
Evaluation of the Site Form as a Site Productive Indicator in Temperate Uneven-Aged Multispecies Forests in Durango, Mexico

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
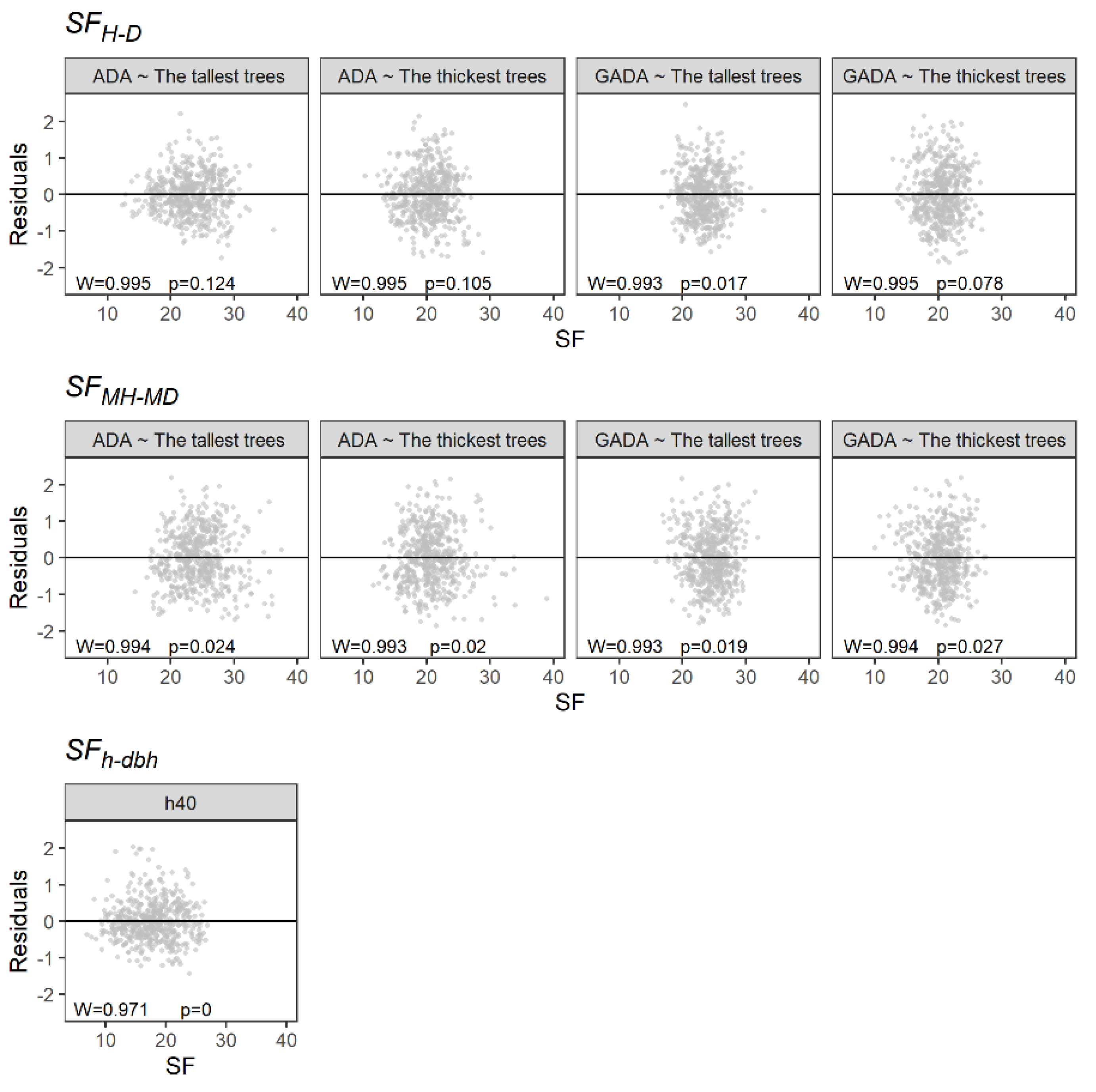
2.1. H-D, MH-MD and h-dbh Model Fitting
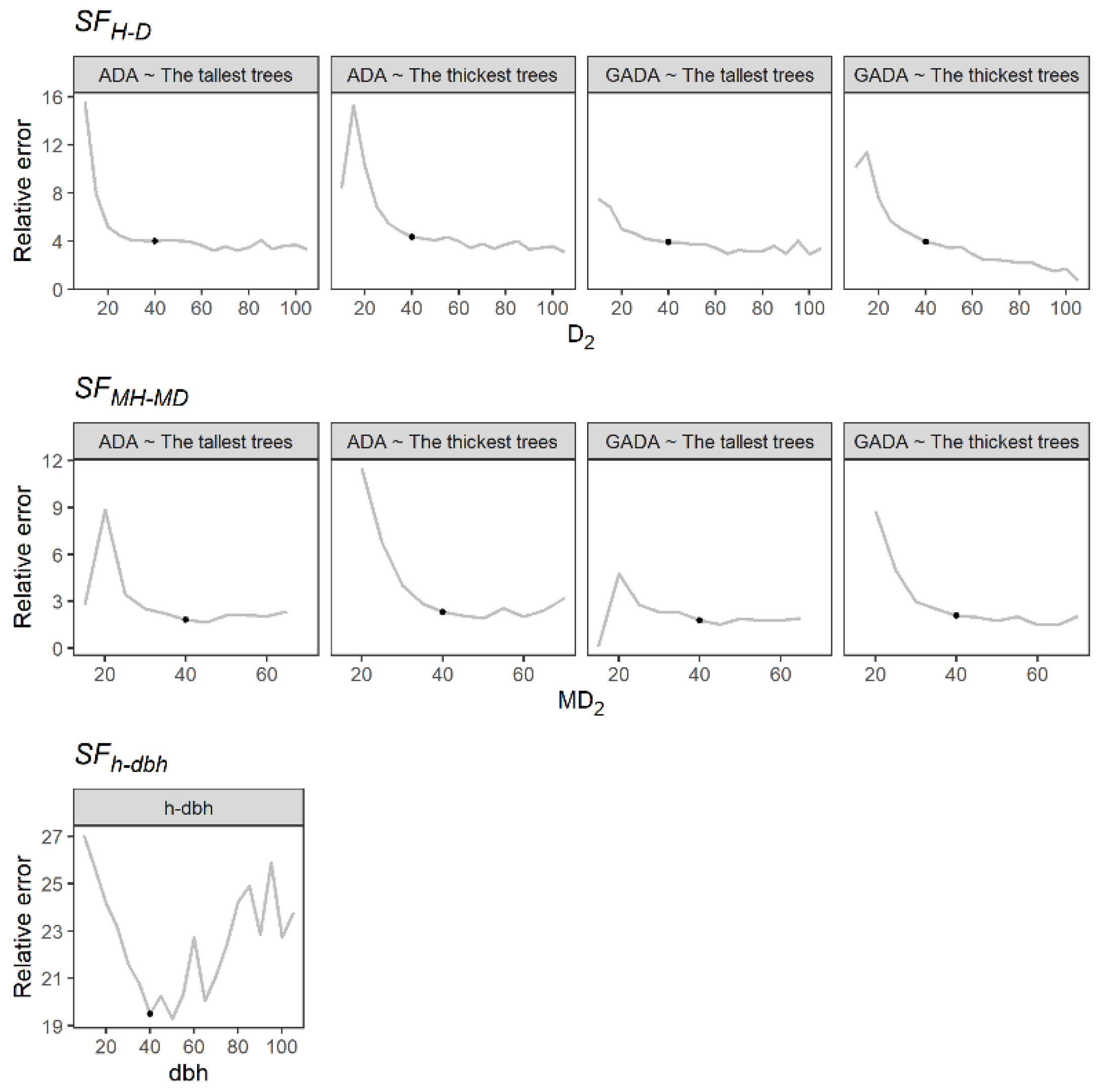
2.2. Reference Diameter Selection
2.3. Evaluation of SF as a Site Quality Indicator
3. Discussion
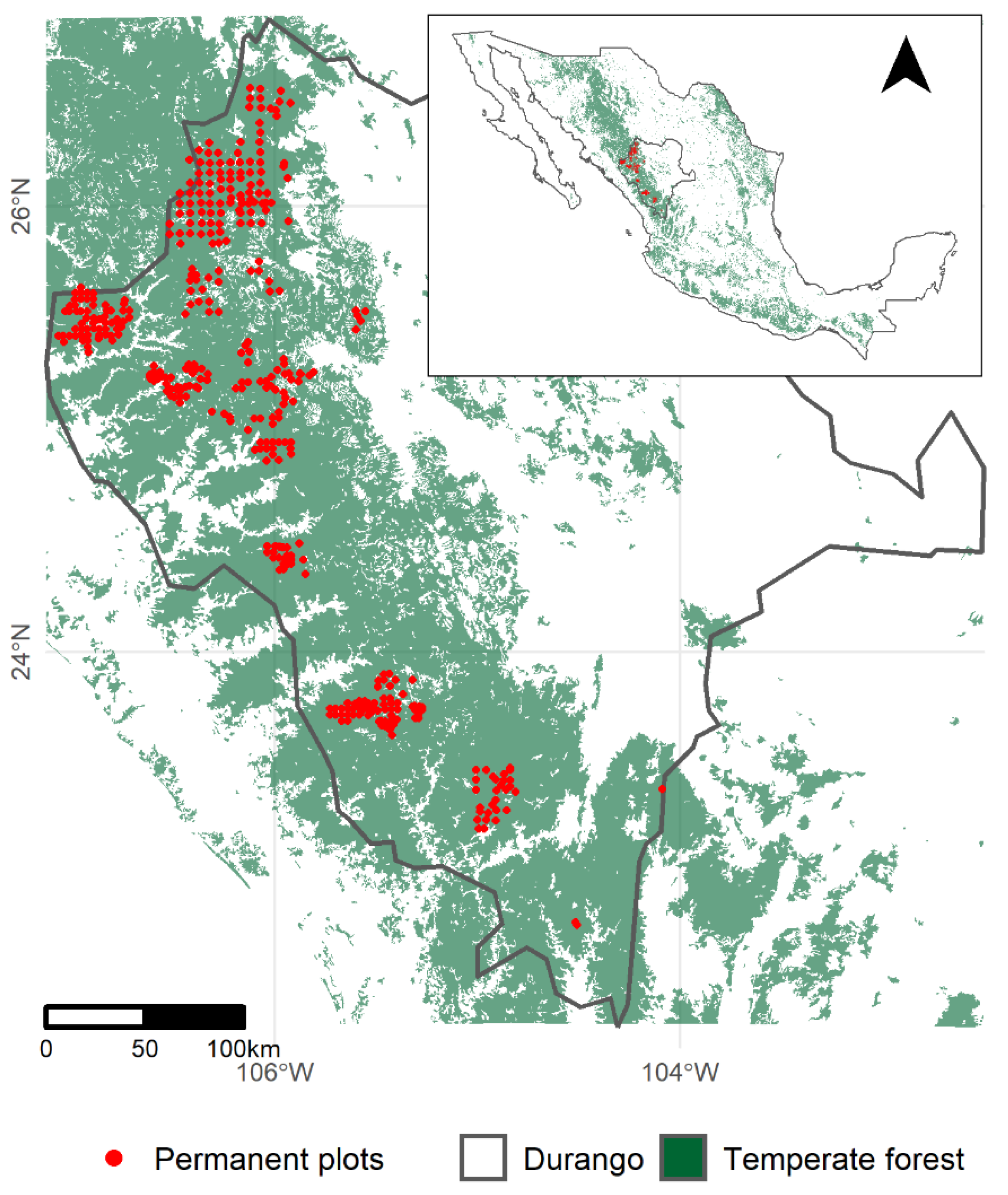
4. Materials and Methods
4.1. H-D, MH-MD and h-dbh Models
4.2. Reference Diameter Selection
4.3. Evaluation of SF as a Site Quality Indicator
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blanco, M.M.E.; Hoppus, M.; Lister, A.; Westfall, J.A. Is there a better metric than site to indicate the productivity of forested land? In Proceedings of the Eighth Annual Forest Inventory and Analysis Symposium, Monterey, CA, USA, 16–19 October 2006; McRoberts, R.E., Reams, G.A., Van Deusen, P.C., McWilliams, W.H., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 2006; pp. 355–362. [Google Scholar]
- Bontemps, J.D.; Bouriaud, O. Predictive approaches to forest site productivity: Recent trends, challenges, and future perspectives. Forestry 2014, 87, 109–128. [Google Scholar] [CrossRef]
- Skovsgaard, J.P.; Vanclay, J.K. Forest site productivity: A review of spatial and temporal variability in natural site conditions. Forestry 2013, 86, 305–315. [Google Scholar] [CrossRef]
- del Río, M.; Pretzsch, H.; Alberdi, I.; Bielak, K.; Bravo, F.; Brunner, A.; Condés, S.; Ducey, M.J.; Fonseca, T.; von Lüpke, N.; et al. Characterization of the structure, dynamics, and productivity of mixed-species stands: A review and perspectives. Eur. J. For. Res. 2015, 135, 23–49. [Google Scholar] [CrossRef]
- Eichhorn, F. Beziehungen zwischen Bestandshöhe und Bestandsmasse. Allg. Forst. Jagdztg. 1904, 80, 45–49. [Google Scholar]
- Burkhart, H.E.; Tomé, M. Modeling Forest Trees and Stands; Springer: Amsterdam, The Netherlands, 2012; p. 458. [Google Scholar]
- Huang, S.; Titus, S.J. An index of site productivity for uneven-aged or mixed-species stands. Can. J. For. Res. 1993, 23, 558–562. [Google Scholar] [CrossRef]
- Flury, P. Über den Aufbau des Plenterwaldes. Mitt. Der Schweiz. Cent. Für Das Vers. 1929, 15, 305–357. [Google Scholar]
- von Gadow, K.; Sánchez, O.S.; Álvarez, G.J.G. Estructura y Crecimiento del Bosque; Universidad de Santiago de Compostela: Lugo, Spain, 2007; p. 282. [Google Scholar]
- Vanclay, J.K.; Henry, N.B. Assessing site productivity of indigenous cypress pine forest in southern Queensland. Commonw. For. Rev. 1988, 67, 53–64. [Google Scholar]
- Wang, G.G. Is height of dominant trees at a reference diameter an adequate measure of site quality? For. Ecol. Manag. 1998, 112, 49–54. [Google Scholar] [CrossRef]
- Fu, L.; Lei, X.; Sharma, R.P.; Li, H.; Zhu, G.; Hong, L.; You, L.; Duan, G.; Guo, H.; Lei, Y.; et al. Comparing height-age and height-diameter modelling approaches for estimating site productivity of natural uneven-aged forests. Forestry 2018, 91, 419–433. [Google Scholar] [CrossRef]
- Molina-Valero, J.A.; Diéguez-Aranda, U.; Álvarez-González, J.G.; Castedo-Dorado, F.; Pérez-Cruzado, C. Assessing site form as an indicator of site quality in even-aged Pinus Radiata D. Don stands in north-western Spain. Ann. For. Sci. 2019, 76, 113. [Google Scholar] [CrossRef] [Green Version]
- Do, H.T.T.; Zimmer, H.C.; Vanclay, J.K.; Grant, J.C.; Trinh, B.N.; Nguyen, H.H.; Nichols, J.D. Site form classification– a practical tool for guiding site-specific tropical forest landscape restoration and management. Forestry 2021, 95, 261–273. [Google Scholar] [CrossRef]
- Asthon, M.S.; Kelty, M.J. The Practice of Silviculture Applied Forest Ecology; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2018; p. 758. [Google Scholar]
- Sharma, M.; Amateis, R.L.; Burkhart, H.E. Top height definition and its index determination in thinned and unthinned loblolly pine plantations. For. Ecol. Manag. 2002, 168, 163–175. [Google Scholar] [CrossRef]
- Kuuluvainen, T. Forest management and biodiversity conservation based on natural ecosystem dynamics in the northern Europe: The complexity challenge. Ambio 2009, 38, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, aaf8957. [Google Scholar] [CrossRef] [Green Version]
- Torres-Rojo, J.M.; Moreno-Sánchez, R.; Mendoza-Briseño, M.A. Sustainable forest management in Mexico. Curr. For. Rep. 2016, 2, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Reich, R.M.; Bravo, V.A. New approach for modeling climatic data with applications in modeling tree species distributions in the states of Jalisco and Colima, Mexico. J. Arid. Environ. 2008, 72, 1343–1357. [Google Scholar] [CrossRef]
- Mohamed, A.; Reich, R.M.; Khosla, R.; Aguirre-Bravo, C.; Mendoza, B.M. Influence of climatic, topography and soil attributes on the spatial distribution of site productivity index of the species rich forests of Jalisco, Mexico. J. For. Res. 2014, 25, 87–95. [Google Scholar] [CrossRef]
- Alfaro, R.T.; Martínez-Vilalta, J.; Retana, J. Regeneration patterns in Mexican pine-oak forests. For. Ecosyst. 2019, 6, 50. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Larreta, B.; López-Martínez, J.O.; González, E.J.; Corral-Rivas, J.J.; Hernández, F.J. Assessing above-ground biomass-functional diversity relationships in temperate forests in northern Mexico. For. Ecosyst. 2021, 8, 8. [Google Scholar] [CrossRef]
- Pretzsch, H.; Forrester, D.I.; Bauhus, J. Mixed-Species Forest, Ecology and Management; Springer: Berlin/Heidelberg, Germany, 2017; p. 653. [Google Scholar]
- Corral-Rivas, J.J.; Torres-Rojo, J.M.; Lujan-Soto, J.E.; Nava-Miranda, M.G.; Aguirre-Calderón, O.A.; von Gadow, K. Density and production in the natural forests of Durango/Mexico. Allg. Forst. Jagdztg. 2016, 187, 93–103. [Google Scholar]
- Navar, J. Modeling tree diversity, stand structure and productivity of northern temperate coniferous of Mexico. PeerJ 2019, 7, e7051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, M.; Kiviste, A.; von Gadow, K. A spatially explicit height-diameter model for Scots pine in Estonia. Eur. J. For. Res. 2011, 130, 303–315. [Google Scholar] [CrossRef] [Green Version]
- Temesgen, H.; Zhang, C.H.; Zhao, X.H. Modelling tree height-diameter relationships in multi-species and multi-layered forests: A large observational study from Northeast China. For. Ecol. Manag. 2014, 316, 78–89. [Google Scholar] [CrossRef]
- Shen, J.; Lei, X.; Lei, Y.; Li, Y. Comparison between site index and site form for site quality evaluation of Larix olgensis plantation. J. Beijing For. Univ. 2018, 40, 1–8. [Google Scholar]
- Kelty, M.J. Comparative productivity of monocultures and mixed species stands. In The Ecology and Silviculture of Mixed-Species Forests; Kelty, M.J., Larson, B.C., Oliver, C.D., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1992. [Google Scholar]
- Garber, S.M.; Maguire, D.A. Stand productivity and development in two mixed-species spacing trials in the Central Oregon Cascades. For. Sci. 2004, 50, 92–105. [Google Scholar]
- Jacob, M.; Leuschner, C.; Thomas, F.M. Productivity of temperate broad-leaved forest stands differing in tree species diversity. Ann. For. Sci. 2010, 67, 503. [Google Scholar] [CrossRef]
- Berrill, J.P.; O’Hara, K.L. Estimating site productivity in irregular stand structures by indexing the basal area or volume increment of the dominant species. Can. J. For. Res. 2014, 44, 92–100. [Google Scholar] [CrossRef]
- Antón-Fernández, C.; Burkhart, H.E.; Strub, M.; Amateis, R.L. Effects of initial spacing on height development of loblolly pine. For. Sci. 2011, 53, 201–211. [Google Scholar]
- Trouvé, R.; Bontemps, J.D.; Collet, C.; Seynave, I.; Lebourgeois, F. When do dendrometric rules fail? Insights from 20 years of experimental thinnings on sessile oak in the GIS Coop network. For. Ecol. Manag. 2019, 433, 276–286. [Google Scholar] [CrossRef]
- Toraño-Caicoya, A.; Pretzsch, H. Stand density biases the estimation of the site index especially on dry sites. Can. J. For. Res. 2020, 51, 1050–1064. [Google Scholar] [CrossRef]
- Peracca, G.G.; O’Hara, K.L. Effects of growing space on growth for 20-year-old giant sequoia, ponderosa pine and Douglas-fir in the Sierra Nevada. West. J. Appl. For. 2008, 23, 156–165. [Google Scholar] [CrossRef] [Green Version]
- Vospernik, S.; Monserud, R.A.; Sterba, H. Do individual-tree growth models correctly represent height:diameter ratios of Norway spruce and Scots pine? For. Ecol. Manag. 2010, 260, 1735–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalho, F.M.G.; Pimineta, E.M.; Goulart, C.P.; de Almeida, M.N.F.; Vidaurre, G.B.; Hein, P.R.G. Effect of stand density on longitudinal variation of wood and bark growth in fast-growing Eucalyptus plantations. iForest 2019, 12, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Macfarlane, D.W.; Green, E.J.; Burkhart, H.E. Population density influences assessment and application of site index. Can. J. For. Res. 2000, 30, 1472–1475. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Dinh, T.T.; Shen, H.L. Height-diameter relationship for Pinus koraiensis in Mengjiagang forest farm of northeast China using nonlinear regressions and artificial neural network models. J. For. Sci. 2019, 65, 134–143. [Google Scholar] [CrossRef] [Green Version]
- Wiedemann, E. Die Rotbuche 1931; M. & H. Schaper: Hannover, Germany, 1932; p. 191. [Google Scholar]
- Assmann, E. Grundflächen- und Volumzuwachs der Rotbuche bei verschiedenen Durchforstungsgraden. Forstwiss. Centralbl. 1950, 69, 256–286. [Google Scholar] [CrossRef]
- Moller, C.M. The effect of thinning, age, and site on foliage, increment, and loss of dry matter. J. For. 1947, 45, 393–404. [Google Scholar]
- Torres-Rojo, J.M. Exploring volume growth-density of mixed multiaged stands in northern Mexico. Agrociencia 2014, 48, 447–461. [Google Scholar]
- Padilla-Martínez, J.R.; Corral-Rivas, J.J.; Briseño-Reyes, J.; Paul, C.; López-Serrano, P.M.; von Gadow, K. Patterns of density and production in the community forests of the Sierra Madre Occidental, Mexico. Forests 2020, 11, 307. [Google Scholar] [CrossRef]
- Corral, R.J.J.; Álvarez, G.J.G.; Ruíz, G.A.A.; von Gadow, K. Compatible height and site index models for five pine species in El Salto, Durango (Mexico). For. Ecol. Manag. 2004, 2, 145–160. [Google Scholar] [CrossRef]
- Castillo, L.A.; Vargas-Larreta, B.; Corral, R.J.J.; Nájera, L.J.A.; Cruz, C.F.; Hernández, F.J. Modelo compatible altura- índice de sitio para cuatro especies de pino en Santiago Papasquiaro, Durango. Rev. Mex. Cienc. For. 2013, 4, 86–103. [Google Scholar]
- Quiñonez-Barraza, G.; De los Santos-Posadas, H.M.; Cruz-Cobos, F.; Velázquez-Martínez, A.; Ángeles-Pérez, G.; Ramírez-Valverde, G. Índice de sitio con polimorfismo complejo para masas forestales de Durango, México. Agrociencia 2015, 49, 439–454. [Google Scholar]
- Corral-Rivas, J.J.; Vargas-Larreta, B.; Wehenkel, C.; Aguirre-Calderón, O.A.; Crecente-Campo, F. Guía Para el Establecimiento, Seguimiento y Evaluación de Sitios Permanentes de Monitoreo de Paisajes Productivos Forestales; Fondo Sectorial para la Investigación, el Desarrollo y la Innovación Tecnológica Forestal: Distrito Federal, Mexico, 2013; p. 82. [Google Scholar]
- Silva-Flores, R.; Pérez-Verdín, G.; Wehenkel, C. Patterns of trees diversity in relation climatic factors on the Sierra Madre Occidental, Mexico. PLoS ONE 2014, 9, e105034. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, F. A new growth curve and its application to timber yield studies. J. For. 1939, 37, 819–820. [Google Scholar]
- Martínez-Salvador, M.; Mata-Gonzalez, R.; Pinedo-Alvarez, A.; Morales-Nieta, C.R.; Prieto-Amparán, J.A.; Vázquez-Quintero, G.; Villareal-Guerrero, F. A spatial forestry productivity potential model for Pinus arizonica Engelm, a key timber species from Northwest Mexico. Sustainability 2019, 11, 829. [Google Scholar] [CrossRef] [Green Version]
- Hernández, J.F.; Meraz, A.J.C.; Vargas, L.B.; Nájera, L.J.A. Diameter, height, basal area and volume growth of three pine species from Chihuahua, Mexico. Rev. Mex. Cienc. For. 2020, 11, 120–143. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org (accessed on 8 September 2022).
- Bailey, R.L.; Clutter, J.L. Base-age invariant polymorphic site curves. For. Sci. 1974, 20, 155–159. [Google Scholar]
- Cieszewski, C.J.; Bailey, R.L. Generalized algebraic difference approach: Theory based derivation of dynamic site equations with polymorphism and variable asymptotes. For. Sci. 2000, 46, 116–126. [Google Scholar]
- Trim, K.R.; Coble, D.W.; Weng, Y.; Stovall, J.P.; Hung, I. A new site index model for intensively managed loblolly pine (Pinus taeda) plantations in the West Gulf Coastal Plain. For. Sci. 2020, 66, 2–13. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models. 2020. Available online: https://CRAN.R-project.org/package=nlme (accessed on 8 September 2022).
- Simental-Cano, B.; López-Sánchez, C.A.; Wehenkel, C.; Vargas-Larreta, B.; Álvarez-González, J.G.; Corral-Rivas, J.J. Species-specific and regional volume models for 12 forest species in Durango, Mexico. Rchscfa 2016, 23, 155–171. [Google Scholar] [CrossRef]
- Reineke, L.H. Perfecting a stand-density index for even-aged forests. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Model | Trees Selected | Parameter | Estimator | SE | R2 | RMSE |
|---|---|---|---|---|---|---|---|
| SFH-D | Sc. ADA | The 100 tallest trees | 23.02 | 0.23 | 0.98 | 0.86 | |
| SFH-D | Sc. GADA | The 100 tallest trees | 534.60 | 38.45 | 0.98 | 0.83 | |
| −144.44 | 9.83 | ||||||
| SFH-D | Sc. ADA | The 100 thickest trees | 39.67 | 0.36 | 0.98 | 0.89 | |
| SFH-D | Sc. GADA | The 100 thickest trees | 845.25 | 112.33 | 0.99 | 0.77 | |
| −209.41 | 26.29 | ||||||
| SFMH-MD | Sc. ADA | The 100 tallest trees | 43.34 | 0.99 | 0.99 | 0.49 | |
| SFMH-MD | Sc. GADA | The 100 tallest trees | 347.70 | 66.13 | 0.99 | 0.43 | |
| −91.35 | 15.29 | ||||||
| SFMH-MD | Sc. ADA | The 100 thickest trees | 63.17 | 1.30 | 0.99 | 0.52 | |
| SFMH-MD | Sc. GADA | The 100 thickest trees | 394.75 | 75.84 | 0.99 | 0.44 | |
| −100.81 | 16.54 | ||||||
| SFh-dbh | Schumacher | All trees | 0.62 | 2.86 |
| Pearson Correlation | ||||||
|---|---|---|---|---|---|---|
| Method | Model | Trees Selected | Coniferous Individuals’ Proportion | |||
| 0–0.25 | 0.26–0.5 | 0.51–0.75 | 0.76–1 | |||
| SFH-D | Sc. ADA | The 100 tallest trees | 0.74 | 0.71 | 0.58 | 0.75 |
| SFH-D | Sc. GADA | The 100 tallest trees | 0.42 | 0.46 | 0.44 | 0.6 |
| SFH-D | Sc. ADA | The 100 thickest trees | 0.21 | 0.53 | 0.43 | 0.48 |
| SFH-D | Sc. GADA | The 100 thickest trees | 0.05 | 0.4 | 0.33 | 0.4 |
| SFMH-MD | Sc. ADA | The 100 tallest trees | 0.37 | 0.15 | 0.14 | 0.35 |
| SFMH-MD | Sc. GADA | The 100 tallest trees | 0.28 | −0.01 | 0.08 | 0.21 |
| SFMH-MD | Sc. ADA | The 100 thickest trees | −0.08 | −0.16 | −0.08 | −0.04 |
| SFMH-MD | Sc. GADA | The 100 thickest trees | −0.05 | −0.15 | −0.11 | −0.08 |
| SFh-dbh | Schumacher | All trees | 0.41 | 0.79 | 0.7 | 0.66 |
| Method | Model | Trees Selected | Intercept | Slope | R2 | RMSE |
|---|---|---|---|---|---|---|
| SFH-D | Sc. ADA | The 100 tallest trees | −0.597 | 0.139 | 0.46 | 0.552 |
| SFH-D | Sc. GADA | The 100 tallest trees | −1.146 | 0.160 | 0.27 | 0.642 |
| SFH-D | Sc. ADA | The 100 thickest trees | 0.242 ns | 0.117 | 0.22 | 0.665 |
| SFH-D | Sc. GADA | The 100 thickest trees | 1.350 | 0.062 | 0.04 | 0.738 |
| SFMH-MD | Sc. ADA | The 100 tallest trees | 1.391 | 0.050 | 0.07 | 0.729 |
| SFMH-MD | Sc. GADA | The 100 tallest trees | 1.621 | 0.040 | 0.02 | 0.746 |
| SFMH-MD | Sc. ADA | The 100 thickest trees | 2.847 | −0.012 ns | 0.00 | 0.753 |
| SFMH-MD | Sc. GADA | The 100 thickest trees | 3.053 | −0.022 ns | 0.01 | 0.751 |
| SFh-dbh | Schumacher | All trees | 0.290 | 0.131 | 0.49 | 0.541 |
| Variable | 1st Inventory (423 Plots) 2007 | 2nd Inventory (423 Plots) 2012 | 3rd Inventory (107 Plots) 2017 | |||
|---|---|---|---|---|---|---|
| Mean (Min–Max) | S.D. | Mean (Min–Max) | S.D. | Mean (Min–Max) | S.D. | |
| S | 8 (2–15) | 2 | 8 (2–16) | 3 | 8 (2–14) | 2 |
| N | 622 (120–2148) | 272 | 606 (108–2060) | 269 | 692 (140–2092) | 318 |
| dbh | 17.9 (7.5–104) | 10.6 | 18.8 (7.5–105.5) | 10.8 | 18.7 (7.5–106) | 10.9 |
| h | 10.7 (1.5–48.2) | 5.6 | 11.8 (1.5–49.4) | 5.8 | 12.9 (2.1–49.7) | 6 |
| G | 21.4 (3.1–53.9) | 8.09 | 22.3 (3.7–55.9) | 8.51 | 25.2 (4.4–58.7) | 10.5 |
| V | 194.7 (12.1–697.3) | 109.0 | 215.5 (15.7–786.2) | 119.6 | 261.6 (19.7–851.3) | 151.1 |
| Dq | 21.6 (12.4–51.0) | 4.7 | 22.3 (12.9–52.2) | 4.8 | 22.1 (13.4–41.8) | 4.90 |
| SDI | 449.5 (71.7–1047.5) | 155.4 | 462.8 (84.4–988.6) | 162.2 | 523.2 (95.9–1044.3) | 199.7 |
| PAI | 7.3 (0.7–23.5) | 4.2 | 9.2 (0.6–22.7) | 4.9 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Padilla-Martínez, J.R.; Paul, C.; Corral-Rivas, J.J.; Husmann, K.; Diéguez-Aranda, U.; von Gadow, K. Evaluation of the Site Form as a Site Productive Indicator in Temperate Uneven-Aged Multispecies Forests in Durango, Mexico. Plants 2022, 11, 2764. https://doi.org/10.3390/plants11202764
Padilla-Martínez JR, Paul C, Corral-Rivas JJ, Husmann K, Diéguez-Aranda U, von Gadow K. Evaluation of the Site Form as a Site Productive Indicator in Temperate Uneven-Aged Multispecies Forests in Durango, Mexico. Plants. 2022; 11(20):2764. https://doi.org/10.3390/plants11202764
Chicago/Turabian StylePadilla-Martínez, Jaime Roberto, Carola Paul, José Javier Corral-Rivas, Kai Husmann, Ulises Diéguez-Aranda, and Klaus von Gadow. 2022. "Evaluation of the Site Form as a Site Productive Indicator in Temperate Uneven-Aged Multispecies Forests in Durango, Mexico" Plants 11, no. 20: 2764. https://doi.org/10.3390/plants11202764
APA StylePadilla-Martínez, J. R., Paul, C., Corral-Rivas, J. J., Husmann, K., Diéguez-Aranda, U., & von Gadow, K. (2022). Evaluation of the Site Form as a Site Productive Indicator in Temperate Uneven-Aged Multispecies Forests in Durango, Mexico. Plants, 11(20), 2764. https://doi.org/10.3390/plants11202764