
Chemical Constituents of Callistemon subulatus and Their Anti-Pancreatic Cancer Activity against Human PANC-1 Cell Line

 ,
, 

Abstract
:1. Introduction
2. Results
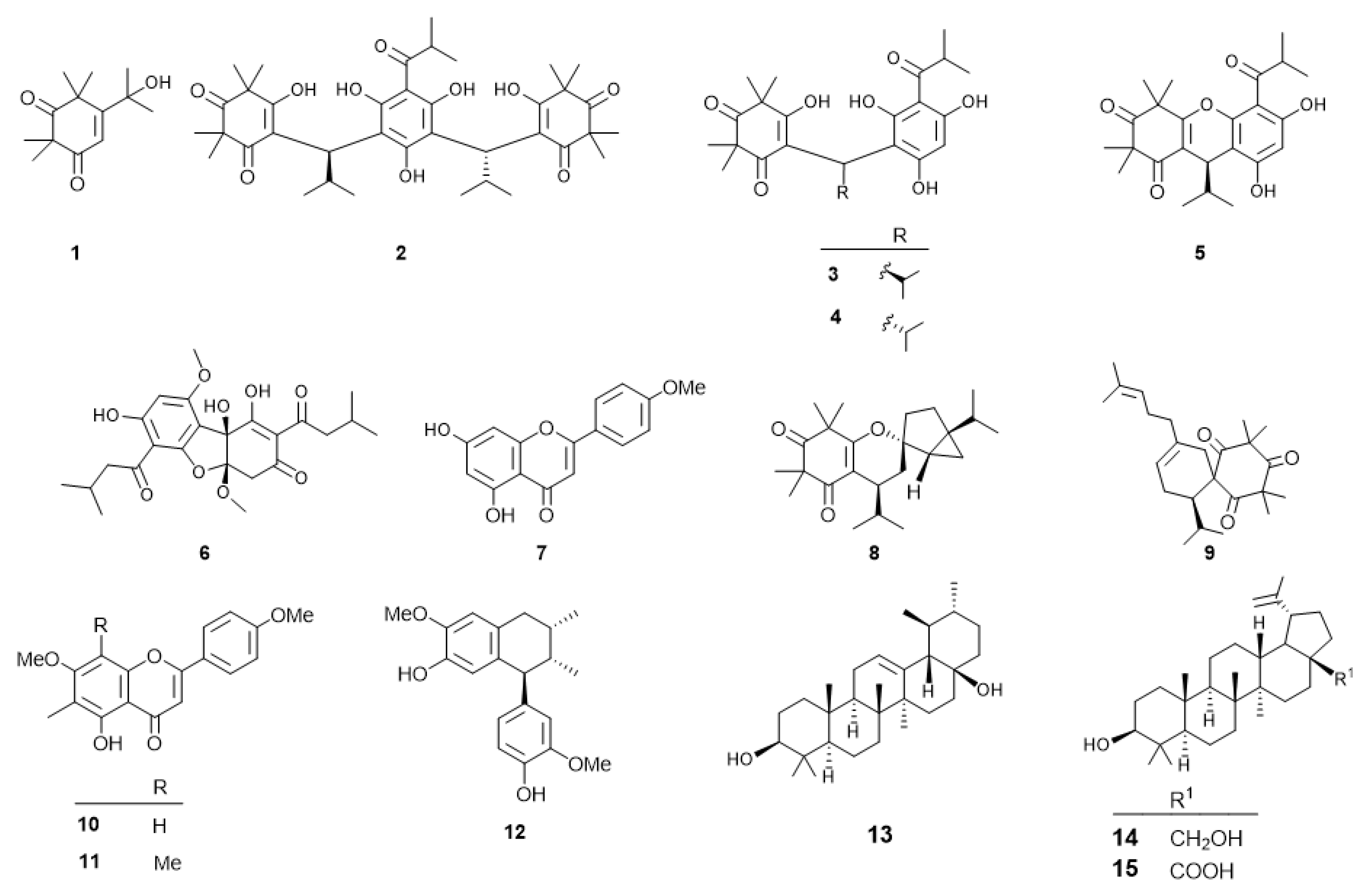
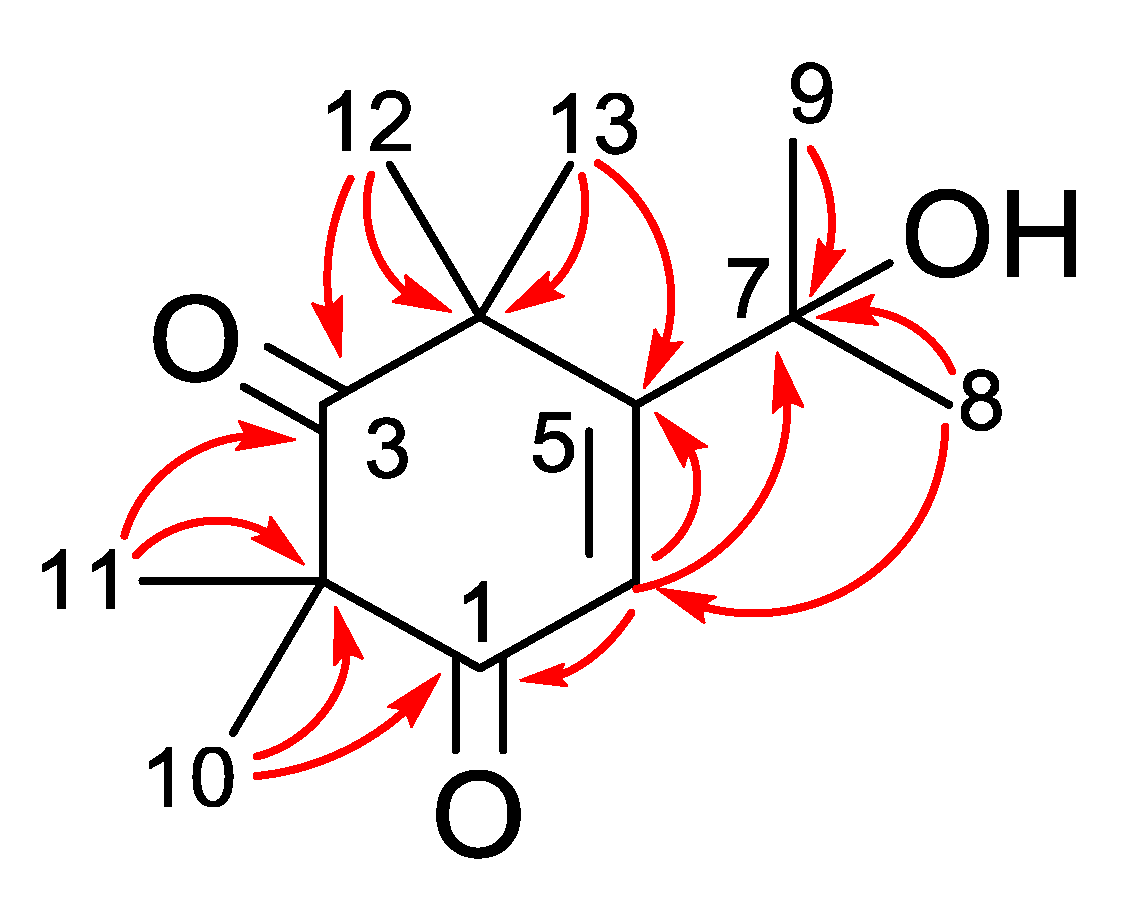
2.1. Phytochemical Investigation of Callistemon Subulatus
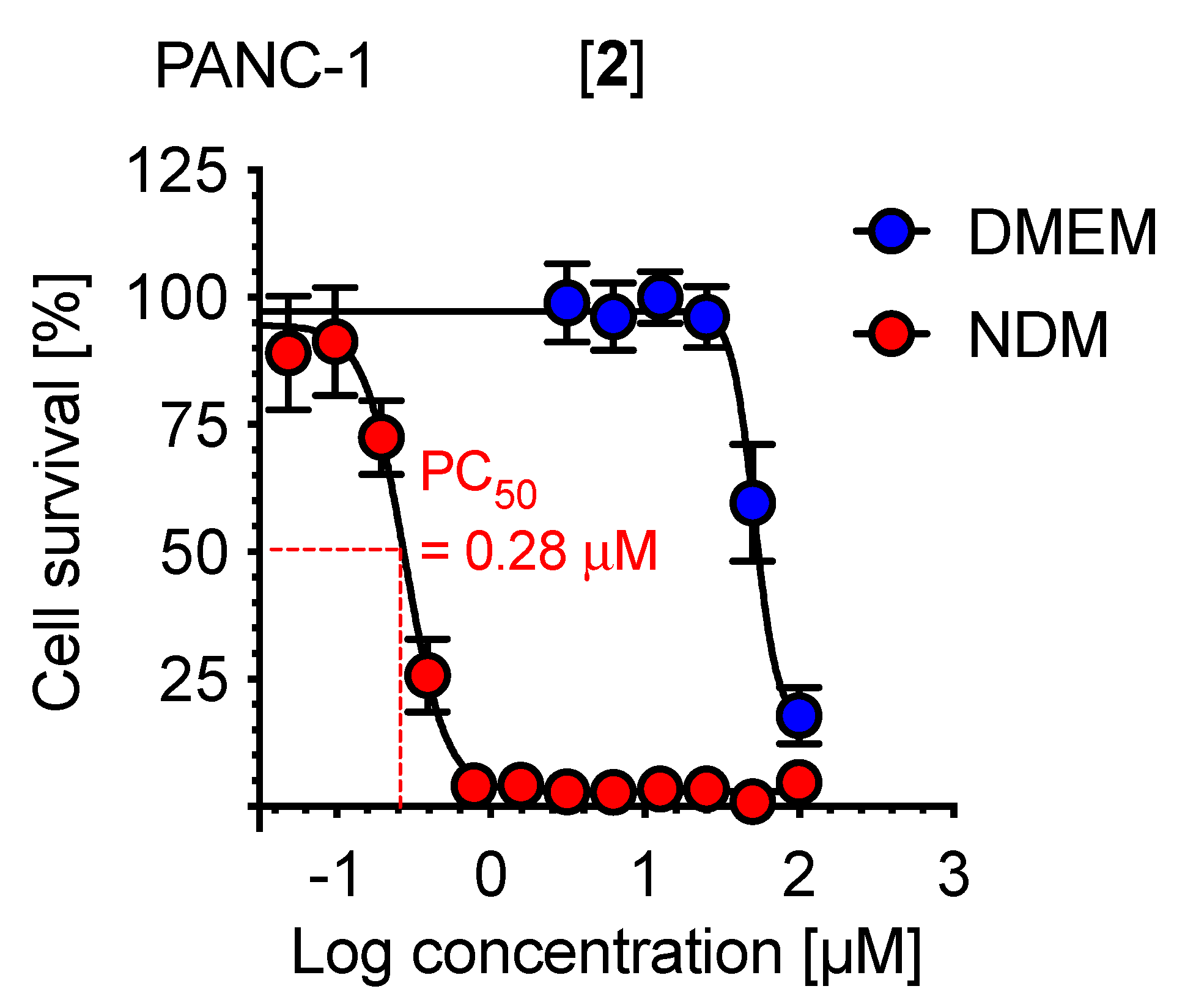
2.2. Preferential Cytotoxicity against PANC-1 Cells
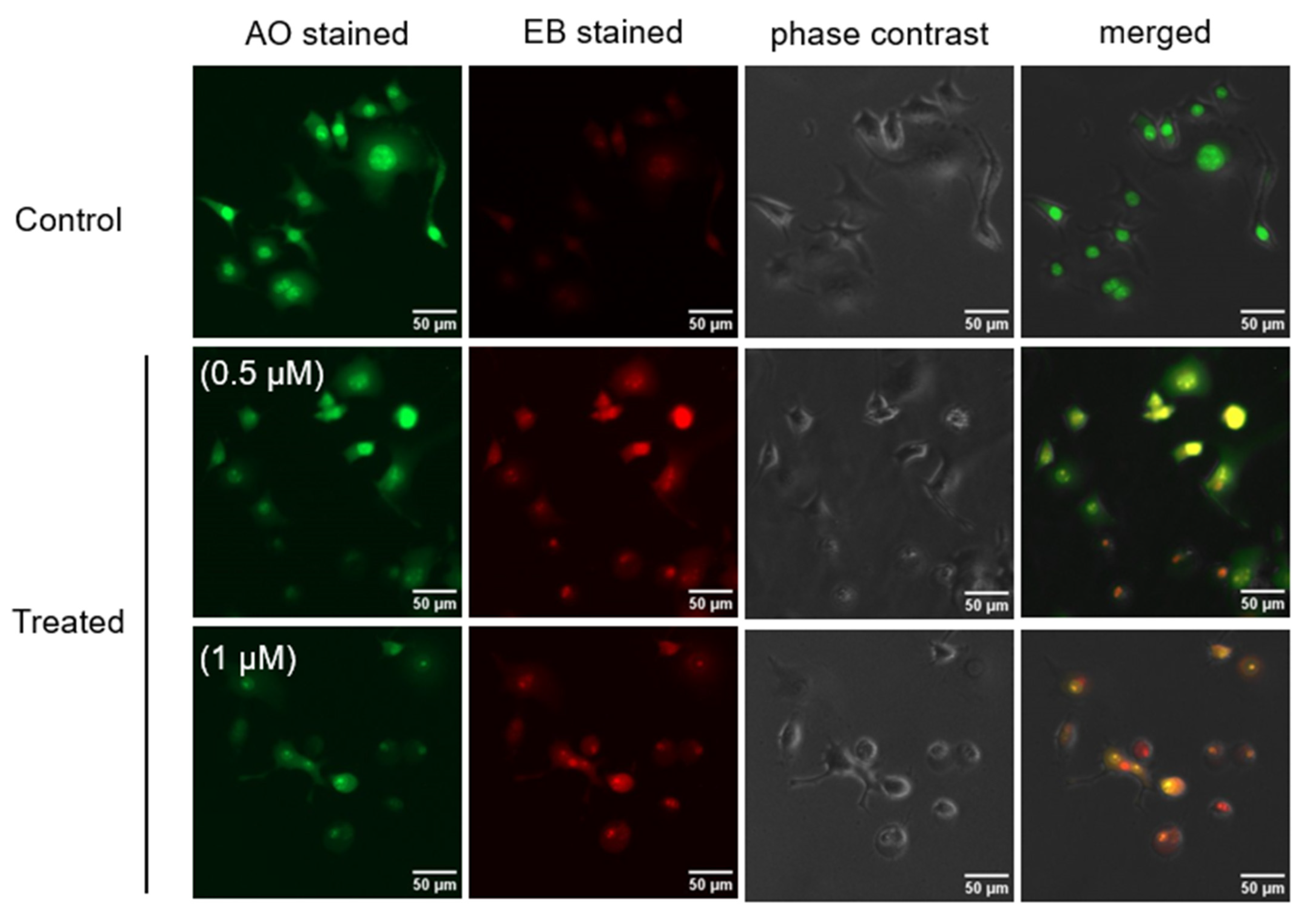
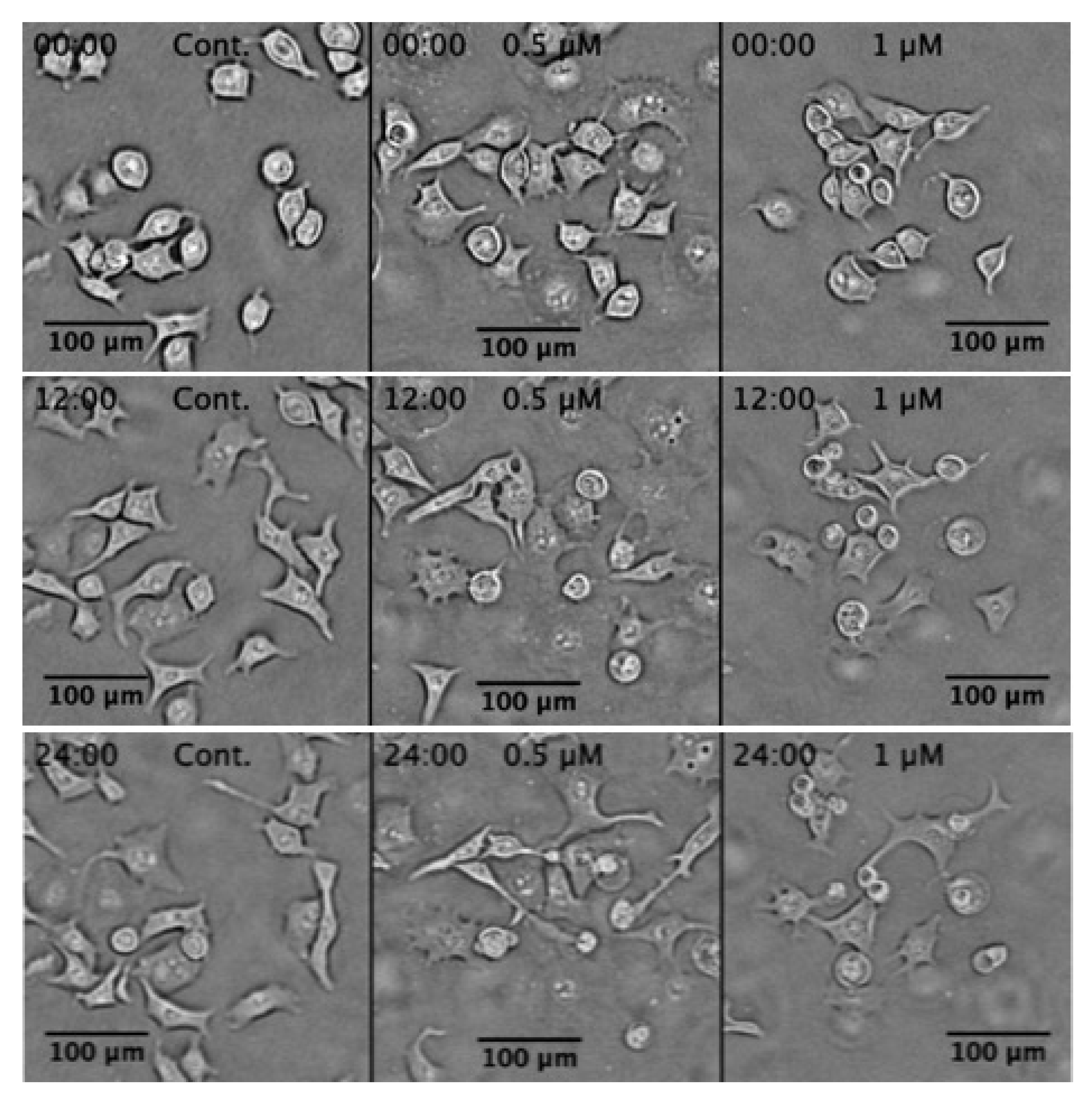
2.3. Morphological Change in PANC-1 Cells in NDM
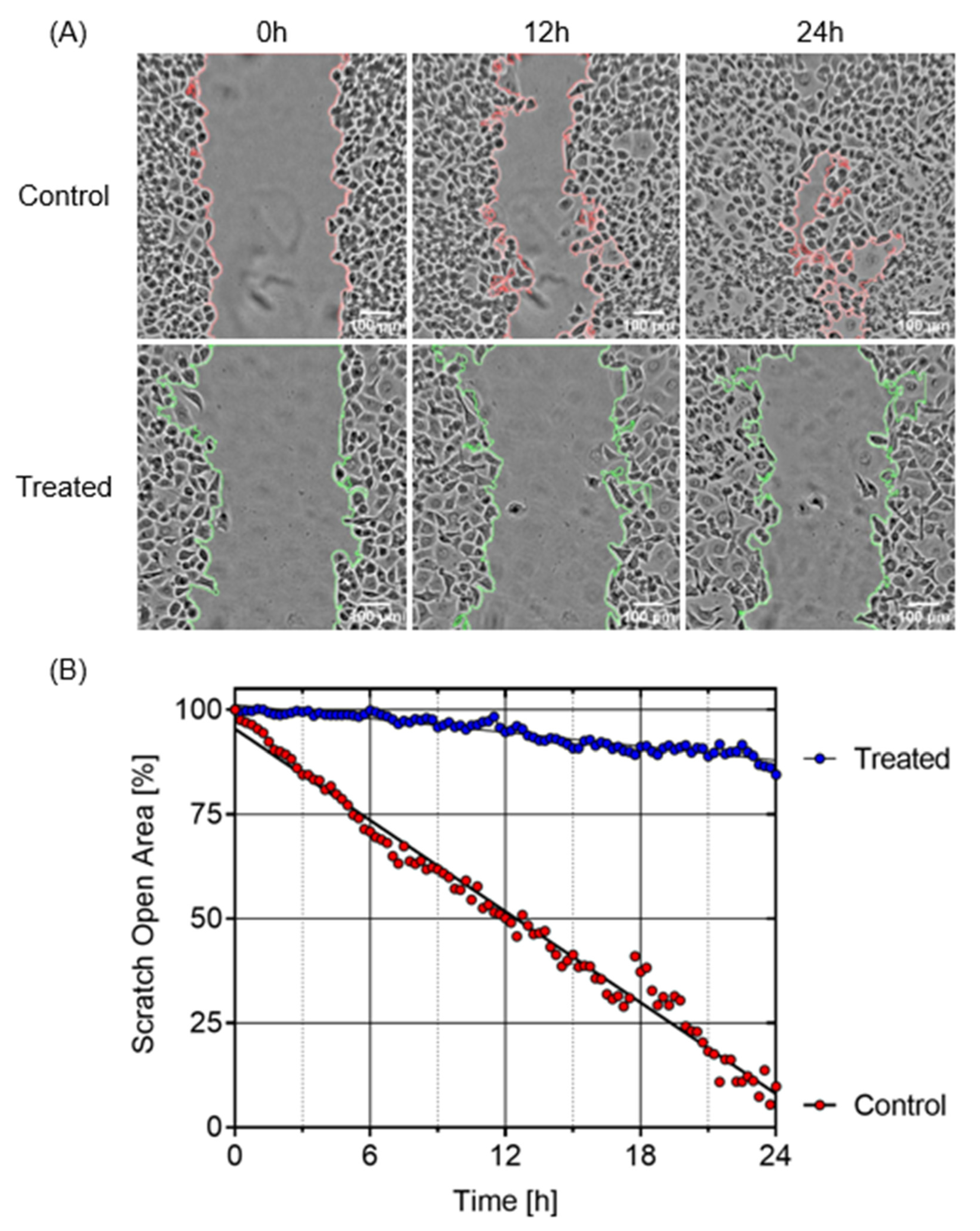
2.4. Inhibition of PANC-1 Cell Migration in DMEM
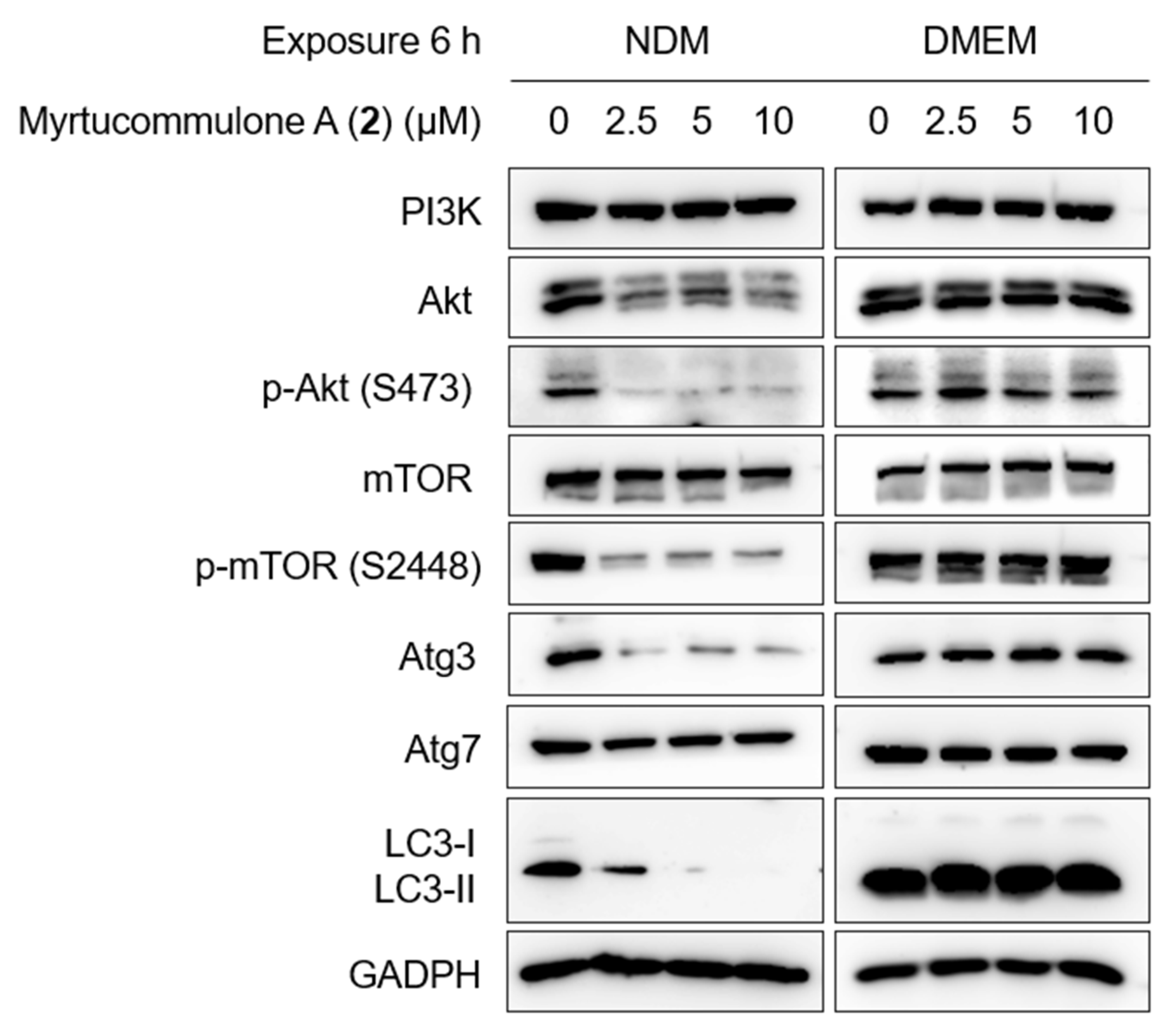
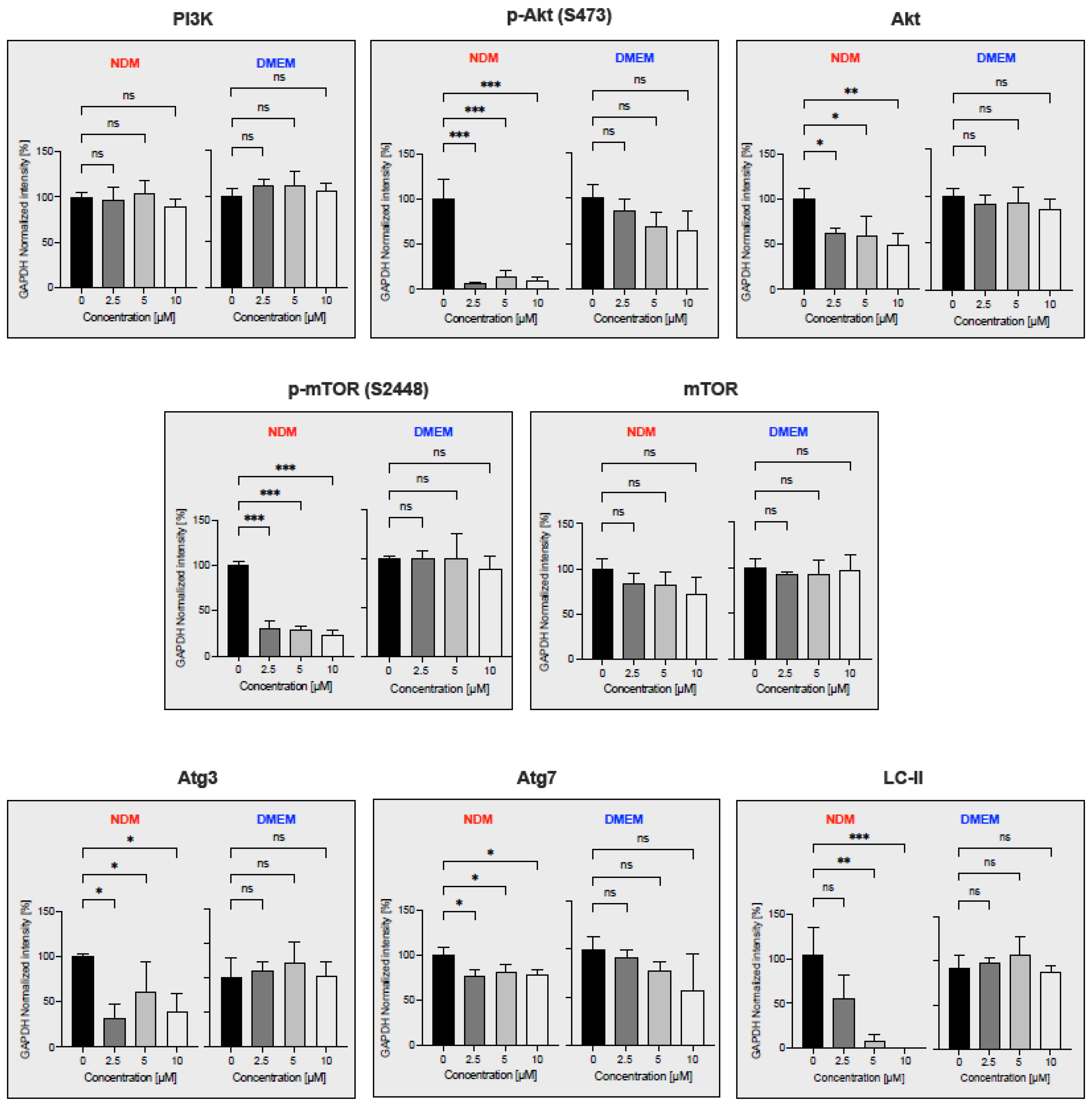
2.5. Inhibition of PI3K/Akt/mTOR and Autophagy Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedure
4.2. Chemical and Antibodies
4.3. Extraction and Isolation
4.4. Cell Line and Cell Culture of PANC-1
4.5. Preferential Cytotoxicity Assay against PANC-1 Cells
4.6. Morphological Assessment of PANC-1 Cells
4.7. Live Cell imaging of PANC-1 Cells
4.8. Cell Migration Assay
4.9. Western Blot Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bengtsson, A.; Andersson, R.; Ansari, D. The actual 5-year survivors of pancreatic ductal adenocarcinoma based on real-world data. Sci. Rep. 2020, 10, 16425. [Google Scholar] [CrossRef]
- Ushio, J.; Kanno, A.; Ikeda, E.; Ando, K.; Nagai, H.; Miwata, T.; Kawasaki, Y.; Tada, Y.; Yokoyama, K.; Numao, N.; et al. Pancreatic ductal adenocarcinoma: Epidemiology and risk factors. Diagnostics 2021, 11, 562. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Rawla, P.; Sunkara, T.; Gaduputi, V. Epidemiology of pancreatic cancer: Global trends, etiology and risk factors. World. J. Oncol. 2019, 10, 10–27. [Google Scholar] [CrossRef] [PubMed]
- Klatte, D.C.F.; Wallace, M.B.; Löhr, M.; Bruno, M.J.; van Leerdam, M.E. Hereditary pancreatic cancer. Best Pract. Res. Clin. Gastroenterol. 2022, 58–59, 101783. [Google Scholar] [CrossRef]
- Iyengar, S.; Nevala-Plagemann, C.; Garrido-Laguna, I. Updates on adjuvant and neoadjuvant treatment strategies for surgically resectable and borderline resectable pancreatic ductal adenocarcinoma. Ther. Adv. Med. Oncol. 2021, 13, 175883592110458601. [Google Scholar] [CrossRef]
- Fine, R.L.; Fogelman, D.R.; Schreibman, S.M.; Desai, M.; Sherman, W.; Strauss, J.; Guba, S.; Andrade, R.; Chabot, J. The gemcitabine, docetaxel, and capecitabine (GTX) regimen for metastatic pancreatic cancer: A retrospective analysis. Cancer Chemother. Pharmacol. 2008, 61, 167–175. [Google Scholar] [CrossRef]
- Louvet, C.; Labianca, R.; Hammel, P.; Lledo, G.; Zampino, M.G.; André, T.; Zaniboni, A.; Ducreux, M.; Aitini, E.; Taïeb, J.; et al. Gemcitabine in combination with oxaliplatin compared with gemcitabine alone in locally advanced or metastatic pancreatic cancer: Results of a GERCOR and GISCAD phase III Trial. J. Clin. Oncol. 2005, 23, 3509–3516. [Google Scholar] [CrossRef]
- Herrmann, R.; Bodoky, G.; Ruhstaller, T.; Glimelius, B.; Bajetta, E.; Schüller, J.; Saletti, P.; Bauer, J.; Figer, A.; Pestalozzi, B.; et al. Gemcitabine plus capecitabine compared with gemcitabine alone in advanced pancreatic cancer: A randomized, multicenter, phase III trial of the Swiss Group for clinical cancer research and the Central European Cooperative Oncology group. J. Clin. Oncol. 2007, 25, 2212–2217. [Google Scholar] [CrossRef]
- Moore, M.J.; Goldstein, D.; Hamm, J.; Figer, A.; Hecht, J.R.; Gallinger, S.; Au, H.J.; Murawa, P.; Walde, D.; Wolff, R.A.; et al. Erlotinib plus gemcitabine compared with gemcitabine alone in patients with advanced pancreatic cancer: A phase III trial of the National Cancer Institute of Canada Clinical Trials Group. J. Clin. Oncol. 2007, 25, 1960–1966. [Google Scholar] [CrossRef] [Green Version]
- Colucci, G.; Labianca, R.; Di Costanzo, F.; Gebbia, V.; Cartenì, G.; Massidda, B.; Dapretto, E.; Manzione, L.; Piazza, E.; Sannicolò, M.; et al. Randomized phase III trial of gemcitabine plus cisplatin compared with single-agent gemcitabine as first-line treatment of patients with advanced pancreatic cancer: The GIP-1 study. J. Clin. Oncol. 2010, 28, 1645–1651. [Google Scholar] [CrossRef]
- Adel, N. Current treatment landscape and emerging therapies for pancreatic cancer. Am. J. Manag. Care 2019, 25 (Suppl. 1), S3–S10. [Google Scholar] [PubMed]
- Gargiulo, P.; Dietrich, D.; Herrmann, R.; Bodoky, G.; Ruhstaller, T.; Scheithauer, W.; Glimelius, B.; Berardi, S.; Pignata, S.; Brauchli, P. Predicting mortality and adverse events in patients with advanced pancreatic cancer treated with palliative gemcitabine-based chemotherapy in a multicentre phase III randomized clinical trial: The APC-SAKK risk scores. Ther. Adv. Med. Oncol. 2019, 11, 1758835918818351. [Google Scholar] [CrossRef] [PubMed]
- De Dosso, S.; Siebenhüner, A.R.; Winder, T.; Meisel, A.; Fritsch, R.; Astaras, C.; Szturz, P.; Borner, M. Treatment landscape of metastatic pancreatic cancer. Cancer Treat. Rev. 2021, 96, 102180. [Google Scholar] [CrossRef] [PubMed]
- Awale, S.; Lu, J.; Kalauni, S.K.; Kurashima, Y.; Tezuka, Y.; Kadota, S.; Esumi, H. Identification of arctigenin as an antitumor agent having the ability to eliminate the tolerance of cancer cells to nutrient starvation. Cancer Res. 2006, 66, 1751–1757. [Google Scholar] [CrossRef]
- Awale, S.; Nakashima, E.M.N.; Kalauni, S.K.; Tezuka, Y.; Kurashima, Y.; Lu, J.; Esumi, H.; Kadota, S. Angelmarin, a novel anti-cancer agent able to eliminate the tolerance of cancer cells to nutrient starvation. Bioorg. Med. Chem. Lett. 2006, 16, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Fayez, S.; Feineis, D.; Mudogo, V.; Awale, S.; Bringmann, G. Ancistrolikokines E–H and related 5,8′-coupled naphthylisoquinoline alkaloids from the Congolese liana Ancistrocladus likoko with antiausterity activities against PANC-1 human pancreatic cancer cells. RSC Adv. 2017, 7, 53740–53751. [Google Scholar] [CrossRef]
- Awale, S.; Tawila, A.M.; Dibwe, D.F.; Ueda, J.; Sun, S.; Athikomkulchai, S.; Balachandran, C.; Saiki, I.; Matsumoto, K.; Esumi, H. Highly oxygenated antiausterity agents from the leaves of Uvaria Dac. Bioorg. Med. Chem. Lett. 2017, 27, 1967–1971. [Google Scholar] [CrossRef]
- Nguyen, M.T.T.; Nguyen, H.X.; Le, T.H.; Do, T.N.V.; Dang, P.H.; Pham, T.V.; Giang, T.T.M.; Sun, S.; Kim, M.J.; Tawila, A.M.; et al. A New flavanone derivative from the rhizomes of Boesenbergia pandurata. Nat. Prod. Res. 2022, 36, 1959–1965. [Google Scholar] [CrossRef]
- Sun, S.; Dibwe, D.F.; Kim, M.J.; Omar, A.M.; Phan, N.D.; Fujino, H.; Pongterdsak, N.; Chaithatwatthana, K.; Phrutivorapongkul, A.; Awale, S. A new anti-austerity agent, 4′-O-methylgrynullarin from Derris scandens induces PANC-1 human pancreatic cancer cell death under nutrition starvation via inhibition of Akt/MTOR pathway. Bioorg. Med. Chem. Lett. 2021, 40, 127967. [Google Scholar] [CrossRef]
- Phan, N.D.; Omar, A.M.; Sun, S.; Maneenet, J.; Dibwe, D.F.; Sato, M.; Kalauni, S.K.; Toyooka, N.; Fujii, T.; Awale, S. Abietane diterpenes from Abies spectabilis and their anti-pancreatic cancer activity against the MIA PaCa-2 cell line. Bioorg. Med. Chem. Lett. 2022, 66, 128723. [Google Scholar] [CrossRef]
- Awale, S.; Okada, T.; Dibwe, D.F.; Maruyama, T.; Takahara, S.; Okada, T.; Endo, S.; Toyooka, N. Design and synthesis of functionalized coumarins as potential anti-austerity agents that eliminates cancer cells’ tolerance to nutrition starvation. Bioorg. Med. Chem. Lett. 2019, 29, 1779–1784. [Google Scholar] [CrossRef]
- Tawila, A.M.; Sun, S.; Kim, M.J.; Omar, A.M.; Dibwe, D.F.; Ueda, J.; Toyooka, N.; Awale, S. Chemical constituents of Callistemon citrinus from Egypt and their antiausterity activity against PANC-1 human pancreatic cancer cell line. Bioorg. Med. Chem. Lett. 2020, 30, 127352. [Google Scholar] [CrossRef]
- Omar, A.M.; Sun, S.; Kim, M.J.; Phan, N.D.; Tawila, A.M.; Awale, S. Benzophenones from Betula alnoides with antiausterity activities against the PANC-1 human pancreatic cancer cell line. J. Nat. Prod. 2021, 84, 1607–1616. [Google Scholar] [CrossRef]
- Sun, S.; Kim, M.J.; Dibwe, D.F.; Omar, A.M.; Athikomkulchai, S.; Phrutivorapongkul, A.; Okada, T.; Tsuge, K.; Toyooka, N.; Awale, S. Anti-austerity activity of Thai medicinal plants: Chemical constituents and anti-pancreatic cancer activities of Kaempferia parviflora. Plants 2021, 10, 229. [Google Scholar] [CrossRef]
- Ikeda, M.; Sato, A.; Mochizuki, N.; Toyosaki, K.; Miyoshi, C.; Fujioka, R.; Mitsunaga, S.; Ohno, I.; Hashimoto, Y.; Takahashi, H.; et al. Phase I Trial of GBS-01 for Advanced Pancreatic Cancer Refractory to Gemcitabine. Cancer Sci. 2016, 107, 1818–1824. [Google Scholar] [CrossRef]
- Mady, M.S.; Elsayed, H.E.; El-Sayed, E.K.; Hussein, A.A.; Ebrahim, H.Y.; Moharram, F.A. Polyphenolic profile and ethno pharmacological activities of Callistemon subulatus (cheel) craven leaves cultivated in Egypt. J. Ethnopharmacol. 2022, 284, 114698. [Google Scholar] [CrossRef]
- Ibrahim, N.; Moussa, A.Y. Comparative metabolite profiling of Callistemon macropunctatus and Callistemon subulatus volatiles from different geographical origins. Ind. Crops Prod. 2020, 147, 112222. [Google Scholar] [CrossRef]
- Gad, H.A.; Ayoub, I.M.; Wink, M. Phytochemical profiling and seasonal variation of essential oils of three Callistemon species cultivated in Egypt. PLoS ONE 2019, 14, e0219571. [Google Scholar] [CrossRef]
- Fayemi, P.O.; Öztürk, I.; Özcan, C.; Muguruma, M.; Yetim, H.; Sakata, R.; Ahhmed, A. Antimicrobial activity of extracts of Callistemon citrinus flowers and leaves against listeria monocytogenes in beef burger. J. Food Meas. Charact. 2017, 11, 924–929. [Google Scholar] [CrossRef]
- Kumar, R.; Gupta, A.; Singh, A.K.; Bishayee, A.; Pandey, A.K. The antioxidant and antihyperglycemic activities of bottlebrush plant (Callistemon lanceolatus) stem extracts. Medicines 2020, 7, 11. [Google Scholar] [CrossRef]
- Sudhakar, M.; Rao, C.V.; Rao, A.L.; Ramesh, A.; Srinivas, N.; Raju, D.B.; Murthy, B.K. Antinociceptive and anti-inflammatory effects of the standardized oil of Indian Callistemon lanceolatus leaves in experimental animals. East Cent. Afr. J. Pharm. Sci. 2004, 7, 10–15. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, V.; Prakash, O. Pharmacognostic study and anti-inflammatory activity of Callistemon lanceolatus leaf. Asian Pac. J. Trop. Biomed. 2011, 1, 177–181. [Google Scholar] [CrossRef]
- Ahmed, A.H. Phytochemical and cytotoxicity studies of Callistemon viminalis leaves extract growing in Egypt. Curr. Pharm. Biotechnol. 2020, 21, 219–225. [Google Scholar] [CrossRef]
- Hans, M.; Charpentier, M.; Huch, V.; Jauch, J.; Bruhn, T.; Bringmann, G.; Quandt, D. Stereoisomeric composition of natural myrtucommulone A. J. Nat. Prod. 2015, 78, 2381–2389. [Google Scholar] [CrossRef]
- Fiorini-Puybaret, C.; Aries, M.-F.; Fabre, B.; Mamatas, S.; Luc, J.; Degouy, A.; Ambonati, M.; Mejean, C.; Poli, F. Pharmacological properties of Myrtacine® and its potential value in acne treatment. Planta Med. 2011, 77, 1582–1589. [Google Scholar] [CrossRef]
- Kashman, Y.; Rotstein, A.; Lifshitz, A. The structure determination of two new acylphloroglucinols from Myrtus communis L. Tetrahedron 1974, 30, 991–997. [Google Scholar] [CrossRef]
- Xiang, Y.-Q.; Liu, H.-X.; Zhao, L.-Y.; Xu, Z.-F.; Tan, H.-B.; Qiu, S.-X. Callistemenonone A, a novel dearomatic dibenzofuran-type acylphloroglucinol with antimicrobial activity from Callistemon viminalis. Sci. Rep. 2017, 7, 2363. [Google Scholar] [CrossRef]
- Yang, W.-J.; Liu, C.; Gu, Z.-Y.; Zhang, X.-Y.; Cheng, B.; Mao, Y.; Xue, G.-P. Protective effects of acacetin isolated from Ziziphora clinopodioides Lam. (Xintahua) on neonatal rat cardiomyocytes. Chin. Med. 2014, 9, 28. [Google Scholar] [CrossRef]
- Wu, L.; Wang, X.; Li, R.; Zhang, Y.; Yang, M.; Luo, J.; Kong, L.-Y. Callistiviminenes A-O: Diverse adducts of β-triketone and sesqui- or monoterpene from the fruits of Callistemon viminalis. Phytochemistry 2016, 131, 140–149. [Google Scholar] [CrossRef]
- Cuong, N.M.; Khanh, P.N.; Duc, H.V.; Huong, T.T.; Kim, Y.-C.; Long, P.Q.; Kim, Y.H. Flavonoids and triterpenoids from Callistemon citrinus and their inhibitory effect on no production in LPS-stimulated RAW264.7 macrophages. Vietnam J. Sci. Technol. 2016, 54, 214–223. [Google Scholar] [CrossRef]
- Wang, B.-G.; Hong, X.; Li, L.; Zhou, J.; Hao, X.-J. Chemical constituents of two chinese magnoliaceae plants, Tsoongiodendron odorum and Manglietiastrum sinicum, and their inhibition of platelet aggregation. Planta Med. 2000, 66, 511–515. [Google Scholar] [CrossRef]
- Hatem, N.A.; Najah, Z.M. Isolation and elucidation of some chemical constituents of Lavandula officinalis. J. Chem. Pharm. Res. 2016, 8, 394–401. [Google Scholar]
- Tijjani, A.; Ndukwe, I.G.; Ayo, R.G. Isolation and characterization of lup-20(29)-Ene-3, 28- Diol (betulin) from the stem-bark of Adenium obesum (Apocynaceae). Trop. J. Pharm. Res. 2012, 11, 259–262. [Google Scholar] [CrossRef]
- Khanh, P.N.; Duc, H.V.; Huong, T.T.; Son, N.T.; Ha, V.T.; Van, D.T.; Tai, B.H.; Kim, J.E.; Jo, A.R.; Kim, Y.H.; et al. Alkylphloroglucinol derivatives and triterpenoids with soluble epoxide hydrolase inhibitory activity from Callistemon citrinus. Fitoterapia 2016, 109, 39–44. [Google Scholar] [CrossRef]
- Tawila, A.M.; Sun, S.; Kim, M.J.; Omar, A.M.; Dibwe, D.F.; Ueda, J.-Y.; Toyooka, N.; Awale, S. Highly potent antiausterity agents from Callistemon citrinus and their mechanism of action against the PANC-1 human pancreatic cancer cell line. J. Nat. Prod. 2020, 83, 2221–2232. [Google Scholar] [CrossRef]
- Tawila, A.M.; Omar, A.M.; Phan, N.D.; Takahashi, I.; Maneenet, J.; Awale, S. New Callistrilone epimers from Callistemon citrinus and their antiausterity activity against the PANC-1 human pancreatic cancer cell line. Tetrahedron Lett. 2022, 100, 153881. [Google Scholar] [CrossRef]
- Katsuta, E.; Qi, Q.; Peng, X.; Hochwald, S.N.; Yan, L.; Takabe, K. Pancreatic adenocarcinomas with mature blood vessels have better overall survival. Sci. Rep. 2019, 9, 1310. [Google Scholar] [CrossRef]
- Lippi, G.; Mattiuzzi, C. The global burden of pancreatic cancer. Arch. Med. Sci. 2020, 16, 820–824. [Google Scholar] [CrossRef]
- Sun, S.; Kim, M.J.; Omar, A.M.; Duy Phan, N.; Aoike, M.; Awale, S. GDP induces PANC-1 human pancreatic cancer cell death preferentially under nutrient starvation by inhibiting PI3K/Akt/MTOR/autophagy signaling pathway. Chem. Biodiver. 2021, 18, e2100389. [Google Scholar] [CrossRef]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Sig. Transduct. Target Ther. 2020, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alzahrani, A.S. PI3K/Akt/MTOR Inhibitors in cancer: At the bench and bedside. Semin. Cancer Biol. 2019, 59, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Li, X.; Zhang, J. mTOR signaling in cancer and mTOR inhibitors in solid tumor targeting therapy. Int. J. Mol. Sci. 2019, 20, 755. [Google Scholar] [CrossRef] [PubMed]
- Hoxhaj, G.; Manning, B.D. The PI3K-Akt network at the interface of oncogenic signalling and cancer metabolism. Nat. Rev. Cancer 2020, 20, 74–88. [Google Scholar] [CrossRef]
- Brown, J.R.; Byrd, J.C.; Coutre, S.E.; Benson, D.M.; Flinn, I.W.; Wagner-Johnston, N.D.; Spurgeon, S.E.; Kahl, B.S.; Bello, C.; Webb, H.K.; et al. Idelalisib, an inhibitor of phosphatidylinositol 3-kinase P110δ, for relapsed/refractory chronic lymphocytic leukemia. Blood 2014, 123, 3390–3397. [Google Scholar] [CrossRef]
- Scott, W.J.; Hentemann, M.F.; Rowley, R.B.; Bull, C.O.; Jenkins, S.; Bullion, A.M.; Johnson, J.; Redman, A.; Robbins, A.H.; Esler, W.; et al. Discovery and SAR of novel 2,3-dihydroimidazo [1,2-c]quinazoline PI3K inhibitors: Identification of copanlisib (BAY 80-6946). Chem. Med. Chem. 2016, 11, 1517–1530. [Google Scholar] [CrossRef]
- Rodrigues, D.A.; Sagrillo, F.S.; Fraga, C.A.M. Duvelisib: A 2018 novel FDA-approved small molecule inhibiting phosphoinositide 3-kinases. Pharmaceuticals 2019, 12, 69. [Google Scholar] [CrossRef]
- Murphy, A.G.; Zahurak, M.; Shah, M.; Weekes, C.D.; Hansen, A.; Siu, L.L.; Spreafico, A.; LoConte, N.; Anders, N.M.; Miles, T.; et al. A phase I study of dinaciclib in combination with MK-2206 in patients with advanced pancreatic cancer. Clin. Transl. Sci. 2020, 13, 1178–1188. [Google Scholar] [CrossRef]
- Piffoux, M.; Eriau, E.; Cassier, P.A. Autophagy as a therapeutic target in pancreatic cancer. Br. J. Cancer 2021, 124, 333–344. [Google Scholar] [CrossRef]
- Fang, D.; Xie, H.; Hu, T.; Shan, H.; Li, M. Binding features and functions of ATG3. Front. Cell Dev. Biol. 2021, 9, 685625. [Google Scholar] [CrossRef]
- Görgülü, K.; Diakopoulos, K.N.; Kaya-Aksoy, E.; Ciecielski, K.J.; Ai, J.; Lesina, M.; Algül, H. The role of autophagy in pancreatic cancer: From bench to the dark bedside. Cells 2020, 9, 1063. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, X.; Kang, R.; Zeh, H.; Klionsky, D.J.; Tang, D. Regulation and function of autophagy in pancreatic cancer. Autophagy 2021, 17, 3275–3296. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Subulatone A (1) | |
|---|---|---|
| δC | δH | |
| 1 | 199.0 a | - |
| 2 | 55.1 | - |
| 3 | 210.9 a | - |
| 4 | 51.7 | - |
| 5 | 97.5 | - |
| 6 | 143.1 | 7.13, s |
| 7 | 79.5 | - |
| 8 | 24.0 | 1.49, s |
| 9 | 24.2 | 1.35, s |
| 10 | 20.0 | 1.03, s |
| 11 | 15.3 | 1.31, s |
| 12 | 26.7 | 1.37, s |
| 13 | 23.8 | 1.37, s |
| Compound | PC50, µM | Compound | PC50, µM |
|---|---|---|---|
| 1 | 16.5 | 9 | 0.02 |
| 2 | 0.28 | 10 | 6.1 |
| 3 | 4.4 | 11 | 1.0 |
| 4 | 6.0 | 12 | 88.3 |
| 5 | 9.2 | 13 | 5.2 |
| 6 | 42.8 | 14 | 0.7 |
| 7 | 7.8 | 15 | 15.2 |
| 8 | 10.0 | Gemcitabine a | >100 |
| Arctigenin b | 0.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maneenet, J.; Tawila, A.M.; Omar, A.M.; Phan, N.D.; Ojima, C.; Kuroda, M.; Sato, M.; Mizoguchi, M.; Takahashi, I.; Awale, S. Chemical Constituents of Callistemon subulatus and Their Anti-Pancreatic Cancer Activity against Human PANC-1 Cell Line. Plants 2022, 11, 2466. https://doi.org/10.3390/plants11192466
Maneenet J, Tawila AM, Omar AM, Phan ND, Ojima C, Kuroda M, Sato M, Mizoguchi M, Takahashi I, Awale S. Chemical Constituents of Callistemon subulatus and Their Anti-Pancreatic Cancer Activity against Human PANC-1 Cell Line. Plants. 2022; 11(19):2466. https://doi.org/10.3390/plants11192466
Chicago/Turabian StyleManeenet, Juthamart, Ahmed M. Tawila, Ashraf M. Omar, Nguyen Duy Phan, Chiharu Ojima, Masahiro Kuroda, Mao Sato, Mio Mizoguchi, Ikue Takahashi, and Suresh Awale. 2022. "Chemical Constituents of Callistemon subulatus and Their Anti-Pancreatic Cancer Activity against Human PANC-1 Cell Line" Plants 11, no. 19: 2466. https://doi.org/10.3390/plants11192466
APA StyleManeenet, J., Tawila, A. M., Omar, A. M., Phan, N. D., Ojima, C., Kuroda, M., Sato, M., Mizoguchi, M., Takahashi, I., & Awale, S. (2022). Chemical Constituents of Callistemon subulatus and Their Anti-Pancreatic Cancer Activity against Human PANC-1 Cell Line. Plants, 11(19), 2466. https://doi.org/10.3390/plants11192466