
The Amount of the Rare Sugar Tagatose on Tomato Leaves Decreases after Spray Application under Greenhouse Conditions

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
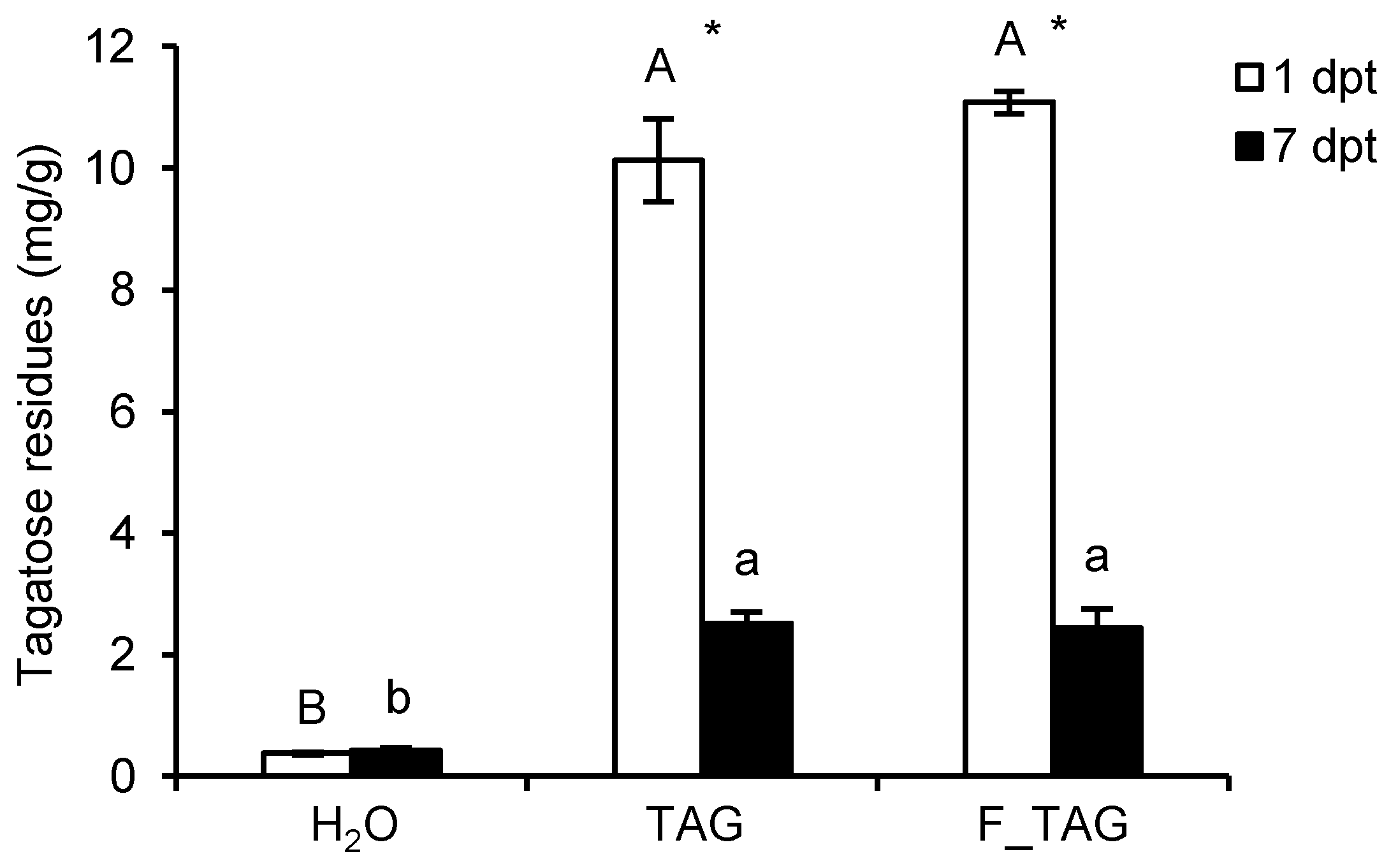
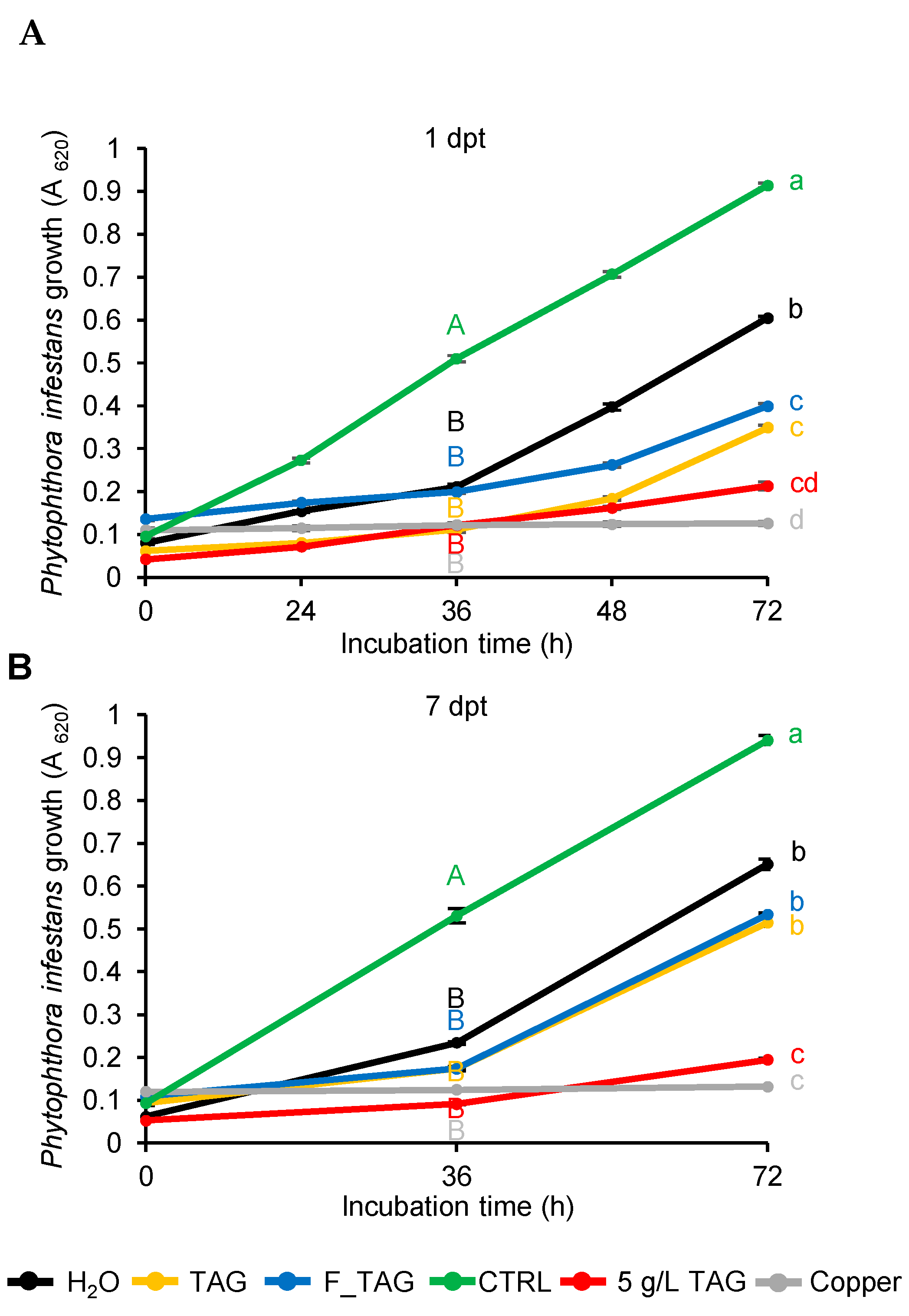
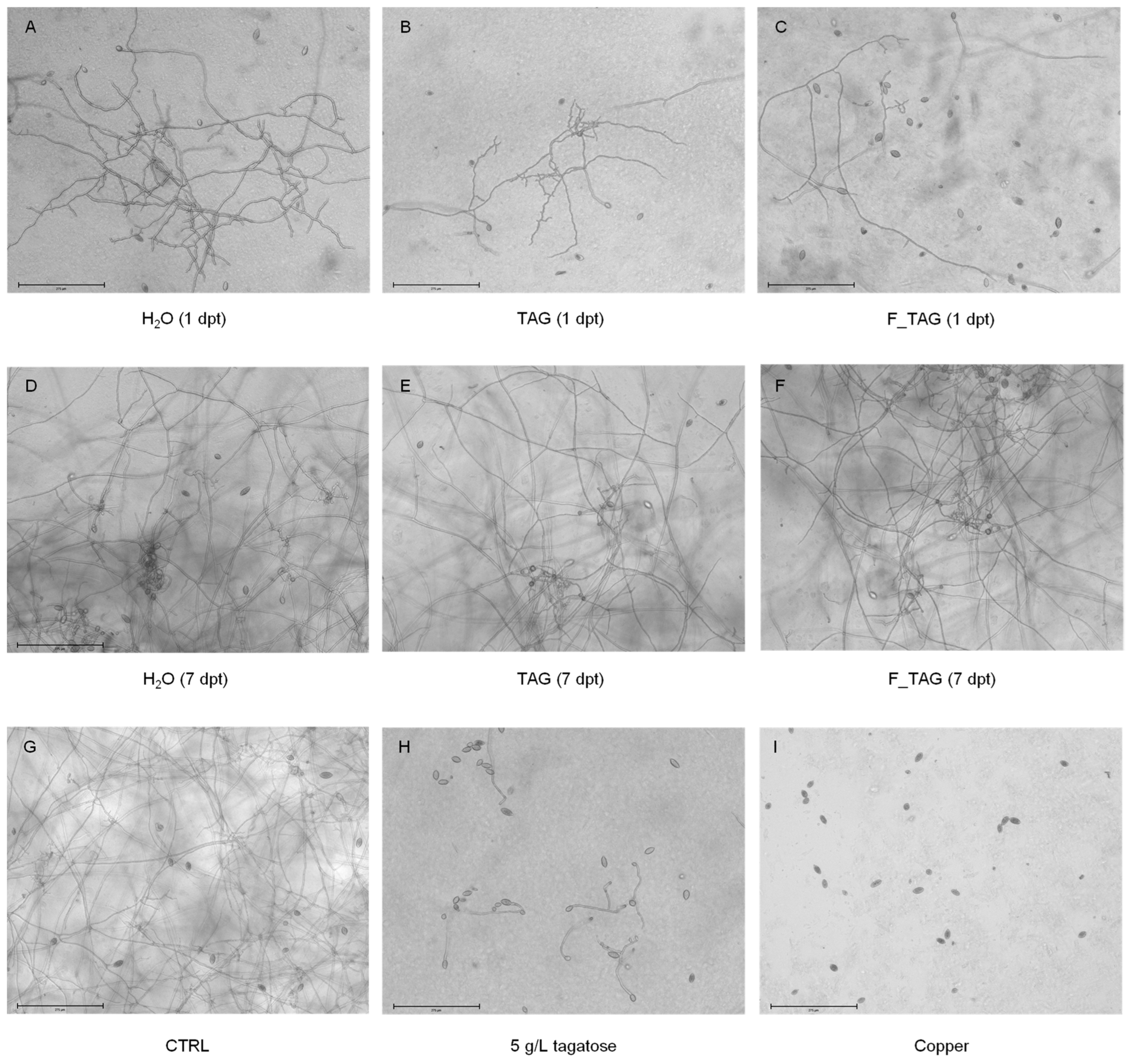
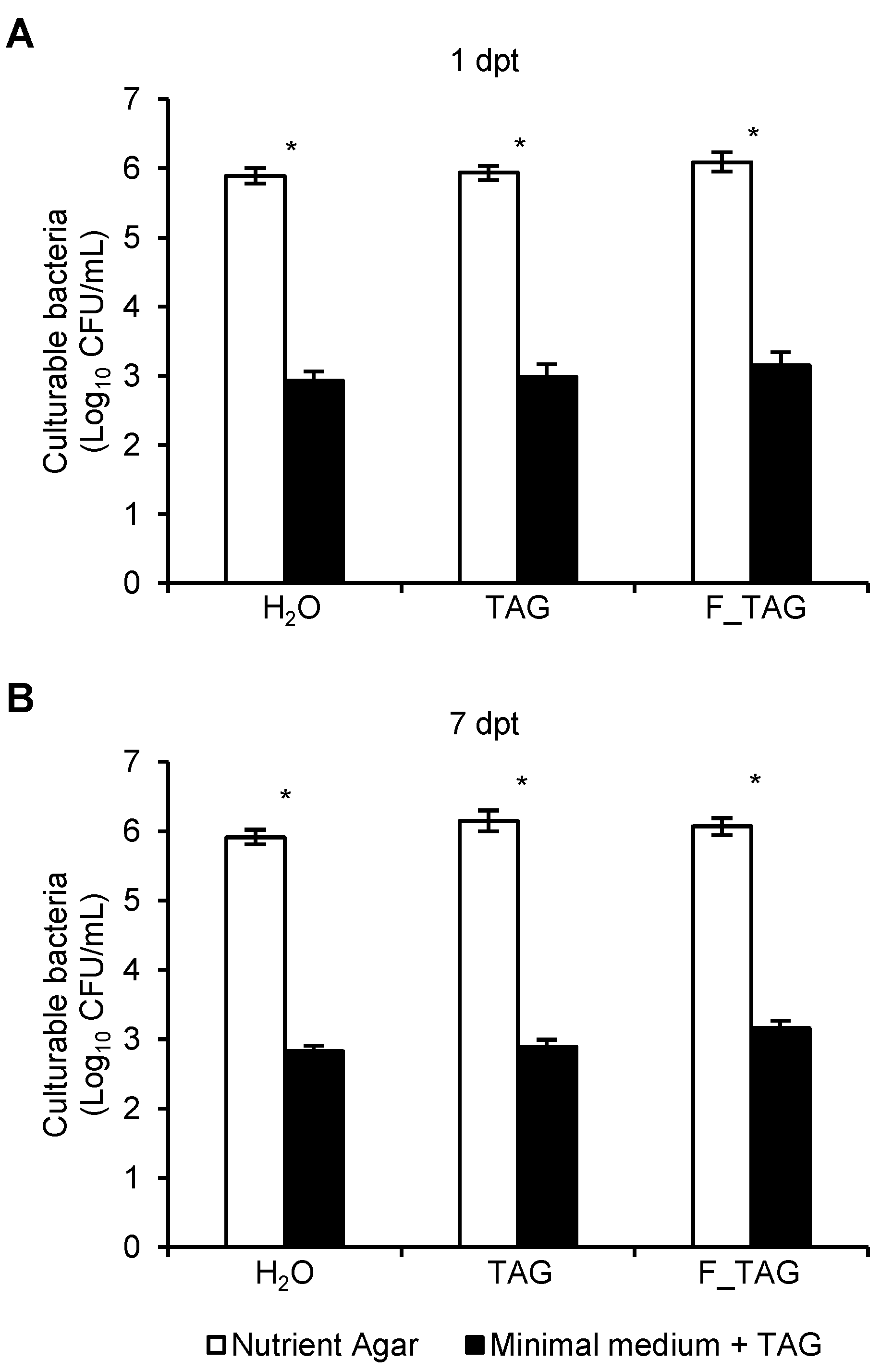
2.1. Tagatose Persistence on Tomato Leaves
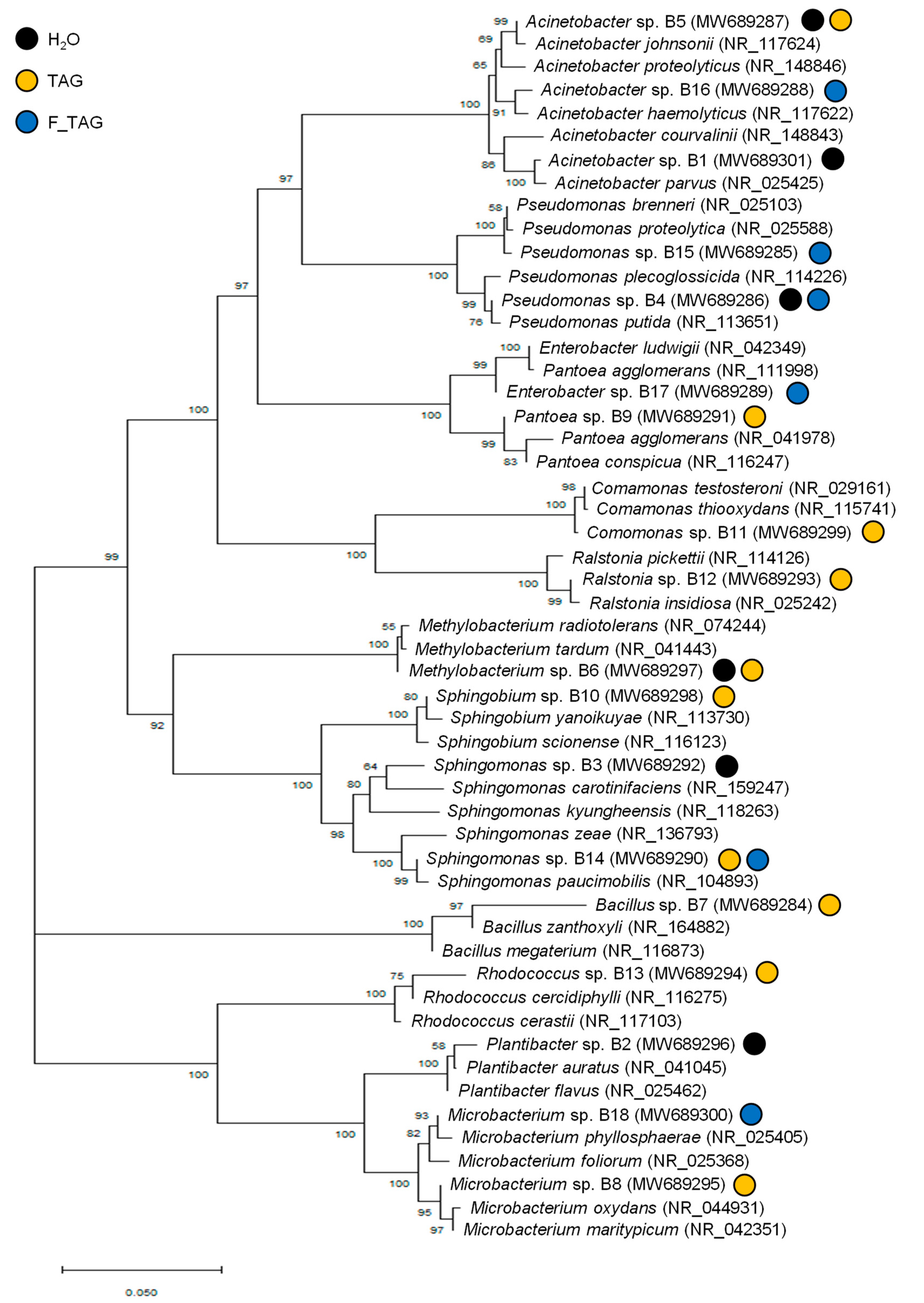
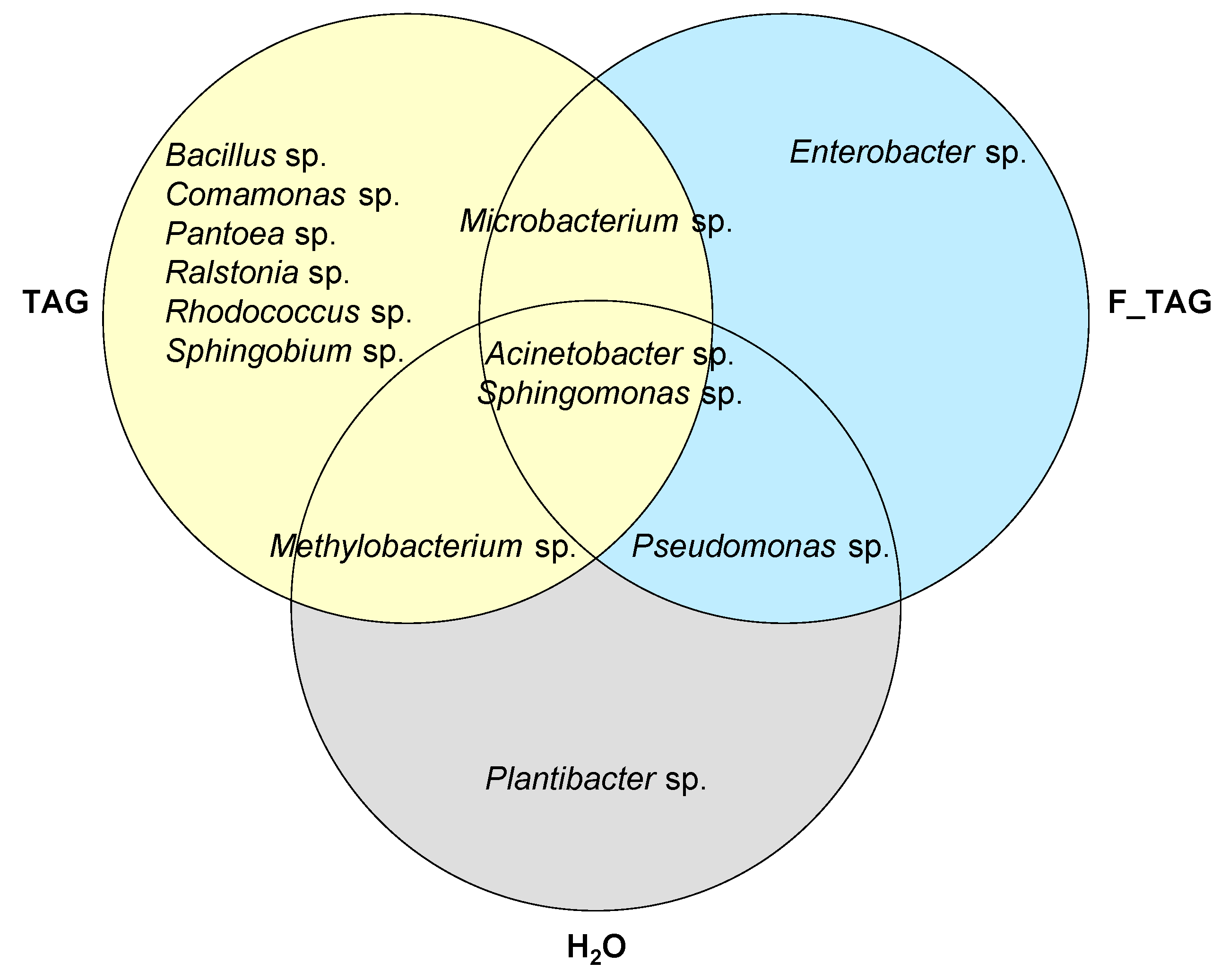
2.2. Potential Tagatose-Degrading Bacteria of Tomato Phyllosphere
3. Discussion
4. Materials and Methods
4.1. Biological Material and Growth Conditions
4.2. Plant Treatments and Collection of Leaf Washing Suspensions
4.3. Tagatose Quantification
4.4. Efficacy Test of Leaf Washing Suspensions against Phytophthora Infestans Growth
4.5. Isolation of Culturable Bacteria
4.6. Identification of Bacterial Isolates
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Granström, T.B.; Takata, G.; Tokuda, M.; Izumori, K. Izumoring: A novel and complete strategy for bioproduction of rare sugars. J. Biosci. Bioeng. 2004, 97, 89–94. [Google Scholar] [CrossRef]
- Li, Z.; Gao, Y.; Nakanishi, H.; Gao, X.; Cai, L. Biosynthesis of rare hexoses using microorganisms and related enzymes. Beilstein J. Org. Chem. 2013, 9, 2434–2445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mijailovic, N.; Nesler, A.; Perazzolli, M.; Aït Barka, E.; Aziz, A. Rare sugars: Recent advances and their potential role in sustainable crop protection. Molecules 2021, 26, 1720. [Google Scholar] [CrossRef]
- Izumori, K. Izumoring: A strategy for bioproduction of all hexoses. J. Biotechnol. 2006, 124, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Vastenavond, C.; Bertelsen, H.; Hansen, S.R.; Laursen, R.; Saunders, J.; Eriknauer, K. Tagatose (D-tagatose). In Alternative Sweeteners, 4th ed.; O’Brien-Nabors, L., Ed.; CRC Press: New York, NY, USA, 2011; Volume 4, pp. 197–222. [Google Scholar]
- Bissett, D.L.; Anderson, R.L. Lactose and D-galactose metabolism in group N streptococci: Presence of enzymes for both the D-galactose 1-phosphate and D-tagatose 6-phosphate pathways. J. Bacteriol. 1974, 117, 318–320. [Google Scholar] [CrossRef] [Green Version]
- Levin, G.V. Tagatose, the new GRAS sweetener and health product. J. Med. Food 2002, 5, 23–36. [Google Scholar] [CrossRef]
- Chahed, A.; Lazazzara, V.; Moretto, M.; Nesler, A.; Corneo, P.E.; Barka, E.A.; Pertot, I.; Puopolo, G.; Perazzolli, M. The differential growth inhibition of Phytophthora spp. caused by the rare sugar tagatose is associated with species-specific metabolic and transcriptional changes. Front. Microbiol. 2021, 12, 711545. [Google Scholar] [CrossRef]
- Chahed, A.; Nesler, A.; Aziz, A.; Barka, E.A.; Pertot, I.; Perazzolli, M. A review of knowledge on the mechanisms of action of the rare sugar d-tagatose against phytopathogenic oomycetes. Plant Pathol. 2021, 70, 1979–1986. [Google Scholar] [CrossRef]
- Chahed, A.; Nesler, A.; Navazio, L.; Baldan, B.; Busato, I.; Ait-Barka, E.; Pertot, I.; Puopolo, G.; Perazzolli, M. The rare sugar tagatose selectively inhibits the growth of Phytophthora infestans and not Phytophthora cinnamomi by interfering with mitochondrial processes. Front. Microbiol. 2020, 11, 128. [Google Scholar] [CrossRef]
- Corneo, P.E.; Jermini, M.; Nadalini, S.; Giovannini, O.; Nesler, A.; Perazzolli, M.; Pertot, I. Foliar and root applications of the rare sugar tagatose control powdery mildew in soilless grown cucumbers. Crop Prot. 2021, 149, 105753. [Google Scholar] [CrossRef]
- Corneo, P.E.; Nesler, A.; Lotti, C.; Chahed, A.; Vrhovsek, U.; Pertot, I.; Perazzolli, M. Interactions of tagatose with the sugar metabolism are responsible for Phytophthora infestans growth inhibition. Microbiol. Res. 2021, 247, 126724. [Google Scholar] [CrossRef] [PubMed]
- Perazzolli, M.; Nesler, A.; Giovannini, O.; Antonielli, L.; Puopolo, G.; Pertot, I. Ecological impact of a rare sugar on grapevine phyllosphere microbial communities. Microbiol. Res. 2020, 232, 126387. [Google Scholar] [CrossRef] [PubMed]
- Ohara, T.; Ishida, Y.; Kudou, R.; Kakibuchi, K.; Akimitsu, K.; Izumori, K.; Tajima, S. Plant Disease Control Agent Comprising D-Tagatose as Active Ingredient, and Plant Disease Control Method. US Patent EP 2 329 713 A1, 18 August 2008. [Google Scholar]
- Mochizuki, S.; Fukumoto, T.; Ohara, T.; Ohtani, K.; Yoshihara, A.; Shigematsu, Y.; Tanaka, K.; Ebihara, K.; Tajima, S.; Gomi, K.; et al. The rare sugar D-tagatose protects plants from downy mildews and is a safe fungicidal agrochemical. Commun. Biol. 2020, 3, 423. [Google Scholar] [CrossRef] [PubMed]
- Mijailovic, N.; Nesler, A.; Perazzolli, M.; Aziz, A.; Essaïd, A.B. Foliar application of a tagatose-based product reduces downy mildew symptoms through induction of grapevine resistance and anti-oomycete action. Agronomy 2022, 12, 498. [Google Scholar] [CrossRef]
- Mijailovic, N.; Richet, N.; Villaume, S.; Nesler, A.; Perazzolli, M.; Aït Barka, E.; Aziz, A. D-tagatose-based product triggers sweet immunity and resistance of grapevine to downy mildew, but not to gray mold disease. Plants 2022, 11, 296. [Google Scholar] [CrossRef]
- Fry, J.C. Natural low-calorie sweeteners. In Natural Food Additives, Ingredients and Flavourings; Baines, D., Seal, R., Eds.; Woodhead Publishing: Sawston, UK, 2012; pp. 41–75. [Google Scholar]
- Giovannini, O.; Roman, T.; Nesler, A.; Pertot, I.; Perazzolli, M. Tagatose suppresses grapevine powdery mildew and downy mildew under field conditions with no severe impacts on grape must fermentation. Aust. J. Grape Wine Res. 2022; accepted. [Google Scholar]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef] [Green Version]
- Muller, T.; Ruppel, S. Progress in cultivation-independent phyllosphere microbiology. FEMS Microbiol. Ecol. 2014, 87, 2–17. [Google Scholar] [CrossRef]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef]
- Martinussen, J.; Solem, C.; Holm, A.K.; Jensen, P.R. Engineering strategies aimed at control of acidification rate of lactic acid bacteria. Curr. Opin. Biotechnol. 2013, 24, 124–129. [Google Scholar] [CrossRef]
- Raichand, R.; Pareek, S.; Singh, N.K.; Mayilraj, S. Exiguobacterium aquaticum sp. nov., a member of the genus Exiguobacterium. Int. J. Syst. Evol. Microbiol. 2012, 62, 2150–2155. [Google Scholar] [CrossRef]
- Van der Heiden, E.; Delmarcelle, M.; Lebrun, S.; Freichels, R.; Brans, A.; Vastenavond, C.M.; Galleni, M.; Joris, B. A pathway closely related to the D-tagatose pathway of Gram-negative enterobacteria identified in the Gram-positive bacterium Bacillus licheniformis. Appl. Environ. Microbiol. 2013, 79, 3511–3515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Shah, N.P. The potential of species-specific tagatose-6-phosphate (T6P) pathway in Lactobacillus casei group for galactose reduction in fermented dairy foods. Food Microbiol. 2017, 62, 178–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, M.V.; Brenner, D.J.; Steigerwalt, A.G.; Kosako, Y.; Komagata, K. Erwinia persicinus, a new species isolated from plants. Int. J. Syst. Bacteriol. 1990, 40, 379–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koskella, J.; Stotzky, G. Microbial utilization of free and clay-bound insecticidal toxins from Bacillus thuringiensis and their retention of insecticidal activity after incubation with microbes. Appl. Environ. Microbiol. 1997, 63, 3561–3568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, C.J.; Wang, L.L.; Li, Q.; Shang, Q.M. Bacterial communities in the rhizosphere, phyllosphere and endosphere of tomato plants. PLoS ONE 2019, 14, e0223847. [Google Scholar] [CrossRef]
- Toju, H.; Okayasu, K.; Notaguchi, M. Leaf-associated microbiomes of grafted tomato plants. Sci. Rep. 2019, 9, 1787. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xia, K.; Yang, X.; Tang, C. Growth strategy of microbes on mixed carbon sources. Nat. Commun. 2019, 10, 1279. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Ding, Y.; Liu, M.; Xian, M.; Zhao, G. Comparison of glucose, acetate and ethanol as carbon resource for oroduction of ooly(3-hydroxybutyrate) and other acetyl-CoA derivatives. Front. Bioeng. Biotechnol. 2020, 8, 833. [Google Scholar] [CrossRef]
- Araújo, W.J.; Oliveira, J.S.; Araújo, S.C.S.; Minnicelli, C.F.; Silva-Portela, R.C.B.; da Fonseca, M.M.B.; Freitas, J.F.; Silva-Barbalho, K.K.; Napp, A.P.; Pereira, J.E.S.; et al. Microbial culture in minimal medium with oil favors enrichment of biosurfactant producing genes. Front. Bioeng. Biotechnol. 2020, 8, 962. [Google Scholar] [CrossRef]
- Izumori, K.; Tsusaki, K. L-Ribose Isomerase, Its Preparation and Uses. European Patent EP 0 807 682 A2, 14 May 1997. [Google Scholar]
- Itoh, H.; Okaya, H.; Khan, A.R.; Tajima, S.; Hayakawa, S.; Izumori, K. Purification and characterization of D-tagatose 3-epimerase from Pseudomonas sp. ST-24. Biosci. Biotechnol. Biochem. 1994, 58, 2168–2171. [Google Scholar] [CrossRef]
- Cappelletti, M.; Perazzolli, M.; Antonielli, L.; Nesler, A.; Torboli, E.; Bianchedi, P.L.; Pindo, M.; Puopolo, G.; Pertot, I. Leaf treatments with a protein-based resistance inducer partially modify phyllosphere microbial communities of grapevine. Front. Plant. Sci. 2016, 7, 1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, I.S.; Santos Eda, C.; Menezes, C.R.; Faria, A.F.; Franciscon, E.; Grossman, M.; Durrant, L.R. Bioremediation of a polyaromatic hydrocarbon contaminated soil by native soil microbiota and bioaugmentation with isolated microbial consortia. Bioresour. Technol. 2009, 100, 4669–4675. [Google Scholar] [CrossRef] [PubMed]
- Indraningrat, A.A.G.; Micheller, S.; Runderkamp, M.; Sauerland, I.; Becking, L.E.; Smidt, H.; Sipkema, D. Cultivation of sponge-associated bacteria from Agelas sventres and Xestospongia muta collected from different depths. Mar. Drugs 2019, 17, 578. [Google Scholar] [CrossRef] [Green Version]
- Esmaeel, Q.; Jacquard, C.; Clement, C.; Sanchez, L.; Ait Barka, E. Genome sequencing and traits analysis of Burkholderia strains reveal a promising biocontrol effect against grey mould disease in grapevine (Vitis vinifera L.). World J. Microbiol. Biotechnol. 2019, 35, 40. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number 1 | Isolate Code 2 | Plant Treatment 3 | Taxonomic Identification 4 | BLAST Best Hit 5 | Sequence Identity (%) 6 |
| MW689301 | B1 | H2O | Acinetobacter sp. | Acinetobacter parvus | 99.25 |
| MW689296 | B2 | H2O | Plantibacter sp. | Plantibacter flavus | 99.58 |
| MW689292 | B3 | H2O | Sphingomonas sp. | Sphingomonas kyungheensis | 96.75 |
| MW689286 | B4 | H2O and F_TAG | Pseudomonas sp. | Pseudomonas putida | 99.90 |
| MW689287 | B5 | H2O and TAG | Acinetobacter sp. | Acinetobacter johnsonii | 99.92 |
| MW689297 | B6 | H2O and TAG | Methylobacterium sp. | Methylobacterium radiotolerans | 99.92 |
| MW689284 | B7 | TAG | Bacillus sp. | Bacillus megaterium | 99.88 |
| MW689295 | B8 | TAG | Microbacterium sp. | Microbacterium oxydans | 99.46 |
| MW689291 | B9 | TAG | Pantoea sp. | Pantoea agglomerans | 98.88 |
| MW689298 | B10 | TAG | Sphingobium sp. | Sphingobium yanoikuyae | 99.69 |
| MW689299 | B11 | TAG | Comamonas sp. | Comamonas testosteroni | 99.39 |
| MW689293 | B12 | TAG | Ralstonia sp. | Ralstonia insidiosa | 99.63 |
| MW689294 | B13 | TAG | Rhodococcus sp. | Rhodococcus cercidiphylli | 99.59 |
| MW689290 | B14 | TAG and F_TAG | Sphingomonas sp. | Sphingomonas paucimobilis | 99.71 |
| MW689285 | B15 | F_TAG | Pseudomonas sp. | Pseudomonas brenneri | 99.71 |
| MW689288 | B16 | F_TAG | Acinetobacter sp. | Acinetobacter haemolyticus | 98.94 |
| MW689289 | B17 | F_TAG | Enterobacter sp. | Enterobacter ludwigii | 99.54 |
| MW689300 | B18 | F_TAG | Microbacterium sp. | Microbacterium phyllosphaerae | 99.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chahed, A.; Nesler, A.; Esmaeel, Q.; Barka, E.A.; Perazzolli, M. The Amount of the Rare Sugar Tagatose on Tomato Leaves Decreases after Spray Application under Greenhouse Conditions. Plants 2022, 11, 2781. https://doi.org/10.3390/plants11202781
Chahed A, Nesler A, Esmaeel Q, Barka EA, Perazzolli M. The Amount of the Rare Sugar Tagatose on Tomato Leaves Decreases after Spray Application under Greenhouse Conditions. Plants. 2022; 11(20):2781. https://doi.org/10.3390/plants11202781
Chicago/Turabian StyleChahed, Abdessalem, Andrea Nesler, Qassim Esmaeel, Essaid Ait Barka, and Michele Perazzolli. 2022. "The Amount of the Rare Sugar Tagatose on Tomato Leaves Decreases after Spray Application under Greenhouse Conditions" Plants 11, no. 20: 2781. https://doi.org/10.3390/plants11202781
APA StyleChahed, A., Nesler, A., Esmaeel, Q., Barka, E. A., & Perazzolli, M. (2022). The Amount of the Rare Sugar Tagatose on Tomato Leaves Decreases after Spray Application under Greenhouse Conditions. Plants, 11(20), 2781. https://doi.org/10.3390/plants11202781