
Genome-Wide Identification and Expression Analysis of HSF Transcription Factors in Alfalfa (Medicago sativa) under Abiotic Stress

Abstract
:1. Introduction
2. Results
2.1. Identification of HSF Genes in M. sativa
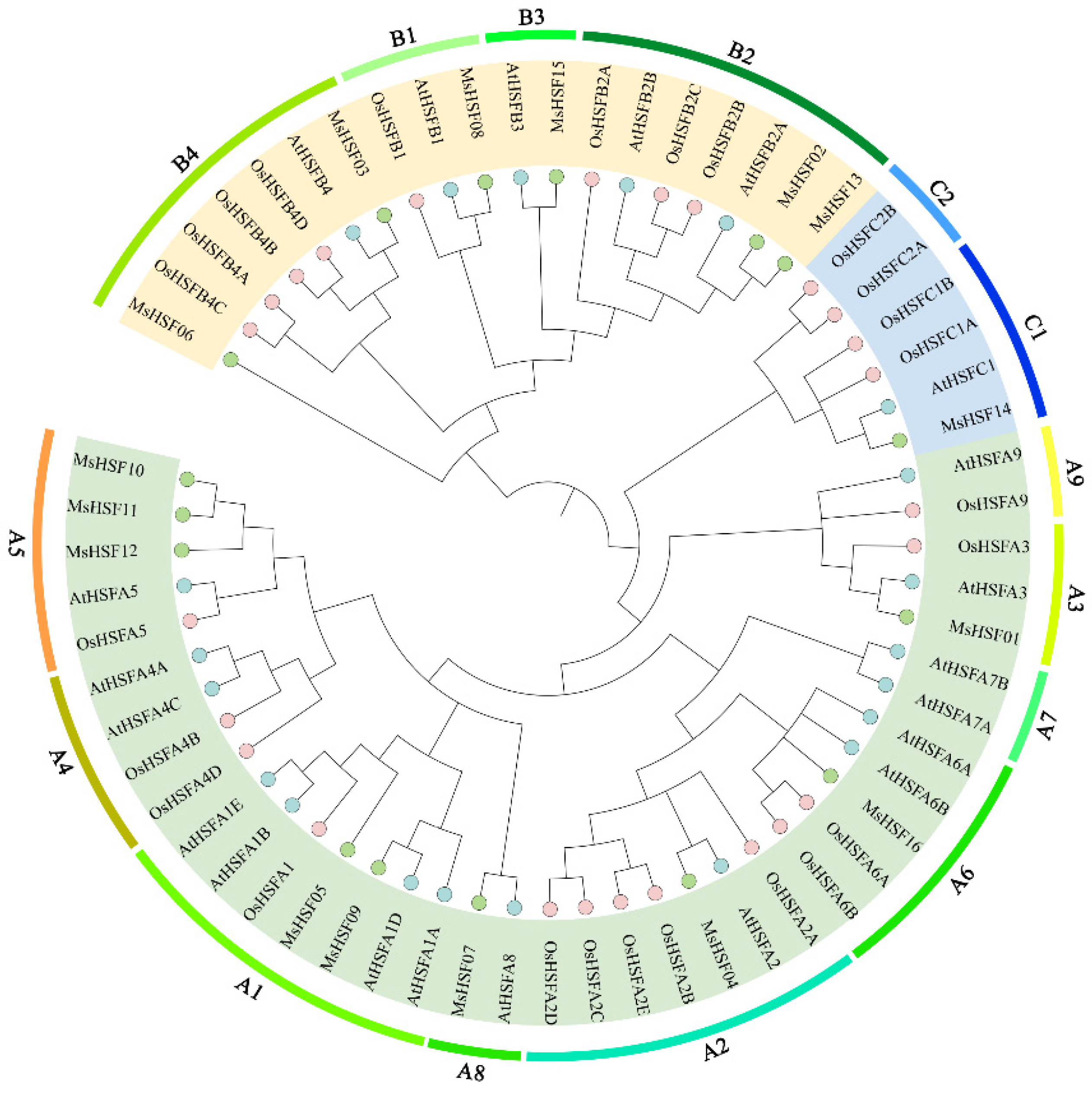
2.2. Phylogenetic Analysis of HSF Genes in M. sativa
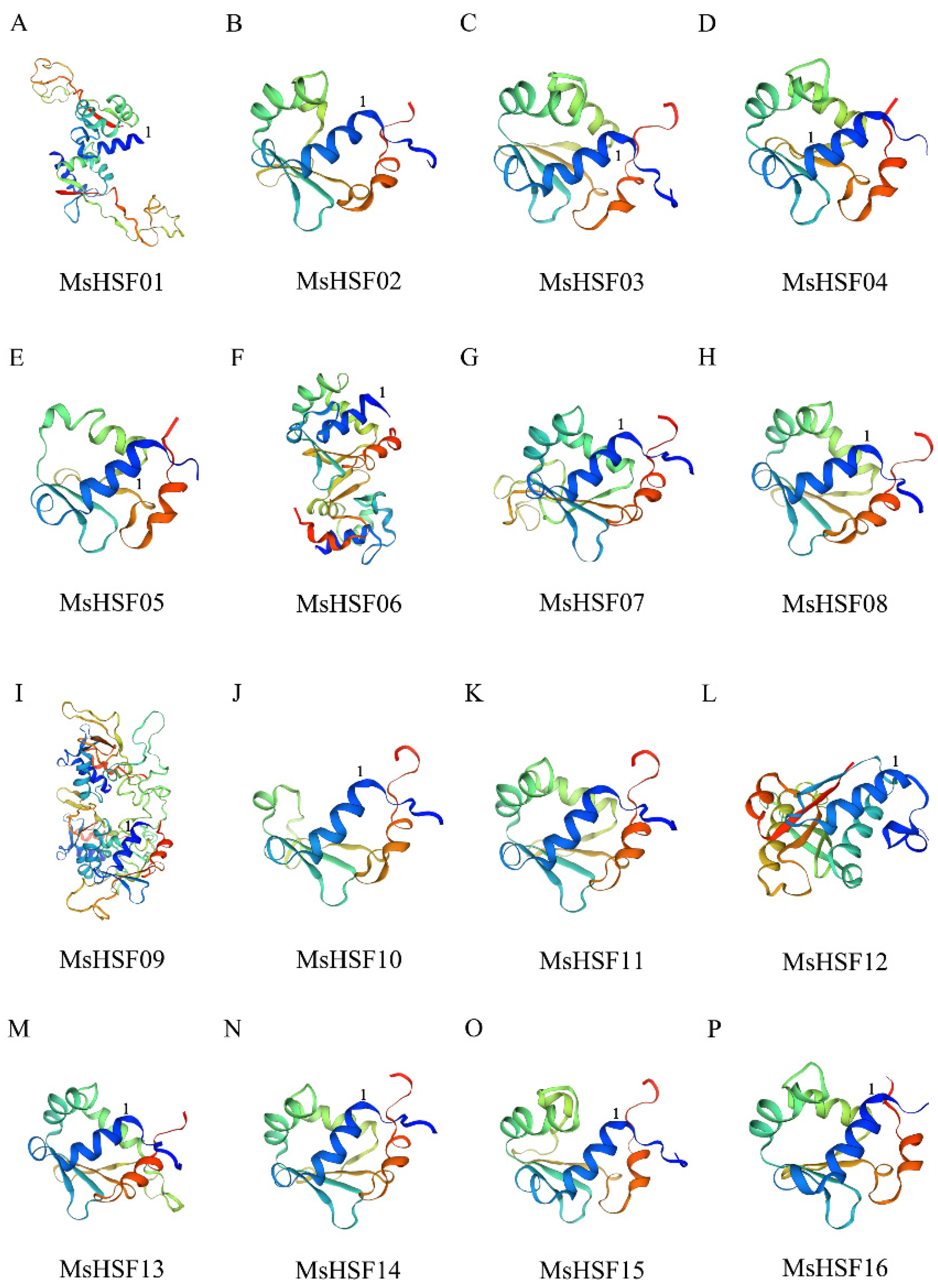
2.3. Multiple Sequence Alignment and Protein Modeling Analysis of the HSF Gene in M. sativa
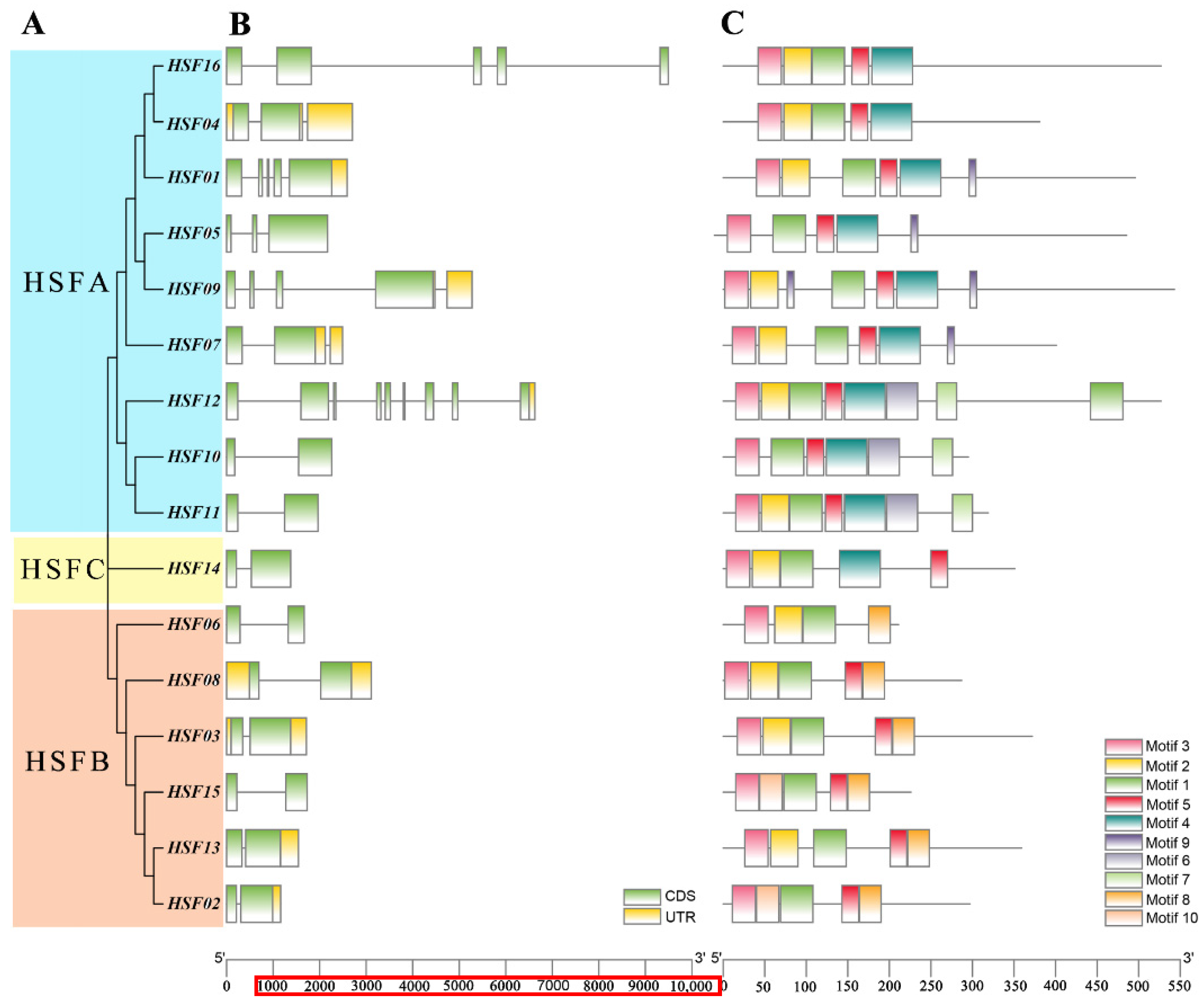
2.4. Gene Structure and Conserved Motif Analysis of the HSF Gene in M. sativa
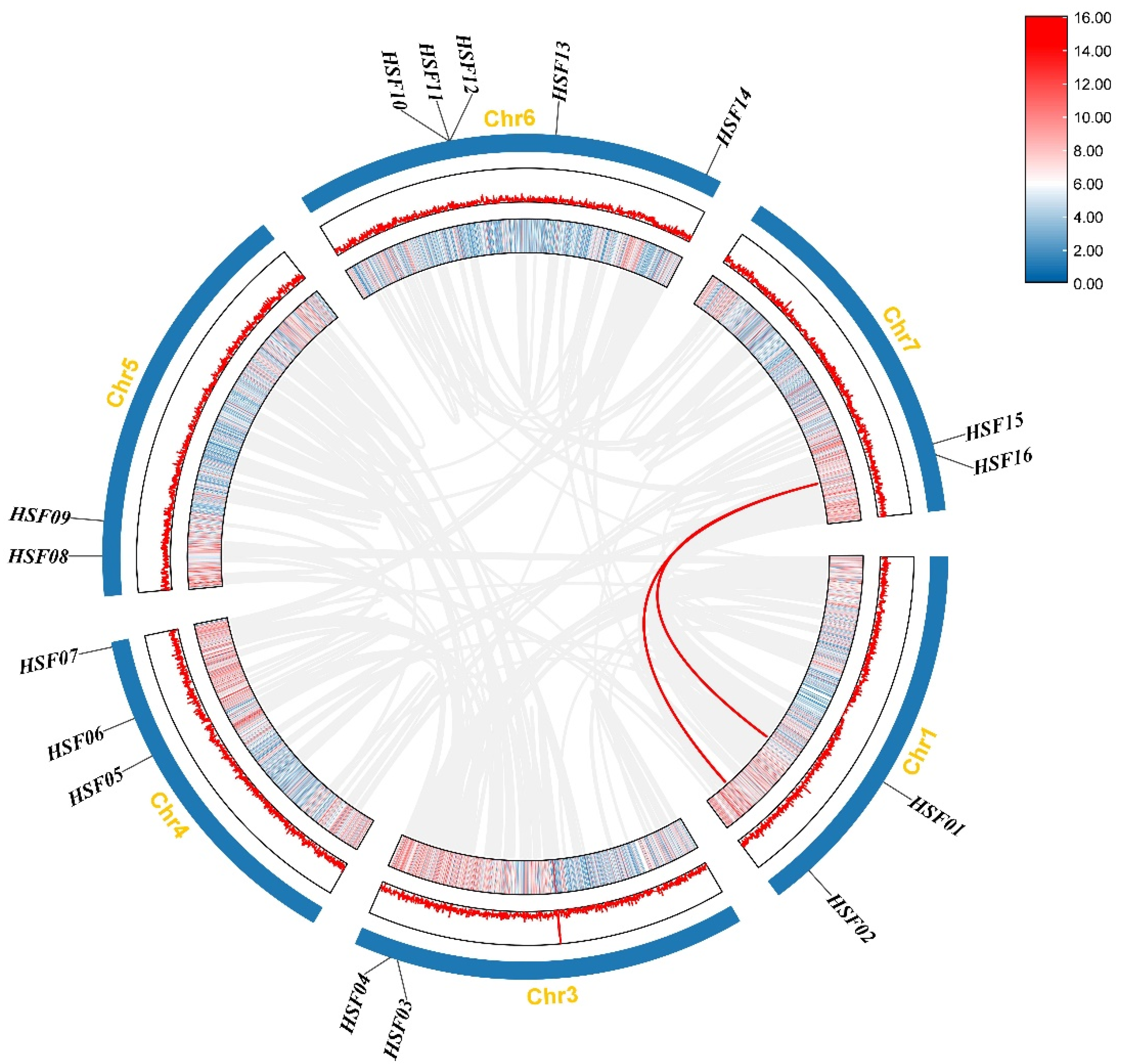
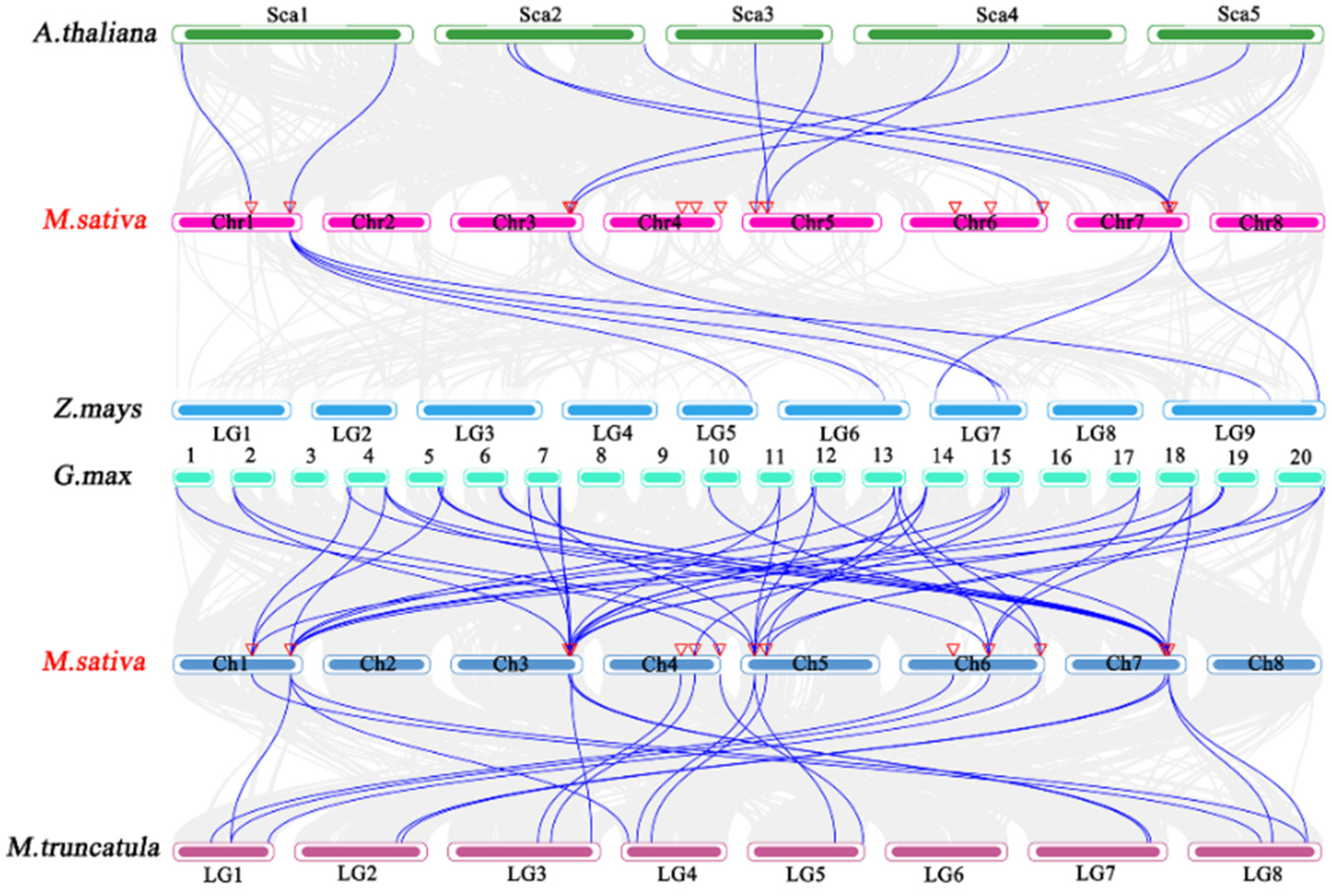
2.5. Chromosome Distribution and Covariance Analysis of HSF Genes in M. sativa
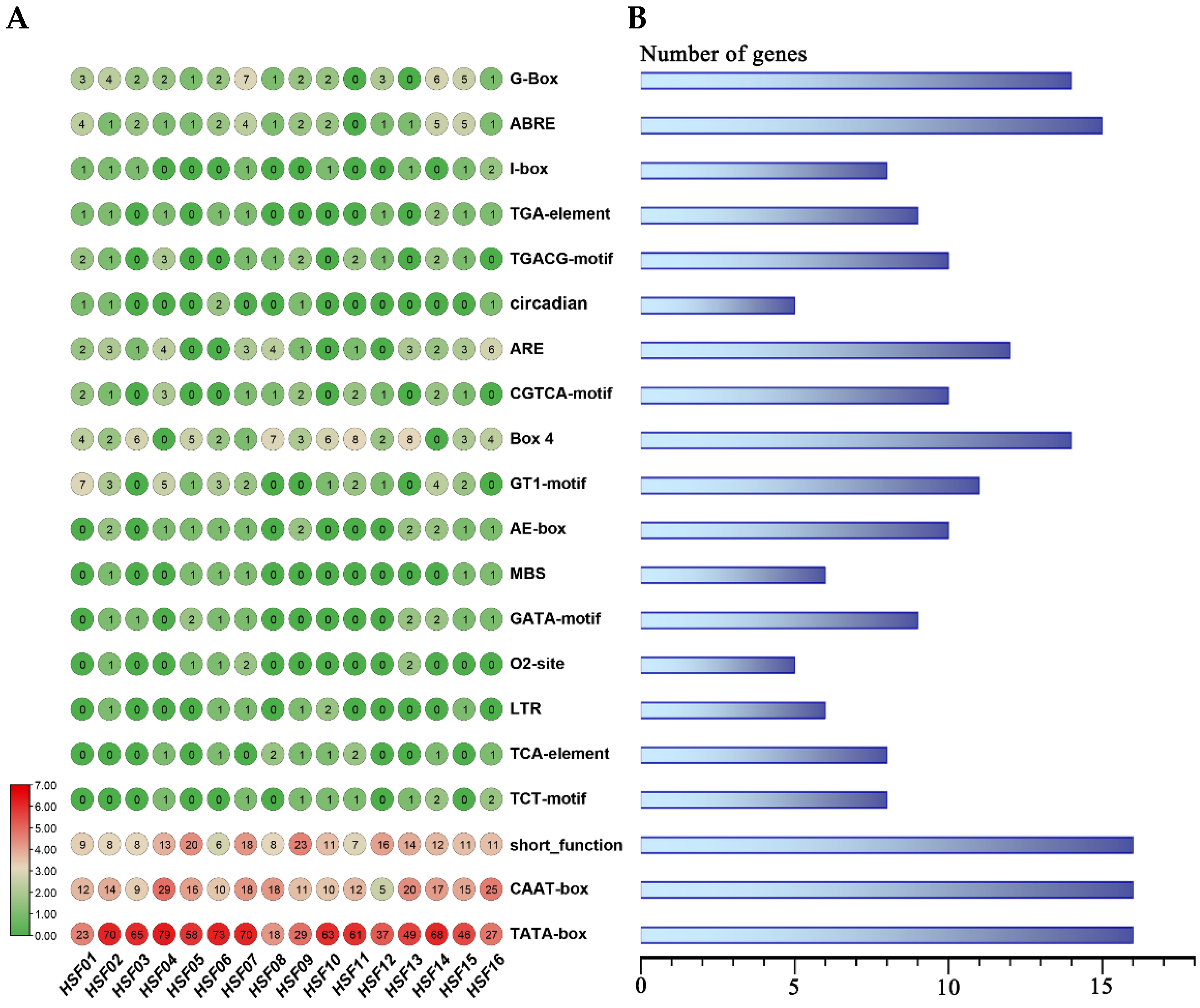
2.6. Analysis of the Promoter Cis-Element of the HSF Gene in M. sativa
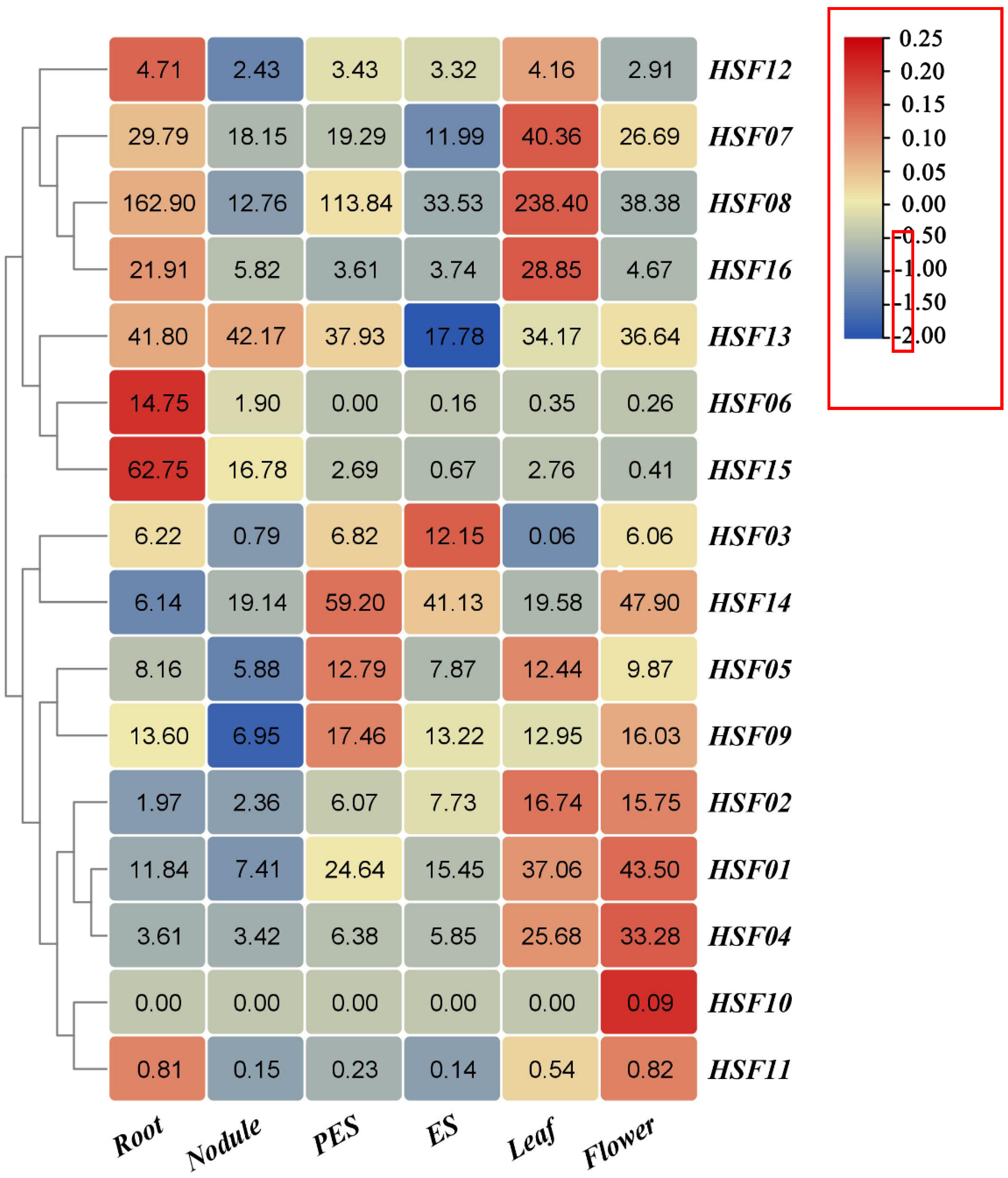
2.7. Expression Profiling of the M. sativa HSF Gene in Different Tissues
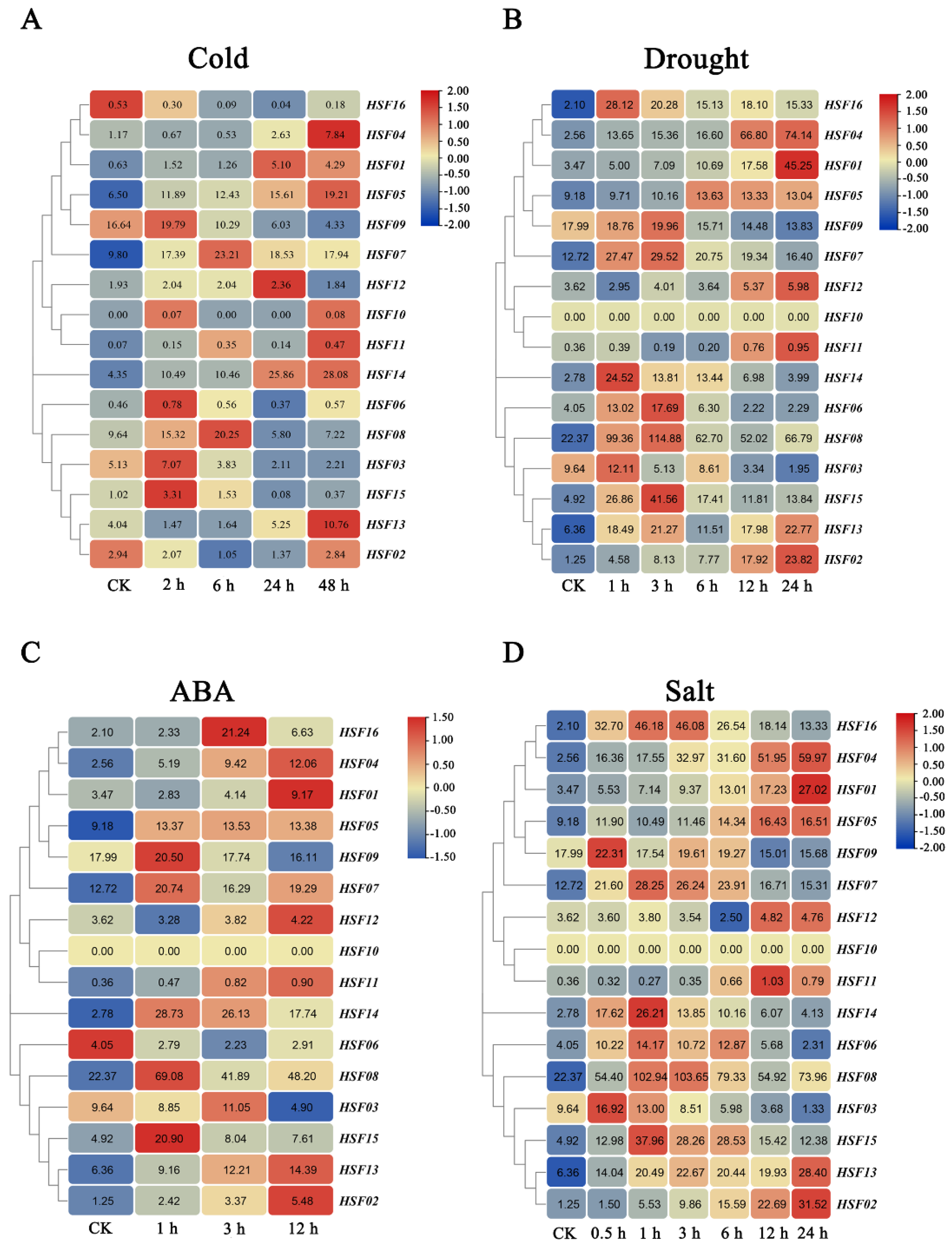
2.8. Expression Analysis of HSF Genes in M. sativa in Response to Abiotic Stresses
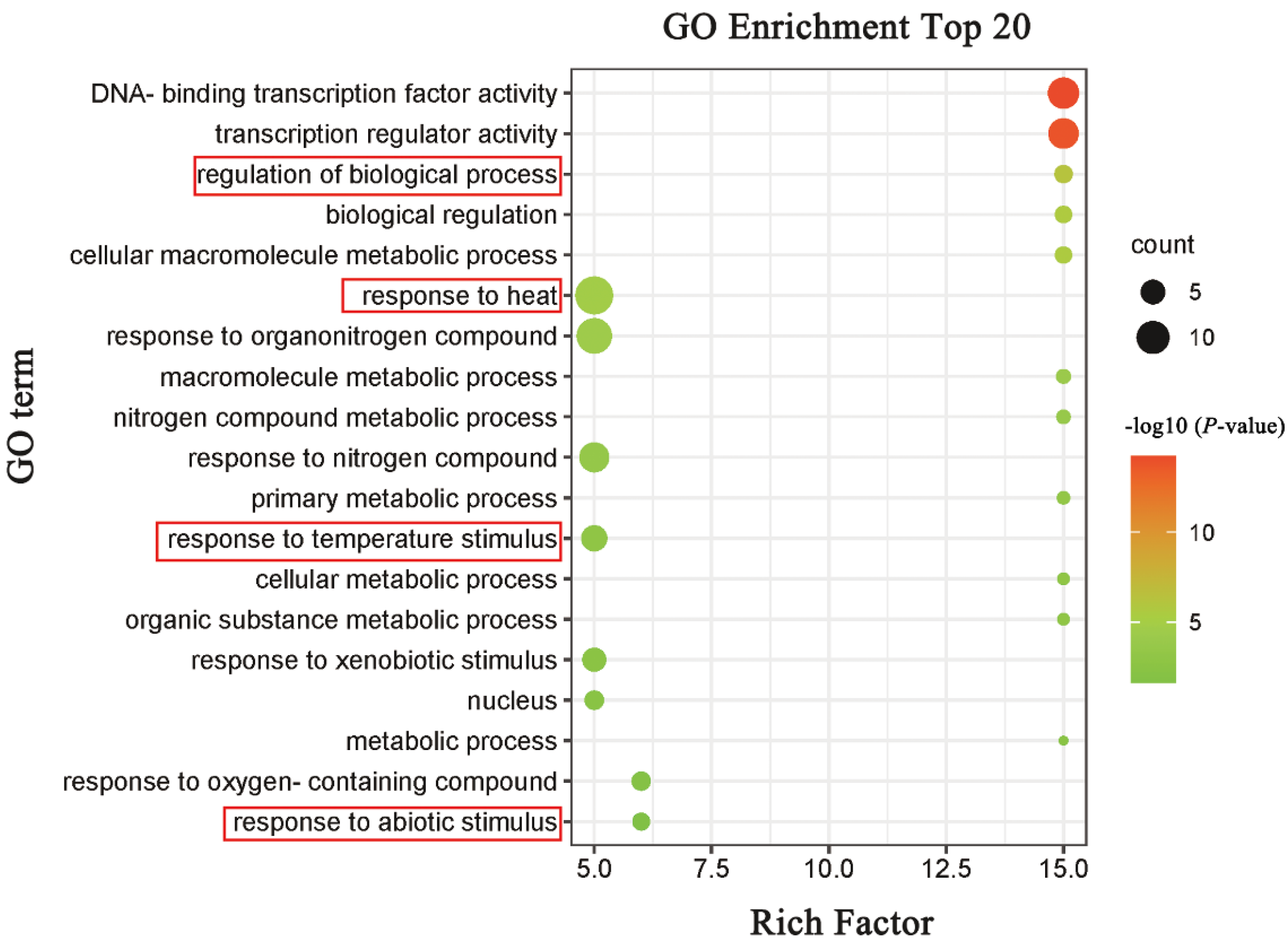
2.9. GO Annotation and Enrichment Analysis of M. sativa HSF Protein
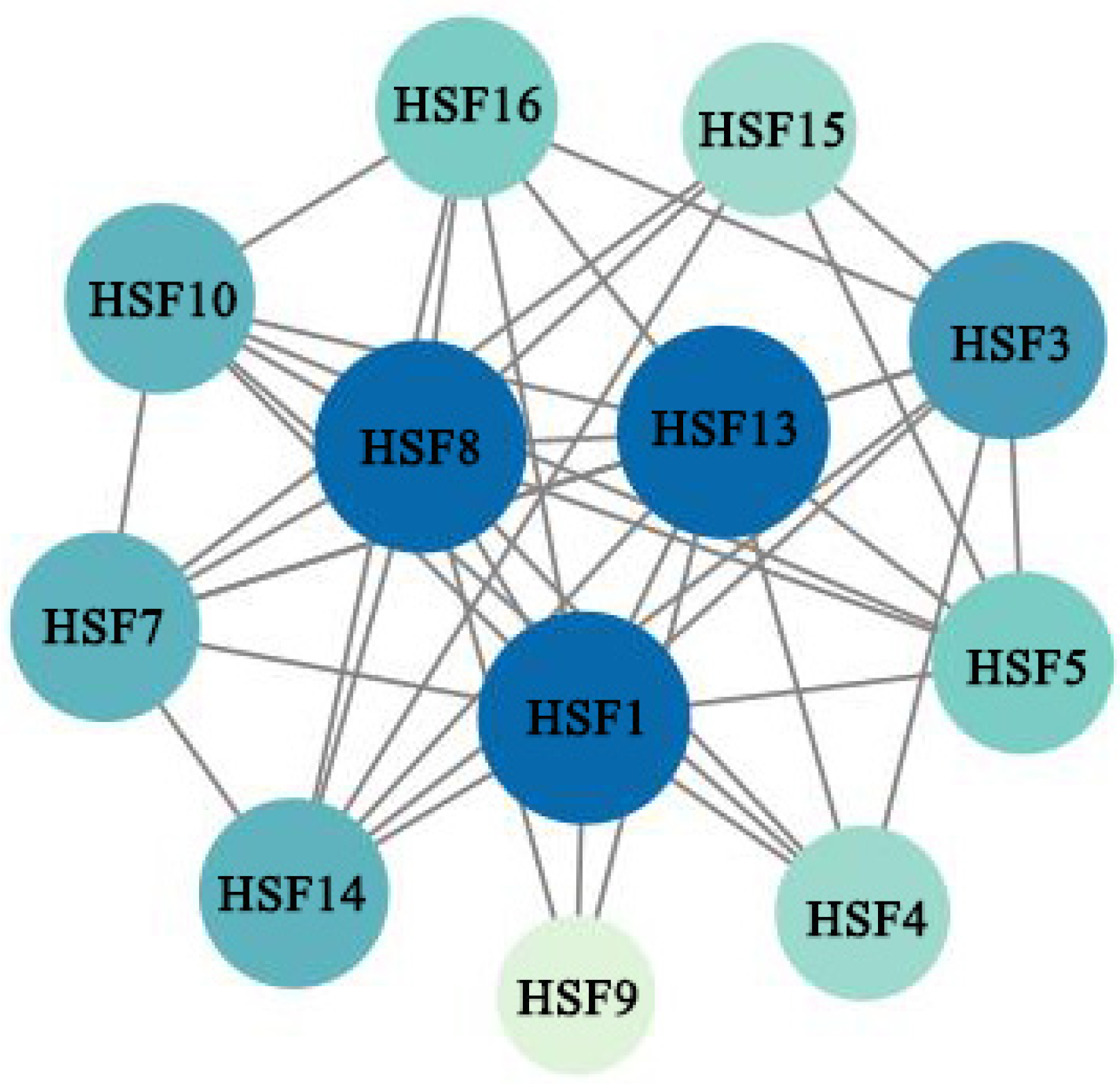
2.10. Interaction Network Analysis of HSF Proteins in M. sativa
3. Discussion
3.1. The Characteristics of HSF Gene Family in M. sativa
3.2. The Potential Roles of Differentially Expressed MsHSF Genes
4. Materials and Methods
4.1. Identification and Sequence Analysis of HSF Genes in M. sativa
4.2. Construction of Phylogenetic Tree and Sequence Comparison
4.3. Gene Structure and Motif Identification
4.4. Chromosome Location and Covariance Analysis
4.5. Identification of Cis-Acting Elements
4.6. Analysis of Tissue-Specific Expression and Abiotic Stress Transcriptome Data
4.7. Protein Interaction Network Prediction and GO Enrichment Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pu, J.; Li, M.; Mao, P.; Zhou, Q.; Liu, W.; Liu, Z. Genome-Wide Identification of the Q-type C2H2 Transcription Factor Family in Alfalfa (Medicago sativa) and Expression Analysis under Different Abiotic Stresses. Genes 2021, 12, 1906. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Lv, J.; Ma, Z.; Dong, W. The mechanism of alfalfa (Medicago sativa L.) response to abiotic stress. Plant Growth Regul. 2019, 89, 239–249. [Google Scholar] [CrossRef]
- Mao, P.; Jin, X.; Bao, Q.; Mei, C.; Zhou, Q.; Min, X.; Liu, Z. WRKY Transcription Factors in Medicago sativa L.: Genome-Wide Identification and Expression Analysis Under Abiotic Stress. DNA Cell Biol. 2020, 39, 2212–2225. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Yu, J.; Kong, L.; Zhang, W.; Hou, X.; Cui, G. Genome-wide investigation of the PLD gene family in alfalfa (Medicago sativa L.): Identification, analysis and expression. BMC Genom. 2022, 23, 243. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Qi, L.; Nian, L.; Zhu, X.; Yi, X.; Jiyu, Z.; Qiu, J. Genome-Wide Identification and Expression Analysis of the SRS Gene Family in Medicago sativa. DNA Cell Biol. 2021, 40, 1539–1553. [Google Scholar] [CrossRef]
- Dong, X.; Deng, H.; Ma, W.; Zhou, Q.; Liu, Z. Genome-wide identification of the MADS-box transcription factor family in autotetraploid cultivated alfalfa and expression analysis under abiotic stress. BMC Genom. 2021, 22, 603. [Google Scholar] [CrossRef]
- Sheng, S.; Guo, X.; Wu, C.; Xiang, Y.; Duan, S.; Yang, W.; Li, W.; Cao, F.; Liu, L. Genome-wide identification and expression analysis of DREB genes in alfalfa (Medicago sativa) in response to cold stress. Plant Signal. Behav. 2022, 17, 2081420. [Google Scholar] [CrossRef]
- He, F.; Wei, C.; Zhang, Y.; Long, R.; Li, M.; Wang, Z.; Yang, Q.; Kang, J.; Chen, L. Genome-Wide Association Analysis Coupled With Transcriptome Analysis Reveals Candidate Genes Related to Salt Stress in Alfalfa (Medicago sativa L.). Front. Plant Sci. 2022, 12, 826584. [Google Scholar] [CrossRef]
- Westerheide, S.D.; Raynes, R.; Powell, C.; Xue, B.; Uversky, V.N. HSF Transcription Factor Family, Heat Shock Response, and Protein Intrinsic Disorder. Curr. Protein Pept. Sci. 2012, 13, 86–103. [Google Scholar] [CrossRef]
- Nian, L.; Zhang, X.; Yi, X.; Liu, X.; Ain, N.U.; Yang, Y.; Li, X.; Haider, F.U.; Zhu, X. Genome-wide identification of ABA receptor PYL/RCAR gene family and their response to cold stress in Medicago sativa L. Physiol. Mol. Biol. Plants 2021, 27, 1979–1995. [Google Scholar] [CrossRef]
- Nian, L.; Liu, X.; Yang, Y.; Zhu, X.; Yi, X.; Haider, F.U. Genome-wide identification, phylogenetic, and expression analysis under abiotic stress conditions of LIM gene family in Medicago sativa L. PLoS ONE 2021, 16, e0252213. [Google Scholar] [CrossRef] [PubMed]
- Panzade, K.P.; Kale, S.S.; Kapale, V.; Chavan, N.R. Genome-Wide Analysis of Heat Shock Transcription Factors in Ziziphus jujuba Identifies Potential Candidates for Crop Improvement Under Abiotic Stress. Appl. Biochem. Biotechnol. 2021, 193, 1023–1041. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, W.; Shi, B. Genome-wide analysis and expression profiling of the heat shock transcription factor gene family in Physic Nut (Jatropha curcas L.). PeerJ 2020, 8, e8467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Song, H.; Li, C.; Li, P.; Li, A.; Guan, H.; Hou, L.; Wang, X. Genome-wide dissection of the heat shock transcription factor family genes in Arachis. Front. Plant Sci. 2017, 8, 106. [Google Scholar] [CrossRef] [Green Version]
- Wan, X.; Yang, J.; Guo, C.; Bao, M.; Zhang, J. Genome-wide identification and classification of the Hsf and sHsp gene families in Prunus mume, and transcriptional analysis under heat stress. PeerJ 2019, 7, e7312. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Xie, F.; Li, Y.; Gong, L.; Luo, Y.; Zhang, Y.; Chen, Q.; Wang, Y.; Lin, Y.; Zhang, Y.; et al. Genome-Wide Analysis of the Heat Shock Transcription Factor Gene Family in Brassica juncea: Structure, Evolution, and Expression Profiles. DNA Cell Biol. 2020, 39, 1990–2004. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Pang, Q.; Li, Z.; Li, Z.; Chen, R.; Sun, G.; Sun, B. Genome-Wide Identification and Characterization of Hsf and Hsp Gene Families and Gene Expression Analysis under Heat Stress in Eggplant (Solanum melongema L.). Horticulturae 2021, 7, 149. [Google Scholar] [CrossRef]
- Rehman, A.; Atif, R.M.; Azhar, M.T.; Peng, Z.; Li, H.; Qin, G.; Jia, Y.; Pan, Z.; He, S.; Qayyum, A.; et al. Genome wide identification, classification and functional characterization of heat shock transcription factors in cultivated and ancestral cottons (Gossypium spp.). Int. J. Biol. Macromol. 2021, 182, 1507–1527. [Google Scholar] [CrossRef]
- Huang, B.; Huang, Z.; Ma, R.; Chen, J.; Zhang, Z.; Yrjälä, K. Genome-wide identification and analysis of the heat shock transcription factor family in moso bamboo (Phyllostachys edulis). Sci. Rep. 2021, 11, 16492. [Google Scholar] [CrossRef]
- Song, X.; Liu, G.; Duan, W.; Liu, T.; Huang, Z.; Ren, J.; Li, Y.; Hou, X. Genome-wide identification, classification and expression analysis of the heat shock transcription factor family in Chinese cabbage. Mol. Genet. Genom. 2014, 289, 541–551. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, B.; Li, J.; Zhang, L.; Wang, Y.; Zheng, H.; Lu, M.; Chen, J. Hsf and Hsp gene families in Populus: Genome-wide identification, organization and correlated expression during development and in stress responses. BMC Genom. 2015, 16, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, G.-P.; Sadat, S.; Drenth, J.; McIntyre, C.L. The heat shock factor family from Triticum aestivum in response to heat and other major abiotic stresses and their role in regulation of heat shock protein genes. J. Exp. Bot. 2014, 65, 539–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Cheng, Y.; Jin, J.; Jin, X.; Jiang, H.; Yan, H.; Cheng, B. Genome Duplication and Gene Loss Affect the Evolution of Heat Shock Transcription Factor Genes in Legumes. PLoS ONE 2014, 9, e102825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.-C.; Li, G.-L.; Liu, Z.-H.; Zhang, H.-M.; Zhang, Y.-M.; Guo, X.-L. Cloning, Localization and Expression Analysis of ZmHsf-like Gene in Zea mays. J. Integr. Agric. 2014, 13, 1230–1238. [Google Scholar] [CrossRef]
- Zhao, L.; Pan, H.; Sun, M.; Zhang, Q. Molecular cloning of Arabidopsis thaliana HSFA2 gene and agrobacterium-mediated genetic transformation of chrysanthemum morifolium ramat. Adv. Intell. Soft Comput. 2010, 134, 827–834. [Google Scholar] [CrossRef]
- Guo, M.; Liu, J.-H.; Ma, X.; Luo, D.-X.; Gong, Z.-H.; Lu, M.-H. The Plant Heat Stress Transcription Factors (HSFs): Structure, Regulation, and Function in Response to Abiotic Stresses. Front. Plant Sci. 2016, 7, 114. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Ren, L.; Li, C.; Zhang, D.; Zhang, X.; Zhou, G.; Gao, D.; Chen, R.; Chen, Y.; Wang, Z.; et al. The genome of a wild Medicago species provides insights into the tolerant mechanisms of legume forage to environmental stress. BMC Biol. 2021, 19, 96. [Google Scholar] [CrossRef]
- Zhang, L.; Jia, X.; Zhao, J.; Hasi, A.; Niu, Y. Molecular characterisation and expression analysis of NAC transcription factor genes in wild Medicago falcata under abiotic stresses. Funct. Plant Biol. 2020, 47, 327–341. [Google Scholar] [CrossRef]
- Xu, C.; He, C.G.; Wang, Y.J.; Bi, Y.F.; Jiang, H. Effect of drought and heat stresses on photosynthesis, pigments, and xanthophyll cycle in alfalfa (Medicago sativa L.). Photosynthetica 2020, 58, 1226–1236. [Google Scholar] [CrossRef]
- Guiza, M.; Benabdelrahim, M.A.; Brini, F.; Haddad, M.; Saibi, W. Assessment of Alfalfa (Medicago sativa L.) Cultivars for Salt Tolerance Based on Yield, Growth, Physiological, and Biochemical Traits. J. Plant Growth Regul. 2021, 10, 1435-8107. [Google Scholar] [CrossRef]
- He, K.; Li, C.; Zhang, Z.; Zhan, L.; Cong, C.; Zhang, D.; Cai, H. Genome-wide investigation of the ZF-HD gene family in two varieties of alfalfa (Medicago sativa L.) and its expression pattern under alkaline stress. BMC Genom. 2022, 23, 150. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Xu, L.; Wang, Y.; Cheng, W.; Luo, X.; Xie, Y.; Fan, L.; Liu, L. Genome-wide characterization and evolutionary analysis of heat shock transcription factors (HSFs) to reveal their potential role under abiotic stresses in radish (Raphanus sativus L.). BMC Genom. 2019, 20, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, M.; Lu, J.-P.; Zhai, Y.-F.; Chai, W.-G.; Gong, Z.-H.; Lu, M.-H. Genome-wide analysis, expression profile of heat shock factor gene family (CaHsfs) and characterisation of CaHsfA2 in pepper (Capsicum annuum L.). BMC Plant Biol. 2015, 15, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, G.; Shen, J.; Yu, H.; Huang, X.; Deng, X.; Hu, Z.; Amee, M.; Chen, L.; Cao, L. Genome-wide identification and functional analyses of heat shock transcription factors involved in heat and drought stresses in ryegrass. Environ. Exp. Bot. 2022, 201, 104968. [Google Scholar] [CrossRef]
- Liu, X.; Meng, P.; Yang, G.; Zhang, M.; Peng, S.; Zhai, M.Z. Genome-wide identification and transcript profiles of walnut heat stress transcription factor involved in abiotic stress. BMC Genom. 2020, 21, 474. [Google Scholar] [CrossRef]
- Zhang, H.Z.; Yang, J.L.; Chen, Y.L.; Mao, X.L.; Wang, Z.C.; Li, C.H. Identification and expression analysis of the heat shock transcription factor (HSF) gene family in Populus trichocarpa. PLANT OMICS 2013, 6, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Fan, K.; Mao, Z.; Ye, F.; Pan, X.; Li, Z.; Lin, W.; Zhang, Y.; Huang, J.; Lin, W. Genome-wide identification and molecular evolution analysis of the heat shock transcription factor (HSF) gene family in four diploid and two allopolyploid Gossypium species. Genomics 2021, 113, 3112–3127. [Google Scholar] [CrossRef]
- Li, W.; Wan, X.-L.; Yu, J.-Y.; Wang, K.-L.; Zhang, J. Genome-wide Identification, classification, and expression analysis of the HSF gene family in carnation (Dianthus caryophyllus). Int. J. Mol. Sci. 2019, 20, 5233. [Google Scholar] [CrossRef] [Green Version]
- Xue, H.; Slavov, D.; Wischmeyer, P. Glutamine activates heat shock transcription factor-1 (HSF1) gene transcription. FASEB J. 2012, 26, 646.27. [Google Scholar] [CrossRef]
- Duan, S.; Liu, B.; Zhang, Y.; Li, G.; Guo, X. Genome-wide identification and abiotic stress-responsive pattern of heat shock transcription factor family in Triticum aestivum L. BMC Genom. 2019, 20, 257. [Google Scholar] [CrossRef]
- Li, P.-S.; Yu, T.-F.; He, G.-H.; Chen, M.; Zhou, Y.-B.; Chai, S.-C.; Xu, Z.-S.; Ma, Y.-Z. Genome-wide analysis of the Hsf family in soybean and functional identification of GmHsf-34 involvement in drought and heat stresses. BMC Genom. 2014, 15, 1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Geng, J.; Du, Y.; Zhao, Q.; Zhang, W.; Fang, Q.; Yin, Z.; Li, J.; Yuan, X.; Fan, Y.; et al. Heat shock transcription factor (Hsf) gene family in common bean (Phaseolus vulgaris): Genome-wide identification, phylogeny, evolutionary expansion and expression analyses at the sprout stage under abiotic stress. BMC Plant Biol. 2022, 22, 33. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.-Y.; Yao, Y.; Hong, Y.-H.; Hou, P.-Y.; Li, C.-X.; Xia, Z.-Q.; Geng, M.-T.; Chen, Y.-H. Differential expression of the Hsf family in cassava under biotic and abiotic stresses. Genome 2019, 62, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Shyamli, P.S.; Pradhan, S.; Panda, M.; Parida, A. De novo Whole-Genome Assembly of Moringa oleifera Helps Identify Genes Regulating Drought Stress Tolerance. Front. Plant Sci. 2021, 12, 766999. [Google Scholar] [CrossRef]
- Shen, C.; Yuan, J. Genome-wide characterization and expression analysis of the heat shock transcription factor family in pumpkin (Cucurbita moschata). BMC Plant Biol. 2020, 20, 471. [Google Scholar] [CrossRef]
- Agarwal, P.; Khurana, P. Functional characterization of HSFs from wheat in response to heat and other abiotic stress conditions. Funct. Integr. Genom. 2019, 19, 497–513. [Google Scholar] [CrossRef]
- Ducy, P.; Karsenty, G. Two distinct osteoblast-specific cis-acting elements control expression of a mouse osteocalcin gene. Mol. Cell. Biol. 1995, 15, 1858–1869. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhong, Y.; Liu, X.; Zhao, C.; Zhao, J.; Li, M.; Hassan, M.U.; Yang, B.; Li, D.; Liu, R.; et al. Cis- regulation of the amino acid transporter genes ZmAAP2 and ZmLHT1 by ZmPHR1 transcription factors in maize ear under phosphate limitation. J. Exp. Bot. 2021, 72, 3846–3863. [Google Scholar] [CrossRef]
- Binder, R.J.; Blachere, N.E.; Srivastava, P.K. Heat shock protein-chaperoned peptides but not free peptides introduced into the cytosol are presented efficiently by major histocompatibility complex I molecules. J. Biol. Chem. 2001, 276, 17163–17171. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Tu, Y.; Wang, X.; Cao, C.; Hu, Y.; Shao, J.; Weng, L.; Mou, X.; Dong, X. A photo-triggered antifungal nanoplatform with efflux pump and heat shock protein reversal activity for enhanced chemo-photothermal synergistic therapy. Biomater. Sci. 2021, 9, 3293–3299. [Google Scholar] [CrossRef]
- Altaff, K.; Radha, V. Influence of heat shock protein (hsp-70) enhancing compound from red alga (Porphyridium purpureum) for augmenting egg production in copepod culture–A new in silico report. Mar. Sci. Technol. Bull. 2021, 10, 186–192. [Google Scholar] [CrossRef]
- Simon, S.; Aissat, A.; Degrugillier, F.; Simonneau, B.; Fanen, P.; Arrigo, A.P. Small Hsps as Therapeutic Targets of Cystic Fibrosis Transmembrane Conductance Regulator Protein. Int. J. Mol. Sci. 2021, 22, 4252. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Hu, X.-L.; Yao, T.; Yang, X.; Chen, J.-G.; Lu, M.-Z.; Zhang, J. Recent advances in the roles of HSFs and HSPs in heat stress response in woody plants. Front. Plant Sci. 2021, 12, 704905. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yu, X.; Wang, D.; Li, L.; Zhou, W.; Zhang, Q.; Wang, X.; Ye, S.; Wang, Z. Genome-wide identification, classification and expression profile analysis of the HSF gene family in Hypericum perforatum. PeerJ 2021, 9, e11345. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Zhu, W.; Song, X.; Lin, X.; Cai, J.; Wang, M.; Yang, Q. Genome-Wide Identification and Function Analyses of Heat Shock Transcription Factors in Potato. Front. Plant Sci. 2016, 7, 490. [Google Scholar] [CrossRef] [Green Version]
- Samtani, H.; Sharma, A.; Khurana, J.P.; Khurana, P. The heat stress transcription factor family in Aegilops tauschii: Genome-wide identification and expression analysis under various abiotic stresses and light conditions. Mol. Genet. Genom. 2022, 2, 11–15. [Google Scholar] [CrossRef]
- Saha, D.; Mukherjee, P.; Dutta, S.; Meena, K.; Sarkar, S.K.; Mandal, A.B.; Dasgupta, T.; Mitra, J. Genomic insights into HSFs as candidate genes for high-temperature stress adaptation and gene editing with minimal off-target effects in flax. Sci. Rep. 2019, 9, 5581. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.; Yan, L.; Li, H.; Lian, X.; Cheng, J.; Wang, W.; Zheng, X.; Wang, X.; Li, J.; Ye, X.; et al. Genome-wide identification of HSF family in peach and functional analysis of PpHSF5 involvement in root and aerial organ development. PeerJ 2021, 9, e10961. [Google Scholar] [CrossRef]
- Eddy, S.R. What is a hidden Markov model? Nat. Biotechnol. 2004, 22, 1315–1316. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Gene ID | Chromosome Location | Size (aa) | MW (kDa) | pI | Stability | A.I | GRAVY | Predicted Location |
|---|---|---|---|---|---|---|---|---|---|
| HSF01 | MsG0180003410.01.T01 | S1:61939996–61942587 | 496 | 55,750.22 | 4.91 | U | 69.91 | −0.591 | Nuclear |
| HSF02 | MsG0180005479.01.T01 | S1:92250440–92250760 | 297 | 32,972.9 | 6.26 | U | 71.28 | −0.613 | Nuclear |
| HSF03 | MsG0380017150.01.T01 | S3:92635955–92637670 | 372 | 42,094.06 | 8.16 | U | 62.82 | −0.715 | Nuclear |
| HSF04 | MsG0380017281.01.T01 | S3:94430904–94433609 | 381 | 42,838.88 | 4.93 | U | 78.5 | −0.547 | Nuclear |
| HSF05 | MsG0480021648.01.T01 | S4:61250000–61252171 | 480 | 53,424.6 | 4.85 | U | 69.62 | −0.55 | Nuclear |
| HSF06 | MsG0480022400.01.T01 | S4:72054039–72055708 | 211 | 24,541.86 | 6.02 | U | 69.72 | −0.708 | Nuclear |
| HSF07 | MsG0480023951.01.T01 | S4:91618543–91621035 | 401 | 45,748.67 | 4.99 | U | 80.7 | −0.498 | Nuclear |
| HSF08 | MsG0580024826.01.T01 | S5:10417724–10420833 | 287 | 32,168.91 | 6.9 | S | 62.79 | −0.784 | Nuclear |
| HSF09 | MsG0580025495.01.T01 | S5:19600912–19606188 | 543 | 60,982.41 | 5.1 | U | 71.47 | −0.624 | Nuclear |
| HSF10 | MsG0680032444.01.T01 | S6:41477418–41479676 | 295 | 33,653.64 | 5.66 | U | 68.14 | −0.802 | Nuclear |
| HSF11 | MsG0680032449.01.T01 | S6:41520725–41522690 | 319 | 36,439.77 | 5.27 | U | 70.34 | −0.717 | Nuclear |
| HSF12 | MsG0680032451.01.T01 | S6:41583472–41590098 | 527 | 59,504.68 | 6.96 | S | 83.61 | −0.474 | Nuclear |
| HSF13 | MsG0680033612.01.T01 | S6:69476487–69478031 | 359 | 39,836.88 | 5.54 | U | 76.3 | −0.439 | Nuclear |
| HSF14 | MsG0680035659.01.T01 | S6:110235176−110236554 | 351 | 40,233.48 | 5.58 | U | 63.85 | −0.664 | Nuclear |
| HSF15 | MsG0780040480.01.T01 | S7:78201225–78202956 | 226 | 26,119.55 | 7.56 | U | 71.55 | −0.755 | Nuclear |
| HSF16 | MsG0780040676.01.T01 | S7:80984782–80994274 | 527 | 60,447.57 | 5.43 | U | 73.76 | −0.61 | Nuclear |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, J.; Zhang, G.; Ye, Y.; Shang, L.; Hong, S.; Ma, Q.; Zhao, Y.; Gu, C. Genome-Wide Identification and Expression Analysis of HSF Transcription Factors in Alfalfa (Medicago sativa) under Abiotic Stress. Plants 2022, 11, 2763. https://doi.org/10.3390/plants11202763
Ma J, Zhang G, Ye Y, Shang L, Hong S, Ma Q, Zhao Y, Gu C. Genome-Wide Identification and Expression Analysis of HSF Transcription Factors in Alfalfa (Medicago sativa) under Abiotic Stress. Plants. 2022; 11(20):2763. https://doi.org/10.3390/plants11202763
Chicago/Turabian StyleMa, Jin, Guozhe Zhang, Yacheng Ye, Linxue Shang, Sidan Hong, Qingqing Ma, Yu Zhao, and Cuihua Gu. 2022. "Genome-Wide Identification and Expression Analysis of HSF Transcription Factors in Alfalfa (Medicago sativa) under Abiotic Stress" Plants 11, no. 20: 2763. https://doi.org/10.3390/plants11202763
APA StyleMa, J., Zhang, G., Ye, Y., Shang, L., Hong, S., Ma, Q., Zhao, Y., & Gu, C. (2022). Genome-Wide Identification and Expression Analysis of HSF Transcription Factors in Alfalfa (Medicago sativa) under Abiotic Stress. Plants, 11(20), 2763. https://doi.org/10.3390/plants11202763