
Dynamics of 2,4-D and Dicamba Applied to Corn Straw and Their Residual Action in Weeds


Abstract
:1. Introduction
2. Results
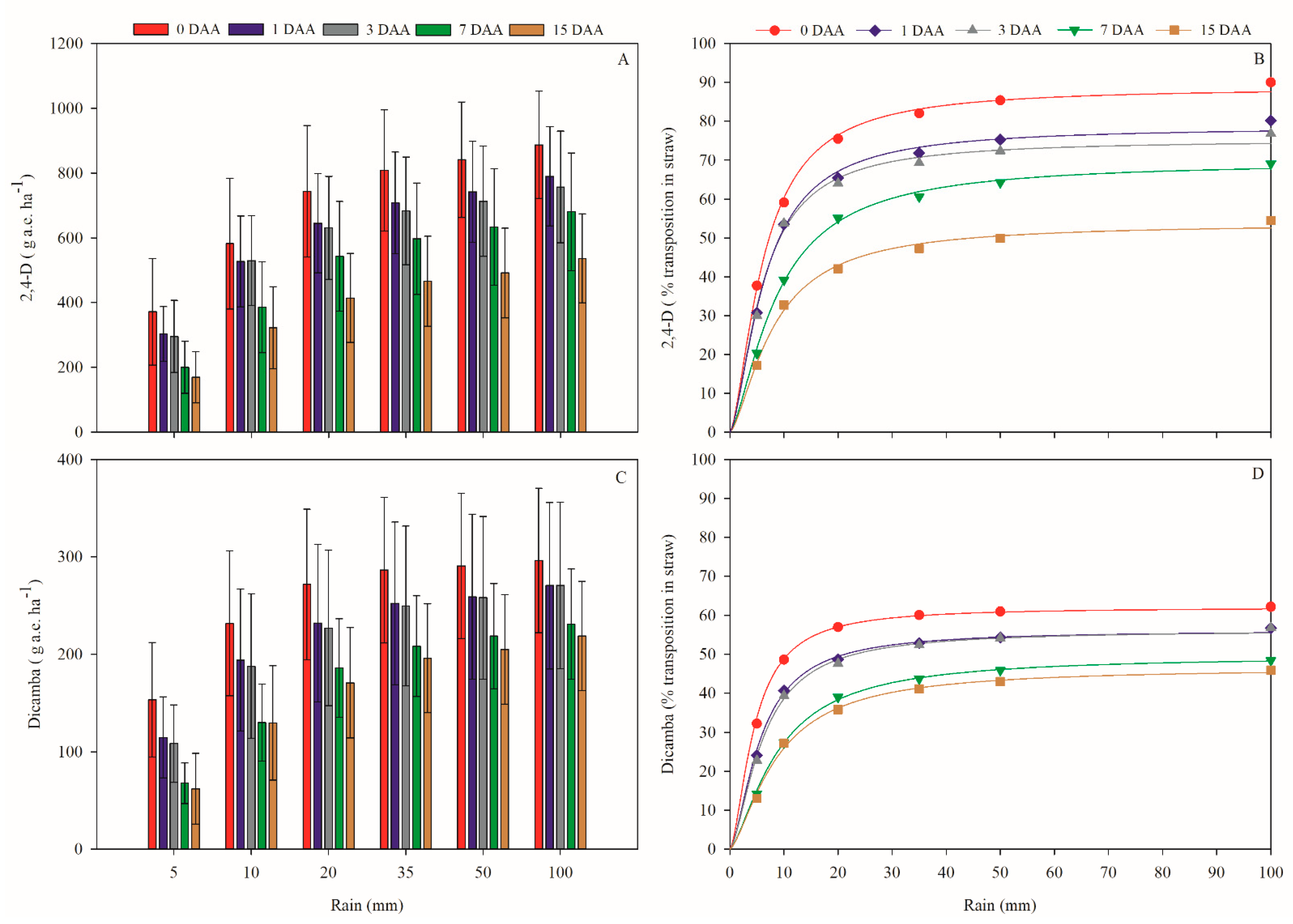
2.1. Dynamics of 2,4-D and Dicamba in Corn Straw
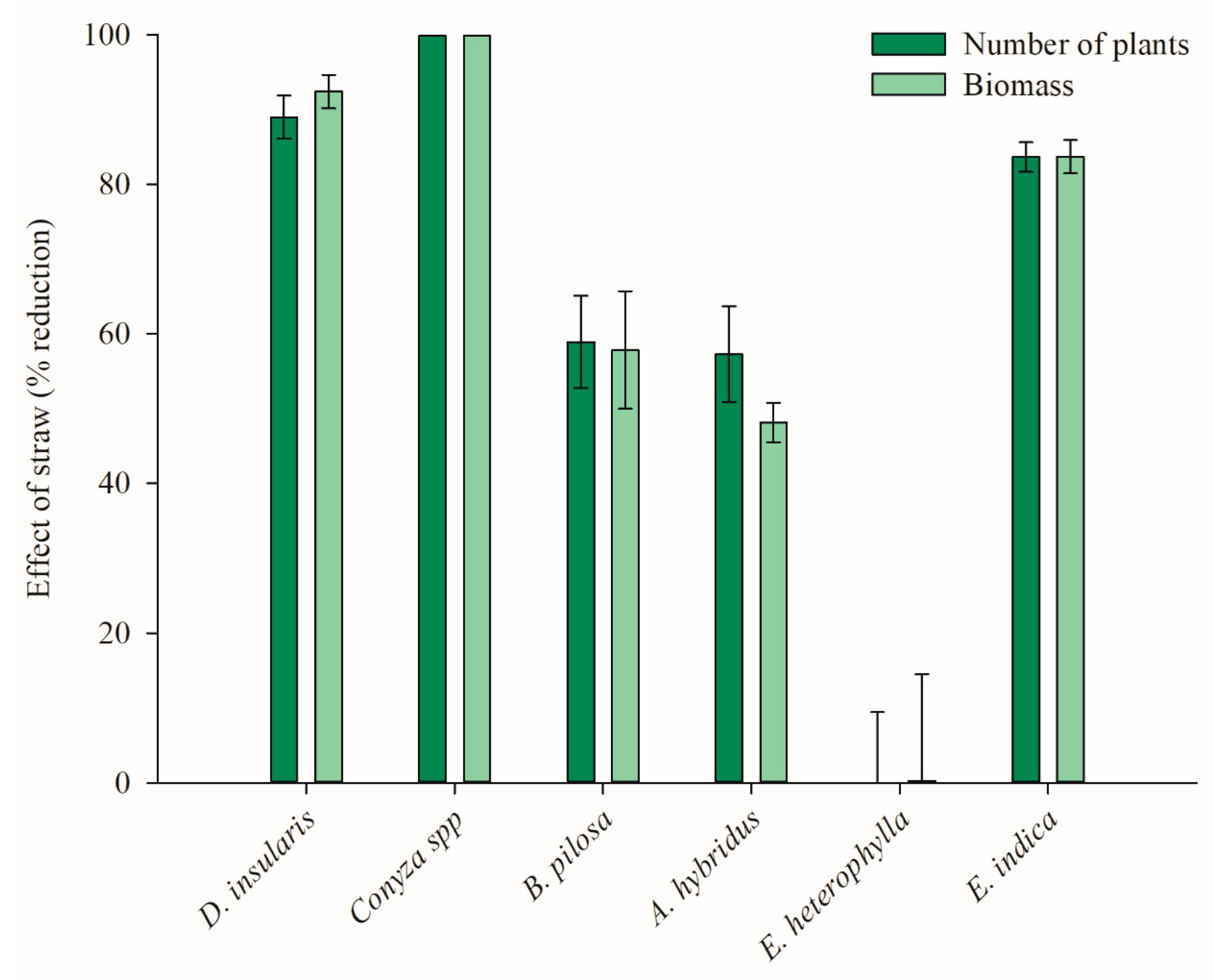
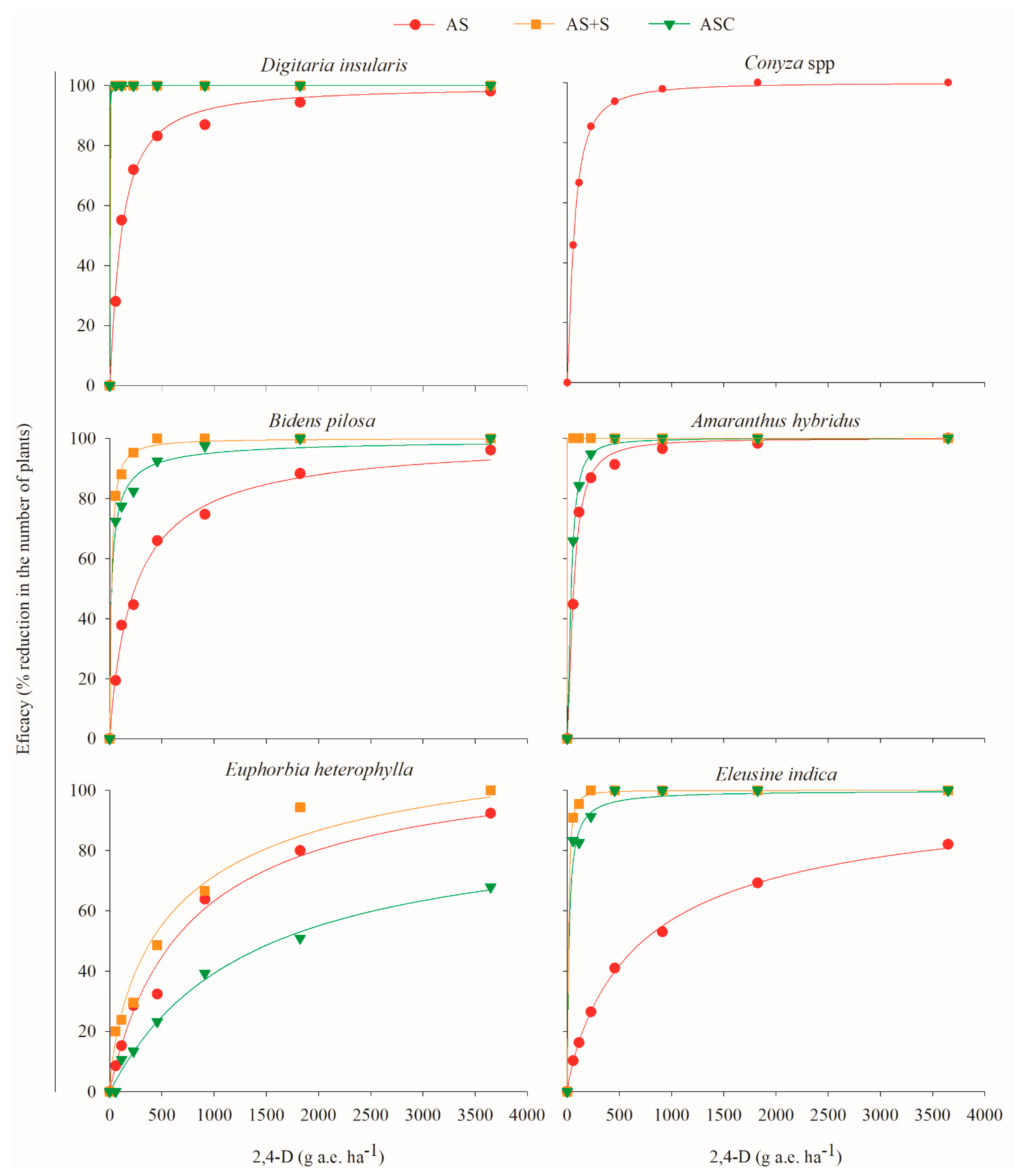
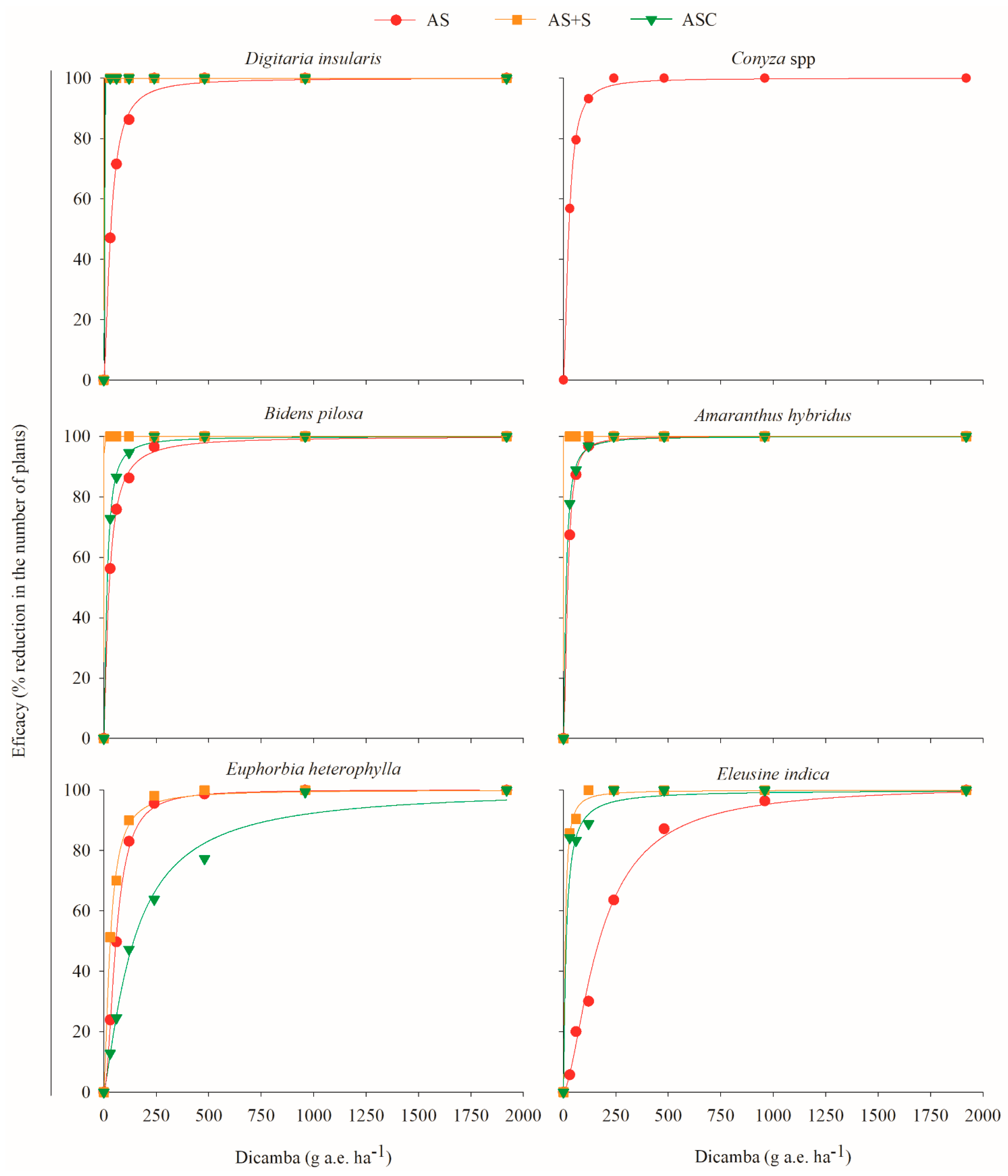
2.2. Effect of Straw and Residual Action of 2,4-D Choline Salt and Dicamba DGA Salt on Weeds
3. Discussion
4. Materials and Methods
4.1. Dynamics of 2,4-D Choline and DGA Salt in Corn Straw
4.2. Residual Action of 2,4-D Choline and Dicamba DGA Salt in Weeds
4.3. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Busi, R.; Goggin, D.E.; Heap, I.M.; Horak, M.J.; Jugulam, M.; Masters, R.A.; Napier, R.M.; Riar, D.S.; Satchivi, N.M.; Torra, J.; et al. Weed Resistance to Synthetic Auxin Herbicides. Pest Manag. Sci. 2018, 74, 2265–2276. [Google Scholar] [CrossRef] [PubMed]
- CTNBIO—Comissão Técnica Nacional de Biossegurança Resumo Geral de Plantas Geneticamente Modificadas Aprovadas Para Comercialização. Available online: http://ctnbio.mctic.gov.br/publicacoes/-/document_library_display/cwksGAQxt1lp/view/1684467 (accessed on 7 August 2022).
- Wiese, A.F.; Rea, H.E. Bindweed (Convolvulus Arvensis L.) Control and Seedling Emergence as Affected by Tillage, 24-D and Competitive Crops. Agron. J. 1959, 51, 672–675. [Google Scholar] [CrossRef]
- Hagood, E.S. Control of Triazine-Resistant Smooth Pigweed (Amaranthus Hybridus) and Common Lambsquarters (Chenopodium Album) in No-till Corn (Zea Mays). Weed Technol. 1989, 3, 136–142. [Google Scholar] [CrossRef]
- Bruce, J.A.; Kells, J.J. Horseweed (Conyza Canadensis) Control in No-Tillage Soybeans (Glycine Max) with Preplant and Preemergence Herbicides. Weed Technol. 1990, 4, 642–647. [Google Scholar] [CrossRef]
- Ou, J.; Thompson, C.R.; Stahlman, P.W.; Jugulam, M. Preemergence Application of Dicamba to Manage Dicamba-Resistant Kochia (Kochia Scoparia). Weed Technol. 2018, 32, 309–313. [Google Scholar] [CrossRef]
- Johnson, B.; Young, B.; Matthews, J.; Marquardt, P.; Slack, C.; Bradley, K.; York, A.; Culpepper, S.; Hager, A.; Al-Khatib, K.; et al. Weed Control in Dicamba-Resistant Soybeans. Crop Manag. 2010, 9, 1–23. [Google Scholar] [CrossRef]
- Velini, E.D. Comportamento de Herbicidas No Solo. In Proceedings of the Simpósio Nacional Sobre Manejo De Plantas Daninhas Em Hortaliças, Botucatu, Brazil, 27–31 August 2018; UNESP: Botucatu, Brazil, 1992; pp. 44–64. [Google Scholar]
- Locke, M.A.; Bryson, C.T. Herbicide-Soil Interaction in Reduced Tillage and Plant Residue Management Systems. Weed Sci. 1997, 45, 307–320. [Google Scholar] [CrossRef]
- Mundt, T.T.; Carbonari, C.A.; Moisinho, I.S.; de Brito, I.P.F.S.; Picoli Junior, G.J.; Ovejero, R.F.L.; Velini, E.D. Dynamics and Weed Control Effectiveness of Dicamba Herbicide When Applied Directly on the Soil and Corn Straw in Brazil. J. Environ. Sci. Health B 2022, 57, 1–11. [Google Scholar] [CrossRef]
- Rossi, C.V.S.; Velini, E.D.; Luchini, L.C.; Negrisoli, E.; Correa, M.R.; Pivetta, J.P.; Costa, A.G.F.; Silva, F.M.L. Dinâmica Do Herbicida Metribuzin Aplicado Sobre Palha de Cana-de-Açúcar (Saccarum Officinarum). Planta Daninha 2013, 31, 223–230. [Google Scholar] [CrossRef]
- de Matos, A.K.A.; Carbonari, C.A.; Gomes, G.L.G.C.; Velini, E.D. Dinâmica de Herbicidas Em Pré-Emergência Em Sistemas de Produção Com Palha. Rev. Bras. Herbic. 2016, 15, 97. [Google Scholar] [CrossRef]
- Carbonari, C.A.; Velini, E.D.; Correa, M.R.; Negrisoli, E.; Rossi, C.V.; Oliveira, C.P. Efeitos de Períodos de Permanência de Clomazone + Hexazinona No Solo e Na Palha de Cana-de-Açúcar Antes Da Ocorrência de Chuvas Na Eficácia de Controle de Plantas Daninhas. Planta Daninha 2010, 28, 197–205. [Google Scholar] [CrossRef] [Green Version]
- Carbonari, C.A.; Gomes, G.L.G.C.; Trindade, M.L.B.; Silva, J.R.M.; Velini, E.D. Dynamics of Sulfentrazone Applied to Sugarcane Crop Residues. Weed Sci. 2016, 64, 201–206. [Google Scholar] [CrossRef]
- Lewis, K.A.; Tzilivakis, J.; Warner, D.J.; Green, A. An International Database for Pesticide Risk Assessments and Management. Hum. Ecol. Risk Assess. Int. J. 2016, 22, 1050–1064. [Google Scholar] [CrossRef] [Green Version]
- Comfort, S.D.; Inskeep, W.P.; Macur, R.E. Degradation and Transport of Dicamba in a Clay Soil. J. Environ. Qual. 1992, 21, 653–658. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: New York, NY, USA, 1984. [Google Scholar]
- Egley, G.H.; Duke, S.O. Physiology of Weed Seed Dormancy and Germination. In Weed Physiology: Reproduction and Ecophysiology; Duke, S.O., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 27–64. [Google Scholar]
- Velini, E.D.; Negrisoli, E. Controle de Plantas Daninhas Em Cana Crua. In Proceedings of the Congresso Brasileiro Da Ciência Das Plantas Daninhas, Foz do Iguaçu, Brazil, 25–28 July 2020; SBCPD: Londrina, Brazil, 2000; p. 55. [Google Scholar]
- Beyaert, R.P.; Schott, J.W.; White, P.H. Tillage Effects on Corn Production in a Coarse-Textured Soil in Southern Ontario. Agron. J. 2002, 94, 767–774. [Google Scholar] [CrossRef]
- Correia, N.M.; Durigan, J.C. Emergência de Plantas Daninhas Em Solo Coberto Com Palha de Cana-de-Açúcar. Planta Daninha 2004, 22, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Gravena, R.; Rodrigues, J.P.R.G.; Spindola, W.; Pitelli, R.A.; Alves, P.L.C.A. Controle de Plantas Daninhas Através Da Palha de Cana-de-Açúcar Associada à Mistura Dos Herbicidas Trifloxysulfuron Sodium + Ametrina. Planta Daninha 2004, 22, 419–427. [Google Scholar] [CrossRef] [Green Version]
- Nagabhusana, G.C.; Worsham, A.D.; Yenish, J.P. Allelopathic Cover Crops to Reduce Herbicide Use in Sustainable Agricultural Systems. Allelopath. J. 2001, 8, 133–146. [Google Scholar]
- Azania, A.A.P.M.; Azania, C.A.M.; Gravena, R.; Pavani, M.C.M.D.; Pitelli, R.A. Interferência Da Palha de Cana-de-Açúcar (Saccharum Spp.) Na Emergência de Espécies de Plantas Daninhas Da Família Convolvulaceae. Planta Daninha 2002, 20, 207–212. [Google Scholar] [CrossRef]
- Trezzi, M.M.; Vidal, R.A. Potencial de Utilização de Cobertura Vegetal de Sorgo e Milheto Na Supressão de Plantas Daninhas Em Condição de Campo: II—Efeitos Da Cobertura Morta. Planta Daninha 2004, 22, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Coelho, S.P.; Galvão, J.C.C.; Trogello, E.; Campos, S.A.; Pereira, L.P.L.; Barrella, T.P.; Cecon, P.R.; Pereira, A.J. Coberturas Vegetais Na Supressão de Plantas Daninhas Em Sistema de Plantio Direto Orgânico de Milho. Rev. Bras. Milho Sorgo 2016, 15, 65–72. [Google Scholar] [CrossRef]
- Tofoli, G.R.; Velini, E.D.; Negrisoli, E.; Cavenaghi, A.L.; Martins, D. Dinâmica Do Tebuthiuron Em Palha de Cana-de-Açúcar. Planta Daninha 2009, 27, 815–821. [Google Scholar] [CrossRef]
- Maciel, C.D.G.; Velini, E.D. Simulação Do Caminhamento Da Água Da Chuva e Herbicidas Em Palhadas Utilizadas Em Sistemas de Plantio Direto. Planta Daninha 2005, 23, 471–481. [Google Scholar] [CrossRef]
- Zhou, X.; Rotondaro, S.L.; Ma, M.; Rosser, S.W.; Olberding, E.L.; Wendelburg, B.M.; Adelfinskaya, Y.A.; Balcer, J.L.; Blewett, T.C.; Clements, B. Metabolism and Residues of 2,4-Dichlorophenoxyacetic Acid in DAS-40278-9 Maize (Zea Mays) Transformed with Aryloxyalkanoate Dioxygenase-1 Gene. J. Agric. Food Chem. 2016, 64, 7438–7444. [Google Scholar] [CrossRef] [PubMed]
- Wright, T.R.; Shan, G.; Walsh, T.A.; Lira, J.M.; Cui, C.; Song, P.; Zhuang, M.; Arnold, N.L.; Lin, G.; Yau, K.; et al. Robust Crop Resistance to Broadleaf and Grass Herbicides Provided by Aryloxyalkanoate Dioxygenase Transgenes. Proc. Natl. Acad. Sci. USA 2010, 107, 20240–20245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrens, M.R.; Mutlu, N.; Chakraborty, S.; Dumitru, R.; Jiang, W.Z.; LaVallee, B.J.; Herman, P.L.; Clemente, T.E.; Weeks, D.P. Dicamba Resistance: Enlarging and Preserving Biotechnology-Based Weed Management Strategies. Science 2007, 316, 1185–1188. [Google Scholar] [CrossRef] [Green Version]
- FEBRAPDP—Federação Brasileira do Sistema Plantio Direto Área Sob Plantio Direto. Available online: https://febrapdp.org.br/area-de-pd (accessed on 8 August 2022).
- Selim, H.M.; Zhou, L.; Zhu, H. Herbicide Retention in Soil as Affected by Sugarcane Mulch Residue. J. Environ. Qual. 2003, 32, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Lamoreaux, R.J.; Jain, R.; Hess, F.D. Efficacy of Dimethenamid, Metolachlor and Encapsulated Alachlor in Soil Covered with Crop Residue, 3rd ed.; British Crop Protection Council (BCPC): Gilroy, CA, USA, 1993. [Google Scholar]
- Watts, D.; Hall, J.K. Tillage and Application Effects on Herbicide Leaching and Runoff. Soil Tillage Res. 1996, 39, 241–257. [Google Scholar] [CrossRef]
- Watkins, D.A.M. Some Implications of the Photochemical Decomposition of Pesticides. Chem. Ind. 1974, 5, 185–190. [Google Scholar]
- Reichardt, K. Dinâmica Da Matéria Orgânica e Da Energia Em Ecossistemas, 2nd ed.; Esalq-Depto de Física e Meteorologia: Piracicaba, Brazil, 1996. [Google Scholar]
- Radosevich, S.; Holt, J.; Ghersa, C. Weed Ecology: Implications for Management, 2nd ed.; Wiley: New York, NY, USA, 1997. [Google Scholar]
- Bailey, G.W.; White, J.L. Soil-Pesticide Relationships, Adsorption and Desorption of Organic Pesticides by Soil Colloids, with Implications Concerning Pesticide Bioactivity. J. Agric. Food Chem. 1964, 12, 324–332. [Google Scholar] [CrossRef]
- Dadazio, T.S. Obtenção e Análise de Espectros de Absorção de Luz Por Herbicidas. Ph.D. Thesis, Universidade Estadual Paulista Júlio de Mesquita Filho, Botucatu, Brazil, 2018. [Google Scholar]
- Campbell, P.J. 2,4-Dichlorophenoxyacetic Acid (2,4-D), Salts and Esters. Available online: http://www.inchem.org/documents/jmpr/jmpmono/v097pr16.htm (accessed on 8 August 2022).
- Sosnoskie, L.M.; Culpepper, A.S.; Braxton, L.B.; Richburg, J.S. Evaluating the Volatility of Three Formulations of 2,4-D When Applied in the Field. Weed Technol. 2015, 29, 177–184. [Google Scholar] [CrossRef]
- Mueller, T.C.; Steckel, L.E. Dicamba Volatility in Humidomes as Affected by Temperature and Herbicide Treatment. Weed Technol. 2019, 33, 541–546. [Google Scholar] [CrossRef]
- Bish, M.D.; Farrell, S.T.; Lerch, R.N.; Bradley, K.W. Dicamba Losses to Air after Applications to Soybean under Stable and Nonstable Atmospheric Conditions. J. Environ. Qual. 2019, 48, 1675–1682. [Google Scholar] [CrossRef] [Green Version]
- Sosnoskie, L.M.; Culpepper, A.S.; Braxton, L.B.; Richburg, J.S. Comparing Volatility of Three Forms of 2,4-D When Applied in the Field. In Proceedings of the Annual Meeting of the Weed, Waikoloa, HI, USA, 6–9 February 2012; WSSA Abstract: Lawrence, KS, USA, 2012; p. 333. [Google Scholar]
- Egan, J.F.; Mortensen, D.A. Quantifying Vapor Drift of Dicamba Herbicides Applied to Soybean. Environ. Toxicol. Chem. 2012, 31, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Mueller, T.C.; Wright, D.R.; Remund, K.M. Effect of Formulation and Application Time of Day on Detecting Dicamba in the Air under Field Conditions. Weed Sci. 2013, 61, 586–593. [Google Scholar] [CrossRef]
- Hemminghaus, J.W.; Macinnes, A.; Zhang, J. Low Volatility Herbicidal Compositions. WO2014071374A2, 8 May 2014. [Google Scholar]
- Abraham, W. The Chemistry Behind Low-Volatility Dicamba. Available online: https://monsanto.com/app/uploads/2018/07/Chemistry-Behind-Low-Vol-Dicamba_Abraham.pdf (accessed on 15 October 2021).
- Carbonari, C.A.; Costa, R.N.; Giovanelli, B.F.; Bevilaqua, N.C.; Palhano, M.; Barbosa, H.; Lopez Ovejero, R.F.; Velini, E.D. Volatilization of Dicamba Diglycolamine Salt in Combination with Glyphosate Formulations and Volatility Reducers in Brazil. Agronomy 2022, 12, 1001. [Google Scholar] [CrossRef]
- Behrens, R.; Lueschen, W.E. Dicamba Volatility. Weed Sci. 1979, 27, 486–493. [Google Scholar] [CrossRef]
- Carbonari, C.A.; Costa, R.N.; Bevilaqua, N.C.; Pereira, V.G.C.; Giovanelli, B.F.; Lopez Ovejero, R.F.; Palhano, M.; Barbosa, H.; Velini, E.D. Volatilization of Standalone Dicamba and Dicamba Plus Glyphosate as Function of Volatility Reducer and Different Surfaces. Agriculture 2020, 10, 495. [Google Scholar] [CrossRef]
- Kerle, E.A.; Vogue, P.A.; Jenkins, J.J. Understanding Pesticide Persistence and Mobility for Groundwater and Surface Water Protection; Oregon State University: Corvalis, OR, USA, 2007. [Google Scholar]
- Correia, N.M.; Rezende, P.M. Manejo Integrado de Plantas Daninhas Na Cultura Da Soja. Available online: http://www.editora.ufla.br/upload/boletim/tecnico/boletim-tecnico-51.pdf (accessed on 7 August 2022).
- Reddy, K.N.; Locke, M.A.; Wagner, S.C.; Zablotowicz, R.M.; Gaston, L.A.; Smeda, R.J. Chlorimuron Ethyl Sorption and Desorption Kinetics in Soils and Herbicide-Desiccated Cover Crop Residues. J. Agric. Food Chem. 1995, 43, 2752–2757. [Google Scholar] [CrossRef]
- Dao, T.H. Field Decay of Wheat Straw and Its Effects on Metribuzin and S-Ethyl Metribuzin Sorption and Elution from Crop Residues. J. Environ. Qual. 1991, 20, 203–208. [Google Scholar] [CrossRef]
- Heap, I. The International Herbicide-Resistant Weed Database. Available online: http://www.weedscience.org (accessed on 8 August 2022).
- Chang, F.Y.; vanden Born, W.H. Dicamba Uptake, Translocation, Metabolism, and Selectivity. Weed Sci. 1971, 19, 113–117. [Google Scholar] [CrossRef]
- Peterson, M.A.; McMaster, S.A.; Riechers, D.E.; Skelton, J.; Stahlman, P.W. 2,4-D Past, Present, and Future: A Review. Weed Technol. 2016, 30, 303–345. [Google Scholar] [CrossRef]
- Sterling, T.M.; Hall, J.C. Mechanism of Action of Natural Auxins and the Auxinic Herbicides. In Herbicide Activity: Toxicology, Biochemistry and Molecular Biology; Roe, R.M., Burton, J.D., Kuhr, R.J., Eds.; IOS Press: Amsterdam, The Netherlands, 1997; pp. 111–141. [Google Scholar]
- De Veylder, L.; Joubès, J.; Inzé, D. Plant Cell Cycle Transitions. Curr. Opin. Plant Biol. 2003, 6, 536–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewitte, W.; Murray, J.A.H. The Plant Cell Cycle. Annu. Rev. Plant Biol. 2003, 54, 235–264. [Google Scholar] [CrossRef] [Green Version]
- Sauer, M.; Robert, S.; Kleine-Vehn, J. Auxin: Simply Complicated. J. Exp. Bot. 2013, 64, 2565–2577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, X.; Calderon-Villalobos, L.I.A.; Sharon, M.; Zheng, C.; Robinson, C.V.; Estelle, M.; Zheng, N. Mechanism of Auxin Perception by the TIR1 Ubiquitin Ligase. Nature 2007, 446, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Grossmann, K. Auxin Herbicide Action: Lifting the Veil Step by Step. Plant Signal. Behav. 2007, 2, 421–423. [Google Scholar] [CrossRef] [Green Version]
- Bewley, J.D.; Black, M. Seeds: Physiology of Development and Germination, 2nd ed.; Plenum Press: New York, NY, USA, 1994. [Google Scholar]
- Hattley, H.O. The Use of Range in Analysis of Variance. Biometrika 1950, 37, 271–280. [Google Scholar] [CrossRef]
- Mitscherlich, E.A. Das Gesetz Des Minimums Und Das Gesetz Des Abnehmenden Bodenertrages. Landwirtschaftliches Jahrbuch 1909, 25, 537–552. [Google Scholar]
- Streibig, J.C. Herbicide Bioassay. Weed Res. 1988, 28, 479–484. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | 2,4-D | Dicamba |
|---|---|---|
| Chemical group | phenoxy-carboxylic acid | benzoic acid |
| Chemical structure |  |  |
| Molecular formula | C8H6Cl2O3 | C8H6Cl2O3 |
| Solubility in water, 20 °C (mg L−1) | 24,300 | 250,000 |
| Dissociation constant—pKa at 25 °C | 3.4 | 1.87 |
| Octanol-water partition coefficient at pH 7, 20 °C (Log P) | −0.82 | −1.88 |
| Soil adsorption coefficient (Koc) (L kg−1) | 39.3 | 1.42 |
| Henry’s law constant at 25 °C (Pa m³ mol⁻¹) | 4.0 × 10−6 | 5.06 × 10−5 |
| Vapour pressure, 20 °C (mPa) | 0.009 | 1.67 |
| Half-life (days) | 4.4 | 9.62 |
| Days without Rain | Model Parameters | F Value | |||
|---|---|---|---|---|---|
| a | b | c | R² | ||
| 2,4-D choline salt | |||||
| 0 | 86.3723 | 0 | 0.0485 | 0.9992 | 2656.9100 ** |
| 1 | 76.4020 | 0 | 0.0469 | 0.9985 | 1334.6100 ** |
| 3 | 73.3179 | 0 | 0.0500 | 0.9982 | 1147.8600 ** |
| 7 | 66.7652 | 0 | 0.0356 | 0.9987 | 1553.2900 ** |
| 15 | 51.7479 | 0 | 0.0381 | 0.9980 | 1003.4900 ** |
| Dicamba salt DGA | |||||
| 0 | 61.0288 | 0 | 0.0662 | 0.9987 | 11,330.2000 ** |
| 1 | 54.8658 | 0 | 0.0531 | 0.9991 | 2809.9900 ** |
| 3 | 54.7578 | 0 | 0.0495 | 0.9989 | 2313.0500 ** |
| 7 | 47.5389 | 0 | 0.0351 | 0.9992 | 3242.8700 ** |
| 15 | 44.6515 | 0 | 0.0356 | 0.9984 | 1573.5600 ** |
| Species | Application | Model Parameters | F Value | CL80 | CL90 | |||
|---|---|---|---|---|---|---|---|---|
| a | b | X0 | R2 | |||||
| 2,4-D choline salt | ||||||||
| Digitaria insularis | AS | 100.8917 | 1.1257 | 107.1734 | 0.9924 | 30.14 ** | 367.2140 | 754.7010 |
| AS + S | 100 | 1.9752 | 0.0418 | 0.8347 | 40.39 ** | 0.0843 | 0.1271 | |
| ASC | 100 | 1.9752 | 0.0390 | 0.8759 | 56.47 ** | 0.0787 | 0.1186 | |
| Conyza spp. | AS | 99.8945 | 1.3821 | 65.9158 | 0.9995 | 74.47 ** | 179.7220 | 323.1610 |
| Bidens pilosa | AS | 99.6850 | 0.9410 | 242.2398 | 0.8843 | 65.67 ** | 1056.9500 | 2505.1800 |
| AS + S | 99.9880 | 1.1655 | 17.1909 | 0.8957 | 69.50 ** | 56.4763 | 113.2500 | |
| ASC | 99.9188 | 0.7508 | 18.5254 | 0.8035 | 33.84 ** | 117.3980 | 345.7290 | |
| Amaranthus hybridus | AS | 100.301 | 1.4895 | 61.6281 | 0.9421 | 142.26 ** | 156.3080 | 269.4170 |
| AS + S | 100.000 | 1.9465 | 0.0290 | 0.9603 | 193.58 ** | 0.0591 | 0.0896 | |
| ASC | 99.9764 | 1.6085 | 38.361 | 0.9243 | 98.41 ** | 90.8216 | 150.3630 | |
| Euphorbia heterophylla | AS | 96.574 | 1.2701 | 657.7352 | 0.9030 | 88.09 ** | 1959.1900 | 3709.9200 |
| AS + S | 91.3852 | 1.3934 | 507.0584 | 0.8608 | 62.97 ** | 1371.3200 | 2454.0800 | |
| ASC | 101.2237 | 0.8922 | 1579.679 | 0.9922 | 24.16 ** | 7470.9000 | 18,539.900 | |
| Eleusine indica | AS | 99.6114 | 0.888 | 732.8578 | 0.9987 | 61.69 ** | 3491.5300 | 8701.9600 |
| AS + S | 99.9991 | 1.5233 | 12.8008 | 0.9997 | 23.65 ** | 31.8032 | 54.1587 | |
| ASC | 99.9616 | 0.9884 | 17.4681 | 0.9950 | 8.90 ** | 71.0185 | 161.3200 | |
| Dicamba salt DGA | ||||||||
| Digitaria insularis | AS | 99.8782 | 1.5825 | 32.9172 | 0.9972 | 23.85 ** | 79.0445 | 131.953 |
| AS + S | 100 | 1.8838 | 0.0189 | 1.0000 | 7.63 ** | 0.0394 | 0.0606 | |
| ASC | 100 | 1.8953 | 0.0404 | 1.0000 | 6.52 ** | 0.0839 | 0.1287 | |
| Conyza spp. | AS | 99.9489 | 1.6951 | 25.8267 | 0.9992 | 19.90 ** | 58.5121 | 94.4084 |
| Bidens pilosa | AS | 99.9519 | 1.2979 | 24.949 | 0.9984 | 22.89 ** | 72.5981 | 135.6040 |
| AS + S | 100 | 1.96 | 0.0173 | 0.9041 | 75.46 ** | 0.0350 | 0.0530 | |
| ASC | 99.9847 | 1.4081 | 15.0433 | 0.9993 | 10.42 ** | 40.2636 | 71.6183 | |
| Amaranthus hybridus | AS | 99.9881 | 1.8351 | 20.2642 | 0.9108 | 81.87 ** | 43.1330 | 67.1003 |
| AS + S | 100 | 1.9676 | 0.0187 | 0.9528 | 161.61 ** | 0.0378 | 0.0512 | |
| ASC | 99.99 | 1.4423 | 12.8027 | 0.9994 | 60.16 ** | 33.4759 | 58.7372 | |
| Euphorbia heterophylla | AS | 99.2256 | 2.0074 | 57.7511 | 0.9401 | 128.51 ** | 115.2070 | 172.5530 |
| AS + S | 99.7938 | 1.4993 | 30.4375 | 0.9971 | 72.66 ** | 76.7308 | 131.7860 | |
| ASC | 99.3794 | 1.2855 | 143.741 | 0.8483 | 48.03 ** | 422.5980 | 794.133 | |
| Eleusine indica | AS | 97.6066 | 1.8034 | 180.5425 | 0.8749 | 58.30 ** | 389.4280 | 610.5410 |
| AS + S | 99.9931 | 1.3438 | 8.2821 | 0.9978 | 32.84 ** | 23.2364 | 42.4862 | |
| ASC | 99.965 | 1.1119 | 13.2203 | 0.9962 | 9.18 ** | 45.9950 | 95.3784 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gazola, T.; Costa, R.N.; Carbonari, C.A.; Velini, E.D. Dynamics of 2,4-D and Dicamba Applied to Corn Straw and Their Residual Action in Weeds. Plants 2022, 11, 2800. https://doi.org/10.3390/plants11202800
Gazola T, Costa RN, Carbonari CA, Velini ED. Dynamics of 2,4-D and Dicamba Applied to Corn Straw and Their Residual Action in Weeds. Plants. 2022; 11(20):2800. https://doi.org/10.3390/plants11202800
Chicago/Turabian StyleGazola, Tiago, Renato Nunes Costa, Caio Antonio Carbonari, and Edivaldo Domingues Velini. 2022. "Dynamics of 2,4-D and Dicamba Applied to Corn Straw and Their Residual Action in Weeds" Plants 11, no. 20: 2800. https://doi.org/10.3390/plants11202800
APA StyleGazola, T., Costa, R. N., Carbonari, C. A., & Velini, E. D. (2022). Dynamics of 2,4-D and Dicamba Applied to Corn Straw and Their Residual Action in Weeds. Plants, 11(20), 2800. https://doi.org/10.3390/plants11202800