
More than Moths: Flower Visitors of a Night-Blooming Plant in South Florida Pine Rocklands, USA

Abstract
:1. Introduction
2. Results
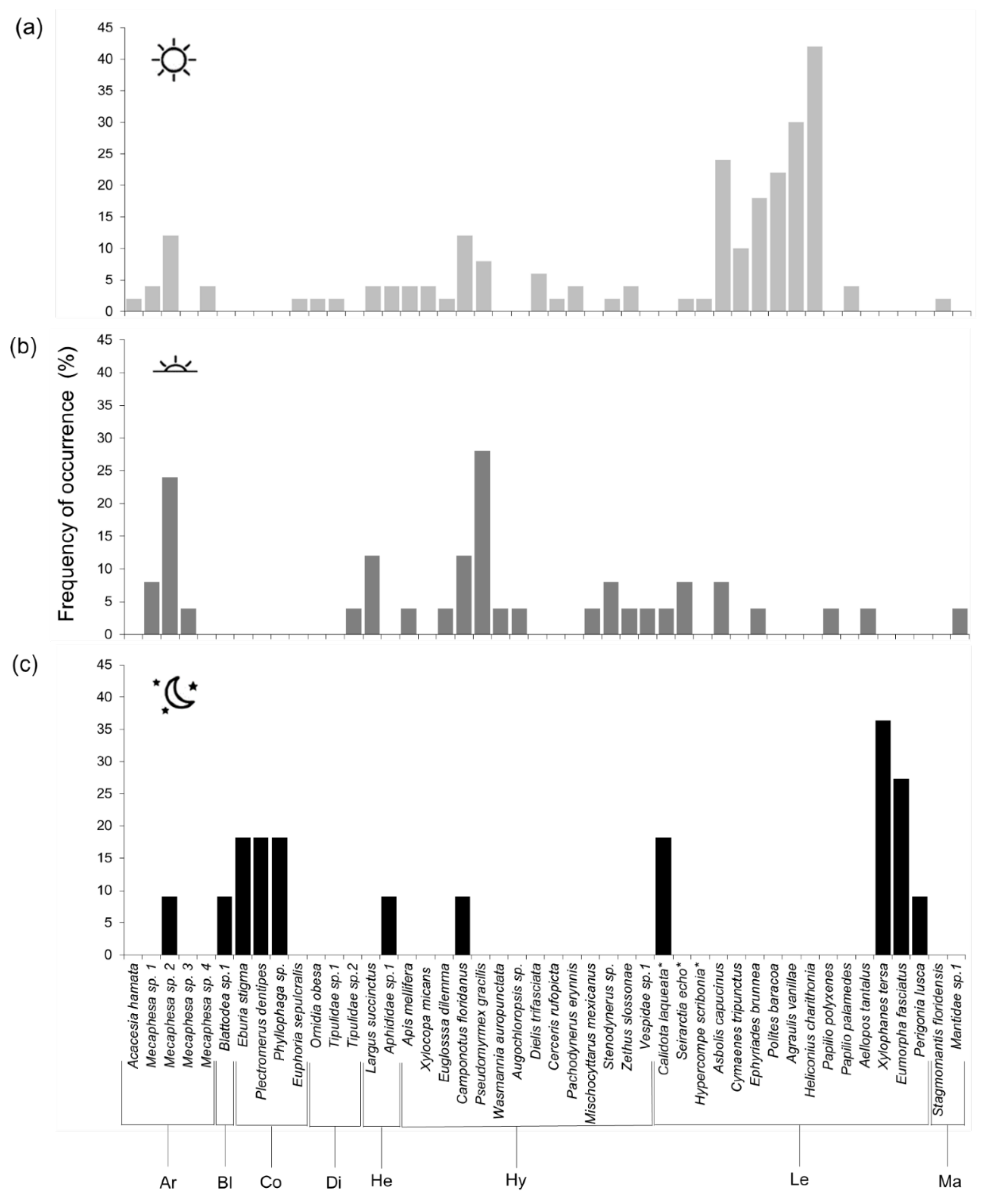
2.1. Occurrence
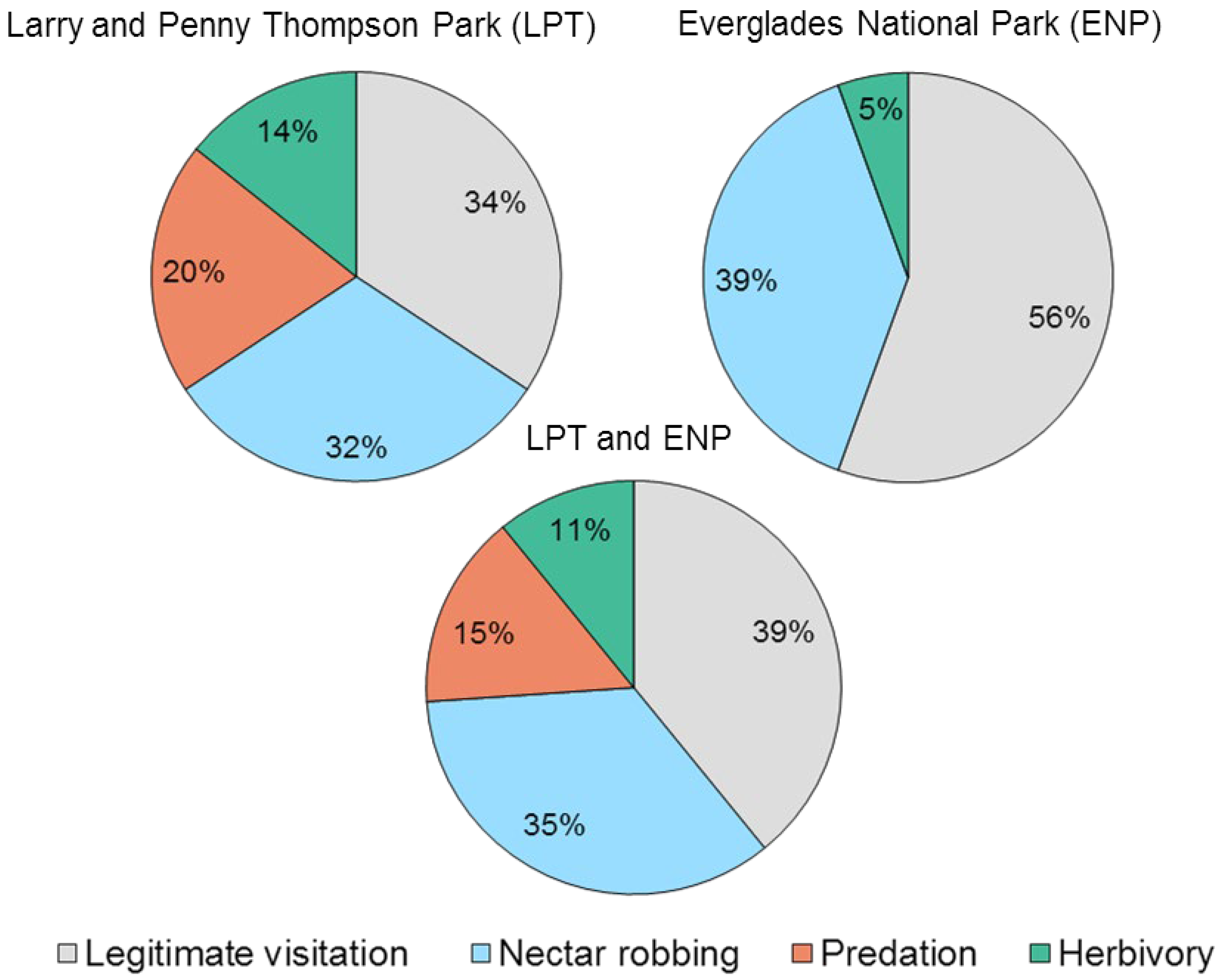
2.2. Visitor Behavior
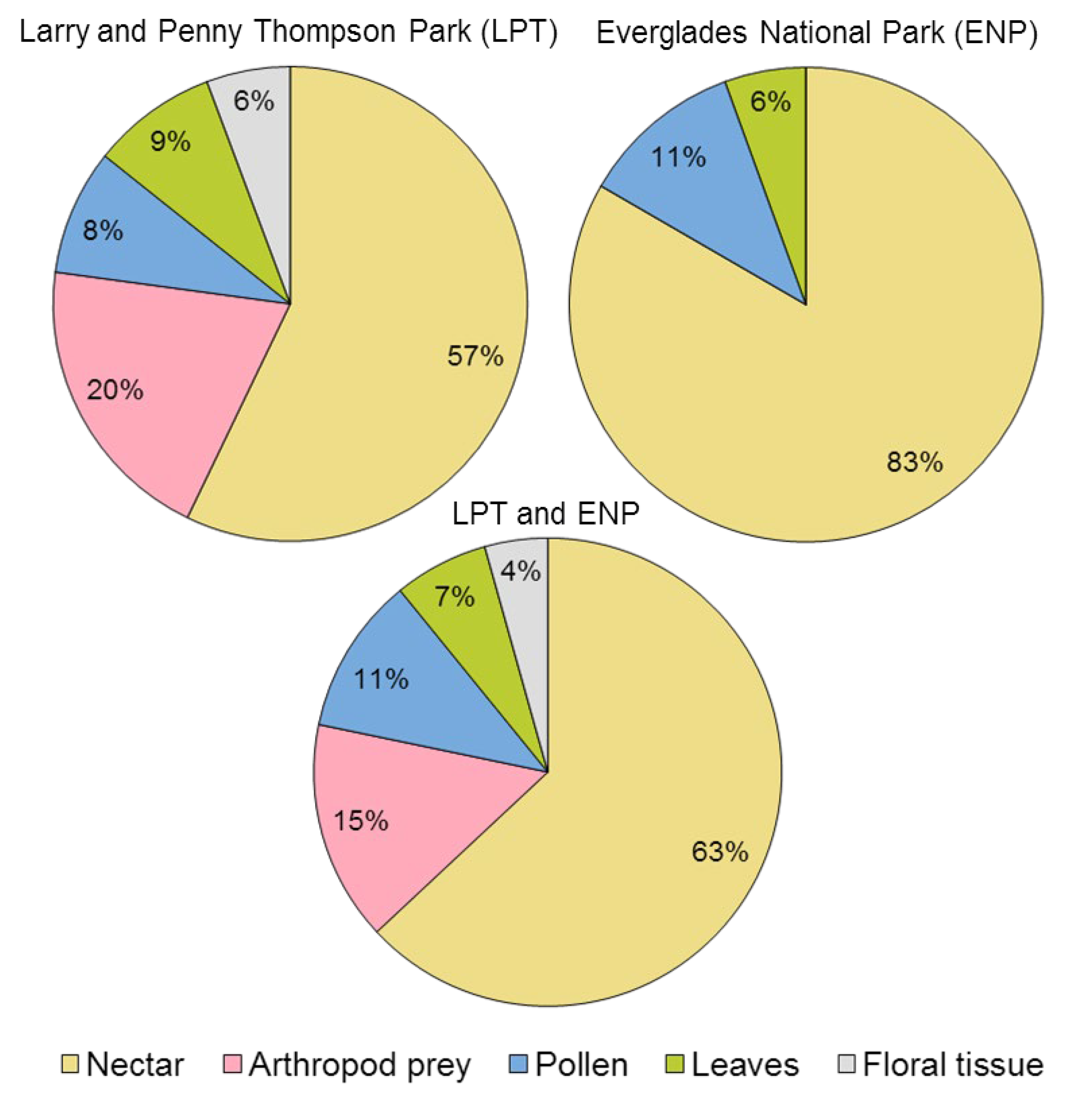
2.3. Resources Consumed by Visitors
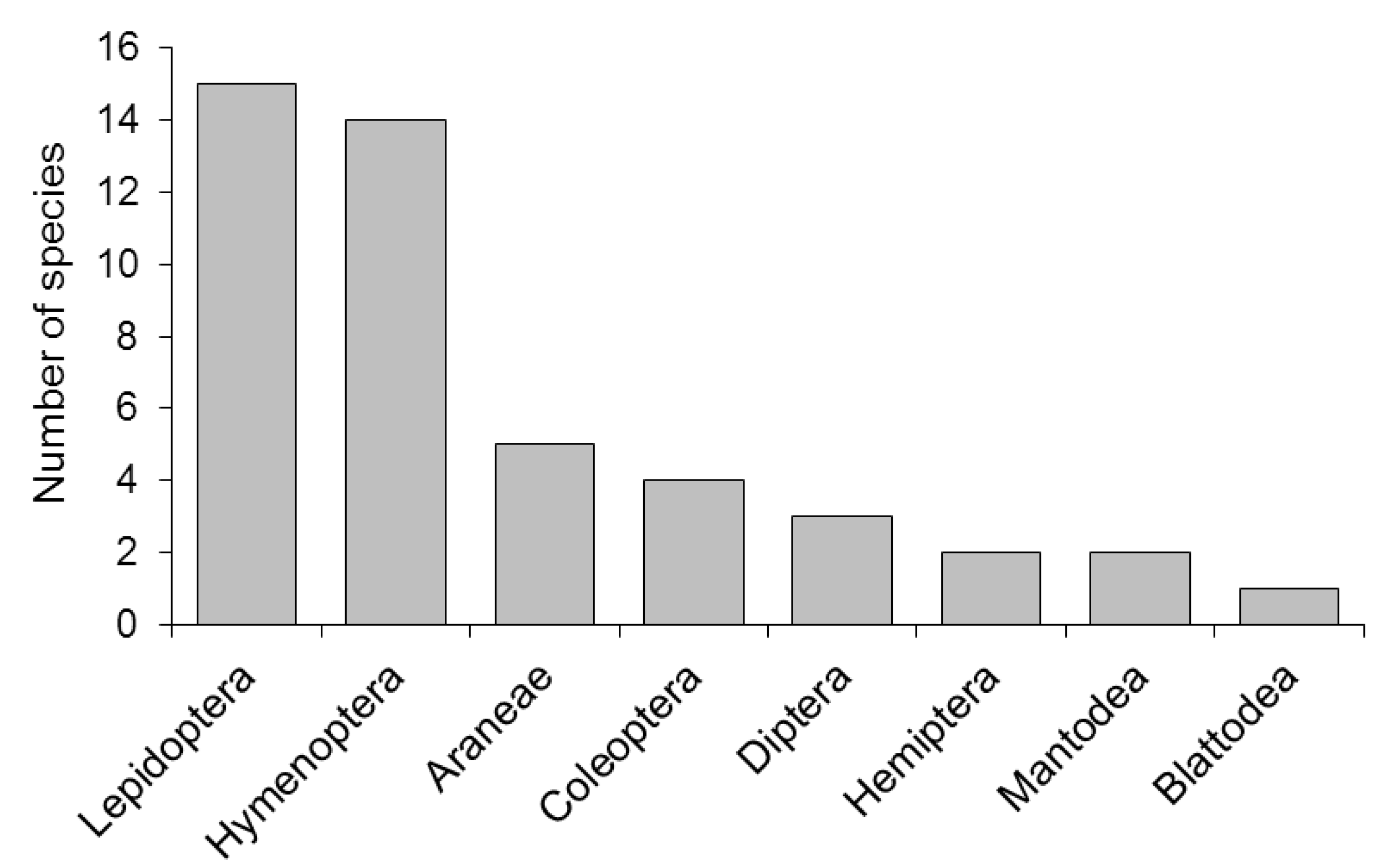
2.4. Taxonomic Diversity and Behaviors of Visitors
2.4.1. Lepidoptera
2.4.2. Hymenoptera
2.4.3. Coleoptera
2.4.4. Other Insect Orders
2.4.5. Aranae
3. Discussion
3.1. Occurrence
3.2. Potential Pollinators
3.3. Nectar Robbing: A Common Behavior
3.4. Guettarda scabra as Food Source for Local Arthropod Fauna
4. Materials and Methods
4.1. Plant Species
4.2. Study Sites
4.3. Flower Visitor Observations
4.4. Statistical Comparisons
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waser, N.M.; Chittka, L.; Price, M.V.; Williams, N.M.; Ollerton, J. Generalization in pollination systems, and why it matters. Ecology 1996, 77, 1043–1060. [Google Scholar] [CrossRef] [Green Version]
- Fleming, T.H.; Sahley, C.T.; Holland, J.N.; Nason, J.D.; Hamrick, J.L. Sonoran Desert columnar cacti and the evolution of generalized pollination systems. Ecol. Monogr. 2001, 71, 511–530. [Google Scholar] [CrossRef]
- De Merxem, D.G.; Borremans, B.; De Jäger, M.L.; Johnson, T.; Jooste, M.; Ros, P.; Zenni, R.D.; Ellis, A.G.; Anderson, B. The importance of flower visitors not predicted by floral syndromes. S. Afr. J. Bot. 2009, 75, 660–667. [Google Scholar] [CrossRef]
- Miyake, T.; Yahara, T. Theoretical evaluation of pollen transfer by nocturnal and diurnal pollinators: When should a flower open? Oikos 1999, 86, 233–240. [Google Scholar] [CrossRef]
- Acevedo-Rodríguez, P.; Strong, M.T. Catalogue of Seed Plants of the West Indies; Smithsonian Institution: Washington, DC, USA, 2012. [Google Scholar] [CrossRef]
- Koptur, S.; Garcia, D. Habitat differences in morphology and reproductive allocation in Guettarda scabra (Rubiaceae). Castanea 2017, 82, 51–57. [Google Scholar] [CrossRef]
- Roberts, A. Guettarda scabra. The IUCN Red List of Threatened Species. 2014. e.T56503696A56503850. Available online: https://www.iucnredlist.org/species/56503696/56503850 (accessed on 19 October 2022).
- WCSP: World Checklist of Selected Plant Families. Facilitated by the Royal Botanic Gardens, Kew. 2022. Available online: https://wcsp.science.kew.org (accessed on 19 October 2022).
- Davis, A.P.; Govaerts, R.; Bridson, D.M.; Ruhsam, M.; Moat, J.; Brummitt, N.A. A global assessment of distribution, diversity, endemism, and taxonomic effort in the Rubiaceae 1. Ann. Mo. Bot. Gard. 2009, 96, 68–78. [Google Scholar] [CrossRef]
- Richards, J.H.; Koptur, S. Floral variation and distyly in Guettarda scabra (Rubiaceae). Am. J. Bot. 1993, 80, 31–40. [Google Scholar] [CrossRef]
- Faegri, K.; van der Pijl, L. The Principles of Pollination Ecology; Pergamon Press: Oxford, UK, 1979. [Google Scholar]
- Tomlinson, P.B. The Biology of Trees Native to Tropical Florida; Harvard University Printing Office: Allston, MA, USA, 1980. [Google Scholar]
- Austin, D.F. Florida Ethnobotany; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Koptur, S. Scientific Note: Insects associated with Guettarda scabra in Everglades National Park, Florida. Castanea 2020, 85, 155–158. [Google Scholar] [CrossRef]
- Wagner, D.L.; Grames, E.M.; Forister, M.L.; Berenbaum, M.R.; Stopak, D. Insect decline in the Anthropocene: Death by a thousand cuts. Proc. Natl. Acad. Sci. USA 2021, 118, e2023989118. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, C.; Pimienta, M.C. Behavior of adult Leucopis sp. (Chamaemyiidae) associated with aphids feeding on flowers of the rough-leaf velvetseed (Guettarda scabra: Rubiaceae) in south Florida. Fly Times 2019, 62, 8–10. Available online: https://www.nadsdiptera.org/News/FlyTimes/issue62.pdf (accessed on 19 October 2022).
- Wardhaugh, C.W. How many species of arthropods visit flowers? Arthropod-Plant Interact. 2015, 9, 547–565. [Google Scholar] [CrossRef]
- Maruyama, P.K.; Amorim, F.W.; Oliveira, P.E. Night and day service: Distyly and mixed pollination system in Faramea cyanea (Rubiaceae). Flora Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 818–824. [Google Scholar] [CrossRef]
- Locatelli, E.; Machado, I.C.S. Floral biology of Cereus fernambucensis: A sphingophilous cactus of restinga. Bradleya 1999, 17, 86–94. [Google Scholar] [CrossRef]
- Walter, H.E. Floral biology of Echinopsis chiloensis ssp. chiloensis (Cactaceae): Evidence for a mixed pollination syndrome. Flora Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 757–763. [Google Scholar] [CrossRef]
- Young, H.J. Diurnal and nocturnal pollination of Silene alba (Caryophyllaceae). Am. J. Bot. 2002, 89, 433–440. [Google Scholar] [CrossRef]
- Haber, W.A.; Frankie, G.W. A tropical hawkmoth community: Costa Rican dry forest Sphingidae. Biotropica 1989, 21, 155–172. [Google Scholar] [CrossRef]
- Miyake, T.; Yahara, T. Why does the flower of Lonicera japonica open at dusk? Can. J. Bot. 1998, 76, 1806–1811. [Google Scholar] [CrossRef]
- Darrault, R.O.; Schlindwein, C. Esfingídeos (Lepidoptera, Sphingidae) no Tabuleiro Paraibano, nordeste do Brasil: Abundância, riqueza e relação com plantas esfingófilas. Rev. Bras. Zool. 2002, 19, 429–443. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Rodríguez, P.A.; Krömer, T.; García-Franco, J.G.; MacSwiney, G.M.C. From dusk till dawn: Nocturnal and diurnal pollination in the epiphyte Tillandsia heterophylla (Bromeliaceae). Plant Biol. 2016, 18, 37–45. [Google Scholar] [CrossRef]
- Wolff, D.; Braun, M.; Liede, S. Nocturnal versus diurnal pollination success in Isertia laevis (Rubiaceae): A sphingophilous plant visited by hummingbirds. Plant Biol. 2003, 5, 71–78. [Google Scholar] [CrossRef]
- Xu, Y.; Luo, Z.; Gao, S.; Zhang, D. Pollination niche availability facilitates colonization of Guettarda speciosa with heteromorphic self-incompatibility on oceanic islands. Sci. Rep. 2018, 8, 13765. [Google Scholar] [CrossRef] [Green Version]
- Martínez, L. Fenología Reproductiva y Efecto del Robo de Néctar en el Éxito Reproductivo de Guettarda clarensis, en. Bachelor’s Thesis, Universidad Central “Marta Abreu” de Las Villas, Santa Clara, Villa Clara, Cuba, 2013. Available online: https://dspace.uclv.edu.cu/handle/123456789/1723 (accessed on 19 October 2022).
- Novo, R.R.; Consolaro, H.; Almeida, N.M.; Castro, C.C. Floral biology of the velvetseed Guettarda platypoda DC. (Rubiaceae): Atypical distyly or style dimorphism? Flora Morphol. Distrib. Funct. Ecol. Plants 2018, 239, 62–70. [Google Scholar] [CrossRef]
- Warriner, M.D. A range extension for the large carpenter bee Xylocopa micans (Hymenoptera: Apidae) with notes on floral and habitat associations. J. Kans. Entomol. Soc. 2010, 83, 267–269. [Google Scholar] [CrossRef]
- Lenczewski, B. Butterflies of Everglades National Park; Report T-588; South Florida Research Center: Homestead, FL, USA, 1980; 110p. [Google Scholar]
- Fishbein, M.; Venable, D.L. Diversity and temporal change in the effective pollinators of Asclepias tuberosa. Ecology 1996, 77, 1061–1073. [Google Scholar] [CrossRef]
- Watanabe, K.; Kato, H.; Kuraya, E.; Sugawara, T. Pollination and reproduction of Psychotria homalosperma, an endangered distylous tree endemic to the oceanic Bonin (Ogasawara) Islands, Japan. Plant Species Biol. 2018, 33, 16–27. [Google Scholar] [CrossRef]
- Pemberton, R.W.; Wheeler, G.S. Orchid bees don’t need orchids: Evidence from the naturalization of an orchid bee in Florida. Ecology 2006, 87, 1995–2001. [Google Scholar] [CrossRef]
- Koptur, S. Flowering phenology and floral biology of Inga (Fabaceae: Mimosoideae). Syst. Bot. 1983, 8, 354–368. [Google Scholar] [CrossRef]
- Koptur, S.; Peña, S.; Barrios, R.B. Do morning butterfly visitors benefit a night-flowering hawkmoth pollinated plant? Castanea 2021, 86, 100–111. [Google Scholar] [CrossRef]
- Genini, J.; Morellato, L.P.C.; Guimarães, P.R., Jr.; Olesen, J.M. Cheaters in mutualism networks. Biol. Lett. 2010, 6, 494–497. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Pérez, L.; Faife-Cabrera, M. Nectar Robbing in Guettarda clarensis (Rubiaceae): Does Floral Neighborhood Matter? Rev. Del. Jardín Botánico Nac. 2019, 40, 47–57. Available online: https://www.rjbn.uh.cu/index.php/RJBN/article/view/443/465 (accessed on 19 October 2022).
- Martínez, L. Relación del Vecindario Floral Con la Frecuencia de Robo de Néctar en Guettarda clarensis (Rubiaceae). Master’s Thesis, Universidad Central “Marta Abreu” de Las Villas, Santa Clara, Villa Clara, Cuba, 2017. Available online: https://dspace.uclv.edu.cu/handle/123456789/10813 (accessed on 19 October 2022).
- Schlindwein, C.; Westerkamp, C.; Carvalho, A.T.; Milet-Pinheiro, P. Visual signalling of nectar-offering flowers and specific morphological traits favour robust bee pollinators in the mass-flowering tree Handro-anthus impetiginosus (Bignoniaceae). Bot. J. Linn. Soc. 2014, 176, 396–407. [Google Scholar] [CrossRef]
- Irwin, R.E.; Bronstein, J.L.; Manson, J.S.; Richardson, L. Nectar robbing: Ecological and evolutionary perspectives. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 271–292. [Google Scholar] [CrossRef]
- Barrows, E.M. Robbing of exotic plants by introduced carpenters and honeybees in Hawaii, with comparative notes. Biotropica 1980, 12, 23–29. [Google Scholar] [CrossRef]
- Dedej, S.; Delaplane, K.S. Nectar-robbing carpenter bees reduce seed-setting capability of honey bees (Hymenoptera: Apidae) in rabbiteye blueberry, Vaccinium ashei, ‘Climax’. Environ. Entomol. 2004, 33, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Guitián, J.; Sánchez, J.M.; Guitián, P. Pollination ecology of Petrocoptis grandiflora Rothm. (Caryophyllaceae); a species endemic to the north-west part of the Iberian Peninsula. Bot. J. Linn. Soc. 1994, 115, 19–27. [Google Scholar] [CrossRef]
- Schmid, S.; Schmid, V.S.; Zillikens, A.; Steiner, J. Diversity of flower visitors and their role for pollination in the ornithophilous bromeliad Vriesea friburgensis in two different habitats in southern Brazil. Ecotropica 2011, 17, 91–102. [Google Scholar]
- Zhang, Y.W.; Robert, G.W.; Wang, Y.; Guo, Y.H. Nectar robbing of a carpenter bee and its effects on the reproductive fitness of Glechoma longituba (Lamiaceae). Plant Ecol. 2007, 193, 1–13. [Google Scholar] [CrossRef]
- Scott, P.E.; Buchmann, S.L.; O’Rourke, M.K. Evidence for mutualism between a flower-piercing carpenter bee and ocotillo: Use of pollen and nectar by nesting bees. Ecol. Entomol. 1993, 18, 234–240. [Google Scholar] [CrossRef]
- Gottsberger, G. Some pollination strategies in neotropical savannas and forests. Plant Syst. Evol. 2004, 152, 29–45. [Google Scholar] [CrossRef]
- Singh, V.K.; Barman, C.; Tandon, R. Nectar Robbing Positively Influences the Reproductive Success of Tecomella undulata (Bignoniaceae). PLoS ONE 2014, 9, e102607. [Google Scholar] [CrossRef]
- Maloof, J. The effects of a bumble bee nectar robber on plant reproductive success and pollinator behavior. Am. J. Bot. 2001, 88, 1960–1965. [Google Scholar] [CrossRef] [Green Version]
- Loope, L.L. Phenology of Flowering and Fruiting in Plant Communities of Everglades National Park and Biscayne National Monument, Florida; Report T-593; South Florida Research Center: Homestead, FL, USA, 1980; 50p. [Google Scholar]
- Gunderson, L.; Taylor, D.; Craig, J. Fire Effects on Flowering and Fruiting Patterns of Understory Plants in Pinelands of Everglades National Park; Report SFRC-83/04; South Florida Research Center: Homestead, FL, USA, 1983; 36p. [Google Scholar]
- Grissell, E.E. Mason Wasps of Florida, Zethus spp. (Insecta: Hymenoptera: Vespidae: Eumeninae); Entomology Circular 153; Florida Department of Agriculture and Consumer Services, Division of Plant Industry, UF/IFAS Extension: Gainesville, FL, USA, 2021; pp. 1–3. [Google Scholar]
- Grissell, E.E. Scoliid Wasps of Florida, Campsomeris, Scolia and Trieliss pp. (Insecta: Hymenoptera: Scoliidae); Entomology Circular 179 and 185; Florida Department of Agriculture and Consumer Services, Division of Plant Industry, UF/IFAS Extension: Gainesville, FL, USA, 2017; pp. 1–14. [Google Scholar]
- Tuttle, J.P. The Hawk Moths of North America: A Natural History Study of the Sphingidae of the United States and Canada; Wedge Entomological Research Foundation: Washington, DC, USA, 2007. [Google Scholar]
- Warren, A.D.; Davis, K.J.; Grishin, N.V.; Pelham, J.P.; Stangeland, E.M. Interactive Listing of American Butterflies. 2012. Available online: https://www.butterfliesofamerica.com/ (accessed on 19 October 2022).
- Daniels, J. Florida butterflies. Wings: Essays Invertebr. Conserv. 2010, 33, 18–21. Available online: https://xerces.org/wings (accessed on 19 October 2022).
- Boggs, C.L. Nutritional and life-history determinants of resource allocation in holometabolous insects. Am. Nat. 1981, 117, 692–709. [Google Scholar] [CrossRef]
- Gilbert, L.E. Pollen feeding and reproductive biology of Heliconius butterflies. Proc. Natl. Acad. Sci. USA 1972, 69, 1403–1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, D.M.; Boggs, C.L.; Fogel, M.L. Pollen feeding in the butterfly Heliconius charitonia: Isotopic evidence for essential amino acid transfer from pollen to eggs. Proc. R. Soc. London. Ser. B Biol. Sci. 2003, 270, 2631–2636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slotten, J.R.; Miller, W. Occurrence of Eupyrrhoglossum sagra and Perigonia lusca in Florida (Lepidoptera: Sphingidae). Holarct. Lepid. 2000, 7, 59–63. [Google Scholar]
- Thomas, M.C. A Flower Beetle, Euphoria Sepulcralis (Fabricius) (Insecta: Coleoptera: Scarabaeidae); Entomology Circular 386; Florida Department of Agriculture and Consumer Services, Division of Plant Industry, UF/IFAS Extension: Gainesville, FL, USA, 2019; pp. 1–6. [Google Scholar]
- Jackson, R.R.; Pollard, S.D.; Nelson, X.J.; Edwards, G.B.; Barrion, A.T. Jumping spiders (Araneae: Salticidae) that feed on nectar. J. Zool. 2001, 255, 25–29. [Google Scholar] [CrossRef]
- Marquínez, X.; Cepeda, J.; Lara, K.; Sarmiento, R. Arañas asociadas a la floración de Drimys granadensis (Winteraceae). Rev. Colomb. Entomol. 2010, 36, 172–175. [Google Scholar] [CrossRef]
- Nahas, L.; Gonzaga, M.O.; Del-Claro, K. Wandering and web spiders feeding on the nectar from extrafloral nectaries in neotropical savanna. J. Zool. 2017, 301, 125–132. [Google Scholar] [CrossRef]
- Nyffeler, M.; Olson, E.J.; Symondson, W.O. Plant-eating by spiders. J. Arachnol. 2016, 44, 15–27. [Google Scholar] [CrossRef]
- Taylor, R.M.; Foster, W.A. Spider nectarivory. Am. Entomol. 1996, 42, 82–86. [Google Scholar] [CrossRef] [Green Version]
- Vogelei, A.; Greissl, R. Survival strategies of the crab spider Thomisus onustus Walckenaer 1806 (Chelicerata, Arachnida, Thomisidae). Oecologia 1989, 80, 513–515. [Google Scholar] [CrossRef] [PubMed]
- Dukas, R. Effects of perceived danger on flower choice by bees. Ecol. Lett. 2001, 4, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Morse, D.H. Choice of hunting site as a consequence of experience in late-instar crab spiders. Oecologia 1999, 120, 252–257. [Google Scholar] [CrossRef]
- Dukas, R.; Morse, D.H. Crab spiders affect flower visitation by bees. Oikos 2003, 101, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Llandres, A.L.; De Mas, E.; Rodriguez-Girones, M.A. Response of pollinators to the tradeoff between resource acquisition and predator avoidance. Oikos 2012, 121, 687–696. [Google Scholar] [CrossRef]
- Robertson, I.C.; Maguire, D.K. Crab spiders deter insect visitations to slickspot peppergrass flowers. Oikos 2005, 109, 577–582. [Google Scholar] [CrossRef]
- Romero, G.Q.; Antiqueira, P.A.; Koricheva, J. A meta-analysis of predation risk effects on pollinator behaviour. PLoS ONE 2011, 6, e20689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suttle, K.B. Pollinators as mediators of top-down effects on plants. Ecol. Lett. 2003, 6, 688–694. [Google Scholar] [CrossRef] [Green Version]
- Gentry, A.H. Anti-Pollinators for Mass-Flowering Plants? Biotropica 1978, 10, 68–69. [Google Scholar] [CrossRef]
- Hopkins, H.C.; Hopkins, M.J. Predation by a snake of a flower-visiting bat at Parkia nitida (Leguminosae: Mimosoideae). Brittonia 1982, 34, 225–227. [Google Scholar] [CrossRef]
- Romero, G.Q.; Vasconcellos-Neto, J. Beneficial effects of flower-dwelling predators on their host plant. Ecology 2004, 85, 446–457. [Google Scholar] [CrossRef]
- Carpenter, J.M. The genus Pachodynerus in North America (Hymenoptera: Vespidae: Eumeninae). Proc. Entomol. Soc. Wash. 1986, 88, 572–577. [Google Scholar]
- Krombein, K.V. Trap-Nesting Wasps and Bees: Life Histories, Nests and Associates; Smithsonian Press: Washington, DC, USA, 1967. [Google Scholar]
- Bradley, J.C. The species of Campsomeris (Hymenoptera-Scoliidae) of the Plumipes Group, inhabiting the United States, the Greater Antilles, and the Bahama Islands. Proc. Acad. Sci. Phila. 1928, 80, 313–337. [Google Scholar]
- Florida Natural Areas Inventory (FNAI). Pine Rockland. In Guide to the Natural Communities of Florida; Florida Natural Areas Inventory: Tallahassee, FL, USA, 2010; pp. 69–72. [Google Scholar]
- Koptur, S. The conservation of specialized and generalized pollination systems in subtropical ecosystems: A case study. In Plant–Pollinator Interactions: From Specialization to Generalization; Waser, N., Ollerton, J., Eds.; University of Chicago Press: Chicago, IL, USA, 2006; pp. 341–361. [Google Scholar]
- Snyder, J.R.; Herndon, A.; Robertson, W.B., Jr. South Florida Rockland. In Ecosystems of Florida; Myers, R.L., Ewel, J.J., Eds.; University of Central Florida Press: Orlando, FL, USA, 1990; pp. 230–274. [Google Scholar]
- Peña, A.L.; Koptur, S. A Historical Floristic Inventory of Pine Rockland Fabaceae (Leguminosae). Nat. Areas J. 2021, 41, 258–272. [Google Scholar] [CrossRef]
- Possley, J.E.; Maschinski, J.M.; Maguire, J.; Guerra, C. Vegetation Monitoring to Guide Management Decisions in Miami’s Urban Pine Rockland Preserves. Nat. Areas J. 2014, 34, 154–165. [Google Scholar] [CrossRef]
- Natureserve. 2022. Available online: https://explorer.natureserve.org/Taxon/ELEMENT_GLOBAL.2.723149/South_Florida_Pine_Rockland (accessed on 19 October 2022).
- Artz, D.R.; Waddington, K.D. The Effects of Neighbouring Tree Islands on Pollinator Density and Diversity, and on Pollination of a Wet Prairie Species, Asclepias lanceolata (Apocynaceae). J. Ecol. 2006, 94, 597–608. [Google Scholar] [CrossRef]
- Pascarella, J.B.; Waddington, K.D.; Neal, P.R. The Bee Fauna (Hymenoptera: Apoidea) of Everglades National Park, Florida and Adjacent Areas: Distribution, Phenology, and Biogeography. J. Kans. Entomol. Soc. 1999, 72, 32–45. [Google Scholar]
- Pascarella, J.B.; Waddington, K.D.; Neal, P.R. Non-apoid flower-visiting fauna of Everglades National Park, Florida. Biodivers. Conserv. 2001, 10, 551–566. [Google Scholar] [CrossRef]
- Pascarella, J.B. Pollination Ecology of Ardisia escallonioides (Myrsinaceae). Castanea 1997, 62, 1–7. [Google Scholar]
- Pascarella, J.B. Hurricane Disturbance, Plant-Animal Interactions, and the Reproductive Success of a Tropical Shrub. Biotropica 1998, 30, 416–424. [Google Scholar] [CrossRef]
- Cardel, Y.; Koptur, S. Effects of Florivory on the Pollination of Flowers: An Experimental Field Study with a Perennial Plant. Int. J. Plant Sci. 2010, 171, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Harris, B.M.; Koptur, S. Facilitated fecundity in sand flax: Pollination in an endangered herb of pine rocklands. Flora 2022, 289, 152041. [Google Scholar] [CrossRef]
- Linares, L.J.; Koptur, S. Floral Biology and Breeding System of the Crenulate Leadplant, Amorpha herbacea var. crenulata, an Endangered South Florida Pine Rockland Endemic. Nat. Areas J. 2010, 30, 138–147. [Google Scholar] [CrossRef]
- Liu, H.; Koptur, S. Breeding System and Pollination of a Narrowly Endemic Herb of the Lower Florida Keys: Impacts of the Urban Wildland Interface. Am. J. Bot. 2003, 90, 1180–1187. [Google Scholar] [CrossRef]
- Royal Botanic Gardens, Kew 2021. The World Checklist of Vascular Plants (WCVP). Checklist Dataset. Available online: https://www.gbif.org/dataset/f382f0ce-323a-4091-bb9f-add557f3a9a2 (accessed on 19 October 2022).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CLASS ORDER Family Species (Author) | Behavior on Plant | Resource Used | Study Site | |
|---|---|---|---|---|
| LPT | ENP | |||
| ARACHNIDA | ||||
| ARANEAE | ||||
| Araneidae | ||||
| Acacesia hamata Hentz | pr | ap | x | |
| Thomisidae | ||||
| Mecaphesa sp. 1 | pr | ap | x | |
| Mecaphesa sp. 2 | pr | ap | x | |
| Mecaphesa sp. 3 | pr | ap | x | |
| Mecaphesa sp. 4 | pr | ap | x | |
| INSECTA | ||||
| BLATTODEA | ||||
| Blattodea sp.1 | 2nr | n | x | |
| COLEOPTERA | ||||
| Cerambycidae | ||||
| Eburia stigma Oliver | lv | p | x | |
| Plectromerus dentipes Oliver | lv | p | x | |
| Scarabaeidae | ||||
| Euphoria sepulcralis Fabricius | lv | p | x | |
| Phyllophaga sp. | h | f | x | |
| DIPTERA | ||||
| Syrphidae | ||||
| Ornidia obesa Fabricius | lv | p | x | |
| Tipulidae | ||||
| Tipulidae sp.1 | 2nr | n | x | |
| Tipulidae sp.2 | 2nr | n | x | |
| HEMIPTERA | ||||
| Aphididae | ||||
| Aphididae sp.1 | h | f | x | |
| Largidae | ||||
| Largus succinctus Linnaeus | 2nr | n | x | x |
| HYMENOPTERA | ||||
| Apidae | ||||
| Apis mellifera Linnaeus | 2nr | n | x | |
| Euglosssa dilemma Bembé & Eltz | lv | n | x | |
| Xylocopa micans Lepeletier | 1nr | n | x | |
| Crabronidae | ||||
| Cerceris rufopicta Smith | 2nr | n | x | |
| Formicidae | ||||
| Camponotus floridanus Buckley | 2nr | n | x | |
| Pseudomyrmex gracilis Fabricius | 2nr | n | x | |
| Wasmannia auropunctata Roger | 2nr | n | x | |
| Halictidae | ||||
| Augochloropsis sp. | 2nr | n | x | |
| Scoliidae | ||||
| Dielis trifasciata Fabricius | lv | p | x | |
| Vespidae | ||||
| Mischocyttarus mexicanus cubicola Richards | 2nr | n | x | |
| Pachodynerus erynnis Lepeletier | 2nr | n | x | |
| Stenodynerus sp. | 1nr, 2nr | n | x | |
| Vespidae sp.1 | 1nr | n | x | |
| Zethus slossonae Fox | 1nr | n | x | x |
| LEPIDOPTERA | ||||
| Erebidae | ||||
| Calidota laqueata Edwards * | h | l | x | x |
| Hypercompe scriboni Stoll * | h | l | x | |
| Seirarctia echo Smith * | h | l | x | |
| Hesperiidae | ||||
| Asbolis capucinus Lucas | lv | n | x | x |
| Cymaenes tripunctus Herrich-Schäffer | lv | n | x | |
| Ephyriades brunnea Herrich-Schäffer | lv | n | x | x |
| Polites baracoa Lucas | lv | n | x | |
| Nymphalidae | ||||
| Agraulis vanillae Linnaeus | lv | n | x | x |
| Heliconius charithonia Linnaeus | lv | n | x | |
| Papilionidae | ||||
| Papilio palamedes Drury | lv | n | x | |
| Papilio polyxenes Fabricius | lv | n | x | |
| Sphingidae | ||||
| Aellopos tantalus Linnaeus | lv | n | x | |
| Eumorpha fasciatus Sulzer | lv | n | x | |
| Perigonia lusca Fabricius | lv | n | x | |
| Xylophanes tersa Linnaeus | lv | n | x | x |
| MANTODEA | ||||
| Mantidae | ||||
| Mantidae sp.1 | pr | ap | x | |
| Stagmomantis floridensis Davis | pr | ap | x | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pimienta, M.C.; Koptur, S. More than Moths: Flower Visitors of a Night-Blooming Plant in South Florida Pine Rocklands, USA. Plants 2022, 11, 2799. https://doi.org/10.3390/plants11202799
Pimienta MC, Koptur S. More than Moths: Flower Visitors of a Night-Blooming Plant in South Florida Pine Rocklands, USA. Plants. 2022; 11(20):2799. https://doi.org/10.3390/plants11202799
Chicago/Turabian StylePimienta, María Cleopatra, and Suzanne Koptur. 2022. "More than Moths: Flower Visitors of a Night-Blooming Plant in South Florida Pine Rocklands, USA" Plants 11, no. 20: 2799. https://doi.org/10.3390/plants11202799
APA StylePimienta, M. C., & Koptur, S. (2022). More than Moths: Flower Visitors of a Night-Blooming Plant in South Florida Pine Rocklands, USA. Plants, 11(20), 2799. https://doi.org/10.3390/plants11202799