
Trehalose-Induced Regulations in Nutrient Status and Secondary Metabolites of Drought-Stressed Sunflower (Helianthus annuus L.) Plants

 ,
,  and
and 
Abstract
:1. Introduction
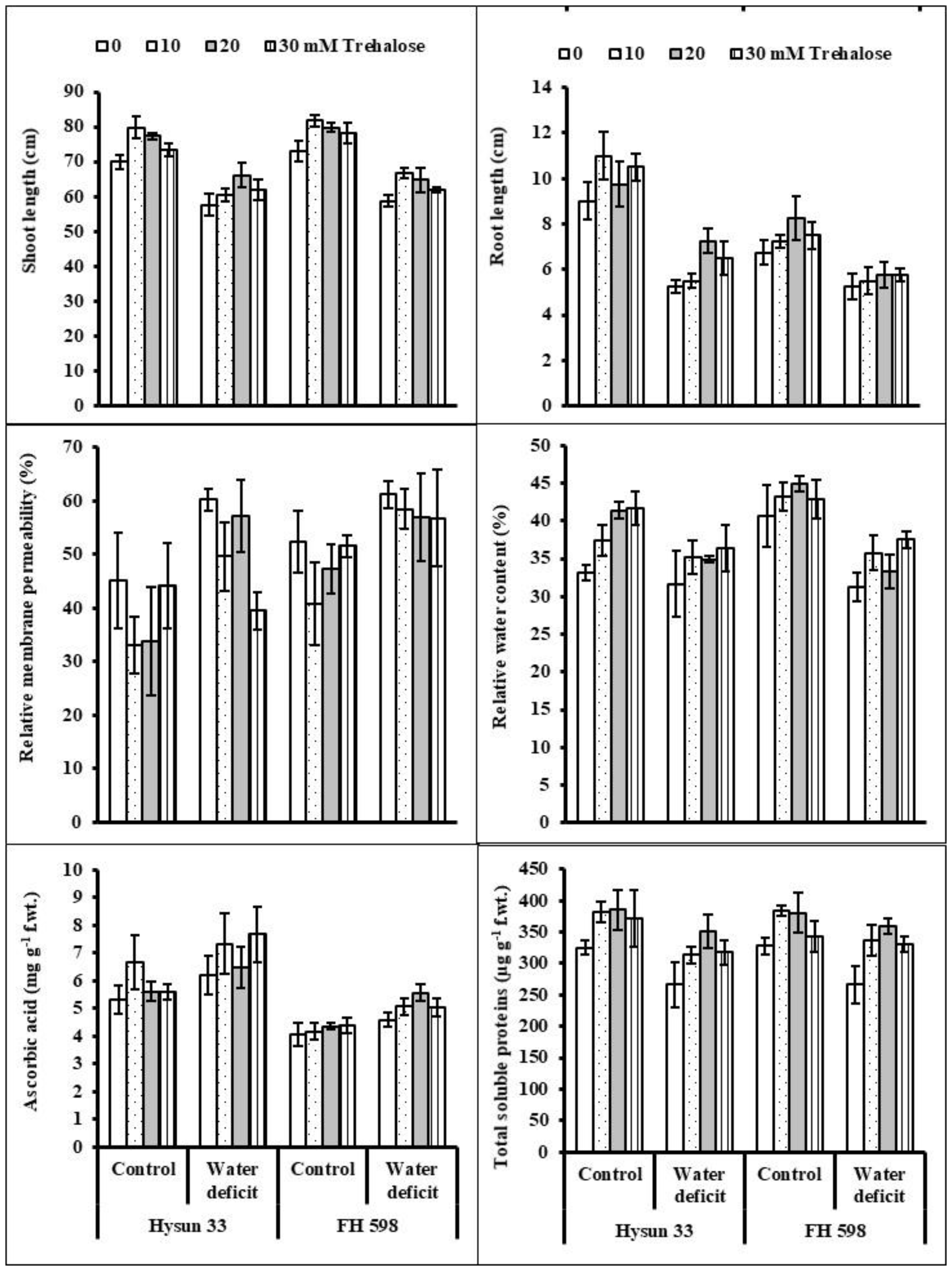
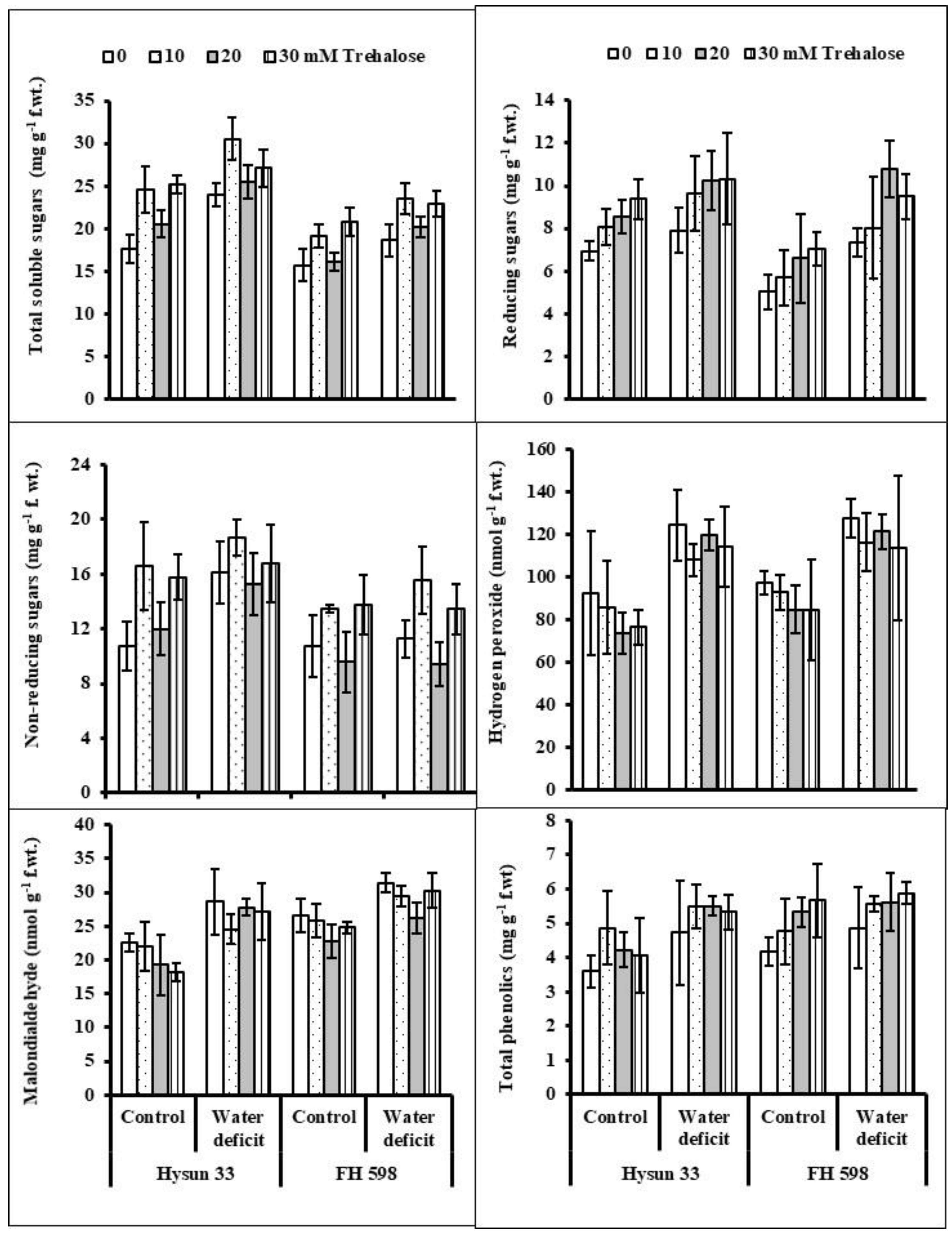
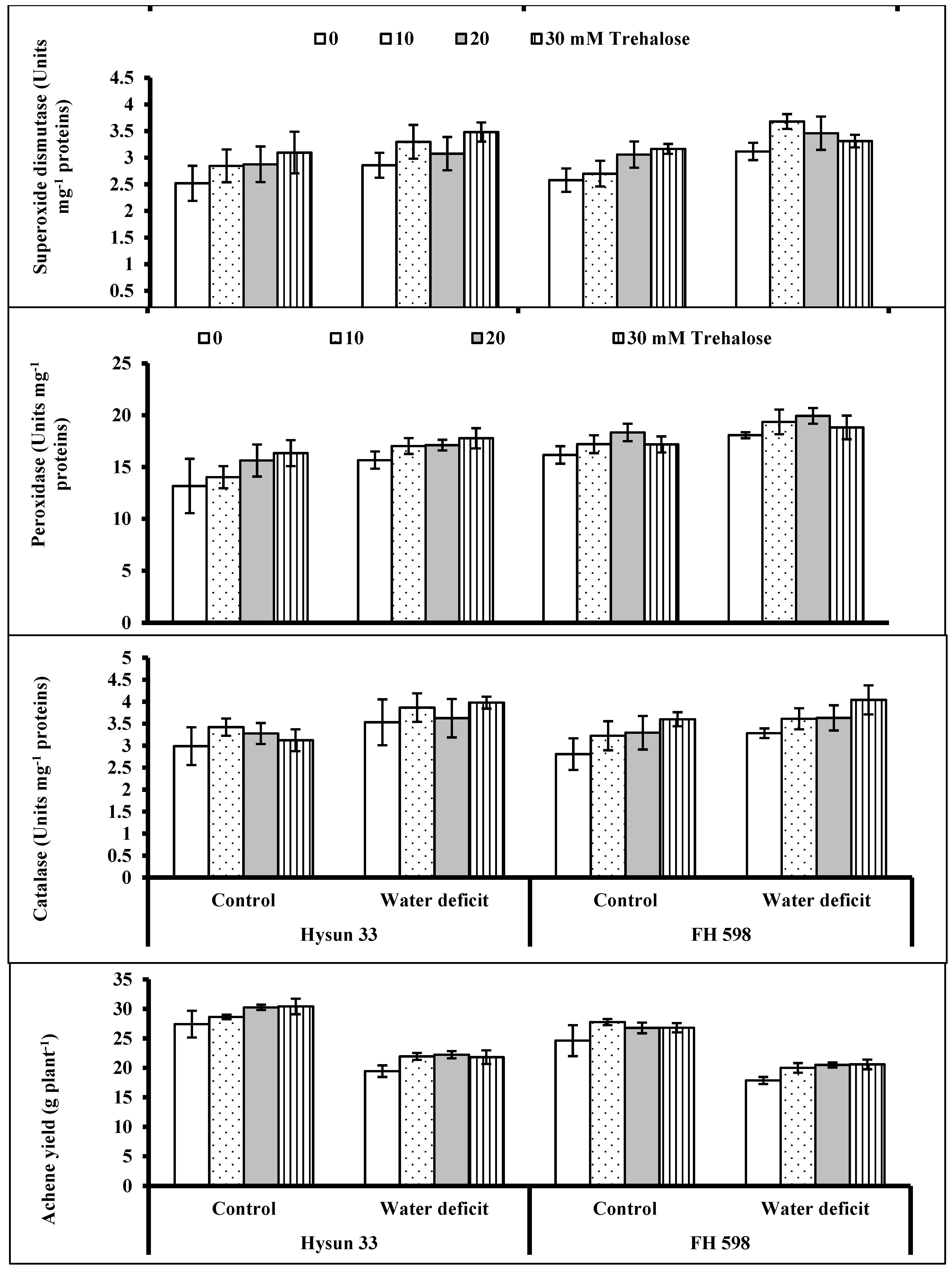
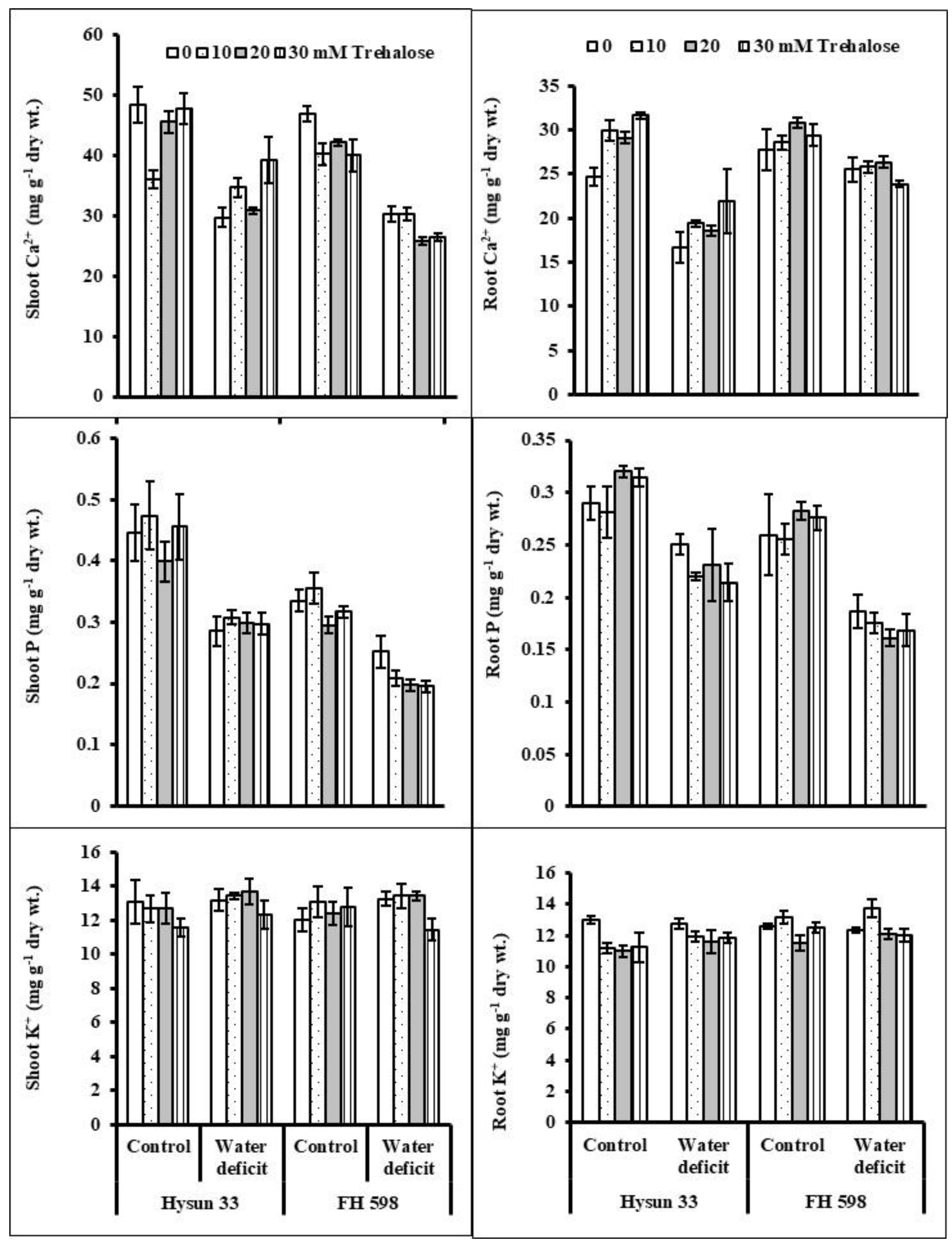
2. Results
3. Discussion
4. Materials and Methods
4.1. Shoot and Root Length
4.2. Relative Membrane Permeability (RMP)
4.3. Relative Water Content (%)
4.4. Ascorbic Acid (AsA)
4.5. Sugar Content
4.6. Hydrogen Peroxide (H2O2)
4.7. Malondialdehyde (MDA)
4.8. Total Phenolics
4.9. Total Soluble Proteins
4.10. Estimation of Antioxidant Enzymes
4.11. Inorganic Nutrients
4.12. Determination of Potassium (K+) and Calcium (Ca2+)
4.13. Determination of Phosphorus (P)
4.14. Achene Yield
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hasan, M.M.U.; Ma, F.; Prodhan, Z.H.; Li, F.; Shen, H.; Chen, Y.; Wang, X. Molecular and physio-biochemical characterization of cotton species for assessing drought stress tolerance. Int. J. Mol. Sci. 2018, 19, 2636. [Google Scholar] [CrossRef] [Green Version]
- Dawood, M.F.A.; Zaid, A.; Abdel Latef, A.A.H.A. Salicylic acid spraying-induced resilience strategies against the damaging impacts of drought and/or salinity stress in two varieties of Vicia faba L. seedlings. J. Plant Growth Regul. 2022, 41, 1919–1942. [Google Scholar] [CrossRef]
- Ashraf, M. Inducing drought tolerance in plants: Recent advances. Biotechnol. Adv. 2010, 28, 169–183. [Google Scholar] [CrossRef]
- Ashraf, M.; Akram, N.A.; Al-Qurainy, F. Drought tolerance: Roles of organic osmolytes, growth regulators and mineral nutrients. Adv. Agron. 2011, 8, 85–96. [Google Scholar]
- Sadiq, M.; Akram, N.A.; Ashraf, M.; Al-Qurainy, F.; Ahmad, P. Alpha-tocopherol-induced regulation of growth and metabolism in plants under non-stress and stress conditions. J. Plant Growth Regul. 2019, 38, 1325–1340. [Google Scholar] [CrossRef]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The impact of drought in plant metabolism: How to exploit tolerance mechanisms to increase crop production. Appl. Sci. 2020, 10, 5692. [Google Scholar] [CrossRef]
- Singh, A.; Sharma, M.K.; Sengar, R.S. Osmolytes: Proline metabolism in plants as sensors of abiotic stress. J. Nat. Appl. Sci. 2017, 9, 2079–2092. [Google Scholar] [CrossRef]
- Shafiq, F.; Iqbal, M.; Ali, M.; Ashraf, M.A. Seed pre-treatment with polyhydroxy fullerene nanoparticles confer salt tolerance in wheat through upregulation of H2O2 neutralizing enzymes and phosphorus uptake. Soil Sci. Plant Nutr. 2019, 19, 734–742. [Google Scholar] [CrossRef]
- Kosar, F.; Akram, N.A.; Ashraf, M.; Ahmad, A.; Alyemeni, M.N.; Ahmad, P. Impact of exogenously applied trehalose on leaf biochemistry, achene yield and oil composition of sunflower under drought stress. Physiol. Plant. 2020, 172, 317–333. [Google Scholar] [CrossRef]
- Wang, X.; Cai, X.; Xu, C.; Wang, Q.; Dai, S. Drought-responsive mechanisms in plant leaves revealed by proteomics. Int. J. Mol. Sci. 2016, 17, 1706. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Li, F.; Wang, G.P.; Yang, X.H.; Wang, W. Exogenously-supplied trehalose protects thylakoid membranes of winter wheat from heat-induced damage. Plant Biol. 2010, 54, 495–501. [Google Scholar] [CrossRef]
- Feng, Y.; Chen, X.; He, Y.; Kou, X.; Xue, Z. Effects of exogenous trehalose on the metabolism of sugar and abscisic acid in tomato seedlings under salt stress. Trans. Tianjin Univ. 2019, 25, 451–471. [Google Scholar] [CrossRef] [Green Version]
- Ismail, A.I.; Arafat, S.M. Quality characteristics of high-oleic sunflower oil extracted from some hybrids cultivated under Egyptian conditions. J. Food Technol. 2014, 1, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Kirsh, V.A.; Hayes, R.B.; Mayne, S.T.; Chatterjee, N.; Subar, A.F.; Dixon, L.B.; Albanes, D.; Andriole, G.L.; Urban, D.A.; Peters, U. Supplemental and dietary vitamin E, β-carotene, and vitamin C intakes and prostate cancer risk. J. Natl. Cancer Inst. 2006, 98, 245–254. [Google Scholar] [CrossRef]
- Kosar, F.; Akram, N.A.; Sadiq, M.; Al-Qurainy, F.; Ashraf, M. Trehalose: A key organic osmolyte effectively involved in plant abiotic stress tolerance. J. Plant Growth Regul. 2019, 38, 606–618. [Google Scholar] [CrossRef]
- Sadiq, M.; Akram, N.A.; Ashraf, M.; Ali, S. Tocopherol confers water stress tolerance: Sugar and osmoprotectant metabolism in mung bean [Vigna radiata (L.) Wilczek]. Agrochimica 2017, 61, 28–42. [Google Scholar]
- Shafiq, S.; Akram, N.A.; Ashraf, M. Does exogenously-applied trehalose alter oxidative defense system in the edible part of radish (Raphanus sativus L.) under water-deficit conditions? Sci. Hortic. 2015, 185, 68–75. [Google Scholar] [CrossRef]
- Kosar, F.; Akram, N.A.; Ashraf, M.; Sadiq, M.; Al-Qurainy, F. Trehalose-induced improvement in growth, photosynthetic characteristics and levels of some key osmoprotectants in sunflower (Helianthus annuus L.) under drought stress. Pak. J. Bot. 2018, 50, 955–961. [Google Scholar]
- Ali, Q.; Ashraf, M. Induction of drought tolerance in maize (Zea mays L.) due to exogenous application of trehalose: Growth, photosynthesis, water relations and oxidative defense mechanism. J. Agron. Crop Sci. 2011, 197, 258–271. [Google Scholar] [CrossRef]
- Räsänen, L.A.; Saijets, S.; Jokinen, K.; Lindström, K. Evaluation of the roles of two compatible solutes, glycine betaine and trehalose, for the Acacia senegal–Sinorhizobium symbiosis exposed to drought stress. Plant Soil 2004, 260, 237–251. [Google Scholar] [CrossRef]
- Quilambo, O.A. Proline content, water retention capability and cell membrane integrity as parameters for drought tolerance in two peanut cultivars. S. Afr. J. Bot. 2004, 70, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Akram, N.A.; Waseem, M.; Ameen, R.; Ashraf, M. Trehalose pretreatment induces drought tolerance in radish (Raphanus sativus L.) plants: Some key physio-biochemical traits. Acta Physiol. Plant. 2016, 38, 1–10. [Google Scholar] [CrossRef]
- Wang, W.B.; Kim, Y.H.; Lee, H.S.; Kim, K.Y.; Deng, X.P.; Kwak, S.S. Analysis of antioxidant enzyme activity during germination of alfalfa under salt and drought stresses. Plant Physiol. Biochem. 2009, 47, 570–577. [Google Scholar] [CrossRef]
- Rapacz, M.; Kościelniak, J.; Jurczyk, B.; Adamska, A.; Wójcik, M. Different patterns of physiological and molecular response to drought in seedlings of malt-and feed-type barleys (Hordeum vulgare). J. Agron. Crop Sci. 2010, 196, 9–19. [Google Scholar] [CrossRef]
- Zeid, I.M. Trehalose as osmoprotectant for maize under salinity-induced stress. J. Agric. Biol. Sci. 2009, 5, 613–622. [Google Scholar]
- Kaya, C.; Tuna, A.L.; Ashraf, M.; Altunlu, H. Improved salt tolerance of melon (Cucumis melo L.) by the addition of proline and potassium nitrate. Environ. Exp. Bot. 2007, 60, 397–403. [Google Scholar] [CrossRef]
- Seminario, A.; Song, L.; Zulet, A.; González, E.M.; Larrainzar, E. Drought stress causes a reduction in the biosynthesis of ascorbic acid in soybean plants. Front. Plant Sci. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.M.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Trehalose-induced drought stress tolerance: A comparative study among different Brassica species. Plant Omics 2014, 7, 271–283. [Google Scholar]
- Bano, S.; Ashraf, M.; Akram, N.A. Salt stress regulates enzymatic and non-enzymatic antioxidative defense system in the edible part of carrot (Daucus carota L.). J. Plant Interact. 2014, 9, 324–329. [Google Scholar] [CrossRef]
- Mahmood, T.; Khalid, S.; Abdullah, M.; Ahmad, Z.; Shah, M.K.N.; Ghafoor, A.; Du, X. Insights into drought stress signaling in plants and the molecular genetic basis of cotton drought tolerance. Cells 2020, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- Aldesuquy, H.; Ghanem, H. Exogenous salicylic acid and trehalose ameliorate short term drought stress in wheat cultivars by up-regulating membrane characteristics and antioxidant defense system. J. Hortic. 2015, 2, 1–10. [Google Scholar] [CrossRef]
- Ibrahim, H.A.; Abdellatif, Y.M. Effect of maltose and trehalose on growth, yield and some biochemical components of wheat plant under water stress. Ann. Agric. Sci. 2016, 61, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Gurrieri, L.; Merico, M.; Trost, P.; Forlani, G.; Sparla, F. Impact of drought on soluble sugars and free proline content in selected Arabidopsis mutants. Biology 2020, 9, 367. [Google Scholar] [CrossRef] [PubMed]
- Rodziewicz, P.; Swarcewicz, B.; Chmielewska, K.; Wojakowska, A.; Stobiecki, M. Influence of abiotic stresses on plant proteome and metabolome changes. Acta Physiol. Plant. 2014, 36, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Yang, Y.; Li, C.; Huang, D.; Zhang, J.; Wang, C.; Li, W.; Wang, N.; Deng, Y.; Liao, W. Research progress on the functions of gasotransmitters in plant responses to abiotic stresses. Plants 2019, 8, 605. [Google Scholar] [CrossRef] [PubMed]
- Ali, Q.; Shahid, S.; Ali, S.; Javed, M.T.; Iqbal, N.; Habib, N.; Hussain, S.M.; Shahid, S.A.; Noreen, Z.; Hussain, A.I.; et al. Trehalose Metabolism in Plants under Abiotic Stresses; CRC Press: Boca Raton, FL, USA, 2019; pp. 349–364. [Google Scholar]
- Rohman, M.; Islam, M.; Monsur, M.B.; Amiruzzaman, M.; Fujita, M.; Hasanuzzaman, M. Trehalose protects maize plants from salt stress and phosphorus deficiency. Plants 2019, 8, 568. [Google Scholar] [CrossRef] [Green Version]
- Darvishan, M.; Tohidi-Moghadam, H.R.; Zahedi, H. The effects of foliar application of ascorbic acid (vitamin C) on physiological and biochemical changes of corn (Zea mays L) under irrigation withholding in different growth stages. Maydica 2013, 58, 195–200. [Google Scholar]
- Selote, D.S.; Khanna-Chopra, R. Antioxidant response of wheat roots to drought acclimation. Protoplasma 2010, 245, 153–163. [Google Scholar] [CrossRef]
- Mishra, A.K.; Rai, R.; Agrawal, S.B. Individual and interactive effects of elevated carbon dioxide and ozone on tropical wheat (Triticum aestivum L.) cultivars with special emphasis on ROS generation and activation of antioxidant defense system. Indian J. Biochem. Biophys. 2013, 50, 139–149. [Google Scholar]
- Waraich, E.A.; Ahmad, R.; Ashraf, M.Y. Role of mineral nutrition in alleviation of drought stress in plants. Aust. J. Crop Sci. 2011, 5, 764–777. [Google Scholar]
- Nounjan, N.; Nghia, P.T.; Theerakulpisut, P. Exogenous proline and trehalose promote recovery of rice seedlings from salt-stress and differentially modulate antioxidant enzymes and expression of related genes. J. Plant Physiol. 2012, 169, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Orooj, A. Salt stress effects on growth, ion accumulation and seed oil concentration in an arid zone traditional medicinal plant ajwain (Trachyspermum ammi [L.] Sprague). J. Arid Environ. 2006, 64, 209–220. [Google Scholar] [CrossRef]
- Shahbaz, M.; Ashraf, M. Does exogenous application of 24-epibrassinolide ameliorate salt induced growth inhibition in wheat (Triticum aestivum L.)? Plant Growth Regul. 2008, 55, 51–64. [Google Scholar] [CrossRef]
- Yang, G.; Rhodes, D.; Joly, R.J. Effect of high temperature on membrane stability and chlorophyll fluorescence in glycinebetaine-containing maize lines. Aust. J. Plant Physiol. 1996, 23, 431–443. [Google Scholar]
- Barrs, H.D.; Weatherley, P.E. A re-examination of the relative turgidity techniques for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar]
- Nelson, N.A. Photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem. 1944, 153, 375–380. [Google Scholar]
- Loomis, W.E.; Shull, C.A. Methods in Plant Physiology; McGraw-Hill: New York, NY, USA, 1937. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant system in acid rain treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar]
- Cakmak, I.; Horst, W.J. Effect of aluminium on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max). Physiol. Plant. 1991, 83, 463–468. [Google Scholar]
- Julkenen-Titto, R. Phenolic constituents in the leaves of northern willows: Methods for the analysis of certain phenolics. J. Agric. Food Chem. 1985, 33, 213–217. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Van Rossum, F.; Vekemans, X.; Meerts, P.; Gratia, E.; Lefèbvre, C. Allozyme variation in relation to ecotypic differentiation and population size in marginal populations of Silene nutans. J. Hered. 1997, 78, 552–560. [Google Scholar] [CrossRef]
- Chance, B.; Maehley, A.C. Assay of catalase and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Luck, H. Estimation of Catalase, Methods in Enzymatic Analysis; Academic Press: New York, NY, USA, 1974. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis; Prentice Hall: Englewood Cliffs, NJ, USA, 1962. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | df | Shoot Length | Root Length | RMP | RWC | Ascorbic Acid | Total Soluble Proteins |
| Drought (D) | 1 | 3291 *** | 135.1 *** | 2105 *** | 611.6 *** | 15.35 *** | 3205 *** |
| Trehalose (Tre) | 3 | 190.7 *** | 4.348 * | 244.1 ns | 89.68 ** | 1.821 ns | 1570 *** |
| Cultivars (Cvs) | 1 | 83.26 * | 40.64 *** | 981.9 ** | 78.48 * | 46.47 *** | 53.93 ns |
| D × Tre | 3 | 14.18 ns | 1.015 ns | 240.9 ns | 15.03 ns | 0.367 ns | 1139 ns |
| D × Cvs | 1 | 9.765 ns | 17.01 *** | 20.55 ns | 85.27 * | 0.389 ns | 1334 ns |
| Tre × Cvs | 3 | 8.807 ns | 0.515 ns | 48.35 ns | 7.092 ns | 1.276 ns | 281.2 ns |
| D × Tre × Cvs | 3 | 16.30 ns | 2.390 ns | 99.87 ns | 10.98 ns | 0.648 ns | 363.3 ns |
| Total Soluble Sugars | Reducing Sugars | Non-Reducing Sugars | Hydrogen Peroxide | MDA | Total Phenolics | ||
| Drought (D) | 1 | 266.5 *** | 67.85 *** | 3794 *** | 1649 *** | 470.5 *** | 9.824 * |
| Trehalose (Tre) | 3 | 111.9 *** | 18.80 * | 489.3 * | 531.3 ns | 29.53 ns | 2.872 ns |
| Cultivars (Cvs) | 1 | 359.1 *** | 30.08 * | 250.2 ns | 466.8 ns | 185.0** | 4.087 ns |
| D × Tre | 3 | 7.674 ns | 1.466 ns | 247.7 ns | 232.0 ns | 12.26 ns | 0.0288 ns |
| D × Cvs | 1 | 8.265 ns | 9.292 ns | 822.5 * | 81.78 ns | 18.55 ns | 1.329 ns |
| Tre × Cvs | 3 | 4.730 ns | 1.166 ns | 119.4 ns | 14.07 ns | 11.85 ns | 0.8396 ns |
| D × Tre × Cvs | 3 | 2.401 ns | 0.537 ns | 188.6 ns | 26.96 ns | 7.537 ns | 0.315 ns |
| Superoxide Dismutase | Peroxidase | Catalase | |||||
| Drought (D) | 1 | 2.964 *** | 61.67 *** | 3.672 *** | |||
| Trehalose (Tre) | 3 | 0.719 * | 12.68 * | 0.803 ns | |||
| Cultivars (Cvs) | 1 | 0.261 ns | 84.04 *** | 0.025 ns | |||
| D × Tre | 3 | 0.167 ns | 1.058 ns | 0.069 ns | |||
| D × Cvs | 1 | 0.119 ns | 0.316 ns | 0.075 ns | |||
| Tre × Cvs | 3 | 0.075 ns | 3.232 ns | 0.218 ns | |||
| D × Tre × Cvs | 3 | 0.099 | 0.272 ns | 0.035 ns |
| Source of Variation | df | Shoot Ca2+ | Root Ca2+ | Shoot K+ | Root K+ |
| Drought (D) | 1 | 2495 *** | 244 *** | 3.672 ns | 1.084 ns |
| Trehalose (Tre) | 3 | 4533 * | 2940 ** | 4.198 ns | 4.394 *** |
| Cultivars (Cvs) | 1 | 2245 *** | 1700 *** | 0.173 ns | 7.333 ** |
| D × Tre | 3 | 1117 *** | 526.7 ns | 1.205 ns | 0.770 ns |
| D × Cvs | 1 | 4435 * | 14,241 *** | 0.444 ns | 0.390 ns |
| Tre × Cvs | 3 | 8907 *** | 2876 ** | 0.437 ns | 3.686 ** |
| D × Tre × Cvs | 3 | 2038 ns | 218.1 ns | 1.809 ns | 0.270 ns |
| Shoot P | Root P | Achene Yield/Plant | |||
| Drought (D) | 1 | 0.2674 *** | 0.112 *** | 851.3 *** | |
| Trehalose (Tre) | 3 | 0.004 ns | 0.0007 ns | 24.98 *** | |
| Cultivars (Cvs) | 1 | 0.163 *** | 0.031 *** | 74.54 *** | |
| D × Tre | 3 | 0.002 ns | 0.002 ns | 0.057 ns | |
| D × Cvs | 1 | 0.004 ns | 0.002 ns | 4.574 ns | |
| Tre × Cvs | 3 | 0.001 ns | 0.0002 ns | 1.095 ns | |
| D × Tre × Cvs | 3 | 0.001 ns | 0.0001 ns | 2.256 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kosar, F.; Alshallash, K.S.; Akram, N.A.; Sadiq, M.; Ashraf, M.; Alkhalifah, D.H.M.; Abdel Latef, A.A.H.; Elkelish, A. Trehalose-Induced Regulations in Nutrient Status and Secondary Metabolites of Drought-Stressed Sunflower (Helianthus annuus L.) Plants. Plants 2022, 11, 2780. https://doi.org/10.3390/plants11202780
Kosar F, Alshallash KS, Akram NA, Sadiq M, Ashraf M, Alkhalifah DHM, Abdel Latef AAH, Elkelish A. Trehalose-Induced Regulations in Nutrient Status and Secondary Metabolites of Drought-Stressed Sunflower (Helianthus annuus L.) Plants. Plants. 2022; 11(20):2780. https://doi.org/10.3390/plants11202780
Chicago/Turabian StyleKosar, Firdos, Khalid S. Alshallash, Nudrat Aisha Akram, Muhammad Sadiq, Muhammad Ashraf, Dalal Hussien M. Alkhalifah, Arafat Abdel Hamed Abdel Latef, and Amr Elkelish. 2022. "Trehalose-Induced Regulations in Nutrient Status and Secondary Metabolites of Drought-Stressed Sunflower (Helianthus annuus L.) Plants" Plants 11, no. 20: 2780. https://doi.org/10.3390/plants11202780
APA StyleKosar, F., Alshallash, K. S., Akram, N. A., Sadiq, M., Ashraf, M., Alkhalifah, D. H. M., Abdel Latef, A. A. H., & Elkelish, A. (2022). Trehalose-Induced Regulations in Nutrient Status and Secondary Metabolites of Drought-Stressed Sunflower (Helianthus annuus L.) Plants. Plants, 11(20), 2780. https://doi.org/10.3390/plants11202780