
Phytochemicals and Inflammation: Is Bitter Better?


Abstract
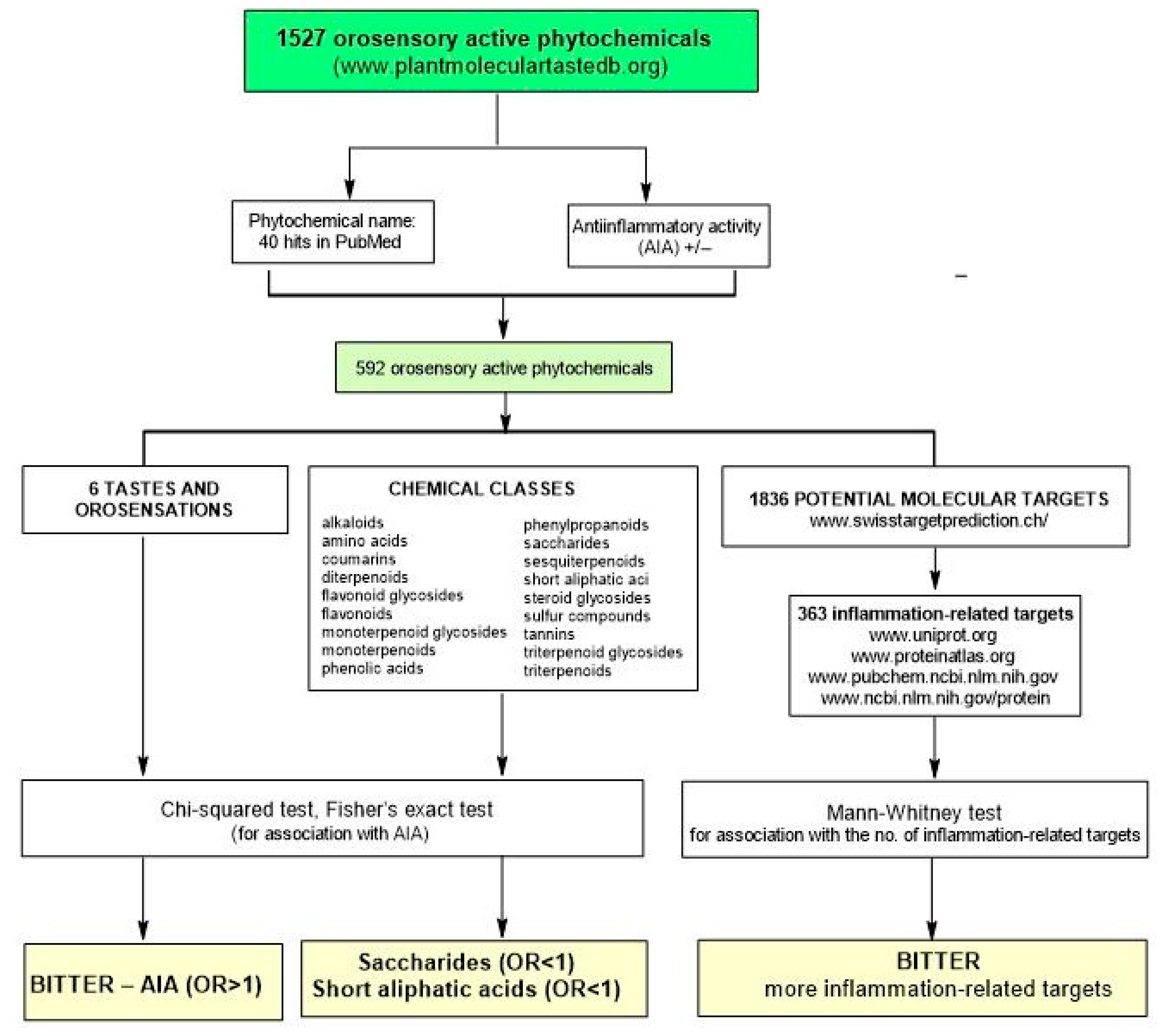
:1. Introduction
2. Results
3. Discussion
3.1. For a Given Phytochemical, Is the Taste Responsible for the Anti-Inflammatory Activity or the Lack Thereof?
3.2. Are Certain Bitter Taste Receptors Involved in the Anti-Inflammatory Activity?
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saqib, S.; Ullah, F.; Naeem, M.; Younas, M.; Ayaz, A.; Ali, S.; Zaman, W. Mentha: Nutritional and Health Attributes to Treat Various Ailments Including Cardiovascular Diseases. Molecules 2022, 27, 6728. [Google Scholar] [CrossRef] [PubMed]
- Gilca, M.; Barbulescu, A. Taste of medicinal plants: A potential tool in predicting ethnopharmacological activities? J. Ethnopharmacol. 2015, 174, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Leonti, M.; Sticher, O.; Heinrich, M. Medicinal plants of the Popoluca, México: Organoleptic properties as indigenous selection criteria. J. Ethnopharmacol. 2002, 81, 307–315. [Google Scholar] [CrossRef]
- Molares, S.; Ladio, A. Medicinal plants in the cultural landscape of a Mapuche-Tehuelche community in arid Argentine Patagonia: An eco-sensorial approach. J. Ethnobiol. Ethnomedicine 2014, 10, 61. [Google Scholar] [CrossRef] [Green Version]
- Ankli, A.; Sticher, O.; Heinrich, M. Yucatec Maya Medicinal Plants Versus Nonmedicinal Plants: Indigenous Characterization and Selection. Hum. Ecol. 1999, 27, 557–580. [Google Scholar] [CrossRef]
- Shepard, G.H., Jr. A Sensory Ecology of Medicinal Plant Therapy in Two Amazonian Societies. Am. Anthr. 2004, 106, 252–266. [Google Scholar] [CrossRef]
- Lu, P.; Zhang, C.-H.; Lifshitz, L.M.; ZhuGe, R. Extraoral bitter taste receptors in health and disease. J. Gen. Physiol. 2017, 149, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Behrens, M.; Meyerhof, W. Gustatory and extragustatory functions of mammalian taste receptors. Physiol. Behav. 2011, 105, 4–13. [Google Scholar] [CrossRef]
- Laffitte, A.; Neiers, F.; Briand, L. Functional roles of the sweet taste receptor in oral and extraoral tissues. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 379–385. [Google Scholar] [CrossRef] [Green Version]
- Gilca, M.; Dragos, D. Extraoral Taste Receptor Discovery: New Light on Ayurvedic Pharmacology. Evidence-Based Complement. Altern. Med. 2017, 2017, 5435831. [Google Scholar] [CrossRef]
- Giuliani, C.; Franceschi, C.; Luiselli, D.; Garagnani, P.; Ulijaszek, S. Ecological Sensing Through Taste and Chemosensation Mediates Inflammation: A Biological Anthropological Approach. Adv. Nutr. Int. Rev. J. 2020, 11, 1671–1685. [Google Scholar] [CrossRef] [PubMed]
- Montinari, M.R.; Minelli, S.; de Caterina, R. The first 3500 years of aspirin history from its roots—A concise summary. Vasc. Pharmacol. 2019, 113, 1–8. [Google Scholar] [CrossRef]
- Baranwal, M.; Gupta, Y.; Dey, P.; Majaw, S. Antiinflammatory phytochemicals against virus-induced hyperinflammatory responses: Scope, rationale, application, and limitations. Phytotherapy Res. 2021, 35, 6148–6169. [Google Scholar] [CrossRef] [PubMed]
- Niu, W.-H.; Wu, F.; Cao, W.-Y.; Wu, Z.-G.; Chao, Y.-C.; Peng, F.; Liang, C. Network pharmacology for the identification of phytochemicals in traditional Chinese medicine for COVID-19 that may regulate interleukin. Biosci. Rep. 2021, 41, BSR20202583. [Google Scholar] [CrossRef] [PubMed]
- Dragos, D.; Gilca, M. Taste of phytocompounds: A better predictor for ethnopharmacological activities of medicinal plants than the phytochemical class? J. Ethnopharmacol. 2018, 220, 129–146. [Google Scholar] [CrossRef]
- Gradinaru, T.-C.; Petran, M.; Dragos, D.; Gilca, M. PlantMolecularTasteDB: A Database of Taste Active Phytochemicals. Front. Pharmacol. 2022, 12, 3804. [Google Scholar] [CrossRef]
- Dragos, D.; Petran, M.; Gilca, M. Bitter taste and astringency- predictors of antiinflammatory activity? In Proceedings of the 4th International Conference on Natural Products Utilization from Plants to Pharmacy Shelf, Albena, Bulgaria, 29 May–1 June 2019. [Google Scholar]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef] [Green Version]
- Wiener, A.; Shudler, M.; Levit, A.; Niv, M.Y. BitterDB: A database of bitter compounds. Nucleic Acids Res. 2012, 40, D413–D419. [Google Scholar] [CrossRef]
- Dagan-Wiener, A.; Di Pizio, A.; Nissim, I.; Bahia, M.S.; Dubovski, N.; Margulis, E.; Niv, M.Y. BitterDB: Taste ligands and receptors database in 2019. Nucleic Acids Res. 2019, 47, D1179–D1185. [Google Scholar] [CrossRef]
- Ahmed, J.; Preissner, S.; Dunkel, M.; Worth, C.; Eckert, A. SuperSweet—A resource on natural and artificial sweetening agents. Nucleic Acids Res. 2011, 39, D377–D382. [Google Scholar] [CrossRef]
- Dragos, D.; Petran, M.; Gradinaru, T.; Gilca, M.P. Do bitter phytocompounds target inflammation-related macromolecules? In Proceedings of the 20th International Congress of International Society for Ethnopharmacology, Virtual Congress, 18–20 April 2021; p. 247. [Google Scholar]
- Bensky, D.; Clavey, S.; Stoger, E. Chinese Herbal Medicine Materia Medica, 3rd ed.; Eastland Press: Seattle, WA, USA, 2004. [Google Scholar]
- Sharma, R.; Dash, B. Caraka Samhita, 2006th ed.; Chowkhamba Sanskrit Series Office: Varanasi, India, 2006. [Google Scholar]
- Patwardhan, B.; Warude, D.; Pushpangadan, P.; Bhatt, N. Ayurveda and Traditional Chinese Medicine: A Comparative Overview. Evidence-Based Complement. Altern. Med. 2005, 2, 465–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, K.; Hankey, A.; Patwardhan, B. Traditional Phytochemistry: Identification of Drug by ‘Taste’. Evidence-Based Complement. Altern. Med. 2007, 4, 145–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murthy, S.K.R. Vagbhata’s Astanga Hrdaya, 2nd ed.; Krishnadas Academy: Varanasi, India, 1994. [Google Scholar]
- Kurihara, K. Umami the Fifth Basic Taste: History of Studies on Receptor Mechanisms and Role as a Food Flavor. BioMed Res. Int. 2015, 2015, 189402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowe, M.S.; Wang, H.; Blakeney, B.A.; Mahavadi, S.; Singh, K.; Murthy, K.S.; Grider, J.R. Expression and function of umami receptors T1R1/T1R3 in gastric smooth muscle. Neurogastroenterol. Motil. 2020, 32, e13737. [Google Scholar] [CrossRef]
- Shi, J.; Wang, S.; Ke, Q.; Lin, J.; Zheng, Y.; Wu, S.; Huang, Z.; Lin, W. T1R1/T1R3 Taste Receptor Suppresses Granulocyte-Mediated Neuroinflammation after Spinal Cord Injury. J. Neurotrauma 2017, 34, 2353–2363. [Google Scholar] [CrossRef]
- Liu, S.; Xu, M.; Zhu, C.; Zhao, Q.; Zhou, F. Taste receptor T1R1/T1R3 promotes the tumoricidal activity of hepatic CD49a+CD49b−natural killer cells. Eur. J. Immunol. 2018, 48, 2031–2041. [Google Scholar] [CrossRef] [Green Version]
- Canivenc-Lavier, M.-C.; Neiers, F.; Briand, L. Plant polyphenols, chemoreception, taste receptors and taste management. Curr. Opin. Clin. Nutr. Metab. Care 2019, 22, 472–478. [Google Scholar] [CrossRef]
- Bayer, S.; Mayer, A.I.; Borgonovo, G.; Morini, G.; Di Pizio, A.; Bassoli, A. Chemoinformatics View on Bitter Taste Receptor Agonists in Food. J. Agric. Food Chem. 2021, 69, 13916–13924. [Google Scholar] [CrossRef]
- O’Connor, L.; Imamura, F.; Brage, S.; Griffin, S.J.; Wareham, N.J.; Forouhi, N.G. Intakes and sources of dietary sugars and their association with metabolic and inflammatory markers. Clin. Nutr. 2018, 37, 1313–1322. [Google Scholar] [CrossRef] [Green Version]
- Laffin, M.; Fedorak, R.; Zalasky, A.; Park, H.; Gill, A.; Agrawal, A.; Keshteli, A.; Hotte, N.; Madsen, K.L. A high-sugar diet rapidly enhances susceptibility to colitis via depletion of luminal short-chain fatty acids in mice. Sci. Rep. 2019, 9, 12294. [Google Scholar] [CrossRef]
- Sakamoto, N.; Kono, S.; Wakai, K.; Fukuda, Y.; Satomi, M.; Shimoyama, T.; Inaba, Y.; Miyake, Y.; Sasaki, S.; Okamoto, K.; et al. Dietary Risk Factors for Inflammatory Bowel Disease: A multicenter case-control study in Japan. Inflamm. Bowel Dis. 2005, 11, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Porto, M.L.; Lírio, L.M.; Dias, A.T.; Batista, A.T.; Campagnaro, B.P.; Mill, J.G.; Meyrelles, S.S.; Baldo, M.P. Increased oxidative stress and apoptosis in peripheral blood mononuclear cells of fructose-fed rats. Toxicol. Vitr. 2015, 29, 1977–1981. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Sun, T. Glycyrrhizin administration ameliorates Streptococcus aureus-induced acute lung injury. Int. Immunopharmacol. 2019, 70, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wu, W.; Li, W.; Huang, S.; Li, Z.; Liu, R.; Shan, Z.; Zhang, C.; Wang, S. Activation of NLRP3 Inflammasome Promotes Foam Cell Formation in Vascular Smooth Muscle Cells and Atherogenesis Via HMGB. J. Am. Hearth Assoc. 2018, 7, e008596. [Google Scholar] [CrossRef] [Green Version]
- Chung, J.-H.; Kong, J.-N.; Choi, H.-E.; Kong, K.-H. Antioxidant, anti-inflammatory, and anti-allergic activities of the sweet-tasting protein brazzein. Food Chem. 2018, 267, 163–169. [Google Scholar] [CrossRef]
- Monier-Williams, M. A Sanskrit-English Dictionary, 15th ed.; Motilal Banarsidass: Delhi, India, 2002. [Google Scholar]
- Pandey, G. Dravyaguṇa Vijñāna: Materia Medica-Vegetable Drugs: English-Sanskrit; Krishnadas Academy: Varanasi, India, 1998. [Google Scholar]
- Peier, A.M.; Moqrich, A.; Hergarden, A.C.; Reeve, A.J.; Andersson, D.A.; Story, G.M.; Earley, T.J.; Dragoni, I.; McIntyre, P.; Bevan, S.; et al. A TRP channel that senses cold stimuli and menthol. Cell 2002, 108, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Kuroki, S.; Hagura, N.; Nishida, S.; Haggard, P.; Watanabe, J. Sanshool on The Fingertip Interferes with Vibration Detection in a Rapidly-Adapting (RA) Tactile Channel. PLoS ONE 2016, 11, e0165842. [Google Scholar] [CrossRef] [Green Version]
- Klein, A.H.; Carstens, M.I.; Carstens, E. Eugenol and carvacrol induce temporally desensitizing patterns of oral irritation and enhance innocuous warmth and noxious heat sensation on the tongue. Pain 2013, 154, 2078–2087. [Google Scholar] [CrossRef] [Green Version]
- Nilius, B.; Owsianik, G.; Voets, T.; Peters, J.A. Transient receptor potential cation channels in disease. Physiol. Rev. 2007, 87, 165–217. [Google Scholar] [CrossRef] [Green Version]
- Gouin, O.; L’Herondelle, K.; Lebonvallet, N.; Le Gall-Ianotto, C.; Sakka, M.; Buhé, V.; Plée-Gautier, E.; Carré, J.-L.; Lefeuvre, L.; Misery, L.; et al. TRPV1 and TRPA1 in cutaneous neurogenic and chronic inflammation: Pro-inflammatory response induced by their activation and their sensitization. Protein Cell 2017, 8, 644–661. [Google Scholar] [CrossRef]
- Kameda, T.; Zvick, J.; Vuk, M.; Sadowska, A.; Tam, W.K.; Leung, V.Y.; Bölcskei, K.; Helyes, Z.; Applegate, L.A.; Hausmann, O.N.; et al. Expression and Activity of TRPA1 and TRPV1 in the Intervertebral Disc: Association with Inflammation and Matrix Remodeling. Int. J. Mol. Sci. 2019, 20, 1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramachandran, R.; Hyun, E.; Zhao, L.; Lapointe, T.K.; Chapman, K.; Hirota, C.L.; Ghosh, S.; McKemy, D.D.; Vergnolle, N.; Beck, P.L.; et al. TRPM8 activation attenuates inflammatory responses in mouse models of colitis. Proc. Natl. Acad. Sci. USA 2013, 110, 7476–7481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stampanoni Bassi, M.; Gentile, A.; Iezzi, E.; Zagaglia, S.; Musella, A.; Simonelli, I.; Gilio, L.; Furlan, R.; Finardi, A.; Marfia, G.A.; et al. Transient Receptor Potential Vanilloid 1 Modulates Central Inflammation in Multiple Sclerosis. Front. Neurol. 2019, 10, 30. [Google Scholar] [CrossRef] [Green Version]
- Silverman, H.A.; Chen, A.; Kravatz, N.L.; Chavan, S.S.; Chang, E.H. Involvement of Neural Transient Receptor Potential Channels in Peripheral Inflammation. Front Immunol. 2020, 11, 2742. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, F.; Aono, H. Role of Transient Receptor Potential Vanilloid 1 in Inflammation and Autoimmune Diseases. Pharmaceuticals 2012, 5, 837–852. [Google Scholar] [CrossRef] [Green Version]
- Calcinoni, O.; Borgonovo, G.; Cassanelli, A.; Banfi, E.; Bassoli, A. Herbs for Voice Database: Developing a Rational Approach to the Study of Herbal Remedies Used in Voice Care. J. Voice 2021, 35, 807.e33–807.e41. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Sun, W.; Feng, X.; Ba, X.; Liu, T.; Guo, J.; Xiao, L.; Jiang, J.; Hao, Y.; Xiong, D.; et al. (−)-menthol increases excitatory transmission by activating both TRPM8 and TRPA1 channels in mouse spinal lamina II layer. Biochem. Biophys. Res. Commun. 2019, 516, 825–830. [Google Scholar] [CrossRef]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef] [Green Version]
- Ekoff, M.; Choi, J.-H.; James, A.; Dahlén, B.; Nilsson, G.; Dahlén, S.-E. Bitter taste receptor (TAS2R) agonists inhibit IgE-dependent mast cell activation. J. Allergy Clin. Immunol. 2014, 134, 475–478. [Google Scholar] [CrossRef]
- Grassin-Delyle, S.; Salvator, H.; Mantov, N.; Abrial, C.; Brollo, M.; Faisy, C.; Naline, E.; Couderc, L.J.; Devillier, P. Bitter Taste Receptors (TAS2Rs) in Human Lung Macrophages: Receptor Expression and Inhibitory Effects of TAS2R Agonists. Front. Physiol. 2019, 10, 1267. [Google Scholar] [CrossRef]
- Yuan, Y.; Fang, X.; Ye, W. Acrid and Bitter Chinese Herbs in Decoction Effectively Relieve Lung Inflammation and Regulation of TRPV1/TAS2R14 Channels in a Rat Asthmatic Model. Evidence-Based Complement. Altern. Med. 2022, 2022, 8061740. [Google Scholar] [CrossRef] [PubMed]
- Tiroch, J.; Sterneder, S.; Di Pizio, A.; Lieder, B.; Hoelz, K.; Holik, A.-K.; Pignitter, M.; Behrens, M.; Somoza, M.; Ley, J.P.; et al. Bitter Sensing TAS2R50 Mediates the trans-Resveratrol-Induced Anti-inflammatory Effect on Interleukin 6 Release in HGF-1 Cells in Culture. J. Agric. Food Chem. 2021, 69, 13339–13349. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Xi, R.; Liu, J.; Peng, X.; Zhao, L.; Zhou, X.; Li, J.; Zheng, X.; Xu, X. TAS2R16 Activation Suppresses LPS-Induced Cytokine Expression in Human Gingival Fibroblasts. Front. Immunol. 2021, 12, 726546. [Google Scholar] [CrossRef] [PubMed]
- Meyerhof, W.; Batram, C.; Kuhn, C.; Brockhoff, A.; Chudoba, E.; Bufe, B.; Appendino, G.B.; Behrens, M. The Molecular Receptive Ranges of Human TAS2R Bitter Taste Receptors. Chem. Senses 2010, 35, 157–170. [Google Scholar] [CrossRef]
- Tran, H.T.T.; Stetter, R.; Herz, C.; Spöttel, J.; Krell, M.; Hanschen, F.S.; Schreiner, M.; Rohn, S.; Behrens, M.; Lamy, E. Allyl Isothiocyanate: A TAS2R38 Receptor-Dependent Immune Modulator at the Interface Between Personalized Medicine and Nutrition. Front. Immunol. 2021, 12, 1255. [Google Scholar] [CrossRef]
- Karaman, R.; Nowak, S.; Di Pizio, A.; Kitaneh, H.; Abu-Jaish, A.; Meyerhof, W.; Niv, M.Y.; Behrens, M. Probing the binding pocket of the broadly tuned human bitter taste receptor TAS2R14 by chemical modification of cognate agonists. Chem. Biol. Drug Des. 2016, 88, 66–75. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2018, 47, D506–D515. [Google Scholar] [CrossRef] [Green Version]
- The Human Protein Atlas. Available online: https://www.proteinatlas.org/ (accessed on 18 September 2022).
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem substance and compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef]
- Fleiss, J.L. The Design and Analysis of Clinical Experiments; John Wiley & Sons: Hoboken, NJ, USA, 1986. [Google Scholar]
- Altman, D.G. Practical Statistics for Medical Research; Chapman and Hall: London, UK, 1991. [Google Scholar]
- Trecca, E.M.; Fortunato, F.; Gelardi, M.; Petrone, P.; Cassano, M. Development of a questionnaire to investigate socio-cultural differences in the perception of smell, taste and flavour. Acta Otorhinolaryngol. Ital. 2021, 41, 336–347. [Google Scholar] [CrossRef]
- Diószegi, J.; Llanaj, E.; Ádány, R. Genetic Background of Taste Perception, Taste Preferences, and Its Nutritional Implications: A Systematic Review. Front. Genet. 2019, 10, 1272. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Taste | Counts (a, b, c, d) | Chi-Square Statistics | p-Value by Chi-Square | p-Value by Fisher’s Exact Test | Odds Ratio (OR) | 95% Confidence Interval for OR |
|---|---|---|---|---|---|---|
| astringent | (73, 14, 379, 126) | 2.7535 | 0.097 | 0.077 | 1.73 | 0.93–3.44 |
| bitter | (322, 68, 130, 72) | 23.435 | 1.3 × 10−6 | 1.4-06 | 2.62 | 1.74–3.94 |
| pungent | (58, 20, 394, 120) | 0.090859 | 0.76 | 0.67 | 0.88 | 0.5–1.62 |
| salty | (1, 2, 451, 138) | N/A | N/A | 0.14 | 0.15 | 0.003–2.97 |
| sour | (24, 18, 428, 122) | 8.1283 | 0.004 | 0.0042 | 0.38 | 0.19–0.77 |
| sweet | (62, 32, 390, 108) | 6.019 | 0.014 | 0.012 | 0.54 | 0.33–0.9 |
| umami | (4, 2, 448, 138) | N/A | N/A | 0.63 | 0.62 | 0.09–6.89 |
| pungent not-bitter | (13, 52, 38, 42) | 11.8943 | 0.00056 | 0.0008 | 0.28 | 0.13–0.58 |
| bitter (flavonoids only) | (53, 4, 6, 2) | N/A | N/A | 0.16 | 4.27 | 0.32–38.32 |
| bitter (flavonoid glycosides only) | (33, 4, 16, 2) | N/A | N/A | 1 | 1.09 | 0.09–8.56 |
| Chemical Class | Counts (a, b, c, d) | Chi-Square Statistics | p-Value by Chi-Square | p-Value by Fisher’s Exact Test | Odds Ratio (OR) | 95% Confidence Interval for OR |
|---|---|---|---|---|---|---|
| alkaloids | (69, 31, 383, 109) | 3.1281 | 0.077 | 0.07 | 0.63 | 0.39–1.06 |
| amino acids | (22, 6, 430, 134) | 0.003071 | 0.96 | 1 | 1.14 | 0.44–3.52 |
| coumarins | (9, 2, 443, 138) | N/A | N/A | 1 | 1.4 | 0.29–13.48 |
| diterpenoids | (12, 1, 440, 139) | N/A | N/A | 0.32 | 3.79 | 0.55–163.1 |
| flavonoid glycosides | (51, 6, 401, 134) | 5.2378 | 0.022 | 0.013 | 2.84 | 1.18–8.27 |
| flavonoids | (59, 6, 393, 134) | 7.5332 | 0.0061 | 0.003 | 3.35 | 1.41–9.7 |
| monoterpenoid glycosides | (12, 1, 440, 139) | N/A | N/A | 0.32 | 3.79 | 0.55–163.1 |
| monoterpenoids | (32, 5, 420, 135) | 1.6864 | 0.19 | 0.16 | 2.06 | 0.77–6.89 |
| phenolic acids | (10, 3, 442, 137) | N/A | N/A | 1 | 1.03 | 0.26–5.92 |
| Phenylpropanoids * | (12, 2, 440, 138) | N/A | N/A | 0.54 | 1.88 | 0.41–17.5 |
| saccharides | (14, 16, 438, 124) | 13.739 | 0.00021 | 0.0003 | 0.25 | 0.11–0.56 |
| sesquiterpenoids | (19, 4, 433, 136) | 0.22099 | 0.64 | 0.62 | 1.49 | 0.48–6.13 |
| short aliphatic acids | (3, 7, 449, 133) | N/A | N/A | 0.0023 | 0.13 | 0.02–0.57 |
| steroid glycosides | (7, 5, 445, 135) | N/A | N/A | 0.17 | 0.43 | 0.11–1.73 |
| sulfur compounds | (9, 6, 443, 134) | N/A | N/A | 0.14 | 0.45 | 0.14–1.58 |
| tannins | (13, 1, 439, 139) | N/A | N/A | 0.21 | 4.11 | 0.61–176.05 |
| triterpenoid glycosides | (7, 2, 445, 138) | N/A | N/A | 1 | 1.09 | 0.2–10.83 |
| triterpenoids | (19, 4, 433, 136) | 0.22099 | 0.64 | 0.62 | 1.49 | 0.48–6.13 |
| Taste | Median [q1–q3] for Yes | Median [q1–q3] for No | W Statistics | p-Value |
|---|---|---|---|---|
| bitter | 25 [21–28] | 20 [14–26] | 19917 | 3 × 10−8 |
| astringent | 23.5 [19–27] | 24 [20–28] | 20372.5 | 0.2 |
| salty | 16.5 [12.25–20.75] | 24 [20–28] | 2565 | 0.02 |
| pungent | 23.5 [20–28.75] | 24 [19.5–28] | 17727.5 | 0.6 |
| sour | 15 [6–21] | 24 [20–28] | 20315.5 | 3 × 10−14 |
| Sweet | 19 [14–24] | 25 [21–29] | 31957.5 | 4–12 |
| Biological Activity | A. Number of Articles Found in PubMed at the Search: Name of the Biological Activity and Phytochemical | Year of the First Published Paper for the Search A | B. Number of Articles Found in PubMed at the Search: Name of the Biological Activity | Year of the First Published Paper for the Search B |
|---|---|---|---|---|
| Antimicrobial OR anti-microbial OR antiinfectious OR anti-infectious OR antiinfective OR infection preventing OR antibacterial OR antiviral OR antiparasitic OR antifungal | 8363 | 1946 | 2,506,227 | 1802 |
| Anticancer OR antineoplastic OR antiproliferative OR cancer preventing | 8032 | 1954 | 1,480,830 | 1934 |
| Anti-inflammatory OR anti-inflammatory OR antiphlogistic OR inflammation preventing | 6449 | 1952 | 756,116 | 1826 |
| Antihypertensive OR hypotensive | 640 | 1959 | 379,075 | 1913 |
| Cardiotonic OR “heart tonic” | 290 | 1954 | 215,217 | 1945 |
| Diuretic | 283 | 1956 | 105,834 | 1807 |
| Immunostimulant OR immune enhancer | 279 | 1972 | 429,982 | 1899 |
| Choleretic | 233 | 1972 | 30,051 | 1945 |
| Cholagogue | 230 | 1972 | 29,742 | 1852 |
| Antacid | 12 | 1969 | 27,690 | 1931 |
| AIA | No AIA | |
|---|---|---|
| Taste/Chemical class | a | b |
| Not Taste/Chemical class | c | d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dragoș, D.; Petran, M.; Gradinaru, T.-C.; Gilca, M. Phytochemicals and Inflammation: Is Bitter Better? Plants 2022, 11, 2991. https://doi.org/10.3390/plants11212991
Dragoș D, Petran M, Gradinaru T-C, Gilca M. Phytochemicals and Inflammation: Is Bitter Better? Plants. 2022; 11(21):2991. https://doi.org/10.3390/plants11212991
Chicago/Turabian StyleDragoș, Dorin, Madalina Petran, Teodora-Cristiana Gradinaru, and Marilena Gilca. 2022. "Phytochemicals and Inflammation: Is Bitter Better?" Plants 11, no. 21: 2991. https://doi.org/10.3390/plants11212991
APA StyleDragoș, D., Petran, M., Gradinaru, T. -C., & Gilca, M. (2022). Phytochemicals and Inflammation: Is Bitter Better? Plants, 11(21), 2991. https://doi.org/10.3390/plants11212991