
Validation of Reference Genes in a Population of Blueberry (Vaccinium corymbosum) Plants Regenerated in Colchicine

 ,
,
Abstract
:1. Introduction
2. Results
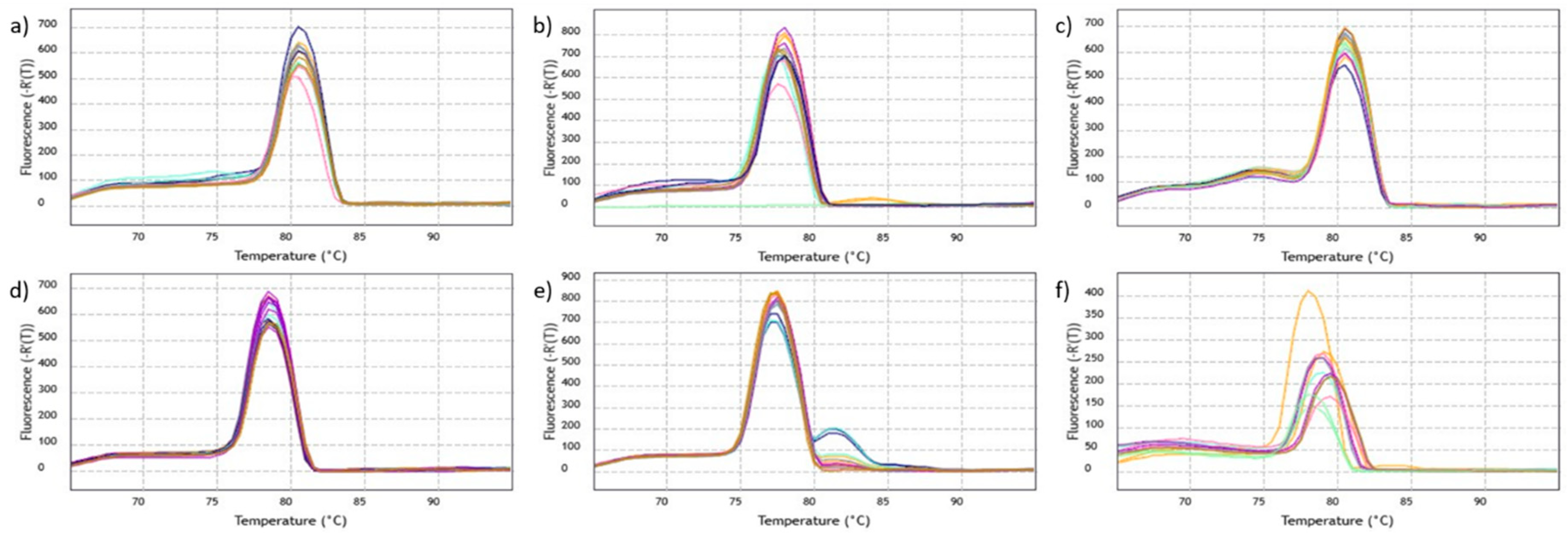
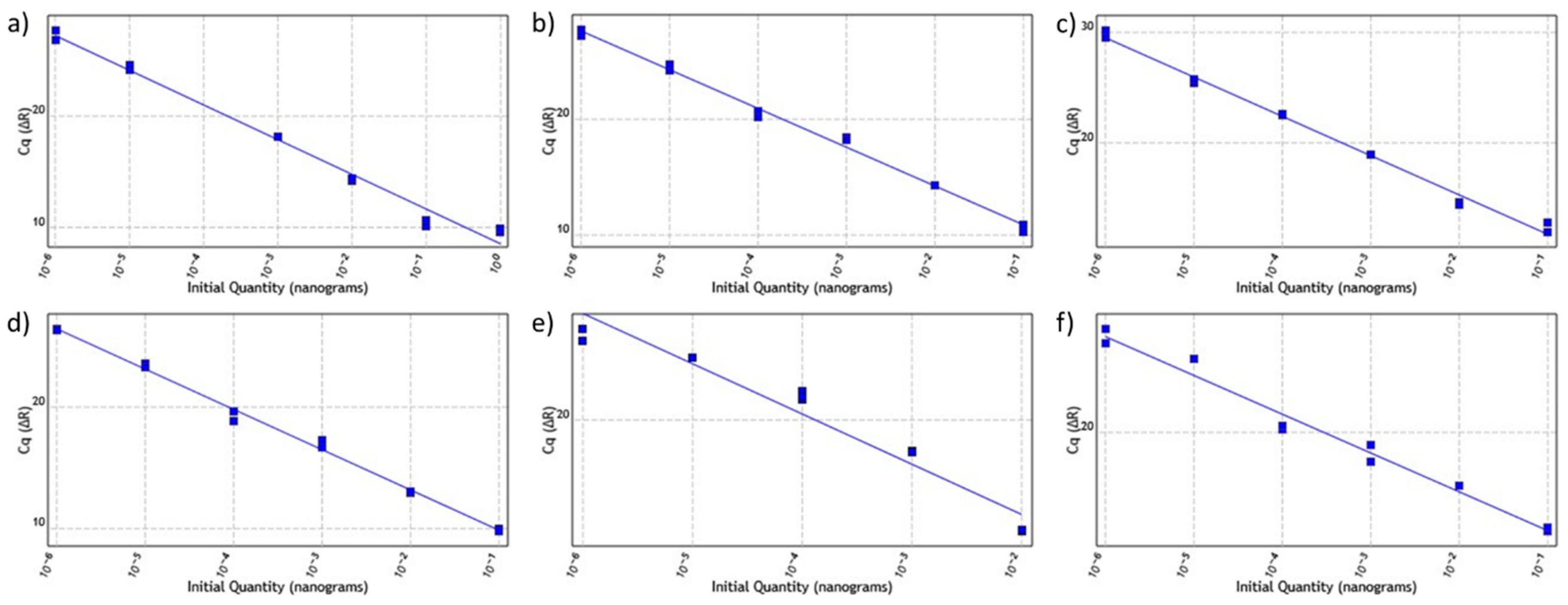
2.1. ATP1, COX2, GAPDH and MATK Gene Primers Showed Higher Efficiency in qPCR Assays
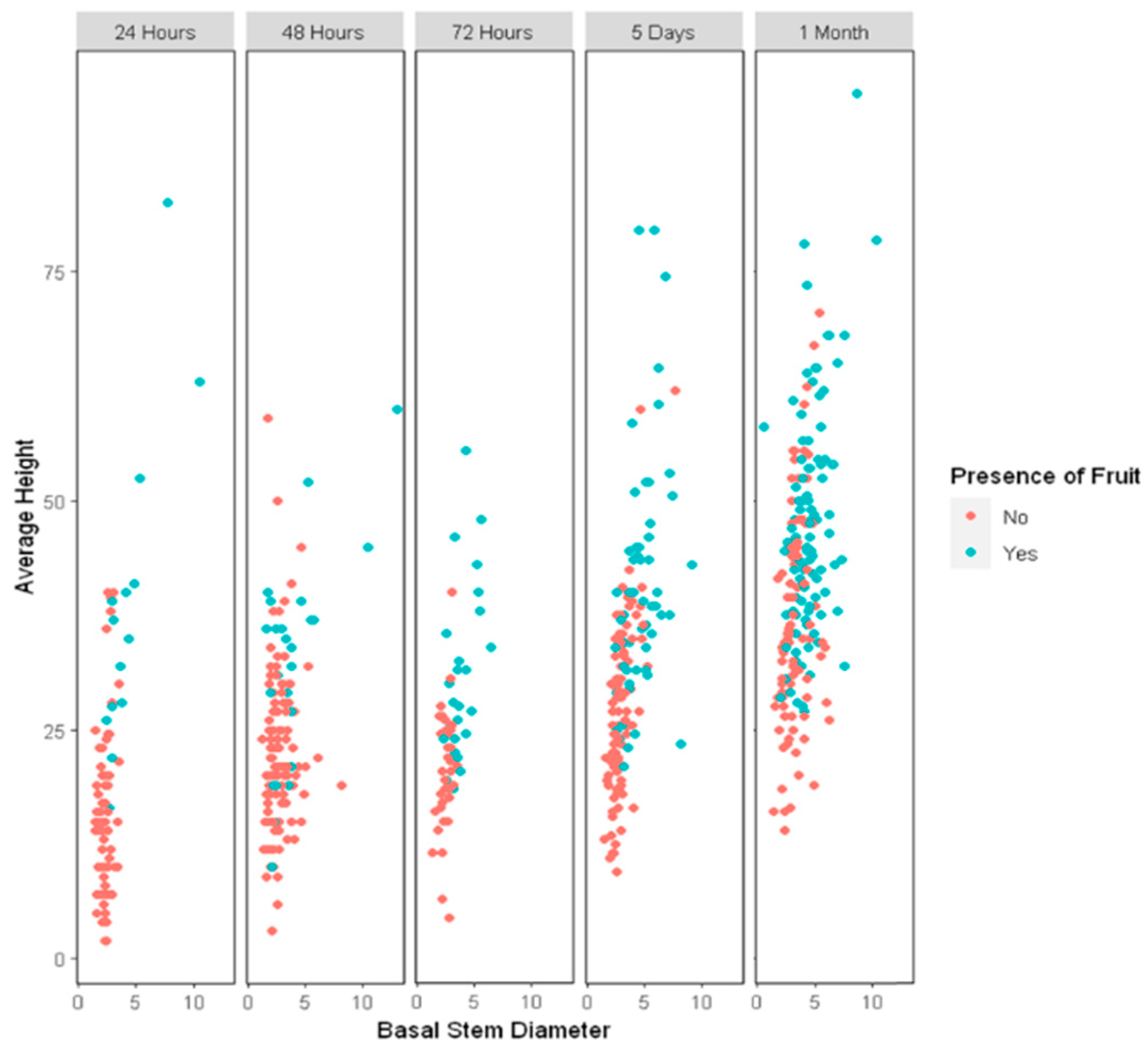
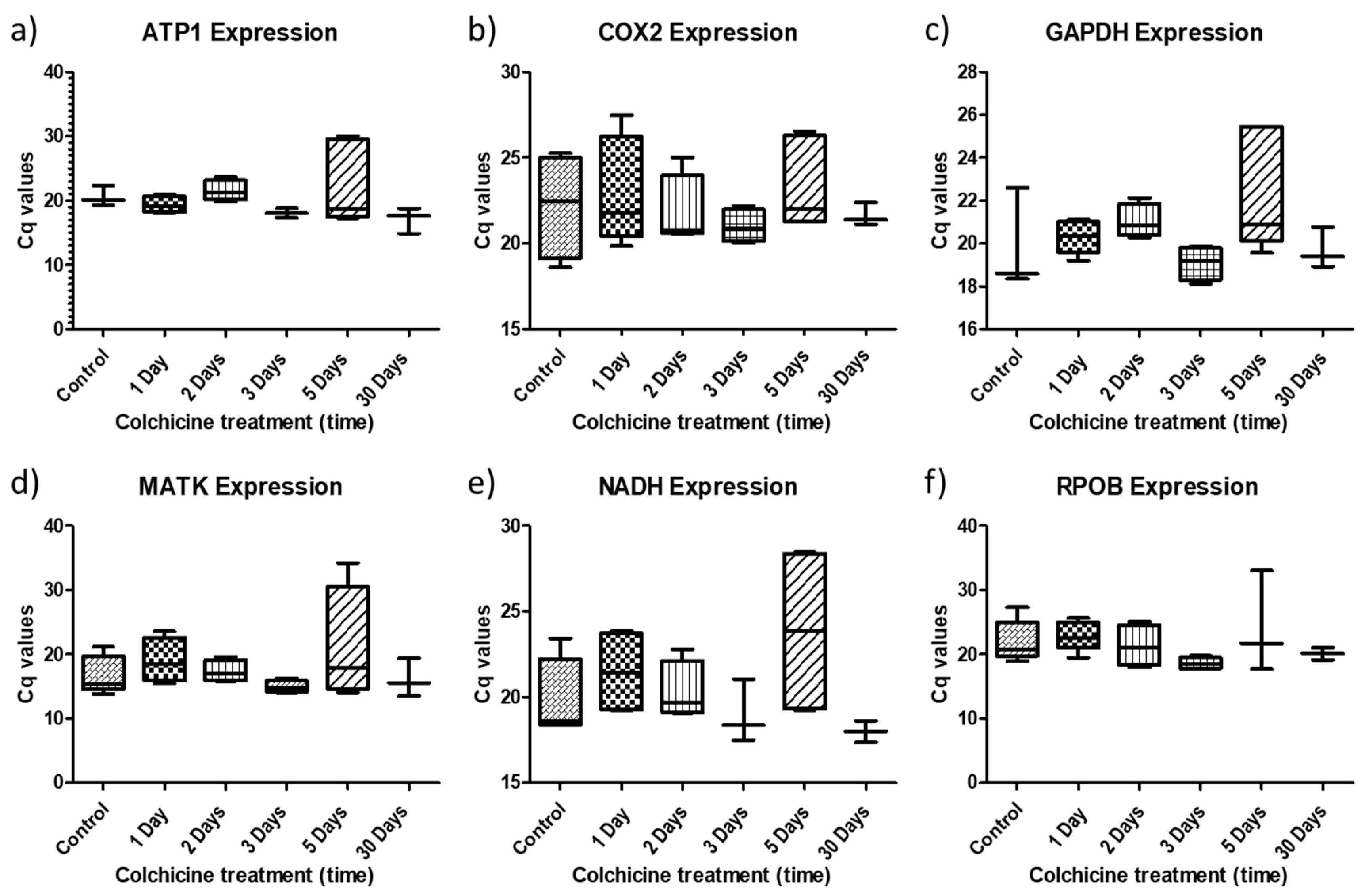
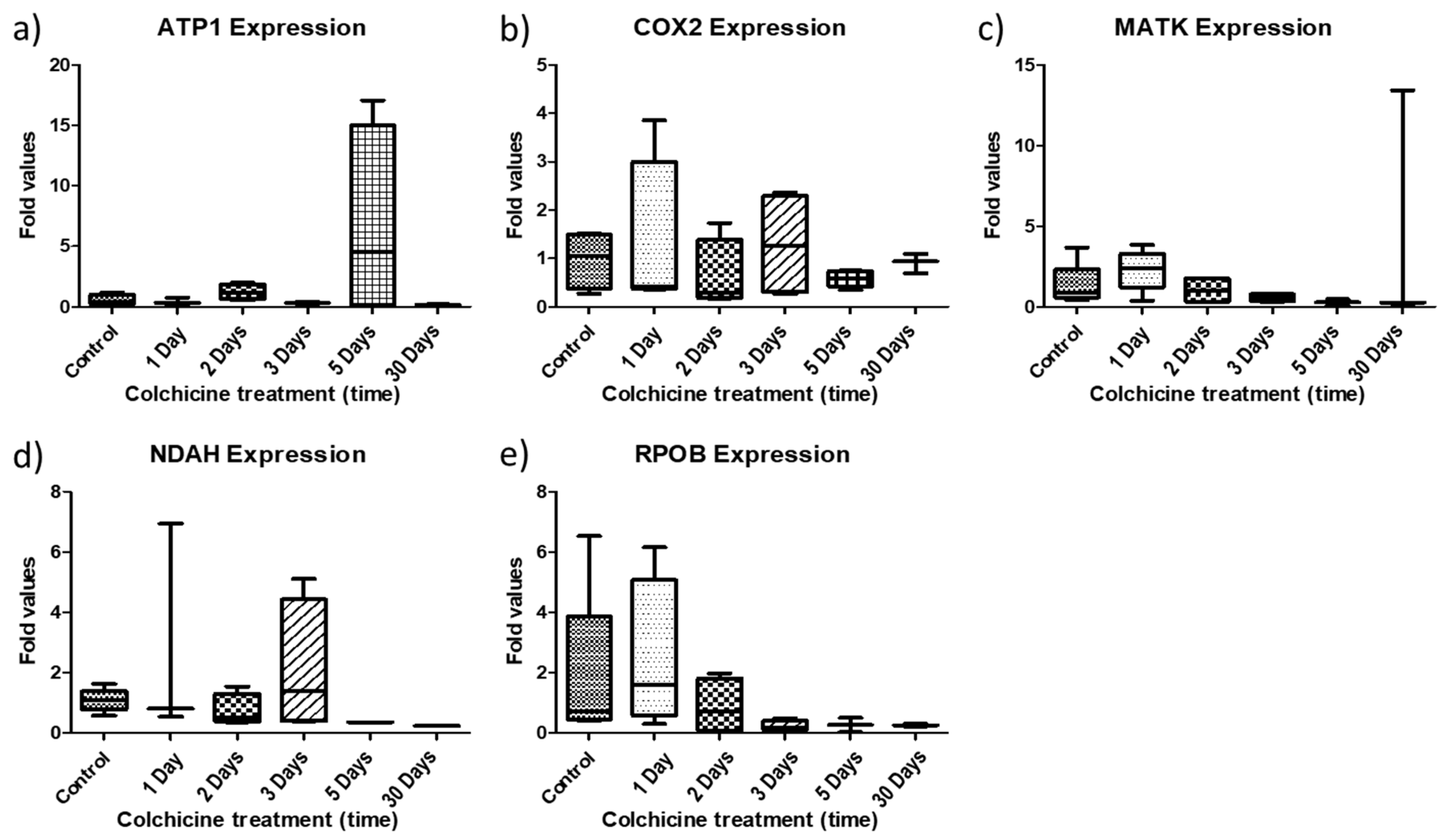
2.2. Gene Expression Behavior in Colchicine-Regenerated Blueberry Clones
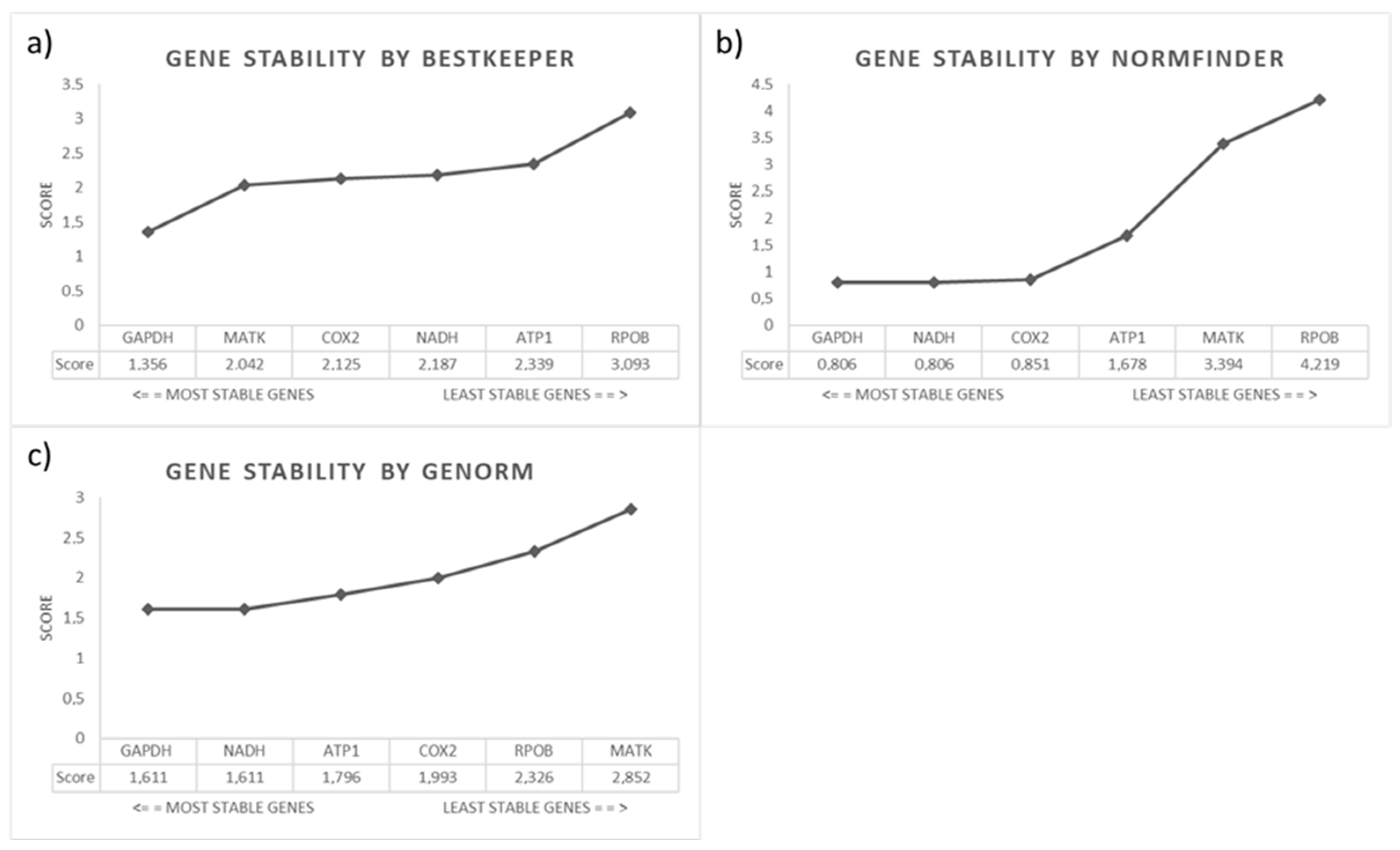
2.3. Are These Genes Stable for qPCR Assays?
3. Discussion
4. Materials and Methods
4.1. Vegetative Material
4.2. Colchicine Treatments and Acclimatization
4.3. Data Processing in the Field Trial
4.4. Sample Processing
4.5. cDNA Synthesis
4.6. Primer Design and cDNA Viability Testing
4.7. Analysis of Primer Activity and Efficiency
4.8. Stability Analysis for Use as Reference Genes
4.9. Statistical Analysis for Gene Expression Assays
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Patel, S. Blueberry as functional food and dietary supplement: The natural way to ensure holistic health. Mediterr. J. Nutr. Metab. 2014, 7, 133–143. [Google Scholar] [CrossRef]
- Ma, L.; Sun, Z.; Zeng, Y.; Luo, M.; Yang, J. Molecular Mechanism and Health Role of Functional Ingredients in Blueberry for Chronic Disease in Human Beings. Int. J. Mol. Sci. 2018, 19, 2785. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, A. Asoex. 8 September 2020. Available online: https://www.asoex.cl/component/content/article/25-noticias/812-temporada-2020-2021-industria-del-arandano-de-chile-proyecta-mantener-volumenes-de-exportacion-y-seguira-potenciando-la-calidad.html (accessed on 19 May 2022).
- Blueberry Breeding: Improving the Unwild Blueberry—ProQuest. Available online: https://www.proquest.com/openview/13fbbf3dd70ebcd17531389ccaba2788/1.pdf?pq-origsite=gscholar&cbl=46215 (accessed on 19 May 2022).
- Caligari, P.; Retamales, J.; Lobos, G. Blueberry breeding for chilean conditions. Acta Hortic. 2009, 810, 125–128. [Google Scholar] [CrossRef]
- Tavan, M.; Mirjalili, M.H.; Karimzadeh, G. In vitro polyploidy induction: Changes in morphological, anatomical and phytochemical characteristics of Thymus persicus (Lamiaceae). Plant Cell Tissue Organ Cult. 2015, 122, 573–583. [Google Scholar] [CrossRef]
- Madani, H.; Escrich, A.; Hosseini, B.; Sanchez-Muñoz, R.; Khojasteh, A.; Palazon, J. Effect of Polyploidy Induction on Natural Metabolite Production in Medicinal Plants. Biomolecules 2021, 11, 899. [Google Scholar] [CrossRef] [PubMed]
- Salman-Minkov, A.; Sabath, N.; Mayrose, I. Whole-genome duplication as a key factor in crop domestication. Nat. Plants 2016, 2, 16115. [Google Scholar] [CrossRef] [PubMed]
- Mo, L.; Chen, J.; Lou, X.; Xu, Q.; Dong, R.; Tong, Z.; Huang, H.; Lin, E. Colchicine-Induced Polyploidy in Rhododendron fortunei Lindl. Plants 2020, 9, 424. [Google Scholar] [CrossRef] [Green Version]
- Fox, D.T.; Soltis, D.E.; Soltis, P.S.; Ashman, T.-L.; Van de Peer, Y. Polyploidy: A Biological Force From Cells to Ecosystems. Trends Cell Biol. 2020, 30, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Jing, Y.; Kang, X. In vitro induction of tetraploid and resulting trait variation in Populus alba × Populus glandulosa clone 84 K. Plant Cell Tissue Organ Cult. 2021, 146, 285–296. [Google Scholar] [CrossRef]
- Jackson, S.; Chen, Z.J. Genomic and expression plasticity of polyploidy. Curr. Opin. Plant Biol. 2010, 13, 153–159. [Google Scholar] [CrossRef]
- Manzoor, A.; Ahmad, T.; Bashir, M.A.; Hafiz, I.A.; Silvestri, C. Studies on Colchicine Induced Chromosome Doubling for Enhancement of Quality Traits in Ornamental Plants. Plants 2019, 8, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arencibia, A.D.; D’Afonseca, V.; Chakravarthi, M.; Castiglione, S. Learning from transgenics: Advanced gene editing technologies should also bridge the gap with traditional genetic selection. Electron. J. Biotechnol. 2019, 41, 22–29. [Google Scholar] [CrossRef]
- Wang, Z.; Fan, G.; Dong, Y.; Zhai, X.; Deng, M.; Zhao, Z.; Liu, W.; Cao, Y. Implications of polyploidy events on the phenotype, microstructure, and proteome of Paulownia australis. PLoS ONE 2017, 12, e0172633. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Zheng, Y.; Song, S.; Huo, B.; Li, D.; Wang, J. In vitro induction of allohexaploid and resulting phenotypic variation in Populus. Plant Cell Tissue Organ Cult. 2018, 134, 183–192. [Google Scholar] [CrossRef]
- Cao, Z.; Deng, Z. Morphological, cytological and molecular marker analyses of ‘Tapestry’ caladium variants reveal diverse genetic changes and enable association of leaf coloration pattern loci with molecular markers. Plant Cell Tissue Organ Cult. 2020, 143, 363–375. [Google Scholar] [CrossRef]
- Maroufi, A.; van Bockstaele, E.; de Loose, M. Validation of reference genes for gene expression analysis in chicory (Cichorium intybus) using quantitative real-time PCR. BMC Mol. Biol. 2010, 11, 15. [Google Scholar] [CrossRef] [Green Version]
- Wan, H.; Zhao, Z.; Qian, C.; Sui, Y.; Malik, A.A.; Chen, J. Selection of appropriate reference genes for gene expression studies by quantitative real-time polymerase chain reaction in cucumber. Anal. Biochem. 2010, 399, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, J.C.; Lopez-Cobollo, R.; Alcazar, R.; Zarza, X.; Koncz, C.; Altabella, T.; Salinas, J.; Tiburcio, A.F.; Ferrando, A. Putrescine Is Involved in Arabidopsis Freezing Tolerance and Cold Acclimation by Regulating Abscisic Acid Levels in Response to Low Temperature. Plant Physiol. 2008, 148, 1094–1105. [Google Scholar] [CrossRef] [Green Version]
- Cruz, F.; Kalaoun, S.; Nobile, P.; Colombo, C.; Almeida, J.; Barros, L.M.G.; Romano, E.; Grossi-De-Sá, M.F.; Vaslin, M.; Alves-Ferreira, M. Evaluation of coffee reference genes for relative expression studies by quantitative real-time RT-PCR. Mol. Breed. 2009, 23, 607–616. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, L.; Mauriat, M.; Pelloux, J.; Bellini, C.; Van Wuytswinkel, O. Towards a Systematic Validation of References in Real-Time RT-PCR. Plant Cell 2008, 20, 1734–1735. [Google Scholar] [CrossRef]
- Nicot, N.; Hausman, J.-F.; Hoffmann, L.; Evers, D. Housekeeping gene selection for real-time RT-PCR normalization in potato during biotic and abiotic stress. J. Exp. Bot. 2005, 56, 2907–2914. [Google Scholar] [CrossRef]
- González-Agüero, M.; García-Rojas, M.; Di Genova, A.; Correa, J.; Maass, A.; Orellana, A.; Hinrichsen, P. Identification of two putative reference genes from grapevine suitable for gene expression analysis in berry and related tissues derived from RNA-Seq data. BMC Genom. 2013, 14, 878. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Gao, Z.; Li, H.; Li, Q.; Zhang, C.; Xu, W.; Song, S.; Ma, C.; Wang, S. Selection of reference genes for miRNA qRT-PCR under abiotic stress in grapevine. Sci. Rep. 2018, 8, 4444. [Google Scholar] [CrossRef]
- Zhu, X.; Li, X.; Chen, W.; Chen, J.; Lu, W.; Chen, L.; Fu, D. Evaluation of New Reference Genes in Papaya for Accurate Transcript Normalization under Different Experimental Conditions. PLoS ONE 2012, 7, e44405. [Google Scholar] [CrossRef]
- Libault, M.; Thibivilliers, S.; Bilgin, D.D.; Radwan, O.; Benitez, M.; Clough, S.J.; Stacey, G. Identification of Four Soybean Reference Genes for Gene Expression Normalization. Plant Genome 2008, 1, 44–54. [Google Scholar] [CrossRef]
- Vashisth, T.; Johnson, L.K.; Malladi, A. An efficient RNA isolation procedure and identification of reference genes for normalization of gene expression in blueberry. Plant Cell Rep. 2011, 30, 2167–2176. [Google Scholar] [CrossRef]
- Die, J.V.; Rowland, L.J. Superior Cross-Species Reference Genes: A Blueberry Case Study. PLoS ONE 2013, 8, e73354. [Google Scholar] [CrossRef]
- Deng, Y.; Li, Y.; Sun, H. Selection of reference genes for RT-qPCR normalization in blueberry (Vaccinium corymbosum × angustifolium) under various abiotic stresses. FEBS Open Bio 2020, 10, 1418–1435. [Google Scholar] [CrossRef]
- Jose, S.; Abbey, J.; Jaakola, L.; Percival, D. Selection and validation of reliable reference genes for gene expression studies from Monilinia vaccinii-corymbosi infected wild blueberry phenotypes. Sci. Rep. 2020, 10, 11688. [Google Scholar] [CrossRef]
- Qiao, G.; Liu, M.; Song, K.; Li, H.; Yang, H.; Yin, Y.; Zhuo, R. Phenotypic and Comparative Transcriptome Analysis of Different Ploidy Plants in Dendrocalamus latiflorus Munro. Front. Plant Sci. 2017, 8, 1371. [Google Scholar] [CrossRef]
- Eng, W.H.; Ho, W.S.; Ling, K.H. In vitro induction and identification of polyploid Neolamarckia cadamba plants by colchicine treatment. PeerJ 2021, 9, e12399. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, K.; Aziz, U.; Zhao, C.; Zhang, M. Evaluation of duplicated reference genes for quantitative real-time PCR analysis in genome unknown hexaploid oat (Avena sativa L.). Plant Methods 2020, 16, 138. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Cao, K.; Deng, C.; Li, Y.; Zhu, G.; Fang, W.; Chen, C.; Wang, X.; Wu, J.; Guan, L.; et al. An integrated peach genome structural variation map uncovers genes associated with fruit traits. Genome Biol. 2020, 21, 258. [Google Scholar] [CrossRef]
- Dorak, M.T. Real-Time PCR; Taylor & Francis: London, UK, 2007; pp. 1–333. [Google Scholar] [CrossRef]
- Lopes, J.M.L.; de Matos, E.M.; de Queiroz Nascimento, L.S.; Viccini, L.F. Validation of reference genes for quantitative gene expression in the Lippia alba polyploid complex (Verbenaceae). Mol. Biol. Rep. 2021, 48, 1037–1044. [Google Scholar] [CrossRef]
- Wang, H.; Chen, S.; Jiang, J.; Zhang, F.; Chen, F. Reference gene selection for cross-species and cross-ploidy level comparisons in Chrysanthemum spp. Sci. Rep. 2015, 5, 8094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, R.; Jouve, N.; De Bustos, A. Comparative analysis of gene expression among species of different ploidy. Mol. Biol. Rep. 2014, 41, 6525–6535. [Google Scholar] [CrossRef] [PubMed]
- Giménez, M.J.; Pistón, F.; Atienza, S.G. Identification of suitable reference genes for normalization of qPCR data in comparative transcriptomics analyses in the Triticeae. Planta 2010, 233, 163–173. [Google Scholar] [CrossRef]
- Wang, H.; Wang, J.; Jiang, J.; Chen, S.; Guan, Z.; Liao, Y.; Chen, F. Reference genes for normalizing transcription in diploid and tetraploid Arabidopsis. Sci. Rep. 2014, 4, 6781. [Google Scholar] [CrossRef] [Green Version]
- Preuten, T.; Cincu, E.; Fuchs, J.; Zoschke, R.; Liere, K.; Börner, T. Fewer genes than organelles: Extremely low and variable gene copy numbers in mitochondria of somatic plant cells. Plant J. 2010, 64, 948–959. [Google Scholar] [CrossRef]
- Regalado, J.J.; Carmona-Martín, E.; Querol, V.; Veléz, C.G.; Encina, C.L.; Pitta-Alvarez, S.I. Production of compact petunias through polyploidization. Plant Cell Tissue Organ Cult. 2017, 129, 61–71. [Google Scholar] [CrossRef]
- Chen, X.; Ge, X.; Wang, J.; Tan, C.; King, G.J.; Liu, K. Genome-wide DNA methylation profiling by modified reduced representation bisulfite sequencing in Brassica rapa suggests that epigenetic modifications play a key role in polyploid genome evolution. Front. Plant Sci. 2015, 6, 836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jonge, H.J.M.; Fehrmann, R.; De Bont, E.S.J.M.; Hofstra, R.; Gerbens, F.; Kamps, W.A.; de Vries, E.; Van Der Zee, A.G.J.; Meerman, G.J.T.; Ter Elst, A. Evidence Based Selection of Housekeeping Genes. PLoS ONE 2007, 2, e898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St-Pierre, J.; Grégoire, J.-C.; Vaillancourt, C. A simple method to assess group difference in RT-qPCR reference gene selection using GeNorm: The case of the placental sex. Sci. Rep. 2017, 7, 16923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, L.; Mauriat, M.; Guénin, S.; Pelloux, J.; Lefebvre, J.-F.; Louvet, R.; Rusterucci, C.; Moritz, T.; Guerineau, F.; Bellini, C.; et al. The lack of a systematic validation of reference genes: A serious pitfall undervalued in reverse transcription-polymerase chain reaction (RT-PCR) analysis in plants. Plant Biotechnol. J. 2008, 6, 609–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, M.; Nijhawan, A.; Tyagi, A.K.; Khurana, J.P. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2006, 345, 646–651. [Google Scholar] [CrossRef]
- Expósito-Rodríguez, M.; Borges, A.A.; Borges-Pérez, A.; Pérez, J.A. Selection of internal control genes for quantita-tive real-time RT-PCR studies during tomato development process. BMC Plant Biol. 2008, 8, 131. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Janská, A.; Hodek, J.; Svoboda, P.; Zámečník, J.; Prášil, I.T.; Vlasáková, E.; Milella, L.; Ovesná, J. The choice of reference gene set for assessing gene expression in barley (Hordeum vulgare L.) under low temperature and drought stress. Mol. Genet. Genom. 2013, 288, 639–649. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Efficiency | Slope | R2 |
|---|---|---|---|
| ATP1 | 109.36 | −3.11 | 0.985 |
| COX2 | 99.02 | −3.34 | 0.994 |
| GAPDH | 91.07 | −3.55 | 0.993 |
| MATK | 100.08 | −3.32 | 0.995 |
| NADH | 113.47 | −3.036 | 0.954 |
| RPOB | 109.74 | −3.10 | 0.980 |
| Gene | Sequence | Protein ID | Organism | Start | End | Primer Size (bp) | CG content (%) | Tm | Product Size |
|---|---|---|---|---|---|---|---|---|---|
| GAPDH F | AGGTTTGGCATCGTTGAGGG | AAM96896.1 | Vaccinium myrtillus | 134 | 153 | 20 | 55 | 60.61 | 174 |
| GAPDH R | AAGGGCAGGCAACACCTTAC | AAM96896.1 | Vaccinium myrtillus | 307 | 288 | 20 | 55 | 60.54 | 174 |
| RPOB F | GTTCACTCGGTTTAGCGGGG | YP_007025937.1 | Vaccinium macrocarpon | 2.6 | 2619 | 20 | 60 | 61.02 | 199 |
| RPOB R | GGATCGCCGGTTCTTCCATC | YP_007025937.1 | Vaccinium macrocarpon | 2798 | 2779 | 20 | 60 | 60.88 | 199 |
| MATK F | CCCTCGACTTTCTGGGCTATC | YP_007025894.1 | Vaccinium macrocarpon | 983 | 1003 | 20 | 57 | 59.93 | 195 |
| MATK R | CCGGCTTACTCATGGGATGT | YP_007025894.1 | Vaccinium macrocarpon | 1177 | 1158 | 20 | 55 | 59.53 | 195 |
| ATP1 F | GACCGCCTTACCCGTCATTG | YP_008999606.1 | Vaccinium macrocarpon | 972 | 991 | 20 | 60 | 60.09 | 181 |
| ATP1 R | TTTCAACTGAGCGGCAGACC | YP_008999606.1 | Vaccinium macrocarpon | 1152 | 1133 | 20 | 55 | 60.88 | 181 |
| COX2 F | GGGTCTTGGTTCGCGCTTTA | YP_008999605.1 | Vaccinium macrocarpon | 158 | 177 | 20 | 55 | 60.95 | 181 |
| COX2 R | GCTGGATCGACTACTACCTCGT | YP_008999605.1 | Vaccinium macrocarpon | 338 | 317 | 20 | 54 | 61.06 | 181 |
| NADH F | TTATTTGGGACCCGCGAGGA | YP_007025953.1 | Vaccinium macrocarpon | 1563 | 1582 | 20 | 55 | 61.27 | 170 |
| NADH R | CGCCACAAATTCCATCCCCA | YP_007025953.1 | Vaccinium macrocarpon | 1732 | 1713 | 20 | 55 | 60.97 | 170 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valenzuela, F.; D’Afonseca, V.; Hernández, R.; Gómez, A.; Arencibia, A.D. Validation of Reference Genes in a Population of Blueberry (Vaccinium corymbosum) Plants Regenerated in Colchicine. Plants 2022, 11, 2645. https://doi.org/10.3390/plants11192645
Valenzuela F, D’Afonseca V, Hernández R, Gómez A, Arencibia AD. Validation of Reference Genes in a Population of Blueberry (Vaccinium corymbosum) Plants Regenerated in Colchicine. Plants. 2022; 11(19):2645. https://doi.org/10.3390/plants11192645
Chicago/Turabian StyleValenzuela, Francisca, Vivían D’Afonseca, Ricardo Hernández, Aleydis Gómez, and Ariel D. Arencibia. 2022. "Validation of Reference Genes in a Population of Blueberry (Vaccinium corymbosum) Plants Regenerated in Colchicine" Plants 11, no. 19: 2645. https://doi.org/10.3390/plants11192645
APA StyleValenzuela, F., D’Afonseca, V., Hernández, R., Gómez, A., & Arencibia, A. D. (2022). Validation of Reference Genes in a Population of Blueberry (Vaccinium corymbosum) Plants Regenerated in Colchicine. Plants, 11(19), 2645. https://doi.org/10.3390/plants11192645