

The Peach (Prunus persica) CBL and CIPK Family Genes: Protein Interaction Profiling and Expression Analysis in Response to Various Abiotic Stresses
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Characteristics of the Peach CBL and CIPK Gene Families
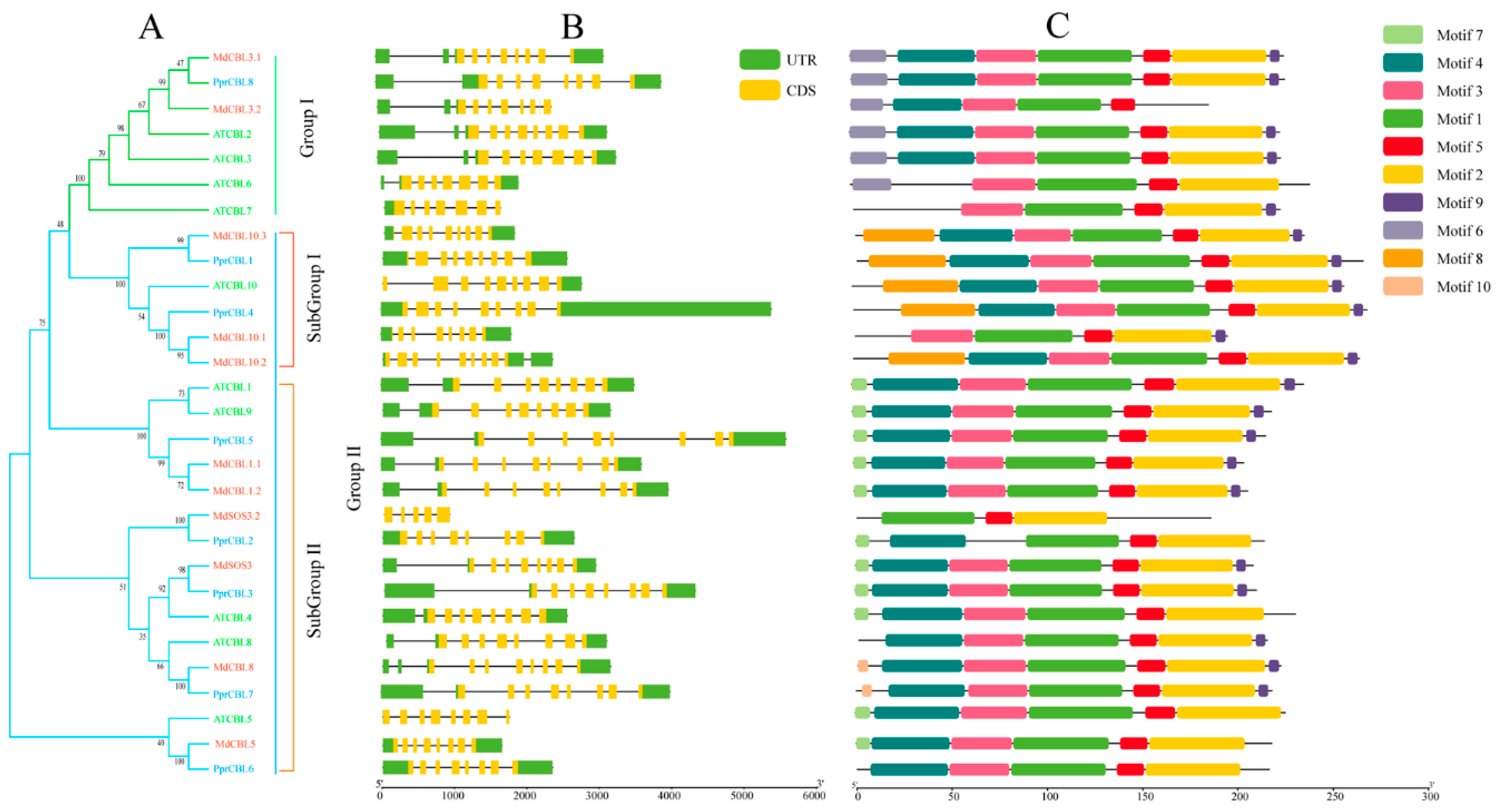
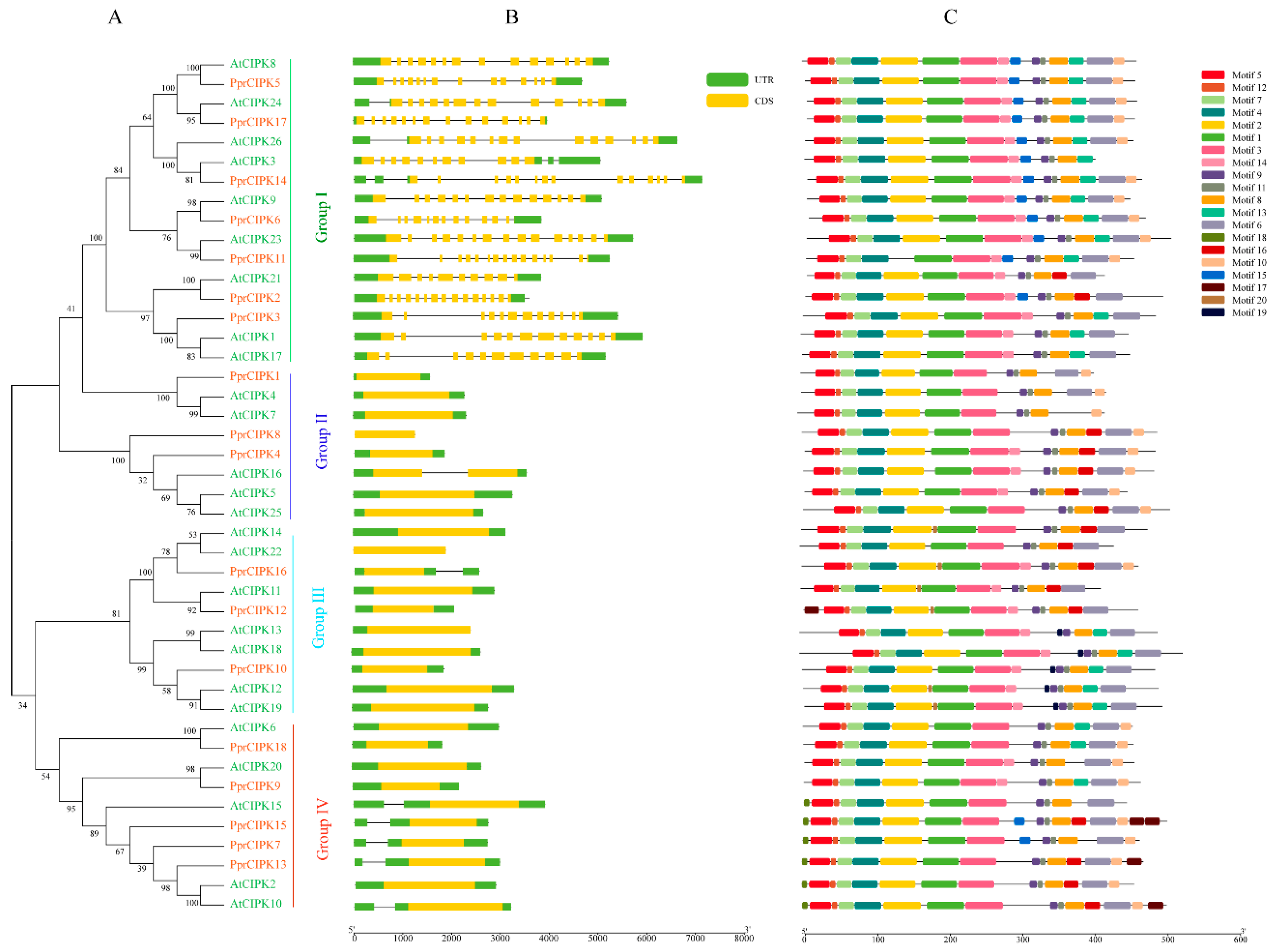
2.2. Phylogenetic, Structure and Motifs Analysis of PprCBL and PprCIPK Family Genes
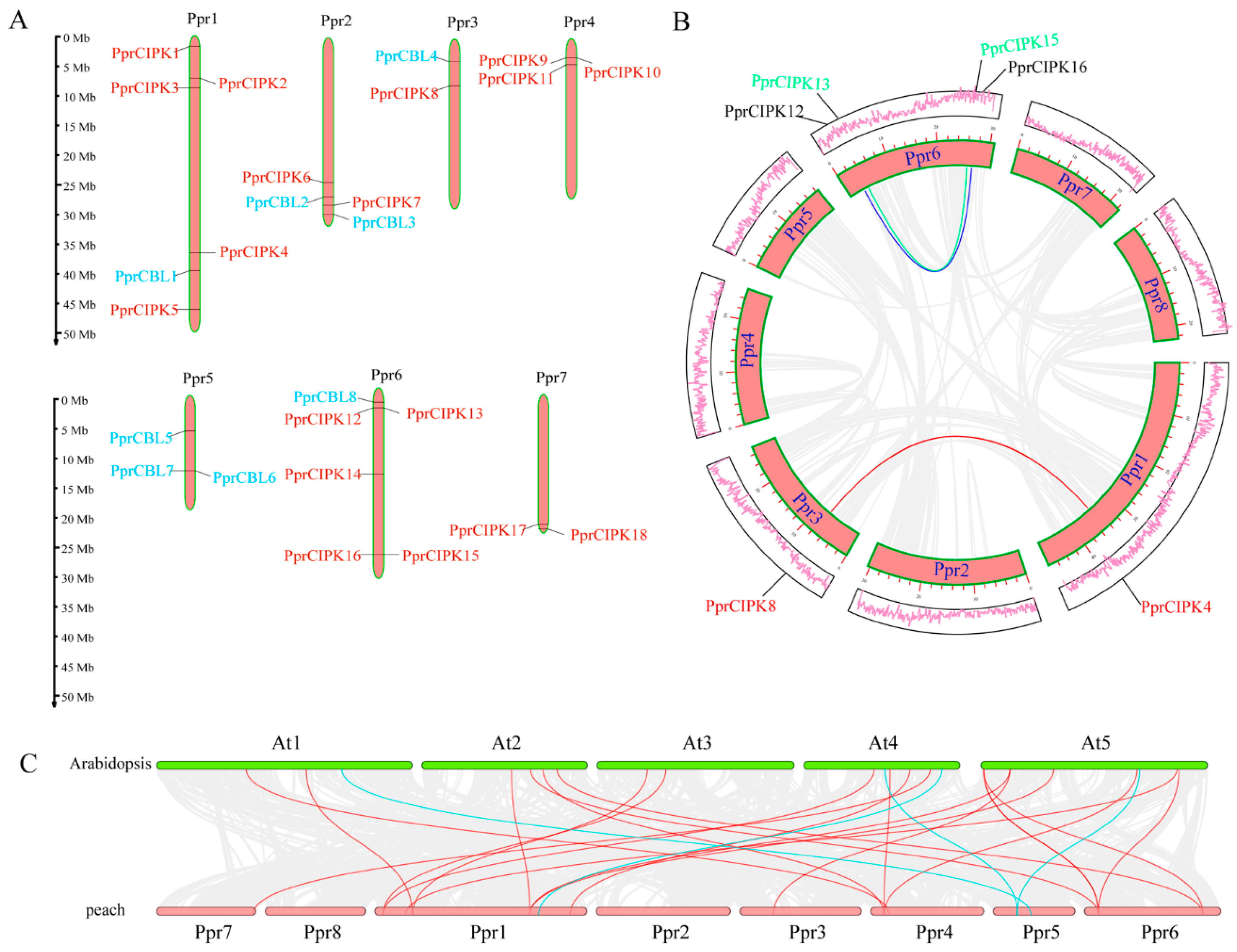
2.3. Expansion Patterns of the Peach CIPK Gene Family
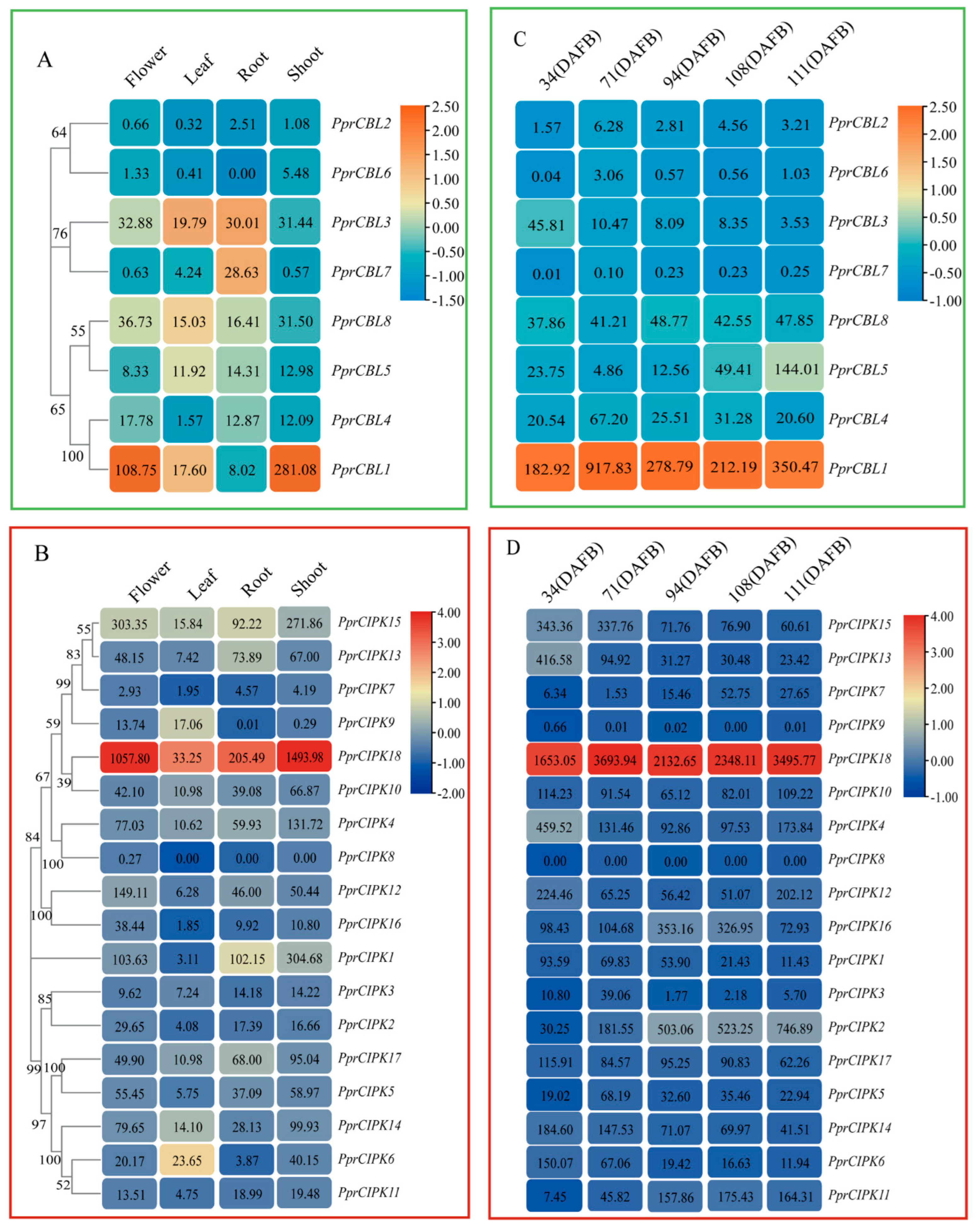
2.4. Expression Profiles of Peach CBL and CIPK Genes in Different Organs and Fruit Developmental Stages
2.5. Expression Pattern Analysis of PprCIPK Genes under Different Treatment Conditions
2.6. Protein Interactions of Peach CBLs and CIPKs
3. Discussion
4. Materials and Methods
4.1. Identification of the Peach CBL and CIPK Family
4.2. Plant Materials and Treatment
4.3. RNA Extraction and RT-qPCR
4.4. Phylogenetic Tree, Collinearity, and Gene Structure Analysis of CBL and CIPK Gene Family Members
4.5. RNA-Seq Data Analysis and Heatmap Construction
4.6. Yeast Two-Hybrid Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tang, R.J.; Wang, C.; Li, K.; Luan, S. The CBL-CIPK Calcium Signaling Network: Unified Paradigm from 20 Years of Discoveries. Trends Plant Sci. 2020, 25, 604–617. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Liu, B.; Sun, X.; Sun, X.; Zheng, C. New functions of CIPK gene family are continue to emerging. Mol. Biol. Rep. 2022, 49, 6647–6658. [Google Scholar] [CrossRef] [PubMed]
- Bailey-Serres, J.; Voesenek, L.A. Life in the balance: A signaling network controlling survival of flooding. Curr. Opin. Plant Biol. 2010, 13, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Verma, P.; Sanyal, S.K.; Pandey, G.K. Ca(2+)-CBL-CIPK: A modulator system for efficient nutrient acquisition. Plant Cell Rep. 2021, 40, 2111–2122. [Google Scholar] [CrossRef]
- Yu, Q.; An, L.; Li, W. The CBL-CIPK network mediates different signaling pathways in plants. Plant Cell Rep. 2014, 33, 203–214. [Google Scholar] [CrossRef]
- Batistic, O.; Waadt, R.; Steinhorst, L.; Held, K.; Kudla, J. CBL-mediated targeting of CIPKs facilitates the decoding of calcium signals emanating from distinct cellular stores. Plant J. 2010, 61, 211–222. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Zhao, R.; Zhou, Y.; Jiao, Y. Evolutionary strategies drive a balance of the interacting gene products for the CBL and CIPK gene families. New Phytol. 2020, 226, 1506–1516. [Google Scholar] [CrossRef]
- Mao, J.; Manik, S.M.; Shi, S.; Chao, J.; Jin, Y.; Wang, Q.; Liu, H. Mechanisms and Physiological Roles of the CBL-CIPK Networking System in Arabidopsis thaliana. Genes 2016, 7, 62. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Lin, J.; Li, H.; Li, X.; Yang, Q.; Cheng, Z.M.; Chang, Y. Characterization of CIPK Family in Asian Pear (Pyrus bretschneideri Rehd) and Co-expression Analysis Related to Salt and Osmotic Stress Responses. Front. Plant Sci. 2016, 7, 1361. [Google Scholar] [CrossRef] [Green Version]
- Weinl, S.; Kudla, J. The CBL-CIPK Ca(2+)-decoding signaling network: Function and perspectives. New Phytol. 2009, 184, 517–528. [Google Scholar] [CrossRef]
- Cui, X.Y.; Du, Y.T.; Fu, J.D.; Yu, T.F.; Wang, C.T.; Chen, M.; Chen, J.; Ma, Y.Z.; Xu, Z.S. Wheat CBL-interacting protein kinase 23 positively regulates drought stress and ABA responses. BMC Plant Biol. 2018, 18, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanyal, S.K.; Kanwar, P.; Yadav, A.K.; Sharma, C.; Kumar, A.; Pandey, G.K. Arabidopsis CBL interacting protein kinase 3 interacts with ABR1, an APETALA2 domain transcription factor, to regulate ABA responses. Plant Sci. 2017, 254, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Q.S.; Guo, Y.; Dietrich, M.A.; Schumaker, K.S.; Zhu, J.K. Regulation of SOS1, a plasma membrane Na+/H+ exchanger in Arabidopsis thaliana, by SOS2 and SOS3. Proc. Natl. Acad. Sci. USA 2002, 99, 8436–8441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Q.S.; Guo, Y.; Quintero, F.J.; Pardo, J.M.; Schumaker, K.S.; Zhu, J.K. Regulation of vacuolar Na+/H+ exchange in Arabidopsis thaliana by the salt-overly-sensitive (SOS) pathway. J. Biol. Chem. 2004, 279, 207–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Li, H.D.; Chen, L.Q.; Wang, Y.; Liu, L.L.; He, L.; Wu, W.H. A protein kinase, interacting with two calcineurin B-like proteins, regulates K+ transporter AKT1 in Arabidopsis. Cell 2006, 125, 1347–1360. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Kim, B.G.; Cheong, Y.H.; Pandey, G.K.; Luan, S. A Ca(2)+ signaling pathway regulates a K(+) channel for low-K response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 12625–12630. [Google Scholar] [CrossRef] [Green Version]
- Tang, R.J.; Zhao, F.G.; Yang, Y.; Wang, C.; Li, K.; Kleist, T.J.; Lemaux, P.G.; Luan, S. A calcium signalling network activates vacuolar K(+) remobilization to enable plant adaptation to low-K environments. Nat. Plants 2020, 6, 384–393. [Google Scholar] [CrossRef]
- Tang, R.J.; Zhao, F.G.; Garcia, V.J.; Kleist, T.J.; Yang, L.; Zhang, H.X.; Luan, S. Tonoplast CBL-CIPK calcium signaling network regulates magnesium homeostasis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2015, 112, 3134–3139. [Google Scholar] [CrossRef] [Green Version]
- Dubeaux, G.; Neveu, J.; Zelazny, E.; Vert, G. Metal Sensing by the IRT1 Transporter-Receptor Orchestrates Its Own Degradation and Plant Metal Nutrition. Mol. Cell 2018, 69, 953–964.e5. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Ren, F.; Zhou, L.; Wang, Q.Q.; Zhong, H.; Li, X.B. The Brassica napus calcineurin B-Like 1/CBL-interacting protein kinase 6 (CBL1/CIPK6) component is involved in the plant response to abiotic stress and ABA signalling. J. Exp. Bot. 2012, 63, 6211–6222. [Google Scholar] [CrossRef]
- Deng, X.; Zhou, S.; Hu, W.; Feng, J.; Zhang, F.; Chen, L.; Huang, C.; Luo, Q.; He, Y.; Yang, G.; et al. Ectopic expression of wheat TaCIPK14, encoding a calcineurin B-like protein-interacting protein kinase, confers salinity and cold tolerance in tobacco. Physiol. Plant 2013, 149, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Li, Y.; Gai, W.X.; Li, C.; Gong, Z.H. The CaCIPK3 gene positively regulates drought tolerance in pepper. Hortic. Res. 2021, 8, 216. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, X.; Duan, W.; Wei, C.; Chen, K.; Grierson, D.; Zhang, B. Genome-Wide Identification and Functional Analysis of Carboxylesterase and Methylesterase Gene Families in Peach (Prunus persica L. Batsch). Front. Plant Sci. 2019, 10, 1511. [Google Scholar] [CrossRef]
- Ksouri, N.; Jimenez, S.; Wells, C.E.; Contreras-Moreira, B.; Gogorcena, Y. Transcriptional Responses in Root and Leaf of Prunus persica under Drought Stress Using RNA Sequencing. Front. Plant Sci. 2016, 7, 1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Cao, H.; Pan, L.; Niu, L.; Wei, B.; Cui, G.; Wang, L.; Yao, J.L.; Zeng, W.; Wang, Z. Two loss-of-function alleles of the glutathione S-transferase (GST) gene cause anthocyanin deficiency in flower and fruit skin of peach (Prunus persica). Plant J. 2021, 107, 1320–1331. [Google Scholar] [CrossRef]
- Yu, D.J.; Jun, S.H.; Park, J.; Kwon, J.H.; Lee, H.J. Transcriptome Analysis of Genes Involved in Cold Hardiness of Peach Tree (Prunus persica) Shoots during Cold Acclimation and Deacclimation. Genes 2020, 11, 611. [Google Scholar] [CrossRef]
- Kolukisaoglu, U.; Weinl, S.; Blazevic, D.; Batistic, O.; Kudla, J. Calcium sensors and their interacting protein kinases: Genomics of the Arabidopsis and rice CBL-CIPK signaling networks. Plant Physiol. 2004, 134, 43–58. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.G.; Waadt, R.; Cheong, Y.H.; Pandey, G.K.; Dominguez-Solis, J.R.; Schultke, S.; Lee, S.C.; Kudla, J.; Luan, S. The calcium sensor CBL10 mediates salt tolerance by regulating ion homeostasis in Arabidopsis. Plant J. 2007, 52, 473–484. [Google Scholar] [CrossRef]
- Xiaolin, Z.; Baoqiang, W.; Xian, W.; Xiaohong, W. Identification of the CIPK-CBL family gene and functional characterization of CqCIPK14 gene under drought stress in quinoa. BMC Genom. 2022, 23, 447. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, B.; Liu, W.Z.; Li, H.; Wang, L.; Wang, B.; Deng, M.; Liang, W.; Deyholos, M.K.; Jiang, Y.Q. Identification and characterization of CBL and CIPK gene families in canola (Brassica napus L.). BMC Plant Biol. 2014, 14, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Gu, Z.; Xin, D.; Hao, L.; Liu, C.; Huang, J.; Ma, B.; Zhang, H. Identification and characterization of putative CIPK genes in maize. J. Genet. Genom. 2011, 38, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Birchler, J.A.; Veitia, R.A. Gene balance hypothesis: Connecting issues of dosage sensitivity across biological disciplines. Proc. Natl. Acad. Sci. USA 2012, 109, 14746–14753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batistic, O.; Kudla, J. Plant calcineurin B-like proteins and their interacting protein kinases. Biochim. Biophys. Acta 2009, 1793, 985–992. [Google Scholar] [CrossRef] [Green Version]
- Edel, K.H.; Kudla, J. Increasing complexity and versatility: How the calcium signaling toolkit was shaped during plant land colonization. Cell Calcium. 2015, 57, 231–246. [Google Scholar] [CrossRef]
- Jiao, Y.; Leebens-Mack, J.; Ayyampalayam, S.; Bowers, J.E.; McKain, M.R.; McNeal, J.; Rolf, M.; Ruzicka, D.R.; Wafula, E.; Wickett, N.J.; et al. A genome triplication associated with early diversification of the core eudicots. Genome Biol. 2012, 13, R3. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Yang, J.; Mei, Q.; Liu, H.; Cheng, Y.; Ma, F.; Mao, K. Genome-Wide Analysis of the Apple CBL Family Reveals That Mdcbl10.1 Functions Positively in Modulating Apple Salt Tolerance. Int. J. Mol. Sci. 2021, 22, 12430. [Google Scholar] [CrossRef]
- Niu, L.; Dong, B.; Song, Z.; Meng, D.; Fu, Y. Genome-Wide Identification and Characterization of CIPK Family and Analysis Responses to Various Stresses in Apple (Malus domestica). Int. J. Mol. Sci. 2018, 19, 2131. [Google Scholar] [CrossRef] [Green Version]
- Cheong, Y.H.; Pandey, G.K.; Grant, J.J.; Batistic, O.; Li, L.; Kim, B.G.; Lee, S.C.; Kudla, J.; Luan, S. Two calcineurin B-like calcium sensors, interacting with protein kinase CIPK23, regulate leaf transpiration and root potassium uptake in Arabidopsis. Plant J. 2007, 52, 223–239. [Google Scholar] [CrossRef]
- Steinhorst, L.; Mahs, A.; Ischebeck, T.; Zhang, C.; Zhang, X.; Arendt, S.; Schultke, S.; Heilmann, I.; Kudla, J. Vacuolar CBL-CIPK12 Ca(2+)-sensor-kinase complexes are required for polarized pollen tube growth. Curr. Biol. 2015, 25, 1475–1482. [Google Scholar] [CrossRef]
- Jiang, H.; Ma, Q.J.; Zhong, M.S.; Gao, H.N.; Li, Y.Y.; Hao, Y.J. The apple palmitoyltransferase MdPAT16 influences sugar content and salt tolerance via an MdCBL1-MdCIPK13-MdSUT2.2 pathway. Plant J. 2021, 106, 689–705. [Google Scholar] [CrossRef] [PubMed]
- Cuellar, T.; Azeem, F.; Andrianteranagna, M.; Pascaud, F.; Verdeil, J.L.; Sentenac, H.; Zimmermann, S.; Gaillard, I. Potassium transport in developing fleshy fruits: The grapevine inward K(+) channel VvK1.2 is activated by CIPK-CBL complexes and induced in ripening berry flesh cells. Plant J. 2013, 73, 1006–1018. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.; Sopory, S.K.; Tuteja, N. Cloning and characterization of CBL-CIPK signalling components from a legume (Pisum sativum). FEBS J. 2006, 273, 907–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.P.; Koster, P.; Drerup, M.M.; Scholz, M.; Li, S.; Edel, K.H.; Hashimoto, K.; Kuchitsu, K.; Hippler, M.; Kudla, J. Fine-tuning of RBOHF activity is achieved by differential phosphorylation and Ca(2+) binding. New Phytol. 2019, 221, 1935–1949. [Google Scholar] [CrossRef]
- Kleist, T.J.; Spencley, A.L.; Luan, S. Comparative phylogenomics of the CBL-CIPK calcium-decoding network in the moss Physcomitrella, Arabidopsis, and other green lineages. Front. Plant Sci. 2014, 5, 187. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.H.; Ban, Y.; Wen, X.P.; Nakajima, I.; Moriguchi, T. Molecular cloning and expression analysis of an arginine decarboxylase gene from peach (Prunus persica). Gene 2009, 429, 10–17. [Google Scholar] [CrossRef]
- Tong, Z.; Gao, Z.; Wang, F.; Zhou, J.; Zhang, Z. Selection of reliable reference genes for gene expression studies in peach using real-time PCR. BMC Mol. Biol. 2009, 10, 71. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 2019, 47, e47. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Chr. | Genomic Location (bp) | CDS Length (bp) | Protein Length (aa) | MW (kDa) | pI | Localization |
|---|---|---|---|---|---|---|---|---|
| PprCBL1 | Prupe.1G412900 | 1 | 35,996,976–35,999,551 | 771 | 256 | 29.73 | 4.65 | Cytoplasmic |
| PprCBL2 | Prupe.2G188700 | 2 | 22,874,062–22,876,782 | 648 | 215 | 24.47 | 4.92 | Cytoplasmic |
| PprCBL3 | Prupe.2G310300 | 2 | 29,368,821–29,373,136 | 639 | 212 | 24.47 | 4.79 | Cytoplasmic |
| PprCBL4 | Prupe.3G051100 | 3 | 3,597,840–3,603,251 | 813 | 270 | 30.96 | 4.77 | Cytoplasmic |
| PprCBL5 | Prupe.5G054400 | 5 | 5,813,999–5,819,707 | 642 | 213 | 24.54 | 4.75 | Cytoplasmic |
| PprCBL6 | Prupe.5G122100 | 5 | 12,135,993–12,138,422 | 642 | 213 | 24.82 | 4.70 | Cytoplasmic |
| PprCBL7 | Prupe.5G122200 | 5 | 12,139,284–12,143,357 | 657 | 218 | 25.20 | 4.93 | Cytoplasmic |
| PprCBL8 | Prupe.6G028500 | 6 | 2,222,085–2,226,043 | 681 | 226 | 26.05 | 4.82 | Cytoplasmic |
| PprCIPK1 | Prupe.1G026400 | 1 | 1,839,105–1,840,662 | 1305 | 434 | 47.74 | 8.85 | Cytoplasmic |
| PprCIPK2 | Prupe.1G091700 | 1 | 6,909,490–6,913,025 | 1410 | 469 | 52.66 | 7.53 | Nuclear |
| PprCIPK3 | Prupe.1G105000 | 1 | 8,458,368–8,464,307 | 1380 | 459 | 51.29 | 6.53 | Cytoplasmic |
| PprCIPK4 | Prupe.1G392900 | 1 | 34,974,805–34,976,767 | 1362 | 453 | 50.88 | 7.99 | Cytoplasmic |
| PprCIPK5 | Prupe.1G539600 | 1 | 44,093,995–44,099,079 | 1347 | 448 | 50.98 | 7.23 | Cytoplasmic |
| PprCIPK6 | Prupe.2G195900 | 2 | 23,340,327–23,344,917 | 1344 | 447 | 51.31 | 8.78 | Cytoplasmic |
| PprCIPK7 | Prupe.2G261600 | 2 | 27,062,570–27,065,556 | 1326 | 441 | 49.87 | 9.15 | Cytoplasmic |
| PprCIPK8 | Prupe.3G097900 | 3 | 7,505,531–7,506,940 | 1410 | 469 | 53.52 | 8.64 | Cytoplasmic |
| PprCIPK9 | Prupe.4G062900 | 4 | 3,012,360–3,014,902 | 1395 | 464 | 52.11 | 8.83 | Cytoplasmic |
| PprCIPK10 | Prupe.4G063100 | 4 | 3,017,616–3,019,702 | 1461 | 486 | 54.49 | 7.58 | Cytoplasmic |
| PprCIPK11 | Prupe.4G085000 | 4 | 4,165,713–4,171,764 | 1377 | 458 | 51.37 | 8.99 | Cytoplasmic |
| PprCIPK12 | Prupe.6G043300 | 6 | 3,165,706–3,167,991 | 1398 | 465 | 51.98 | 8.42 | Cytoplasmic |
| PprCIPK13 | Prupe.6G043700 | 6 | 3,184,043–3,186,693 | 1395 | 464 | 52.91 | 8.52 | Cytoplasmic |
| PprCIPK14 | Prupe.6G156900 | 6 | 13,963,211–13,970,925 | 1320 | 439 | 50.28 | 6.85 | Cytoplasmic |
| PprCIPK15 | Prupe.6G290400 | 6 | 26,800,421–26,803,360 | 1473 | 490 | 55.41 | 8.64 | Cytoplasmic |
| PprCIPK16 | Prupe.6G290700 | 6 | 26,826,847–26,829,608 | 1317 | 438 | 49.61 | 6.50 | Cytoplasmic |
| PprCIPK17 | Prupe.7G244500 | 7 | 20,886,040–20,890,002 | 1341 | 446 | 50.49 | 8.50 | Cytoplasmic |
| PprCIPK18 | Prupe.7G261300 | 7 | 21,709,634–21,711,533 | 1299 | 432 | 48.61 | 8.96 | Cytoplasmic |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, K.; Pan, H.; Sheng, Y.; Wang, Y.; Shi, P.; Xie, Q.; Zhang, J.; Zhou, H. The Peach (Prunus persica) CBL and CIPK Family Genes: Protein Interaction Profiling and Expression Analysis in Response to Various Abiotic Stresses. Plants 2022, 11, 3001. https://doi.org/10.3390/plants11213001
Qiu K, Pan H, Sheng Y, Wang Y, Shi P, Xie Q, Zhang J, Zhou H. The Peach (Prunus persica) CBL and CIPK Family Genes: Protein Interaction Profiling and Expression Analysis in Response to Various Abiotic Stresses. Plants. 2022; 11(21):3001. https://doi.org/10.3390/plants11213001
Chicago/Turabian StyleQiu, Keli, Haifa Pan, Yu Sheng, Yunyun Wang, Pei Shi, Qingmei Xie, Jinyun Zhang, and Hui Zhou. 2022. "The Peach (Prunus persica) CBL and CIPK Family Genes: Protein Interaction Profiling and Expression Analysis in Response to Various Abiotic Stresses" Plants 11, no. 21: 3001. https://doi.org/10.3390/plants11213001
APA StyleQiu, K., Pan, H., Sheng, Y., Wang, Y., Shi, P., Xie, Q., Zhang, J., & Zhou, H. (2022). The Peach (Prunus persica) CBL and CIPK Family Genes: Protein Interaction Profiling and Expression Analysis in Response to Various Abiotic Stresses. Plants, 11(21), 3001. https://doi.org/10.3390/plants11213001