
Sesame as an Alternative Host Plant to Establish and Retain Predatory Mirids in Open-Field Tomatoes


 , ,
, ,  and
and 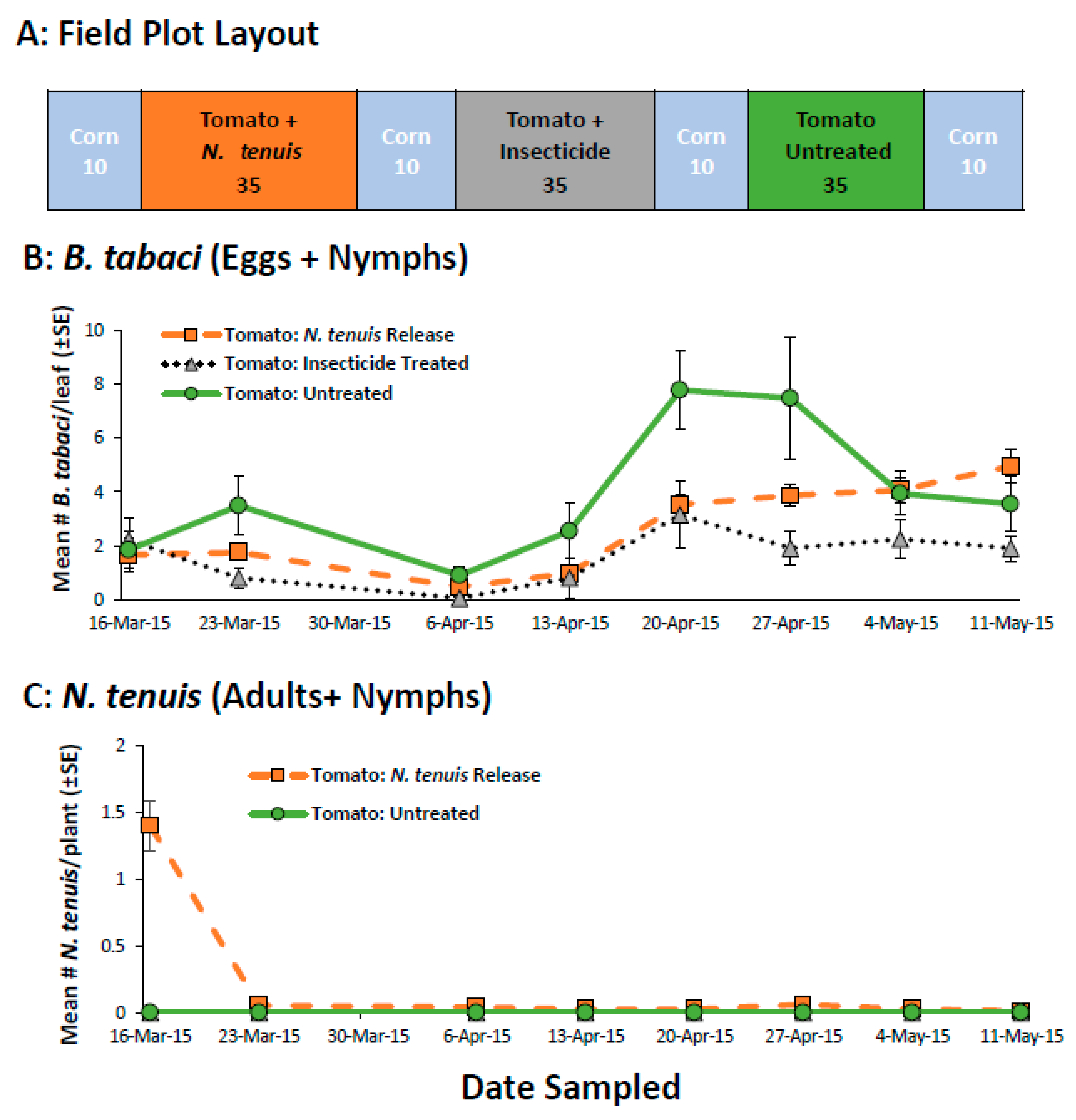{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Pre-Plant Inoculation of Tomato to Establish N. tenuis in Open Field Tomatoes
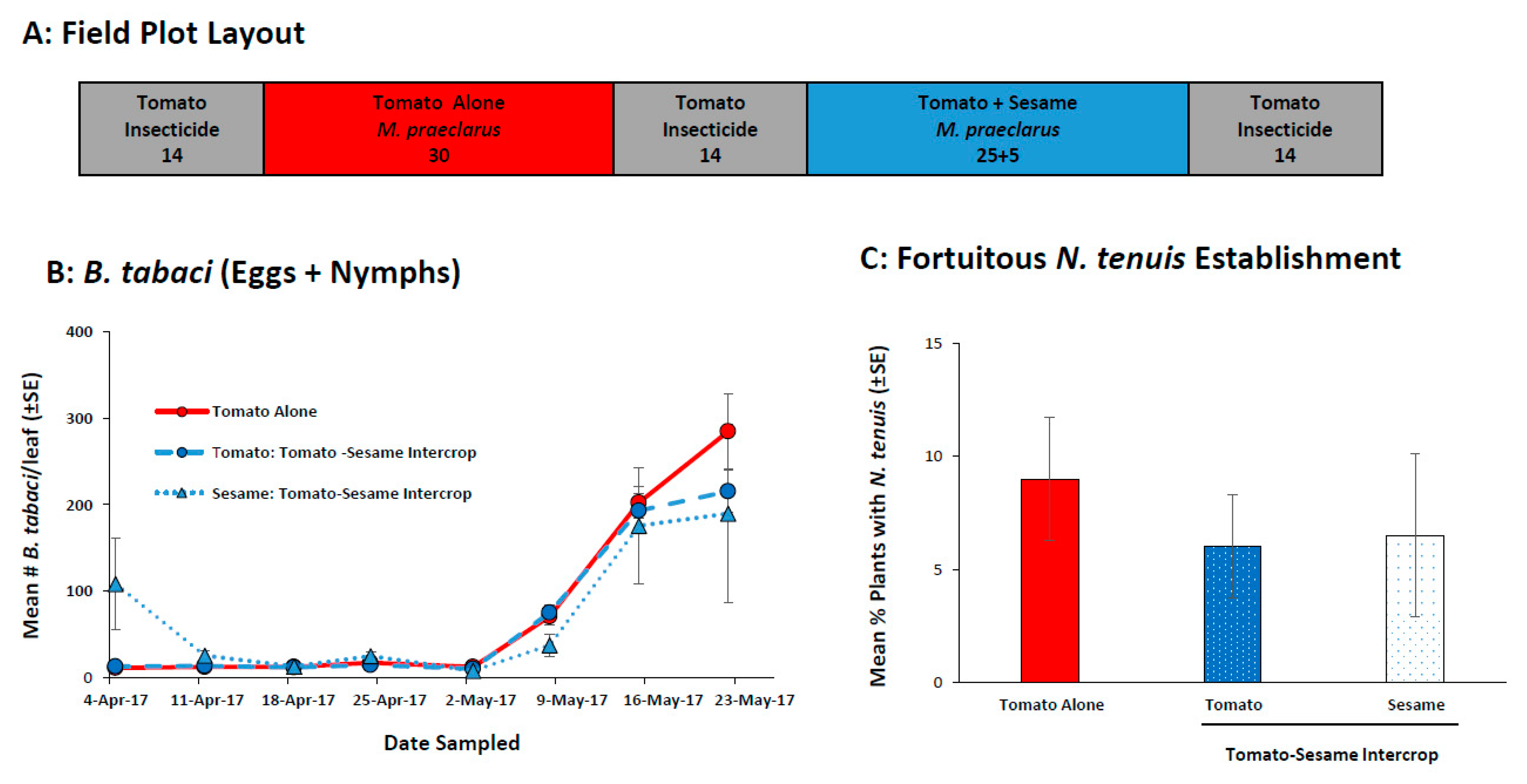
2.2. Tomato-Sesame Intercrop to Retain Zoophyatophagus Mirids
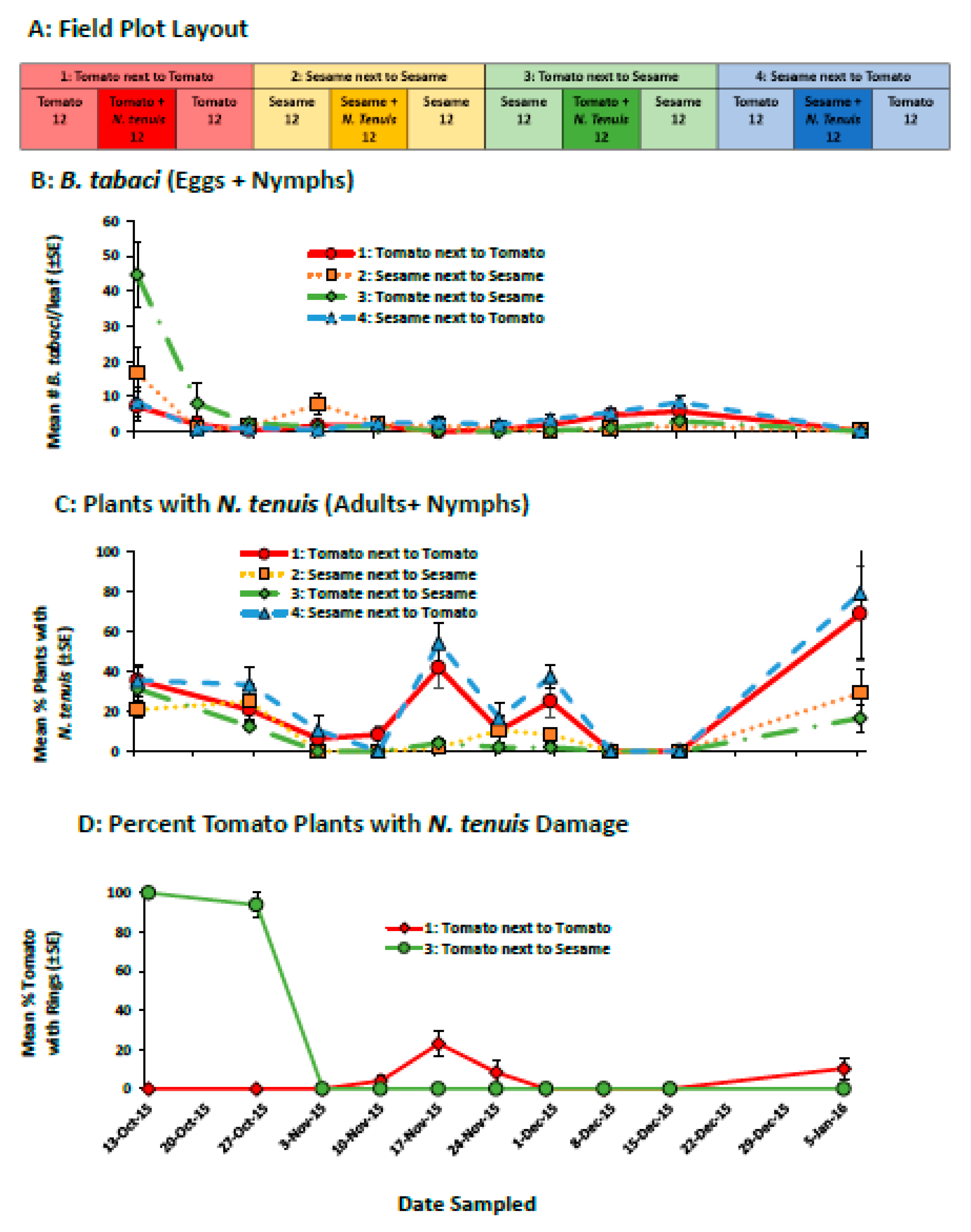
2.2.1. Field Evaluation of Sesame to Retain N. tenuis in Tomato
2.2.2. Field Evaluation of Reduced Amount of Sesame in Tomato-Sesame Interplanting
2.2.3. Field Comparison of Pesticide and N. tenuis in a Tomato-Sesame Intercrop to Control B. tabaci
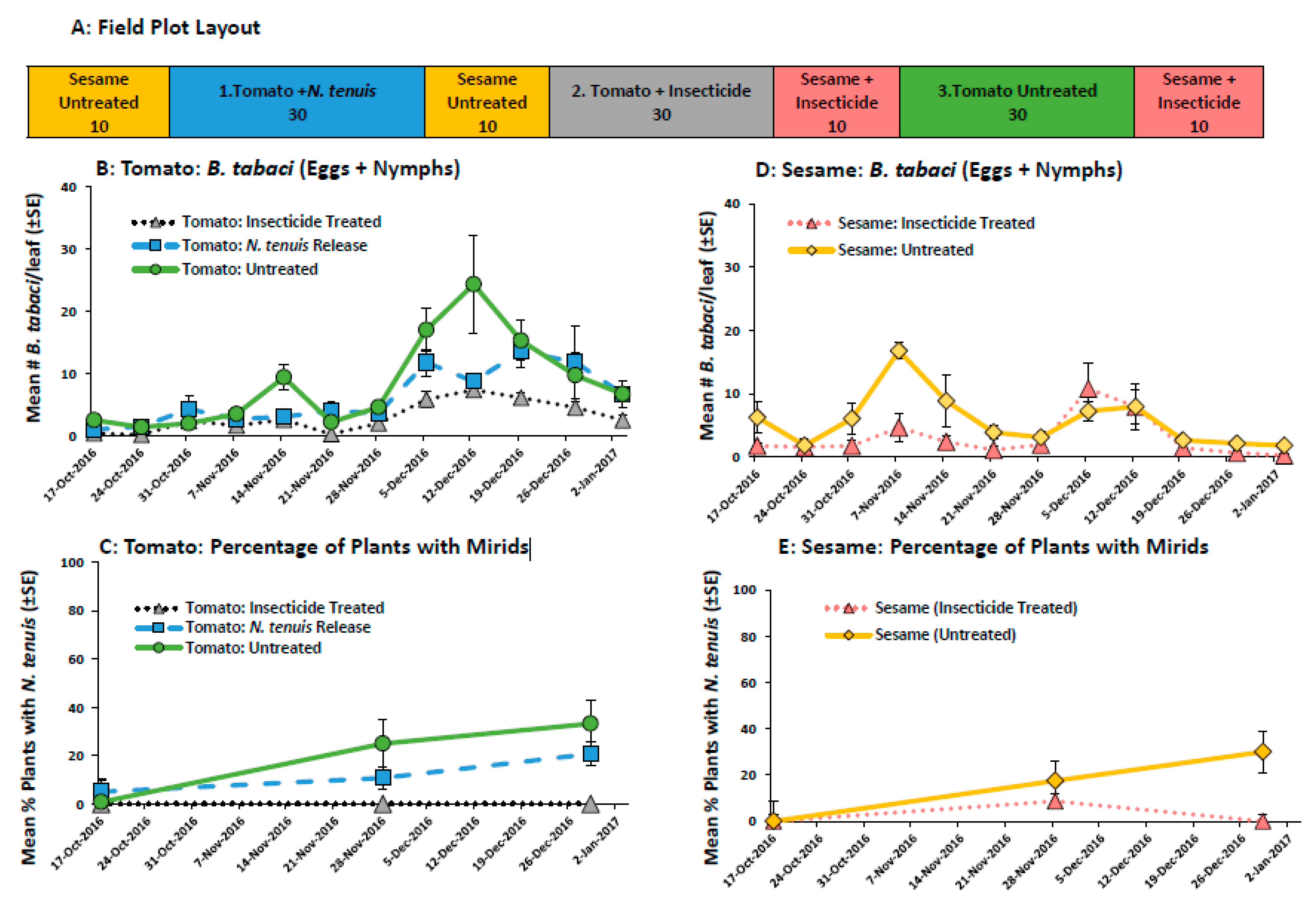
2.2.4. Field Evaluation of M. praeclarus to Control B. tabaci in a Tomato-Sesame Intercrop
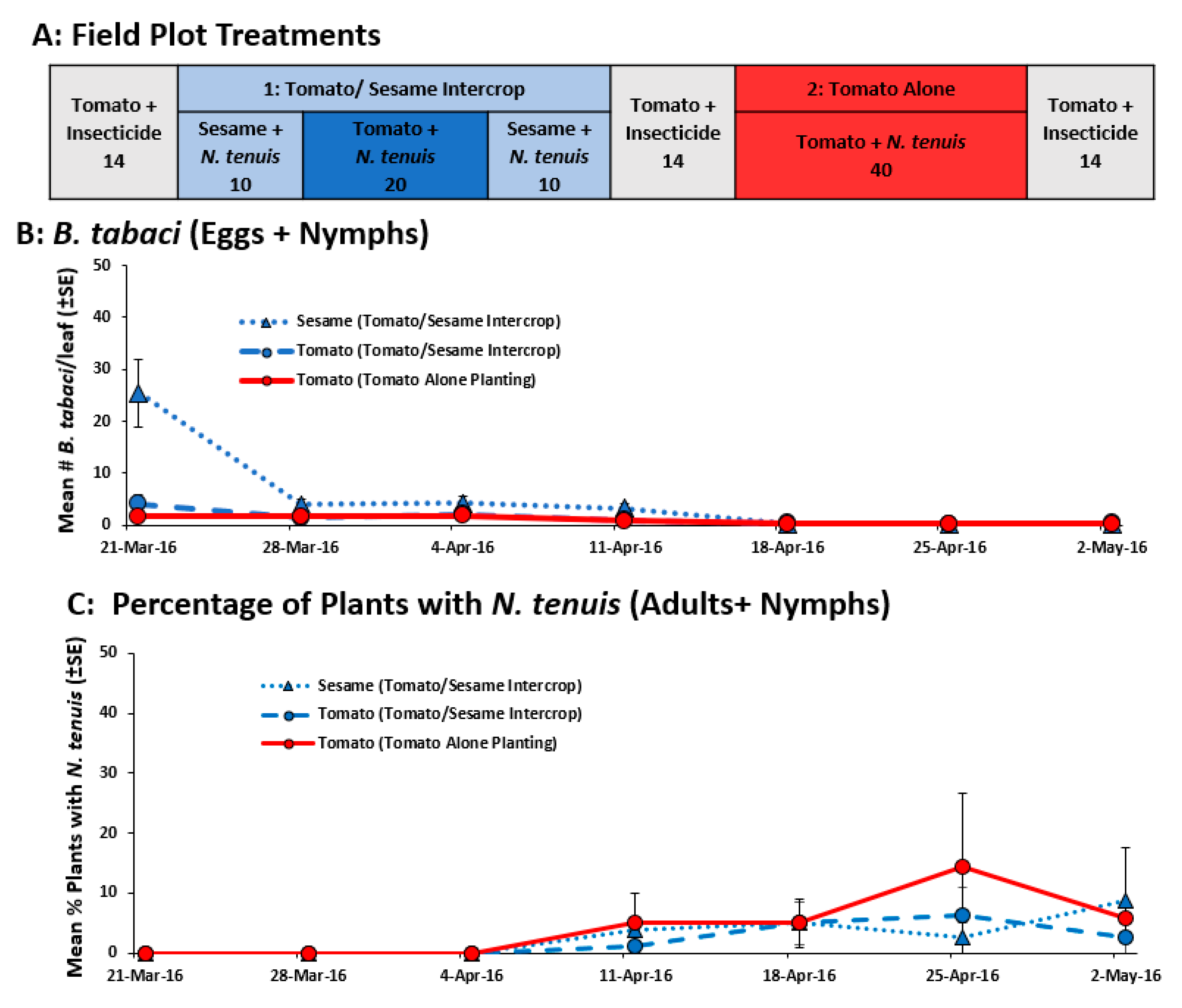
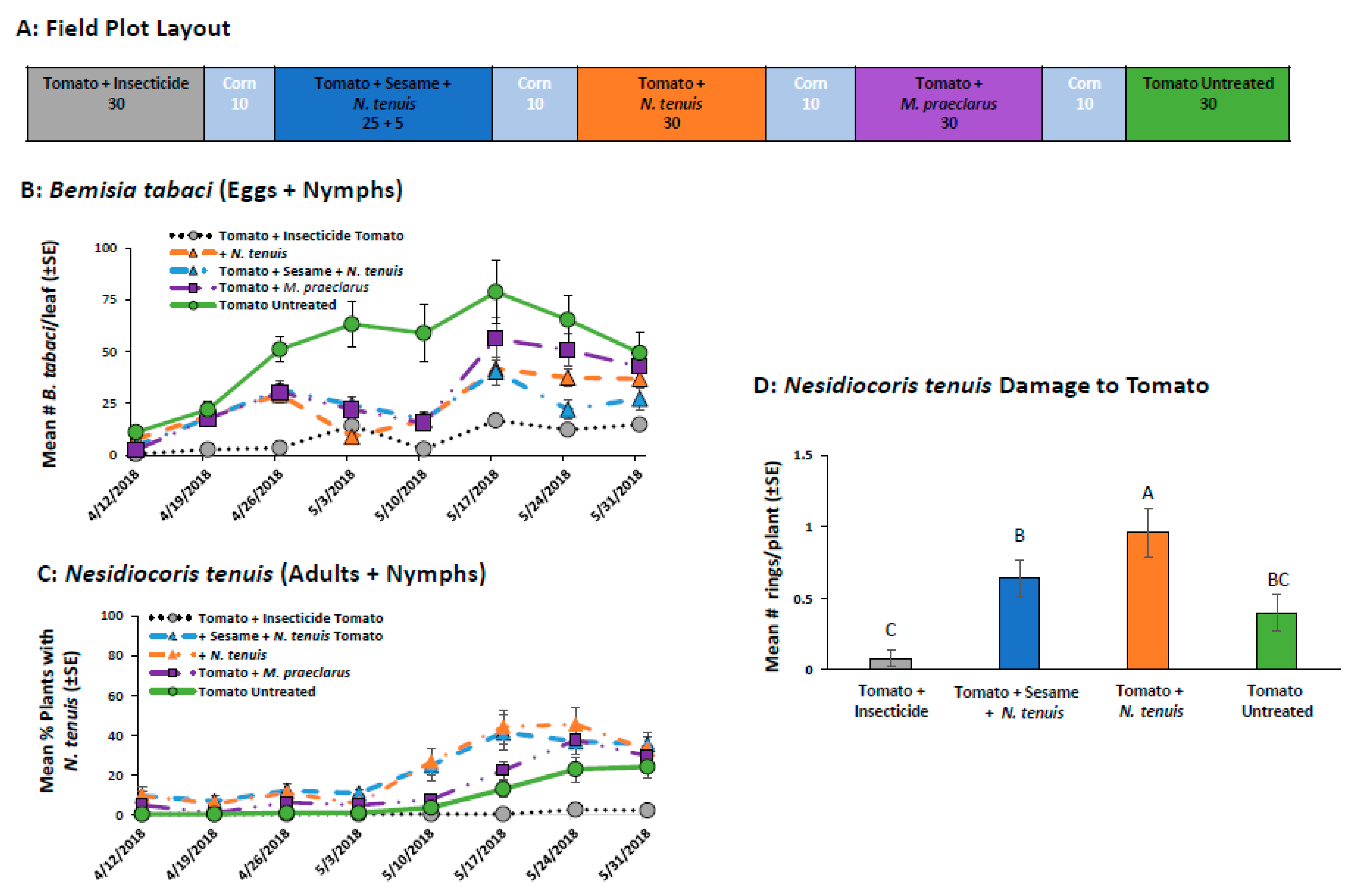
2.2.5. Field Comparison of Pesticide, M. praeclarus and N. tenuis to Control B. tabaci in Tomato Alone and Tomato-Sesame Intercrop
3. Discussion
4. Materials and Methods
4.1. Plants and Colonies
4.2. Field Plot Preparation
4.3. Comparison of N. tenuis and Pesticides to Control Whiteflies
4.3.1. Pre-Plant Inoculation of Tomato to Establish N. tenuis in Open-Field Tomatoes
4.3.2. Cage Evaluation of Pesticide and N. tenuis to Control B. tabaci
4.4. Tomato-Sesame Intercrop to Retain Zoophyatophagus Mirids
4.4.1. Field Evaluation of Sesame-Tomato Interplanting to Retain N. tenuis
4.4.2. Field Evaluation of Reduced Amount of Sesame in Tomato-Sesame Interplanting
4.4.3. Field Comparison of N. tenuis and Pesticide to Control B. tabaci in a Tomato Boarded by Sesame
4.4.4. Field Evaluation of M. praeclarus in a Tomato-Sesame Intercrop
4.4.5. Field Comparison of Pesticide, M. praeclarus and N. tenuis to Control B. tabaci in Tomato Alone and Tomato-Sesame Intercrop
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Research and Markets World-Tomato—Market Analysis, Forecast, Size, Trends and Insights. Available online: https://www.researchandmarkets.com/reports/4701312/world-tomato-market-analysis-forecast-size (accessed on 12 July 2022).
- Desneux, N.; Han, P.; Mansour, R.; Arnó, J.; Brévault, T.; Campos, M.R.; Chailleux, A.; Guedes, R.N.C.; Karimi, J.; Konan, K.A.J.; et al. Integrated Pest Management of Tuta absoluta: Practical Implementations across Different World Regions. J. Pest Sci. 2022, 95, 17–39. [Google Scholar] [CrossRef]
- Stansly, P.A.; Naranjo, S.E. Bemisia: Bionomics and Management of a Global Pest; Stansly, P.A., Naranjo, S.E., Eds.; Springer: Dordrecht, The Netherlands, 2010; ISBN 978-90-481-2459-6. [Google Scholar]
- Desneux, N.; Wajnberg, E.; Wyckhuys, K.A.G.; Burgio, G.; Arpaia, S.; Narváez-Vasquez, C.A.; González-Cabrera, J.; Catalán Ruescas, D.; Tabone, E.; Frandon, J.; et al. Biological Invasion of European Tomato Crops by Tuta absoluta: Ecology, Geographic Expansion and Prospects for Biological Control. J. Pest Sci. 2010, 83, 197–215. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The Sublethal Effects of Pesticides on Beneficial Arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Hemingway, J.; Field, L.; Vontas, J. An Overview of Insecticide Resistance. Science 2002, 298, 96–97. [Google Scholar] [CrossRef]
- Nicolopoulou-Stamati, P.; Maipas, S.; Kotampasi, C.; Stamatis, P.; Hens, L. Chemical Pesticides and Human Health: The Urgent Need for a New Concept in Agriculture. Front. Public Health 2016, 4, 148. [Google Scholar] [CrossRef] [Green Version]
- Calvo-Agudo, M.; González-Cabrera, J.; Picó, Y.; Calatayud-Vernich, P.; Urbaneja, A.; Dicke, M.; Tena, A. Neonicotinoids in Excretion Product of Phloem-Feeding Insects Kill Beneficial Insects. Proc. Natl. Acad. Sci. USA 2019, 116, 16817–16822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tortell, P.D. Earth 2020: Science, Society, and Sustainability in the Anthropocene. Proc. Natl. Acad. Sci. USA 2020, 117, 8683–8691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biondi, A.; Guedes, R.N.C.; Wan, F.; Desneux, N. Ecology, Worldwide Spread, and Management of the Invasive South American Tomato Pinworm, Tuta absoluta: Past, Present, and Future. Annu. Rev. Entomol. 2018, 63, 239–258. [Google Scholar] [CrossRef] [PubMed]
- Guedes, R.N.C.; Roditakis, E.; Campos, M.R.; Haddi, K.; Bielza, P.; Siqueira, H.A.A.; Tsagkarakou, A.; Vontas, J.; Nauen, R. Insecticide Resistance in the Tomato Pinworm Tuta absoluta: Patterns, Spread, Mechanisms, Management and Outlook. J. Pest Sci. 2019, 92, 1329–1342. [Google Scholar] [CrossRef]
- Urbaneja, A.; González-Cabrera, J.; Arnó, J.; Gabarra, R. Prospects for the Biological Control of Tuta absoluta in Tomatoes of the Mediterranean Basin. Pest Manag. Sci. 2012, 68, 1215–1222. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Hedo, M.; Riahi, C.; Urbaneja, A. Use of Zoophytophagous Mirid Bugs in Horticultural Crops: Current Challenges and Future Perspectives. Pest Manag. Sci. 2021, 77, 33–42. [Google Scholar] [CrossRef]
- van Lenteren, J.C.; Lanzoni, A.; Hemerik, L.; Bueno, V.H.P.; Bajonero Cuervo, J.G.; Biondi, A.; Burgio, G.; Calvo, F.J.; de Jong, P.W.; López, S.N.; et al. The Pest Kill Rate of Thirteen Natural Enemies as Aggregate Evaluation Criterion of Their Biological Control Potential of Tuta absoluta. Sci. Rep. 2021, 11, 10756. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Urbaneja-Bernat, P.; Jaques, J.A.; Flors, V.; Urbaneja, A. Defensive Plant Responses Induced by Nesidiocoris tenuis (Hemiptera: Miridae) on Tomato Plants. J. Pest Sci. 2015, 88, 543–554. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Hedo, M.; Bouagga, S.; Jaques, J.A.; Flors, V.; Urbaneja, A. Tomato Plant Responses to Feeding Behavior of Three Zoophytophagous Predators (Hemiptera: Miridae). Biol. Control 2015, 86, 46–51. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Gallego, C.; Roda, A.; Kostyk, B.; Triana, M.; Alférez, F.; Stansly, P.A.; Qureshi, J.; Urbaneja, A. Biological Traits of the Predatory Mirid Macrolophus praeclarus, a Candidate Biocontrol Agent for the Neotropical Region. Bull. Entomol. Res. 2021, 111, 429–437. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Bouagga, S.; Zhang, N.X.; Moerkens, R.; Messelink, G.; Jaques, J.A.; Flors, V.; Broufas, G.; Urbaneja, A.; Pappas, M.L. Induction of Plant Defenses: The Added Value of Zoophytophagous Predators. J. Pest Sci. 2022, 95, 1501–1517. [Google Scholar] [CrossRef]
- Calvo, F.J.; Lorente, M.J.; Stansly, P.A.; Belda, J.E. Preplant Release of Nesidiocoris tenuis and Supplementary Tactics for Control of Tuta absoluta and Bemisa tabaci in Greenhouse Tomato. Entomol. Exp. Appl. 2012, 143, 111–119. [Google Scholar] [CrossRef]
- Calvo, F.J.; Bolckmans, K.; Belda, J.E. Release Rate for a Pre-Plant Application of Nesidiocoris tenuis for Bemisia tabaci Control in Tomato. BioControl 2012, 57, 809–817. [Google Scholar] [CrossRef]
- van Lenteren, J.C.; Van Alomar, O.; Ravensberg, W.J.; Urbaneja, A. Integrated Pest and Disease Management in Greenhouse Crops; Gullino, M.L., Albajes, R., Nicot, P.C., Eds.; Springer International Publishing: Cham, Switzerland, 2020; ISBN 978-3-030-22303-8. [Google Scholar]
- Mollá, O.; Gonzalez-Cabrera, J.; Urbaneja, A. The Combined Use of Bacillus thuringiensis and Nesidiocoris tenuis against the Tomato Borer Tuta absoluta. BioControl 2011, 56, 883–891. [Google Scholar] [CrossRef]
- Calvo, J.; Bolckmans, K.; Stansly, P.A.; Urbaneja, A. Predation by Nesidiocoris tenuis on Bemisia tabaci and Injury to Tomato. BioControl 2009, 54, 237–246. [Google Scholar] [CrossRef]
- van Lenteren, J.C.; Bolckmans, K.; Köhl, J.; Ravensberg, W.J.; Urbaneja, A. Biological Control Using Invertebrates and Microorganisms: Plenty of New Opportunities. BioControl 2018, 63, 39–59. [Google Scholar] [CrossRef] [Green Version]
- Chinchilla-Ramírez, M.; Garzo, E.; Fereres, A.; Gavara-Vidal, J.; ten Broeke, C.J.; van Loon, J.J.A.; Urbaneja, A.; Pérez-Hedo, M. Plant Feeding by Nesidiocoris tenuis: Quantifying Its Behavioral and Mechanical Components. Biol. Control 2021, 152, 104402. [Google Scholar] [CrossRef]
- Moerkens, R.; Pekas, A.; Bellinkx, S.; Hanssen, I.; Huysmans, M.; Bosmans, L.; Wäckers, F. Nesidiocoris tenuis as a Pest in Northwest Europe: Intervention Threshold and Influence of Pepino Mosaic Virus. J. Appl. Entomol. 2020, 144, 566–577. [Google Scholar] [CrossRef]
- Naselli, M.; Zappalà, L.; Gugliuzzo, A.; Tropea Garzia, G.; Biondi, A.; Rapisarda, C.; Cincotta, F.; Condurso, C.; Verzera, A.; Siscaro, G. Olfactory Response of the Zoophytophagous Mirid Nesidiocoris tenuis to Tomato and Alternative Host Plants. Arthropod-Plant Interact. 2017, 11, 121–131. [Google Scholar] [CrossRef]
- Biondi, A.; Zappalà, L.; Di Mauro, A.; Tropea Garzia, G.; Russo, A.; Desneux, N.; Siscaro, G. Can Alternative Host Plant and Prey Affect Phytophagy and Biological Control by the Zoophytophagous Mirid Nesidiocoris tenuis? BioControl 2016, 61, 79–90. [Google Scholar] [CrossRef]
- Urbaneja-Bernat, P.; Bru, P.; González-Cabrera, J.; Urbaneja, A.; Tena, A. Reduced Phytophagy in Sugar-Provisioned Mirids. J. Pest Sci. 2019, 92, 1139–1148. [Google Scholar] [CrossRef]
- Biondi, A.; Zappala, L.; Stark, J.D.; Desneux, N. Do Biopesticides Affect the Demographic Traits of a Parasitoid Wasp and Its Biocontrol Services through Sublethal Effects? PLoS ONE 2013, 8, e76548. [Google Scholar] [CrossRef] [Green Version]
- Konan, K.A.J.; Monticelli, L.S.; Ouali-N’goran, S.W.M.; Ramirez-Romero, R.; Martin, T.; Desneux, N. Combination of Generalist Predators, Nesidiocoris tenuis and Macrolophus pygmaeus, with a Companion Plant, Sesamum indicum: What Benefit for Biological Control of Tuta absoluta? PLoS ONE 2021, 16, e0257925. [Google Scholar] [CrossRef]
- Roda, A.; Castillo, J.; Allen, C.; Urbaneja, A.; Pérez-Hedo, M.; Weihman, S.; Stansly, P.A. Biological Control Potential and Drawbacks of Three Zoophytophagous Mirid Predators against Bemisia tabaci in the United States. Insects 2020, 11, 670. [Google Scholar] [CrossRef]
- Abraços-Duarte, G.; Ramos, S.; Valente, F.; Borges da Silva, E.; Figueiredo, E. Functional Response and Predation Rate of Dicyphus cerastii Wagner (Hemiptera: Miridae). Insects 2021, 12, 530. [Google Scholar] [CrossRef]
- Smith, H.A.; Krey, K.L. Three Release Rates of Dicyphus hesperus (Hemiptera: Miridae) for Management of Bemisia tabaci (Hemiptera: Aleyrodidae) on Greenhouse Tomato. Insects 2019, 10, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, H.A.; Nagle, C.A.; MacVean, C.M.; Vallad, G.E.; van Santen, E.; Hutton, S.F. Comparing Host Plant Resistance, Repellent Mulches, and At-Plant Insecticides for Management of Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae) and Tomato Yellow Leaf Curl Virus. J. Econ. Entomol. 2019, 112, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Johnston, N.; Paris, T.; Paret, M.L.; Freeman, J.; Martini, X. Repelling Whitefly (Bemisia tabaci) Using Limonene-Scented Kaolin: A Novel Pest Management Strategy. Crop Prot. 2022, 154, 105905. [Google Scholar] [CrossRef]
- Castillo, J.A.; Stansly, P.A. Biology of Eretmocerus sudanensis n. sp. Zolnerowich and Rose, Parasitoid of Bemisia tabaci Gennadius. BioControl 2011, 56, 843–850. [Google Scholar] [CrossRef] [Green Version]
- The Florida Tomato Committee Tomato 101 Fact Sheet. Available online: https://www.floridatomatoes.org/tomato-101/ (accessed on 12 February 2022).
- Smith, H.A. Biopesticides for Management of Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae) and Tomato Yellow Leaf Curl Virus. J. Econ. Entomol. 2020, 113, 2310–2318. [Google Scholar] [CrossRef]
- Nakaishi, K.; Fukui, Y.; Arakawa, R. Reproduction of Nesidiocoris tenuis (Reuter) on Sesame. Jpn. J. Appl. Entomol. Zool. 2011, 55, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Arnó, J.; Castañé, C.; Riudavets, J.; Gabarra, R. Risk of Damage to Tomato Crops by the Generalist Zoophytophagous Predator Nesidiocoris tenuis (Reuter) (Hemiptera: Miridae). Bull. Entomol. Res. 2010, 100, 105–115. [Google Scholar] [CrossRef]
- Rozenberg, T.; Shaltiel-Harpaz, L.; Coll, M. Visualizing Eggs of Nesidiocoris tenuis (Heteroptera: Miridae) Embedded in Tomato Plant Tissues. Entomol. Sci. 2015, 18, 400–402. [Google Scholar] [CrossRef]
- Shaltiel-Harpaz, L.; Gerling, D.; Graph, S.; Kedoshim, H.; Azolay, L.; Rozenberg, T.; Nachache, Y.; Steinberg, S.; Allouche, A.; Alon, T. Control of the Tomato Leafminer, Tuta absoluta (Lepidoptera: Gelechiidae), in Open-Field Tomatoes by Indigenous Natural Enemies Occurring in Israel. J. Econ. Entomol. 2016, 109, 120–131. [Google Scholar] [CrossRef]
- Bompard, A.; Jaworski, C.C.; Bearez, P.; Desneux, N. Sharing a Predator: Can an Invasive Alien Pest Affect the Predation on a Local Pest? Popul. Ecol. 2013, 55, 433–440. [Google Scholar] [CrossRef]
- Moerkens, R.; Janssen, D.; Brenard, N.; Reybroeck, E.; del Mar Tellez, M.; Rodríguez, E.; Bosmans, L.; Leirs, H.; Sluydts, V. Simplified Modelling Enhances Biocontrol Decision Making in Tomato Greenhouses for Three Important Pest Species. J. Pest Sci. 2021, 94, 285–295. [Google Scholar] [CrossRef]
- Dilipsundar, N.; Chitra, N.; Gowtham, V. Checklist of Insect Pests of Sesame. Indian J. Entomol. 2019, 81, 928. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Urbaneja, A. The Zoophytophagous Predator Nesidiocoris tenuis: A Successful but Controversial Biocontrol Agent in Tomato Crops. In Advances in Insect Control and Resistance Management; Horowitz, A.R., Ishaaya, I., Eds.; Springer International Publishing: Dordrecht, The Netherlands, 2016; pp. 121–138. ISBN 978-3-319-31798-4. [Google Scholar]
- Molla, O.; Urbaneja, A.; Arnó, J.; Gabarra, R. Preferencia de Presa Por Nesidiocoris tenuis Reuter (Hemiptera: Miridae) Cuando Tuta Absoluta (Meyrick) (Lepidoptera: Gelechiidae) Coexiste Con Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). Phtyoma España 2011, 235, 48–50. [Google Scholar]
- Moayeri, H.R.S.; Ashouri, A.; Brødsgaard, H.F.; Enkegaard, A. Odour-Mediated Preference and Prey Preference of Macrolophus caliginosus between Spider Mites and Green Peach Aphids. J. Appl. Entomol. 2006, 130, 504–508. [Google Scholar] [CrossRef]
- Jaworski, C.C.; Bompard, A.; Genies, L.; Amiens-Desneux, E.; Desneux, N. Preference and Prey Switching in a Generalist Predator Attacking Local and Invasive Alien Pests. PLoS ONE 2013, 8, e82231. [Google Scholar] [CrossRef]
- Urbaneja-Bernat, P.; Mollá, O.; Alonso, M.; Bolkcmans, K.; Urbaneja, A.; Tena, A. Sugars as Complementary Alternative Food for the Establishment of Nesidiocoris tenuis in Greenhouse Tomato. J. Appl. Entomol. 2015, 139, 161–167. [Google Scholar] [CrossRef]
- Messelink, G.J.; Bennison, J.; Alomar, O.; Ingegno, B.L.; Tavella, L.; Shipp, L.; Palevsky, E.; Wäckers, F.L. Approaches to Conserving Natural Enemy Populations in Greenhouse Crops: Current Methods and Future Prospects. BioControl 2014, 59, 377–393. [Google Scholar] [CrossRef]
- Bresch, C.; Ottenwalder, L.; Poncet, C.; Parolin, P. Tobacco as Banker Plant for Macrolophus pygmaeus to Control Trialeurodes Vaporariorum in Tomato Crops. Univers. J. Agric. Res. 2014, 2, 297–304. [Google Scholar] [CrossRef]
- Ozores-Hampton, M.; McAvoy, G. Tomato Varieties for Florida—Florida “Red Rounds,” Plums, Cherries, Grapes, and Heirlooms. HS1189. Horticultural Sciences Department, UF/IFAS Extension. Available online: https://edis.ifas.ufl.edu/pdf%5Carchived%5CHS%5CHS1189%5CHS1189-14561040.pdf (accessed on 30 September 2022).






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castillo, J.; Roda, A.; Qureshi, J.; Pérez-Hedo, M.; Urbaneja, A.; Stansly, P. Sesame as an Alternative Host Plant to Establish and Retain Predatory Mirids in Open-Field Tomatoes. Plants 2022, 11, 2779. https://doi.org/10.3390/plants11202779
Castillo J, Roda A, Qureshi J, Pérez-Hedo M, Urbaneja A, Stansly P. Sesame as an Alternative Host Plant to Establish and Retain Predatory Mirids in Open-Field Tomatoes. Plants. 2022; 11(20):2779. https://doi.org/10.3390/plants11202779
Chicago/Turabian StyleCastillo, Jose, Amy Roda, Jawwad Qureshi, Meritxell Pérez-Hedo, Alberto Urbaneja, and Philip Stansly. 2022. "Sesame as an Alternative Host Plant to Establish and Retain Predatory Mirids in Open-Field Tomatoes" Plants 11, no. 20: 2779. https://doi.org/10.3390/plants11202779
APA StyleCastillo, J., Roda, A., Qureshi, J., Pérez-Hedo, M., Urbaneja, A., & Stansly, P. (2022). Sesame as an Alternative Host Plant to Establish and Retain Predatory Mirids in Open-Field Tomatoes. Plants, 11(20), 2779. https://doi.org/10.3390/plants11202779