
Rhizobacteria Mitigate the Negative Effect of Aluminum on Pea Growth by Immobilizing the Toxicant and Modulating Root Exudation

 , , , , , , ,
, , , , , , ,  ,
,  and
and
Abstract
:1. Introduction
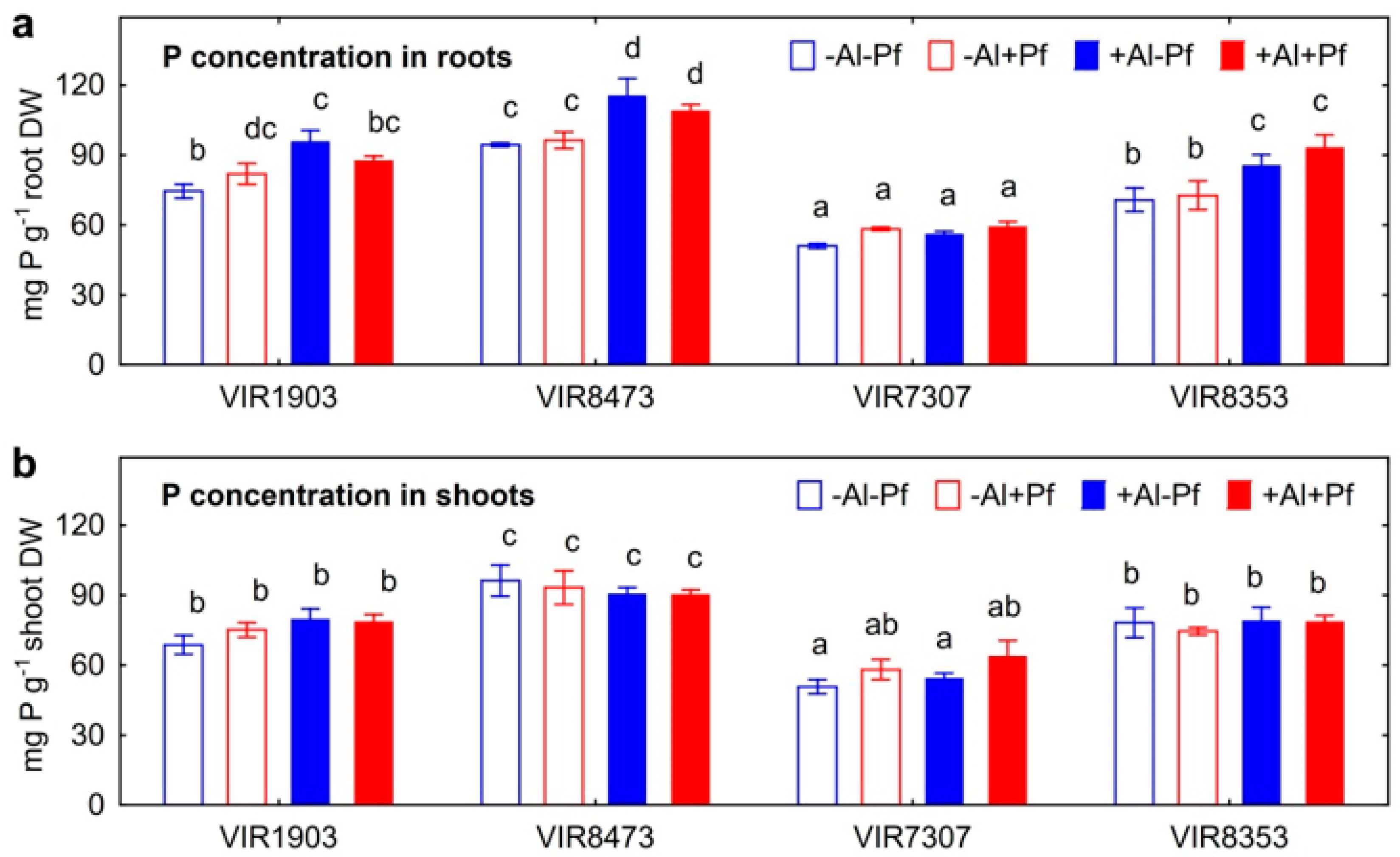
2. Results
2.1. Properties of Ps. fluorescens SPB2137
2.2. Growth of Plants and Bacteria in Hydroponics
2.3. Development of Bacteria in Hydroponics
2.4. Distribution of Al in Hydroponics
2.5. Root Exudation
3. Discussion
3.1. Properties of Ps. fluorescens SPB2137
3.2. Growth of Plants and Bacteria in Hydroponics
3.3. Distribution of Al in Hydroponics
3.4. Root Exudation
3.5. ACC Deaminase
3.6. Significance of Factor’s Effects
4. Materials and Methods
4.1. Plants
4.2. Microorganism
4.3. Bacterial Immobilization of Al in Batch Culture
4.4. Hydroponic Experiments
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Taylor, G.J. Current views of the aluminum stress response: The physiological basis of tolerance. Plant Physiol. Biochem. 1991, 10, 57–93. [Google Scholar]
- Gupta, N.; Gaurav, S.S.; Kumar, A. Molecular basis of aluminium toxicity in plants: A review. Am. J. Plant. Sci. 2013, 4, 21–37. [Google Scholar] [CrossRef]
- Kochian, L.V.; Piñeros, M.A.; Liu, J.; Magalhaes, J.V. Plant adaptation to acid soils: The molecular basis for crop aluminum resistance. Annu. Rev. Plant Biol. 2015, 66, 571–598. [Google Scholar] [CrossRef]
- Ma, J.F.; Ryan, P.R.; Delhaize, E. Aluminium tolerance in plants and the complexing role of organic acids. Trends Plant Sci. 2001, 6, 273–278. [Google Scholar] [CrossRef]
- Kochian, L.V.; Hoekenga, O.A.; Pineros, M.A. How do crop plants tolerate acid soils? Mechanisms of aluminum tolerance and phosphorous efficiency. Annu. Rev. Plant Biol. 2004, 55, 459–493. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Motoda, H. Aluminum toxicity recovery processes in root apexes. Possible association with oxidative stress. Plant Sci. 2012, 185, 1–8. [Google Scholar] [CrossRef]
- Paulino, V.T.; Olivares, J.; Bedmar, E.J. Nodulation and nitrogenase activity of pea nodules as affected by low pH and aluminium. Plant Soil. 1987, 101, 299–302. [Google Scholar] [CrossRef]
- Lazof, D.B.; Holland, M.J. Evaluation of the aluminium-induced root growth inhibition in isolation from low pH effects in Glycine max, Pisum sativum and Phaseolus vulgaris. Funct. Plant Biol. 1999, 26, 147–157. [Google Scholar] [CrossRef]
- Arunakumara, K.K.I.U.; Walpola, B.C.; Yoon, M. Aluminum toxicity and tolerance mechanism in cereals and legumes—A review. J. Korean Soc. Appl. Biol. Chem. 2013, 56, 1–9. [Google Scholar] [CrossRef]
- Motoda, H.; Kano, Y.; Hiragami, F.; Kawamura, K.; Matsumoto, H. Morphological changes in the apex of pea roots during and after recovery from aluminium treatment. Plant Soil. 2010, 333, 49–58. [Google Scholar] [CrossRef]
- Singh, N.B.; Yadav, K.; Amist, N. Phytotoxic effects of aluminum on growth and metabolism of Pisum sativum L. IJBCS 2011, 2, 10–21. [Google Scholar]
- Yu, M.; Shen, R.; Liu, J.; Chen, R.; Xu, M.; Yang, Y.; Xiao, H.; Wang, H.D.; Wang, H.Z.; Wang, C. The role of root border cells in aluminum resistance of pea (Pisum sativum) grown in mist culture. J. Plant Nutr. Soil. Sci. 2009, 172, 528–534. [Google Scholar] [CrossRef]
- Sujkowska-Rybkowska, M.; Borucki, W. Pectins esterification in the apoplast of aluminum-treated pea root nodules. J. Plant Physiol. 2015, 184, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kichigina, N.E.; Pukhalsky, Y.V.; Shaposhnikov, A.I.; Azarova, T.S.; Makarova, N.M.; Loskutov, S.I.; Safronova, V.I.; Tikhonovich, I.A.; Vishniyakova, M.A.; Semenova, E.V.; et al. Aluminum exclusion from root zone and maintenance of nutrient uptake are principal mechanisms of al tolerance in Pisum sativum L. Physiol. Mol. Biol. Plants 2017, 23, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Arato, M.; Borghi, L.; Nouri, E.; Reinhardt, D. Beneficial services of arbuscular mycorrhizal fungi—From ecology to application. Front. Plant Sci. 2018, 9, 1270. [Google Scholar] [CrossRef]
- Bukhat, S.; Imran, A.; Javaid, S.; Shahid, M.; Majeed, A.; Naqqash, T. Communication of plants with microbial world: Exploring the regulatory networks for PGPR mediated defense signaling. Microbiol. Res. 2020, 238, 126486. [Google Scholar] [CrossRef]
- Kumar, M.; Giri, V.P.; Pandey, S.; Gupta, A.; Patel, M.K.; Bajpai, A.B.; Jenkins, S.; Siddique, K.H.M. Plant-Growth-Promoting Rhizobacteria emerging as an effective bioinoculant to improve the growth, production, and stress tolerance of vegetable crops. Int. J. Mol. Sci. 2021, 22, 12245. [Google Scholar] [CrossRef]
- Riaz, M.; Kamran, M.; Fang, Y.; Wang, Q.; Cao, H.; Yang, G.; Deng, L.; Wang, Y.; Zhou, Y.; Anastopoulos, I.; et al. Arbuscular mycorrhizal fungi-induced mitigation of heavy metal phytotoxicity in metal contaminated soils: A critical review. J. Hazard. Mater. 2021, 402, 123919. [Google Scholar] [CrossRef]
- Hawkins, J.P.; Oresnik, I.J. The rhizobium-legume symbiosis: Co-opting successful stress management. Front. Plant Sci. 2021, 12, 796045. [Google Scholar] [CrossRef]
- Koslowsky, S.D.; Boerner, R.E.J. Interactive effects of aluminium, phosphorus and mycorrhizae on growth and nutrient uptake of Panicum virgatum L. (Poaceae). Environ. Pollut. 1989, 61, 107–125. [Google Scholar] [CrossRef]
- Medeiros, C.A.B.; Clark, R.B.; Ellis, J.R. Effects of excess aluminium on mineral uptake in mycorrhizal sorghum. J. Plant Nutr. 1994, 17, 1399–1416. [Google Scholar] [CrossRef]
- Mendoza, J.; Borie, F. Effect of Glomus etunicatum inoculation on aluminium, phosphorus, calcium, and magnesium uptake of two barley genotypes with different aluminium tolerance. Comm. Soil Sci. Plant Anal. 1998, 29, 681–695. [Google Scholar] [CrossRef]
- Lux, H.B.; Cumming, J.R. Mycorrhizae confer aluminium resistance to tulip-poplar seedlings. Can. J. For. Res. 2001, 31, 694–702. [Google Scholar] [CrossRef]
- Seguel, A.; Cumming, J.R.; Klugh-Stewart, K.; Cornejo, P.; Borie, F. The role of arbuscular mycorrhizas in decreasing aluminium phytotoxicity in acidic soils: A review. Mycorrhiza 2013, 23, 167–183. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, M.T.; Bohlool, B.B. Extracellular polysaccharide is not responsible for aluminum tolerance of Rhizobium leguminosarum bv. phaseoli CIAT899. Appl. Environ. Microbiol. 1992, 58, 1095–1101. [Google Scholar] [CrossRef]
- Roy, N.; Chakrabartty, P.K. Effect of aluminum on the production of siderophore by Rhizobium sp. (Cicer arietinum). Curr. Microbiol. 2000, 41, 5–10. [Google Scholar] [CrossRef]
- Avelar Ferreira, P.A.; Bomfeti, C.A.; Lima Soares, B.; de Souza Moreira, F.M. Efficient nitrogen-fixing Rhizobium strains isolated from amazonian soils are highly tolerant to acidity and aluminium. World J. Microbiol. Biotechnol. 2012, 28, 1947–1959. [Google Scholar] [CrossRef]
- Munns, D.; Keyser, H.; Fogle, V.; Hohenberg, J.; Righetti, T.; Lauter, D.; Zaroug, M.; Clarkin, K.; Whitacre, K. Tolerance of soil acidity in symbioses of mung bean with rhizobia. Agron. J. 1979, 71, 256–260. [Google Scholar] [CrossRef]
- Munns, D.N.; Hohenberg, J.S.; Righetti, T.L.; Lauter, D.J. Soil acidity tolerance of symbiotic and nitrogen-fertilized soybeans. Agron. J. 1981, 73, 407–410. [Google Scholar] [CrossRef]
- Arora, P.; Singh, G.; Tiwari, A. Effect of Microbial inoculation in combating the aluminium toxicity effect on growth of Zea mays. Cell. Mol. Biol. (Noisy-le-Grand) 2017, 63, 79–82. [Google Scholar] [CrossRef]
- Thakur, R.; Sharma, K.C.; Gulati, A.; Sud, R.K.; Gulati, A. Stress-tolerant Viridibacillus arenosi strain IHB B 7171 from tea rhizosphere as a potential broad-spectrum microbial inoculant. Indian J. Microbiol. 2017, 57, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Belimov, A.A.; Shaposhnikov, A.I.; Azarova, T.S.; Makarova, N.M.; Safronova, V.I.; Litvinskiy, V.A.; Nosikov, V.V.; Zavalin, A.A.; Tikhonovich, I.A. Microbial consortium of PGPR, rhizobia and arbuscular mycorrhizal fungus makes pea mutant SGECdt comparable with Indian mustard in cadmium tolerance and accumulation. Plants 2020, 9, 975. [Google Scholar] [CrossRef]
- Shaposhnikov, A.I.; Vishnevskaya, N.A.; Shakhnazarova, V.Y.; Belimov, A.A.; Strunnikova, O.K. The role of barley root exudates as a food source in the relationship between Fusarium culmorum and Pseudomonas fluorescens. Mycol. Phytopathol. 2019, 53, 311–331. [Google Scholar] [CrossRef]
- Kuo, Y.H.; Lambein, F.; Ikegami, F.; van Parijs, R. Isoxazolin-5-ones and amino acids in root exudates of pea and sweet pea seedlings. Plant Physiol. 1982, 70, 1283–1289. [Google Scholar] [CrossRef]
- Van Egeraat, A.W.S.M. Exudation of ninhydrin-positive compounds by pea seedling roots: A study of the sites of exudation and of the composition of the exudates. Plant Soil. 1975, 42, 37–47. [Google Scholar] [CrossRef]
- Short, G.E.; Lacy, M.L. Carbohydrate exudation from pea seeds effect of cultivar, seed age, seed color, and temperature. Phytopathology 1976, 66, 182–187. [Google Scholar] [CrossRef]
- Kuzmicheva, Y.V.; Shaposhnikov, A.I.; Petrova, S.N.; Makarova, N.M.; Tychinskaya, I.L.; Puhalsky, J.V.; Parahin, N.V.; Tikhonovich, I.A.; Belimov, A.A. Variety specific relationships between effects of rhizobacteria on root exudation, growth and nutrient uptake of soybean. Plant Soil 2017, 419, 83–96. [Google Scholar] [CrossRef]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Ann. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef]
- Vivanco, J.M.; Baluška, F. Secretions and Exudates in Biological Systems, Signaling and Communication in Plants; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1227–1248. [Google Scholar]
- Belimov, A.A.; Dodd, I.C.; Safronova, V.I.; Shaposhnikov, A.I.; Azarova, T.S.; Makarova, N.M.; Davies, W.J.; Tikhonovich, I.A. Rhizobacteria that produce auxins and contain ACC deaminase decrease amino acid concentrations in the rhizosphere and improve growth and yield of well-watered and water-limited potato (Solanum tuberosum). Ann. Appl. Biol. 2015, 167, 11–25. [Google Scholar] [CrossRef]
- Silambarasan, S.; Logeswari, P.; Valentine, A.; Cornejo, P. Role of Curtobacterium herbarum strain CAH5 on aluminum bioaccumulation and enhancement of Lactuca sativa growth under aluminum and drought stresses. Ecotox. Environ. Saf. 2019, 183, 109573. [Google Scholar] [CrossRef]
- Silambarasan, S.; Logeswari, P.; Cornejo, P.; Kannan, V.R. Role of plant growth-promoting rhizobacterial consortium in improving the Vigna radiata growth and alleviation of aluminum and drought stresses. Environ. Sci. Pollut. Res. 2019, 26, 27647–27659. [Google Scholar] [CrossRef] [PubMed]
- Silambarasan, S.; Logeswari, P.; Cornejo, P.; Kannan, V.R. Evaluation of the production of exopolysaccharide by plant growth promoting yeast Rhodotorula sp. strain CAH2 under abiotic stress conditions. Int. J. Biol. Macromol. 2019, 121, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Appanna, V.D.; Kepes, M.; Rochon, P. Aluminum tolerance in Pseudomonas fluoreseens ATCC 13525: Involvement of a gelatinous lipid-rich residue. FEMS Microbiol. Lett. 1994, 119, 295–301. [Google Scholar] [CrossRef]
- Appanna, V.D.; St Pierre, M. Influence of phosphate on aluminum tolerance in Pseudomonas fluorescens. FEMS Microbiol. Lett. 1994, 124, 327–332. [Google Scholar] [CrossRef]
- Belimov, A.A.; Puhalsky, I.V.; Safronova, V.I.; Shaposhnikov, A.I.; Vishnyakova, M.A.; Semenova, E.V.; Zinovkina, N.Y.; Makarova, N.M.; Wenzel, W.; Tikhonovich, I.A. Role of plant genotype and soil conditions in symbiotic plant-microbe interactions for adaptation of plants to cadmium polluted soils. Water Air Soil Poll. 2015, 226, 264. [Google Scholar] [CrossRef]
- Silva, I.R.; Smyth, T.J.; Israel, D.W.; Raper, C.D.; Rufty, T.W. Magnesium ameliorates aluminum rhizotoxicity in soybean by increasing citric acid production and exudation by roots. Plant Cell Physiol. 2001, 42, 546–554. [Google Scholar] [CrossRef]
- Liao, H.; Wan, H.; Shaff, J.; Wang, X.; Yan, X.; Kochian, L.V. Phosphorus and aluminum interactions in soybean in relation to aluminum tolerance. Exudation of specific organic acids from different regions of the intact root system. Plant Physiol. 2006, 141, 674–684. [Google Scholar] [CrossRef]
- Kikui, S.; Sasaki, T.; Osawa, H.; Matsumoto, H.; Yamamoto, Y. Malate enhances recovery from aluminum-caused inhibition of root elongation in wheat. Plant Soil 2007, 290, 1. [Google Scholar] [CrossRef]
- Chen, Y.T.; Wang, Y.; Yeh, K.C. Role of root exudates in metal acquisition and tolerance. Curr. Opin. Plant Biol. 2017, 39, 66–72. [Google Scholar] [CrossRef]
- Ranjan, A.; Sinha, R.; Lal, S.K.; Bishi, S.K.; Singh, A.K. Phytohormone signalling and cross-talk to alleviate aluminium toxicity in plants. Plant Cell Rep. 2021, 40, 1331–1343. [Google Scholar] [CrossRef]
- Chai, Y.N.; Futrell, S.; Schachtman, D.P. Assessment of bacterial inoculant delivery methods for cereal crops. Front. Microbiol. 2022, 13, 791110. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Yamaya, T. Inhibition of potassium uptake and regulation of membrane-associated Mg2+-ATPase activity of pea roots by aluminium. Soil Sci. Plant Nutr. 1986, 32, 179–188. [Google Scholar] [CrossRef]
- Kamran, M.; Ramesh, S.A.; Gilliham, M.; Tyerman, S.D.; Bose, J. Role of TaALMT1 malate-GABA transporter in alkaline pH tolerance of wheat. Plant Cell Environ. 2020, 43, 2443–2459. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nishizawa, N.K. Iron uptake, translocation, and regulation in higher plants. Ann. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Bi, S.; Wang, S.; Ding, Q. Variation of wheat root exudates under aluminum stress. J. Agric. Food Chem. 2006, 54, 10040–10046. [Google Scholar] [CrossRef] [PubMed]
- Owens, A.G.; Jones, D.L. Competition for amino acids between wheat roots and rhizosphere microorganisms and the role of amino acids in plant N acquisition. Soil Biol. Biochem. 2001, 33, 651–657. [Google Scholar] [CrossRef]
- Ryan, P.R.; Delhaize, E.; Jones, D.L. Function and mechanism of organic anion exudation from plant roots. Ann. Rev. Plant Biol. 2001, 52, 527–560. [Google Scholar] [CrossRef]
- Jones, D.L.; Clode, P.L.; Kilburn, M.R.; Stockdale, E.A.; Murphy, D.V. Competition between plant and bacterial cells at the microscale regulates the dynamics of nitrogen acquisition in wheat (Triticum aestivum). New Phytol. 2013, 200, 796–807. [Google Scholar] [CrossRef]
- Moe, L.A. Amino acids in the rhizosphere: From plants to microbes. Am. J. Bot. 2013, 100, 1692–1705. [Google Scholar] [CrossRef]
- Phillips, D.A.; Fox, T.C.; King, M.D.; Bhuvaneswari, T.V.; Teuber, L.R. Microbial products trigger amino acid exudation from plant roots. Plant Physiol. 2004, 136, 2887–2894. [Google Scholar] [CrossRef]
- Naher, U.A.; Radziah, O.; Halimi, M.S.; Shamsuddin, Z.H.; Mohd Razi, I. Effect of inoculation on root exudates carbon sugar and amino acids production of different rice varieties. Res. J. Microbiol. 2008, 3, 580–587. [Google Scholar] [CrossRef]
- Kudoyarova, G.R.; Melentiev, A.I.; Martynenko, E.V.; Timergalina, L.N.; Arkhipova, T.N.; Shendel, G.V.; Kuz’mina, L.Y.; Dodd, I.C.; Veselov, S.Y. Cytokinin producing bacteria stimulate amino acid deposition from wheat roots. Plant Physiol. Bioch. 2014, 83, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Kravchenko, L.V.; Azarova, T.S.; Makarova, N.M.; Tikhonovich, I.A. The effect of tryptophan present in plant root exudates on the phytostimulating activity of rhizobacteria. Microbiology 2004, 73, 156–158. [Google Scholar] [CrossRef]
- Kravchenko, L.V.; Shapozhnikov, A.I.; Makarova, N.M.; Azarova, T.S.; L’vova, K.A.; Kostyuk, I.I.; Lyapunova, O.A.; Tikonovich, I.A. Exometabolites of bread wheat and tomato affecting the plant–microbe interactions in the rhizosphere. Russ. J. Plant Physiol. 2011, 58, 936–940. [Google Scholar] [CrossRef]
- Kamilova, F.; Kravchenko, L.V.; Shaposhnikov, A.I.; Makarova, N.M.; Azarova, T.S.; Lugtenberg, B. Organic acids, sugars and L-tryptophan in exudates of vegetables growing on stonewool and their effects on activities of rhizosphere bacteria. Mol. Plant–Microbe Interact. 2006, 19, 250–256. [Google Scholar] [CrossRef]
- Wang, P.; Yu, W.; Zhang, J.; Rengel, Z.; Xu, J.; Han, Q.; Chen, L.; Li, K.; Yu, Y.; Chen, Q. Auxin enhances aluminium induced citrate exudation through upregulation of GmMATE and activation of the plasma membrane H+-ATPase in soybean roots. Ann. Bot. 2016, 118, 933–940. [Google Scholar] [CrossRef]
- Wu, D.; Shen, H.; Yokawa, K.; Baluška, F. Alleviation of aluminium induced cell rigidity by overexpression of OsPIN2 in rice roots. J. Exp. Bot. 2014, 65, 5305–5315. [Google Scholar] [CrossRef]
- Zerrouk, I.Z.; Benchabane, M.; Khelifi, L.; Yokawa, K.; Ludwig-Müller, J.; Baluska, F.A. Pseudomonas strain isolated from date-palm rhizospheres improves root growth and promotes root formation in maize exposed to salt and aluminum stress. J. Plant Physiol. 2016, 191, 111–119. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Rossi, M.J.; Glick, B.R. Ethylene and 1-aminocyclopropane-1-carboxylate (ACC) in plant-bacterial interactions. Front. Plant Sci. 2018, 9, 114. [Google Scholar] [CrossRef]
- Gupta, S.; Pandey, S. Unravelling the biochemistry and genetics of ACC deaminase-An enzyme alleviating the biotic and abiotic stress in plants. Plant Gene 2019, 18, 100175. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, V.; Kaur Sidhu, G.; Datta, S.; Singh Dhanjal, D.; Koul, B.; Singh Janeja, H.; Singh, J. Plant growth promoting rhizobacteria from heavy metal contaminated soil promote growth attributes of Pisum sativum L. Biocatal. Agric. Biotechnol. 2019, 17, 665–671. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.D.C.; Glick, B.R.; Santoyo, G. ACC deaminase in plant growth-promoting bacteria (PGPB): An efficient mechanism to counter salt stress in crops. Microbiol. Res. 2020, 235, 126439. [Google Scholar] [CrossRef] [PubMed]
- Paço, A.; da-Silva, J.R.; Torres, D.P.; Glick, B.R.; Brígido, C. Exogenous ACC deaminase is key to improving the performance of pasture legume-rhizobial symbioses in the presence of a high manganese concentration. Plants 2020, 9, 1630. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R.; Penrose, D.M.; Li, J. A model for the lowering of plant ethylene concentrations by plant growth-promoting bacteria. J. Theor. Biol. 1998, 190, 63–68. [Google Scholar] [CrossRef]
- Glick, B.R.; Karaturovic, D.M.; Newell, P.C. A novel procedure for rapid isolation of plant growth promoting pseudomonads. Can. J. Microbiol. 1995, 41, 533–536. [Google Scholar] [CrossRef]
- Suarez, A.; Guttler, A.; Stratz, M.; Staendner, L.H.; Timmis, K.N.; Guzman, C.A. Green fluorescent protein-based reporter systems for genetic analysis of bacteria including monocopy applications. Gene 1997, 196, 69–74. [Google Scholar] [CrossRef]
- Saleh, S.S.; Glick, B.R. Involvement of gacS and rpoS in enhancement of the plant growth-promoting capabilities of Enterobacter cloacae CAL2 and UW4. Can. J. Microbiol. 2001, 47, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–258. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | Ps. fluorescens SPB2137 | Ps. fluorescens SPB2137gfp |
|---|---|---|
| Production of phytohormones (µ mL−1): | ||
| Indole-3-acetic acid | 572 ± 86 a | 395 ± 48 a |
| Indole-3-butyric acid | 1224 ± 183 a | 1326 ± 48 a |
| Indole-3-lactic acid | 41 ± 7 a | 59 ± 6 a |
| Abscisic acid | nd | nd |
| Salicylic acid | nd | nd |
| ACC deaminase activity (µM αKB mg−1 protein h−1) | 6.1 ± 0.4 a | 5.8 ± 0.3 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belimov, A.A.; Shaposhnikov, A.I.; Azarova, T.S.; Syrova, D.S.; Kitaeva, A.B.; Ulyanich, P.S.; Yuzikhin, O.S.; Sekste, E.A.; Safronova, V.I.; Vishnyakova, M.A.; et al. Rhizobacteria Mitigate the Negative Effect of Aluminum on Pea Growth by Immobilizing the Toxicant and Modulating Root Exudation. Plants 2022, 11, 2416. https://doi.org/10.3390/plants11182416
Belimov AA, Shaposhnikov AI, Azarova TS, Syrova DS, Kitaeva AB, Ulyanich PS, Yuzikhin OS, Sekste EA, Safronova VI, Vishnyakova MA, et al. Rhizobacteria Mitigate the Negative Effect of Aluminum on Pea Growth by Immobilizing the Toxicant and Modulating Root Exudation. Plants. 2022; 11(18):2416. https://doi.org/10.3390/plants11182416
Chicago/Turabian StyleBelimov, Andrey A., Alexander I. Shaposhnikov, Tatiana S. Azarova, Darya S. Syrova, Anna B. Kitaeva, Pavel S. Ulyanich, Oleg S. Yuzikhin, Edgar A. Sekste, Vera I. Safronova, Margarita A. Vishnyakova, and et al. 2022. "Rhizobacteria Mitigate the Negative Effect of Aluminum on Pea Growth by Immobilizing the Toxicant and Modulating Root Exudation" Plants 11, no. 18: 2416. https://doi.org/10.3390/plants11182416
APA StyleBelimov, A. A., Shaposhnikov, A. I., Azarova, T. S., Syrova, D. S., Kitaeva, A. B., Ulyanich, P. S., Yuzikhin, O. S., Sekste, E. A., Safronova, V. I., Vishnyakova, M. A., Tsyganov, V. E., & Tikhonovich, I. I. (2022). Rhizobacteria Mitigate the Negative Effect of Aluminum on Pea Growth by Immobilizing the Toxicant and Modulating Root Exudation. Plants, 11(18), 2416. https://doi.org/10.3390/plants11182416