somers, T.C. The polymeric nature of wine pigments. Phytochemistry 1971, 10, 2175–2186. [Google Scholar] [CrossRef]
- Vivar-Quintana, A.M.; Santos-Buelga, C.; Francia-Aricha, E.; Rivas-Gonzalo, J.C. Formation of anthocyanin-derived pigments in experimental red wines. Food Sci Technol Int. 1999, 5, 347–352. [Google Scholar] [CrossRef]
- Regueiro, L.A.; Costas, C.L.; Rubio, J.E.L. Influence of viticultural and enological practices on the development of yeast populations during winemaking. Am. J. Enol. Vitic. 1993, 44, 405–408. [Google Scholar]
- Martini, A.; Vaughan-Martini, A. Grape Must Fermentation: Past and Present. In Yeast Technology; Spencer, J.F.T., Spencer D., M., Eds.; Springer: Berlin/Heidelberg, Germany, 1990; pp. 105–123. [Google Scholar]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Ribéreau-Gayon, J.; Peynaud, E.; Lafourcade, S. The formation of fermentation inhibitory substances by Botrytis cinerea. C. R. Acad. Sci. D Nat. 1952, 234, 478–480. [Google Scholar]
- Ribéreau-Gayon, P.; Lafon-Lafourcade, S.; Dubourdieu, D.; Lucmaret, V.; Larue, F. Métabolisme de Saccharomyces cerevisiae dans les moûts de raisins parasites par Botrytis cinerea. Inhibition de la fermentation; formation d’acide acetique et de glycerol. C. R. Acad. Sci. Ser. D Nat. 1979, 289, 441–444. [Google Scholar]
- Ribereau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. The Chemistry of Wine. In Handbook of Enology, the Microbiology of Wine and Vinifications; Ribereau-Gayon, P., Ed.; John Wiley & Sons, Ltd: Chichester, UK, 2006; pp. 1–228. [Google Scholar]
- Sparapano, L.; Bruno, G.; Graniti, A. Effects on plants of metabolites produced in culture by Phaeoacremonium chlamydosporum, P. aleophilum and Fomitiporia punctata. Phytopathol. Mediterr. 2000, 39, 169–177. [Google Scholar] [CrossRef]
- Kimoto, T.; Shibuya, T.; Shiobara, S. Safety studies of a novel starch, pullulan: Chronic toxicity in rats and bacterial mutagenicity. Food Chem. Toxicol. 1997, 35, 323. [Google Scholar] [CrossRef]
- Leathers, T.D. Pullulan. In Biopolymers. Polysaccharides II: Polysaccharides from Eukariotes; Vandamme, E.J., De Baets, S., Steinbüchel, A., Eds.; Wiley-VCH: Weinheim, Germany, 2002; Volume 6, pp. 1–25. [Google Scholar]
- Rossouw, D.; Jolly, N.; Jacobson, D.; Bauer, F.F. The effect of scale on gene expression: Commercial versus laboratory wine fermentations. Appl Microbiol Biotechnol. 2012, 4, 1207–1219. [Google Scholar] [CrossRef]
- Viana, T.; Loureiro-Dias, M.C. Prista Efficient fermentation of an improved synthetic grape must by enological and laboratory strains of Saccharomyces cerevisiae. AMB Express 2014, 4, 16. [Google Scholar] [CrossRef]
- Graniti, A. Phytotoxins and their involvement in plant diseases. Experientia 1991, 47, 751–755. [Google Scholar] [CrossRef]
- Pontes, J.G.M.; Fernandes, L.S.; Dos Santos, R.V.; Tasic, L.; Fill, T.P. Virulence Factors in the Phytopathogen–Host Interactions: An Overview. J. Agric. Food Chem. 2020, 68, 7555–7570. [Google Scholar] [CrossRef]
- Mathieu, V.; Superchi, S.; Masi, M.; Scafato, P.; Kornienko, A.; Evidente, A. In Vitro Effects of Fungal Phytotoxins on Cancer Cell Viability: First Insight into Structure Activity Relationship of a Potent Metabolite of Cochliobolus australiensis Radicinin. Toxins 2022, 14, 517. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Gopal, J.V.; Ren, S.; Chen, L.; Liu, L.; Gao, Z. Anticancer fungal natural products: Mechanisms of action and biosynthesis. Eur. J. Med. Chem. 2020, 202, 112502. [Google Scholar] [CrossRef]
- Xu, D.; Xue, M.; Shen, Z.; Jia, X.; Hou, X.; Lai, D.; Zhou, L. Phytotoxic Secondary Metabolites from Fungi. Toxins 2021, 13, 261. [Google Scholar] [CrossRef]
- Belinchón, M.M.; Gancedo, J.M. Glucose controls multiple processes in Saccharomyces cerevisiae through diverse combinations of signaling pathways. FEMS Yeast Res. 2007, 7, 808–818. [Google Scholar] [CrossRef] [PubMed]
- Gancedo, J.M. The early steps of glucose signalling in yeast. FEMS Microbiol. Rev. 2008, 32, 673–704. [Google Scholar] [CrossRef]
- Zamora, F. Biochemistry of Alcoholic Fermentation. In Wine Chemistry and Biochemistry; Moreno-Arribas, M.V., Polo, M.C., Eds.; Springer Science+Business Media: New York, NY, USA, 2009; pp. 3–26. [Google Scholar]
- Del Mónaco, S.; Barda, N.; Rubio, N.; Caballero, A. Selection and characterization of a Patagonian Pichia kudriavzevii for wine deacidification. J. Appl. Microbiol. 2014, 117, 451–464. [Google Scholar] [CrossRef]
- Godden, P.; Wilkes, E.; Johnson, D. Trends in the composition of Australian wine 1984–2014. Aust. J. Grape Wine Res. 2015, 21, 741–753. [Google Scholar] [CrossRef]
- Zoecklin, B.W.; Fugelsang, K.C.; Gump, B.H.; Nury, F.S. Hydrogen Ion (pH) and Fixed Acids. In Wine Analysis and Production; Zoecklin, B.W., Fugelsang, K.C., Gump, B.H., Nury, F.S., Eds.; Chapman & Hall: New York, NY, USA, 1995; pp. 76–88. [Google Scholar]
- Redzepovic, S.; Orlic, S.; Majdak, A.; Kozina, B.; Volschenk, H.; Viljoen-Bloom, M. Differential malic acid degradation by selected strains of Saccharomyces during alcoholic fermentation. Int. J. Food Microbiol. 2003, 83, 49–61. [Google Scholar] [CrossRef]
- Shimazu, Y.; Watanabe, M. Effects of yeast strains and environmental conditions on formation of organic acid in must during fermentation. J. Ferment. Technol. 1981, 59, 27–32. [Google Scholar]
- Delfini, C.; Costa, A. Effects of the grape must lees and insoluble materials on the alcoholic fermentation rate and on the production of acetic acid, pyruvic acid and acetaldehyde. Am. J. Enol. Vitic. 1993, 44, 86–92. [Google Scholar]
- Vasserot, Y.; Mornet, F.; Jeandet, P. Acetic acid removal by Saccharomyces cerevisiae during fermentation in enological conditions. Metabolic consequences. Food Chem. 2010, 119, 1220–1223. [Google Scholar] [CrossRef]
- Vigentini, I.; Barrera Cardenas, S.; Valdetara, F.; Faccincani, M.; Panont, C.A.; Picozzi, C.; Foschino, R. Use of native yeast strains for in-bottle fermentation to face the uniformity in sparkling wine production. Front. Microbiol. 2017, 8, 1225. [Google Scholar] [CrossRef]
- Agarbati, A.; Canonico, L.; Ciani, M.; Comitini, F. Fitness of Selected Indigenous Saccharomyces cerevisiae Strains for White Piceno DOC Wines Production. Fermentation 2018, 4, 37. [Google Scholar] [CrossRef]
- Chrzanowski, G. Saccharomyces Cerevisiae-An Interesting Producer of Bioactive Plant Polyphenolic Metabolites. Int. J. Mol. Sci. 2020, 21, 7343. [Google Scholar] [CrossRef] [PubMed]
- Mazza, G.; Fukumoto, L.; Delaquis, P.; Girard, B.; Ewert, B. Anthocyanins, phenolics, and color of Cabernet Franc, Merlot, and Pinot noir wines from British Columbia. J. Agric. Food Chem. 1999, 47, 4009–4017. [Google Scholar] [CrossRef] [PubMed]
- Caridi, A.; Cufari, A.; Ramondino, D. Isolation and clonal pre-selection of enological Saccharomyces. J. Gen. Appl. Microbiol. 2002, 48, 261–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIV. OIV-OENO Resolution, 370-2012. Guidelines for the Characterization of Wine Yeasts of the Genus Saccharomyces Isolated from Vitivinicultural Environments. Available online: https://www.oiv.int/public/medias/1429/oiv-oeno-370-2012-en.pdf (accessed on 11 January 2021).
- OIV. OIV-OENO Resolution, 377-2009. Alcoholic Strength by Volume. Available online: https://www.oiv.int/public/medias/5158/oiv-ma-as312-01a.pdf (accessed on 11 January 2021).



{kind=link}
{kind=link}
| Parameters | Pm33 | Pc25 |
|---|---|---|
| Biomass fresh weight (mg L−1) | 15.3 ± 0.12 | 37.5 ± 0.37 |
| Biomass dry weight (mg L−1) | 4.2 ± 0.02 | 6.8 ± 0.04 |
| Final pH | 3.9 ± 0.2 | 4.0 ± 0.2 |
| Ethyl acetate extracts (mg L−1) | 183.0 ± 12 | 191.0 ± 10 |
| Scytalone (mg L−1) | 23.0 ± 2 | 13.0 ± 1 |
| Isosclerone (mg L−1) | 1.5 ± 0.3 | 2.5 ± 0.6 |
| Ethanol precipitate (g L−1) | 6.5 ± 0.2 | 7.2 ± 0.5 |
| Pullulan (mg L−1) | 430.83 ± 92 | 982.4 ± 0.3 |
| Parameters | Byosal HS1 | IOC 18-2007 | ||||
|---|---|---|---|---|---|---|
| Con | Pul | P-Mix | Con | Pul | P-Mix | |
| pH | 2.40 ± 0.1 a | 2.44 ± 0.1 a | 2.44 ± 0.1 a | 2.40 ± 0.1 a | 2.86 ± 0.1 b | 2.40 ± 0.1 a |
| Residual sugar (g L −1) | 0.1 ± 0.01 a | 0.4 ± 0.01 a | 14.79 ± 0.04 b | 0.29 ± 0.02 a | 0.12 ± 0.01 a | 35.31 ± 0.05 c |
| Utilized sugar (g L −1) | 229.93 ± 0.01 a | 229.61 ± 0.1 a | 215.21 ± 0.4 b | 229.71 ± 0.2 a | 229.92 ± 0.1 a | 194.69 ± 0.2 c |
| Ethanol (%) | 7.5 ± 0.1 a | 4.1 ± 0.1 c | 2.44 ± 0.1 d | 7.3 ± 0.1 a | 5.7 ± 0.1 b | 6.9 ± 0.1 a |
| Ethanol yield 2 | 0.32 ± 0.02 a | 0.17 ± 0.02 c | 0.11 ± 0.02 c | 0.31 ± 0.02 a | 0.24 ± 0.02 a | 0.35 ± 0.02 a |
| Titratable acidity (g L −1) | 10.27 ± 0.1 a | 6.6 ± 0.1 b | 9.82 ± 0.1 a | 10.05 ± 0.1 a | 7.5 ± 0.1 b | 9.75 ± 0.1 a |
| Acetic acid (volatile acidity) (g L −1) | 0.51 ± 0.01 a | 0.25 ± 0.02 c | 0.53 ± 0.02 a | 0.43 ± 0.02 b | 0.40 ± 0.01 b | 0.45 ± 0.01 b |
| Tartaric acid (g L −1) | 4.14 ± 0.07 a | 3.08 ± 0.02 b | 4.29 ± 0.03 a | 4.27 ± 0.03 a | 3.25 ± 0.01 b | 4.29 ± 0.08 a |
| L-malic acid (g L −1) | 1.46 ± 0.07 a | 0.95 ± 0.02 c | 1.43 ± 0.01 a | 1.45 ± 0.02 a | 1.05 ± 0.02 b | 1.51 ± 0.03 a |
| Lactic acid (g L −1) | 0.05 ± 0.02 a | 0.01 ± 0.001 b | 0.05 ± 0.001 a | 0.06 ± 0.001 a | 0.02 ± 0.001 b | 0.06 ± 0.001 a |
| Total polyphenols (mg L −1) | 464.04 ± 5.2 a | 269.88 ± 3.1 b | 257.91 ± 3.6 d | 261.75 ± 3.2 b | 248.88 ± 3.5 c | 273.63 ± 3.7 b |
| Fermentation vigour 3 | 3.17 ± 0.01 a | 0.35 ± 0.07 c | 0.01 ± 0.001 d | 3.16 ± 0.001 a | 1.35 ± 0.01 b | 0.09 ± 0.001 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scarano, L.; Mazzone, F.; Mannerucci, F.; D’Amico, M.; Bruno, G.L.; Marsico, A.D. Preliminary Studies on the In Vitro Interactions Between the Secondary Metabolites Produced by Esca-Associated Fungi and Enological Saccharomyces cerevisiae Strains. Plants 2022, 11, 2277. https://doi.org/10.3390/plants11172277
Scarano L, Mazzone F, Mannerucci F, D’Amico M, Bruno GL, Marsico AD. Preliminary Studies on the In Vitro Interactions Between the Secondary Metabolites Produced by Esca-Associated Fungi and Enological Saccharomyces cerevisiae Strains. Plants. 2022; 11(17):2277. https://doi.org/10.3390/plants11172277
Chicago/Turabian StyleScarano, Leonardo, Francesco Mazzone, Francesco Mannerucci, Margherita D’Amico, Giovanni Luigi Bruno, and Antonio Domenico Marsico. 2022. "Preliminary Studies on the In Vitro Interactions Between the Secondary Metabolites Produced by Esca-Associated Fungi and Enological Saccharomyces cerevisiae Strains" Plants 11, no. 17: 2277. https://doi.org/10.3390/plants11172277
APA StyleScarano, L., Mazzone, F., Mannerucci, F., D’Amico, M., Bruno, G. L., & Marsico, A. D. (2022). Preliminary Studies on the In Vitro Interactions Between the Secondary Metabolites Produced by Esca-Associated Fungi and Enological Saccharomyces cerevisiae Strains. Plants, 11(17), 2277. https://doi.org/10.3390/plants11172277