

Unraveling the Physiological Mechanisms Underlying the Intracultivar Variability of Water Use Efficiency in Vitis vinifera “Grenache”

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Meteorological Conditions
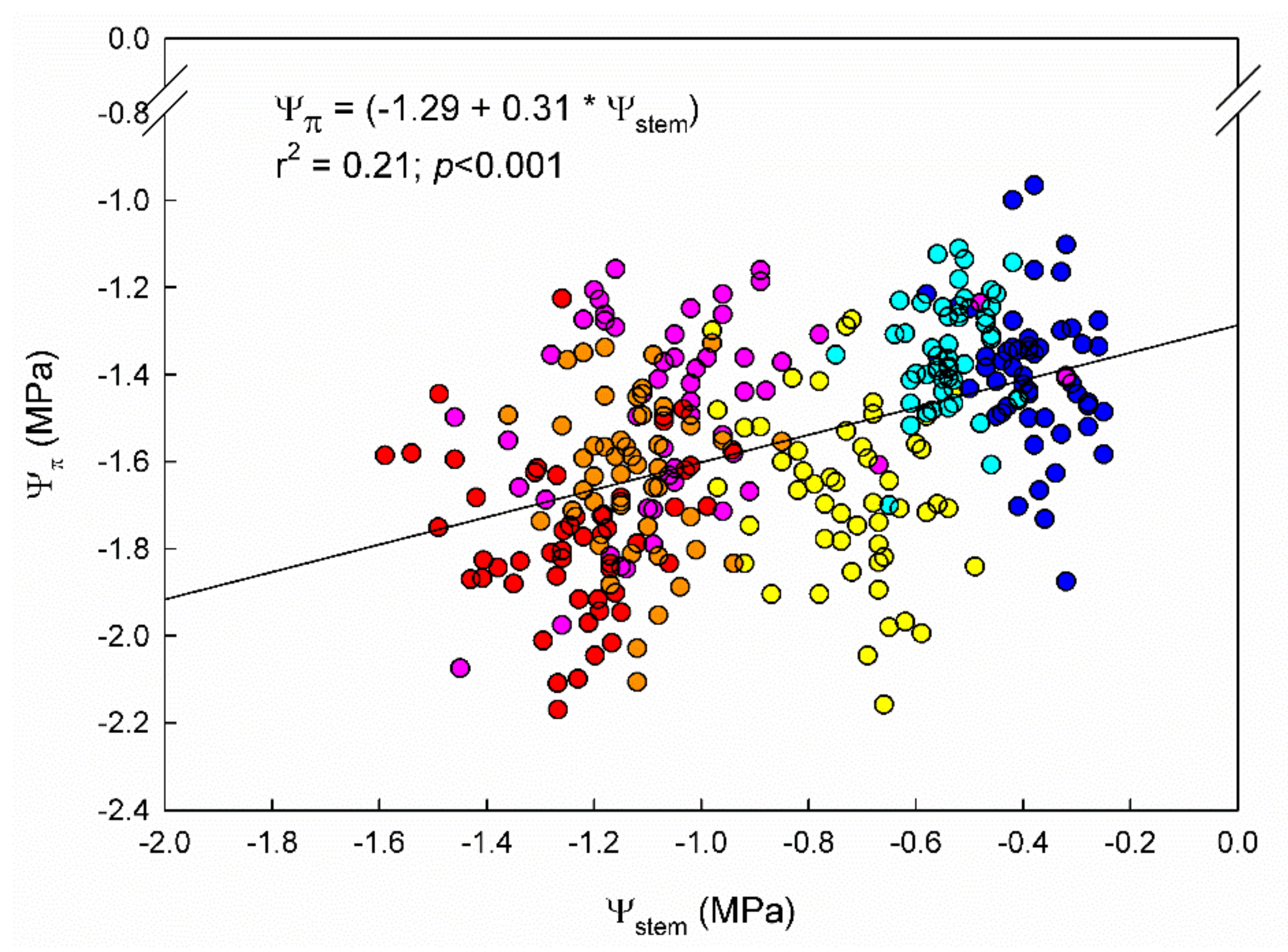
2.2. Plant Water Relations and Net Photosynthesis Rates
2.3. Vine Growth
2.4. Total Biomass
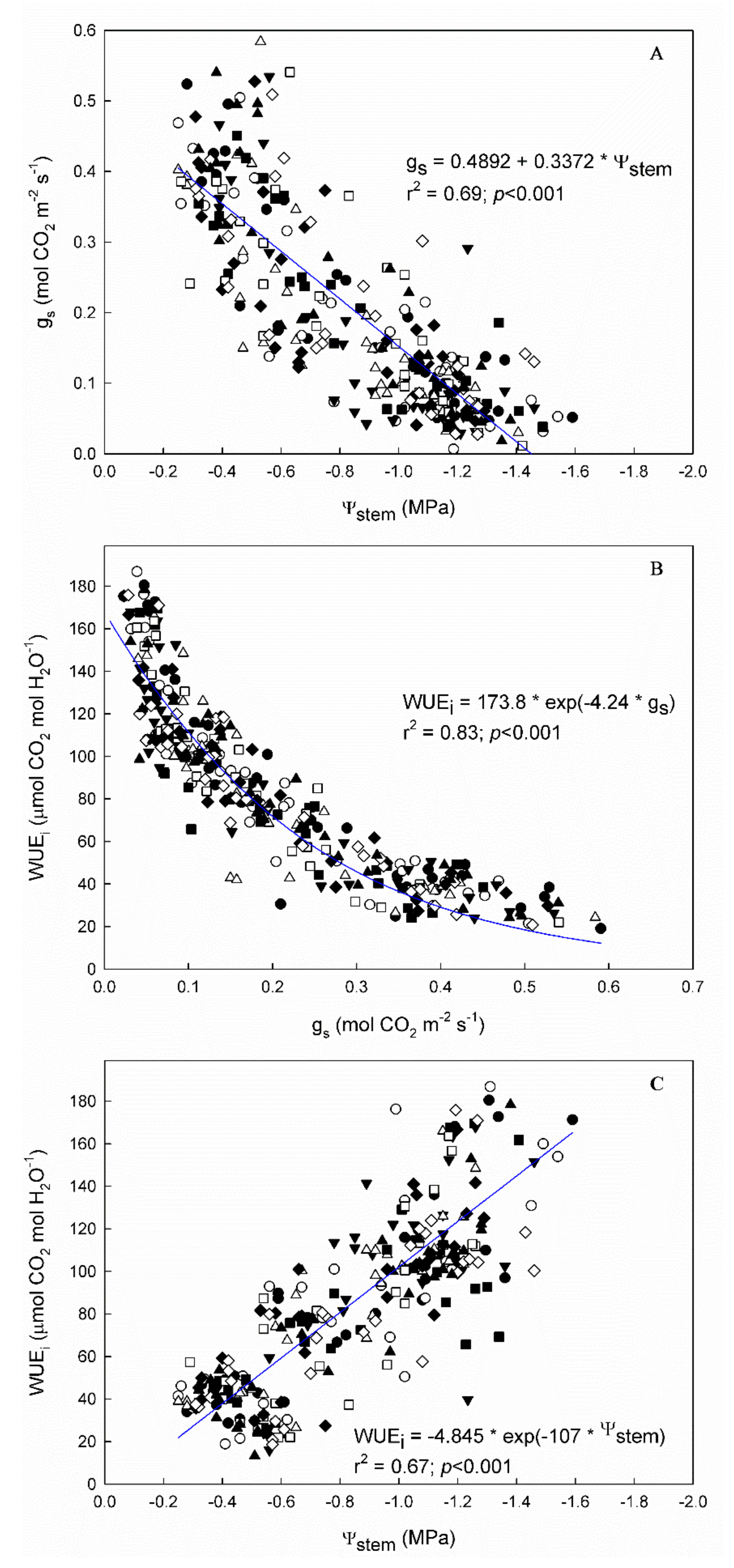
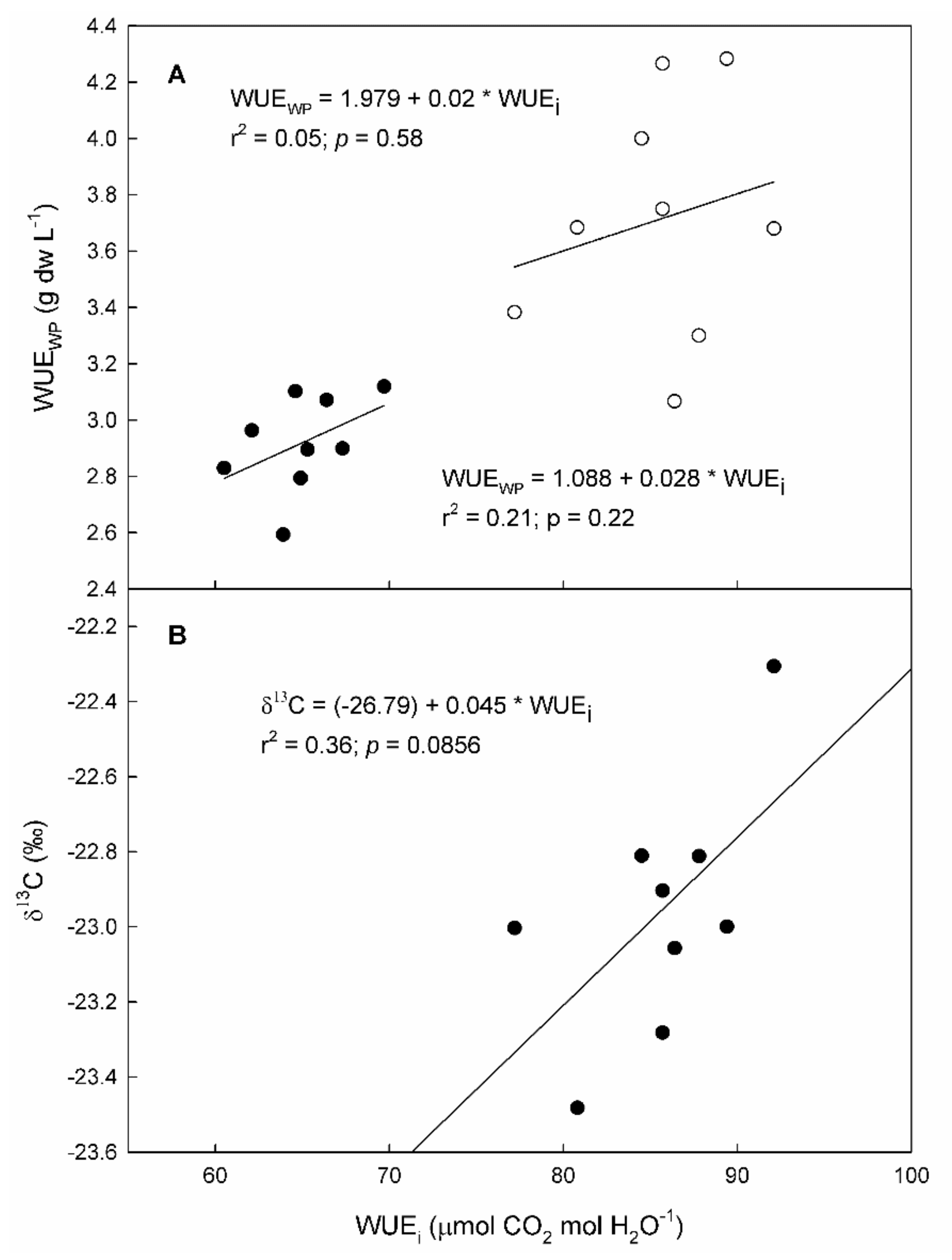
2.5. Water Use Efficiency
2.6. Carbon Isotope Ratio
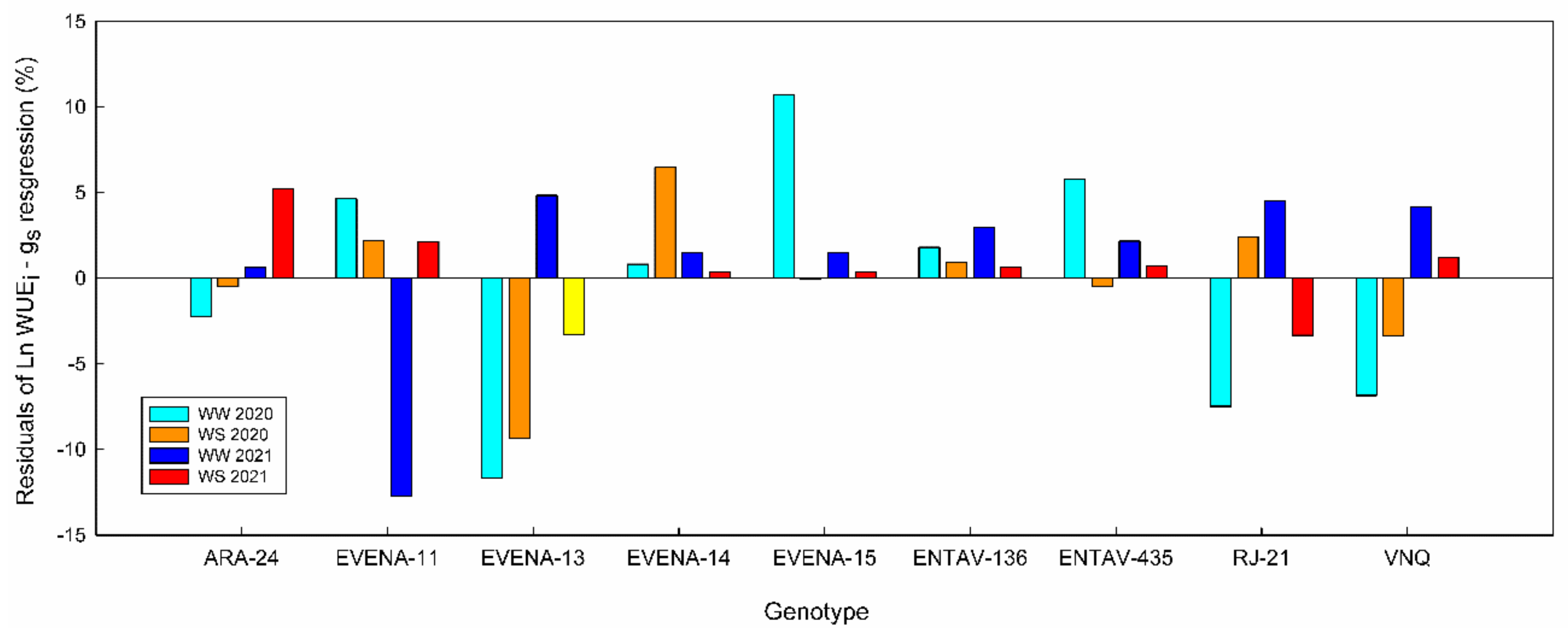
2.7. Ranking Genotypes in WUE
3. Discussion
4. Materials and Methods
4.1. Site Description and Plant Material
4.2. Experimental Design
4.3. Vegetative Growth
4.4. Water Relations
4.5. Leaf Gas Exchange
4.6. Carbon Isotope Ratios
4.7. Ranking Genotypes in WUE
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaves, M.M.; Santos, T.P.; Souza, C.R.; Ortuno, M.F.; Rodrigues, M.L.; Lopes, C.M.; Maroco, J.P.; Pereira, J.S. Deficit irrigation in grapevine improves water-use efficiency while controlling vigour and production quality. Ann. Appl. Biol. 2007, 150, 237–252. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2021: The Physical Science Basis. In Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; 2391p. [Google Scholar] [CrossRef]
- Medrano, H.; Tomás, M.; Martorell, S.; Escalona, J.M.; Pou, A.; Fuentes, S.; Flexas, J.; Bota, J. Improving water use efficiency of vineyards in semi-arid regions. A review. Agron. Sustain. Dev. 2015, 35, 499–517. [Google Scholar] [CrossRef] [Green Version]
- Schultz, H.R. Issues to be considered for strategic adaptation to climate evolution–is atmospheric evaporative demand changing? OENO One 2017, 51, 107–114. [Google Scholar] [CrossRef]
- van Leeuwen, C.; Destrac-Irvine, A.; Dubernet, M.; Duchêne, E.; Gowdy, M.; Marguerit, E.; Pieri, P.; Parker, A.; de Rességuier, L.; Ollat, N. An update on the impact of climate change in viticulture and potential adaptations. Agronomy 2019, 9, 514. [Google Scholar] [CrossRef] [Green Version]
- Buesa, I.; Mirás-Avalos, J.M.; Intrigliolo, D.S. Row orientation effects on potted-vines performance and water-use efficiency. Agr. For. Meteorol. 2020, 294, 108148. [Google Scholar] [CrossRef]
- Buesa, I.; Mirás-Avalos, J.M.; De Paz, J.M.; Visconti, F.; Sanz, F.; Yeves, A.; Guerra, D.; Intrigliolo, D.S. Soil management in semi-arid vineyards: Combined effects of organic mulching and no-tillage under different water regimes. Eur. J. Agron. 2021, 123, 126198. [Google Scholar] [CrossRef]
- Cifre, J.; Bota, J.; Escalona, J.; Medrano, H.; Flexas, J. Physiological tools for irrigation scheduling in grapevine (Vitis vinifera L.): An open gate to improve water-use efficiency? Agric. Ecosyst. Environ. 2005, 106, 159–170. [Google Scholar] [CrossRef]
- Pou, A.; Flexas, J.; Alsina Mdel, M.; Bota, J.; Carambula, C.; de Herralde, F.; Galmes, J.; Lovisolo, C.; Jimenez, M.; Ribas-Carbo, M. Adjustments of water use efficiency by stomatal regulation during drought and recovery in the drought-adapted Vitis hybrid Richter-110 (V. berlandieri × V. rupestris). Physiol. Plant. 2008, 134, 313–323. [Google Scholar] [CrossRef]
- Medrano, H.; Escalona, J.M.; Bota, J.; Gulías, J.; Flexas, J. Regulation of Photosynthesis of C3 Plants in Response to Progressive Drought: Stomatal Conductance as a Reference Parameter. Ann. Bot. 2002, 89, 895–905. [Google Scholar] [CrossRef]
- Gambetta, G.A.; Herrera, J.C.; Dayer, S.; Feng, Q.; Hochberg, U.; Castellarin, S.D. The physiology of drought stress in grapevine: Towards an integrative definition of drought tolerance. J. Exp. Bot. 2020, 71, 4658–4676. [Google Scholar] [CrossRef]
- Pou, A.; Medrano, H.; Tomas, M.; Martorell, S.; Ribas-Carbo, M.; Flexas, J. Anisohydric behaviour in grapevines results in better performance under moderate water stress and recovery than isohydric behaviour. Plant Soil 2012, 359, 335–349. [Google Scholar] [CrossRef]
- Bota, J.; Tomás, M.; Flexas, J.; Medrano, H.; Escalona, J.M. Differences among grapevine cultivars in their stomatal behavior and water use efficiency under progressive water stress. Agric. Water Manag. 2016, 164, 91–99. [Google Scholar] [CrossRef]
- Dayer, S.; Herrera, J.C.; Dai, Z.; Burlett, R.; Lamarque, L.J.; Delzon, S.; Bortolami, G.; Cochard, H.; Gambetta, G.A. The sequence and thresholds of leaf hydraulic traits underlying grapevine varietal differences in drought tolerance. J. Exp. Bot. 2020, 71, 4333–4344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tortosa, I.; Escalona, J.M.; Bota, J.; Tomás, M.; Hernández, E.; Escudero, E.G.; Medrano, H. Exploring the genetic variability in water use efficiency: Evaluation of inter and intra cultivar genetic diversity in grapevines. Plant Sci. 2016, 251, 35–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrizabalaga-Arriazu, M.; Gomès, E.; Morales, F.; Irigoyen, J.J.; Pascual, I.; Hilbert, G. High Temperature and Elevated Carbon Dioxide Modify Berry Composition of Different Clones of Grapevine (Vitis vinifera L.) cv. Tempranillo. Front. Plant Sci. 2020, 11, 603687. [Google Scholar] [CrossRef]
- Tortosa, I.; Douthe, C.; Pou, A.; Balda, P.; Hernandez-Montes, E.; Toro, G.; Escalona, J.M.; Medrano, H. Variability in Water Use Efficiency of Grapevine Tempranillo Clones and Stability over Years at Field Conditions. Agronomy 2019, 9, 701. [Google Scholar] [CrossRef] [Green Version]
- Tortosa, I.; Escalona, J.M.; Toro, G.; Douthe, C.; Medrano, H. Clonal behavior in response to soil water availability in Tempranillo grapevine cv: From plant growth to water use efficiency. Agronomy 2020, 10, 862. [Google Scholar] [CrossRef]
- Tortosa, I.; Escalona, J.M.; Opazo, I.; Douthe, C.; Medrano, H. Genotype Variations in Water Use Efficiency Correspond with Photosynthetic Traits in Tempranillo Grapevine Clones. Agronomy 2022, 12, 1874. [Google Scholar] [CrossRef]
- Buesa, I.; Escalona, J.M.; Tortosa, I.; Marín, D.; Loidi, M.; Santesteban, L.G.; Douthe, C.; Medrano, H. Intracultivar genetic diversity in grapevine: Water use efficiency variability within cv. Grenache. Physiol. Plant. 2021, 173, 2226–2237. [Google Scholar] [CrossRef]
- Pou, A.; Medrano, H.; Flexas, J.; Tyerman, S.D. A putative role for TIP and PIP aquaporins in dynamics of leaf hydraulic and stomatal conductances in grapevine under water stress and re-watering. Plant Cell Environ. 2013, 36, 828–843. [Google Scholar] [CrossRef]
- Ibáñez, J.; de Andrés, M.T.; Molino, A.; Borrego, J. Genetic study of key spanish grapevine varieties using microsatellite analysis. Am. J. Enol. Vitic. 2003, 54, 22–30. [Google Scholar]
- Ibáñez, J.; Muñoz-Organero, G.; Zinelabidine, L.H.; de Andrés, M.T.; Cabello, F.; Martínez-Zapater, J.M. Genetic Origin of the Grapevine Cultivar Tempranillo. Am. J. Enol. Vitic. 2012, 63, 549–553. [Google Scholar] [CrossRef]
- Ibáñez, J.; Carreño, J.; Yuste, J.; Martínez-Zapater, J. Grapevine Breeding and Clonal Selection Programmes in Spain; Woodhead Publishing Series in Food Science, Technology and Nutrition; Elsevier: Amsterdam, The Netherlands, 2015; pp. 183–209. [Google Scholar] [CrossRef]
- Carbonell-Bejerano, P.; Rodríguez-Lorenzo, M.; Royo, C.; Mauri, N.; Hilbert, G.; Renaud, C.; Rambla, J.; Diretto, G.; Granell, A.; Delrot, S.; et al. Grape color variation involves genetic and micro-environmental changes that alter berry phenolic and aromatic composition. Acta Hortic. 2019, 1248, 471–478. [Google Scholar] [CrossRef]
- Schultz, H.R. Differences in hydraulic architecture account for near-isohydric and anisohydric behaviour of two field-grown Vitis vinifera L. cultivars during drought. Plant Cell Environ. 2003, 26, 1393–1405. [Google Scholar] [CrossRef]
- Martorell, S.; Diaz-Espejo, A.; Tomàs, M.; Pou, A.; El Aou-ouad, H.; Escalona, J.M.; Vadell, J.; Ribas-Carbó, M.; Flexas, J.; Medrano, H. Differences in water-use-efficiency between two Vitis vinifera cultivars (Grenache and Tempranillo) explained by the combined response of stomata to hydraulic and chemical signals during water stress. Agric. Water Manag. 2015, 156, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Montes, E.; Tomàs, M.; Escalona, J.; Bota, J.; Medrano, H. Leaf growth rate and nitrogen content determine respiratory costs during leaf expansion in grapevines. Physiol. Plant. 2018, 165, 746–754. [Google Scholar] [CrossRef]
- Buesa, I.; Yeves, A.; Sanz, F.; Chirivella, C.; Intrigliolo, D.S. Effect of delaying winter pruning of Bobal and Tempranillo grapevines on vine performance, grape and wine composition. Aust. J. Grape Wine Res. 2021, 27, 94–105. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Escalona, J.M.; Sampol, B.; Medrano, H. Effects of drought on photosynthesis in grapevines under field conditions: An evaluation of stomatal and mesophyll limitations. Funct. Plant Biol. 2002, 29, 461–471. [Google Scholar] [CrossRef] [Green Version]
- Roig-Oliver, M.; Nadal, M.; Clemente-Moreno, M.J.; Bota, J.; Flexas, J. Cell wall components regulate photosynthesis and leaf water relations of Vitis vinifera cv. Grenache acclimated to contrasting environmental conditions. J. Plant Physiol. 2020, 244, 153084. [Google Scholar] [CrossRef]
- García-Tejera, O.; López-Bernal, Á.; Orgaz, F.; Testi, L.; Villalobos, F. The pitfalls of water potential for irrigation scheduling. Agric. Water Manag. 2020, 243, 106522. [Google Scholar] [CrossRef]
- Lavoie-Lamoureux, A.; Sacco, D.; Risse, P.-A.; Lovisolo, C. Factors influencing stomatal conductance in response to water availability in grapevine: A meta-analysis. Physiol. Plant. 2017, 159, 468–482. [Google Scholar] [CrossRef] [PubMed]
- Soar, C.J.; Speirs, J.; Maffei, S.M.; Penrose, A.B.; McCarthy, M.G.; Loveys, B.R. Grape vine varieties Shiraz and Grenache differ in their stomatal response to VPD: Apparent links with ABA physiology and gene expression in leaf tissue. Aust. J. Grape Wine Res. 2006, 12, 2–12. [Google Scholar] [CrossRef]
- Merli, M.C.; Magnanini, E.; Gatti, M.; Pirez, F.J.; Buesa, I.; Intrigliolo, D.S.; Poni, S. Water stress improves whole-canopy water use efficiency and berry composition of cv. Sangiovese (Vitis vinifera L.) grapevines grafted on the new drought-tolerant rootstock M4. Agric. Water Manag. 2016, 169, 106–114. [Google Scholar] [CrossRef]
- Barrios-Masias, F.; Knipfer, T.; Walker, M.A.; McElrone, A. Differences in hydraulic traits of grapevine rootstocks are not conferred to a common Vitis vinifera scion. Funct. Plant Biol. 2018, 46, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Hinckley, T.M.; Duhme, F.; Hinckley, A.R.; Richter, H. Water relations of drought hardy shrubs: Osmotic potential and stomatal reactivity. Plant Cell Environ. 1980, 3, 131–140. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Gago, J.; Douthe, C.; Florez-Sarasa, I.; Escalona, J.M.; Galmes, J.; Fernie, A.R.; Flexas, J.; Medrano, H. Opportunities for improving leaf water use efficiency under climate change conditions. Plant Sci. 2014, 226, 108–119. [Google Scholar] [CrossRef]
- Tomás, M.; Medrano, H.; Brugnoli, E.; Escalona, J.M.; Martorell, S.; Pou, A.; Ribas-Carbó, M.; Flexas, J. Variability of mesophyll conductance in grapevine cultivars under water stress conditions in relation to leaf anatomy and water use efficiency. Aust. J. Grape Wine Res. 2014, 20, 272–280. [Google Scholar] [CrossRef]
- Medrano, H.; Escalona, J.M.; Cifre, J.; Bota, J.; Flexas, J. A ten-year study on the physiology of two Spanish grapevine cultivars under field conditions: Effects of water availability from leaf photosynthesis to grape yield and quality. Funct. Plant Biol. 2003, 30, 607–619. [Google Scholar] [CrossRef]
- Tomás, M.; Medrano, H.; Pou, A.; Escalona, J.M.; Martorell, S.; Ribas-Carbó, M.; Flexas, J. Water-use efficiency in grapevine cultivars grown under controlled conditions: Effects of water stress at the leaf and whole-plant level. Aust. J. Grape Wine Res. 2012, 18, 164–172. [Google Scholar] [CrossRef]
- Hernández-Montes, E.; Escalona, J.M.; Tomás, M.; Martorell, S.; Bota, J.; Tortosa, I.; Medrano, H. Carbon balance in grapevines (Vitis vinifera L.): Effect of environment, cultivar and phenology on carbon gain, losses and allocation. Aust. J. Grape Wine Res. 2022, 28, 534–544. [Google Scholar] [CrossRef]
- Mairata, A.; Tortosa, I.; Douthe, C.; Escalona, J.M.; Pou, A.; Medrano, H. Comparing Selection Criteria to Select Grapevine Clones by Water Use Efficiency. Agronomy 2022, 12, 1963. [Google Scholar] [CrossRef]
- Bchir, A.; Escalona, J.M.; Gallé, A.; Hernández-Montes, E.; Tortosa, I.; Braham, M.; Medrano, H. Carbon isotope discrimination (δ13C) as an indicator of vine water status and water use efficiency (WUE): Looking for the most representative sample and sampling time. Agric. Water Manag. 2016, 167, 11–20. [Google Scholar] [CrossRef]
- Tomás, M.; Medrano, H.; Escalona, J.M.; Martorell, S.; Pou, A.; Ribas-Carbó, M.; Flexas, J. Variability of water use efficiency in grapevines. Environ. Exp. Bot. 2014, 103, 148–157. [Google Scholar] [CrossRef]
- Galle, A.; Esper, J.; Feller, U.; Ribas-Carbo, M.; Fonti, P. Responses of wood anatomy and carbon isotope composition of Quercus pubescens saplings subjected to two consecutive years of summer drought. Ann. For. Sci. 2010, 67, 809. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Ψstem (MPa) | Ψπ (MPa) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2020 | 2021 | 2020 | 2021 | ||||||||
| WW | WD | WW | WD | WW | WD | WW | WD | ||||
| ARA-24 | −0.52 | −1.13 | abc | −0.38 | −1.05 | ab | −1.33 | abc | −1.65 | −1.39 | −1.65 |
| EVENA-11 | −0.55 | −1.10 | abc | −0.37 | −0.94 | b | −1.47 | a | −1.68 | −1.46 | −1.61 |
| EVENA-13 | −0.58 | −1.23 | a | −0.40 | −1.01 | ab | −1.43 | ab | −1.79 | −1.30 | −1.64 |
| EVENA-14 | −0.55 | −1.11 | abc | −0.39 | −0.99 | b | −1.24 | c | −1.61 | −1.28 | −1.61 |
| EVENA-15 | −0.57 | −1.10 | bc | −0.36 | −0.96 | b | −1.37 | abc | −1.61 | −1.40 | −1.71 |
| ENTAV-136 | −0.51 | −1.03 | c | −0.34 | −1.11 | a | −1.33 | abc | −1.54 | −1.47 | −1.63 |
| ENTAV-435 | −0.53 | −1.19 | ab | −0.43 | −1.00 | ab | −1.30 | bc | −1.60 | −1.49 | −1.63 |
| RJ21 | −0.58 | −1.18 | ab | −0.38 | −1.04 | ab | −1.43 | ab | −1.65 | −1.40 | −1.65 |
| VNQ | −0.49 | −1.12 | abc | −0.40 | −0.97 | b | −1.23 | c | −1.61 | −1.39 | −1.63 |
| Genotype | gs (mol CO2 m−2 s−1) | AN (µmol CO2 m−2 s−1) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2020 | 2021 | 2020 | 2021 | |||||||||||
| WW | WD | WW | WD | WW | WD | WW | WD | |||||||
| ARA-24 | 0.400 | ab | 0.094 | ab | 0.428 | b | 0.131 | ab | 10.9 | 9.5 | ab | 17.9 | d | 12.9 |
| EVENA-11 | 0.291 | a | 0.103 | ab | 0.387 | ab | 0.119 | ab | 9.2 | 10.7 | ab | 15.3 | bcd | 11.8 |
| EVENA-13 | 0.372 | ab | 0.079 | ab | 0.341 | ab | 0.136 | ab | 9.8 | 7.6 | a | 14.5 | ab | 11.3 |
| EVENA-14 | 0.482 | ab | 0.101 | ab | 0.334 | a | 0.131 | ab | 12.1 | 10.8 | ab | 12.3 | a | 11.3 |
| EVENA-15 | 0.439 | ab | 0.123 | b | 0.356 | ab | 0.118 | ab | 12.9 | 12.0 | b | 16.1 | bcd | 11.4 |
| ENTAV-136 | 0.459 | ab | 0.127 | b | 0.389 | ab | 0.103 | a | 12.0 | 12.1 | b | 16.7 | bcd | 10.0 |
| ENTAV-435 | 0.496 | ab | 0.070 | a | 0.377 | ab | 0.098 | a | 12.6 | 7.4 | a | 17.2 | cd | 9.5 |
| RJ21 | 0.546 | b | 0.092 | ab | 0.339 | ab | 0.125 | ab | 12.3 | 9.6 | ab | 15.1 | bc | 11.2 |
| VNQ | 0.520 | ab | 0.080 | ab | 0.382 | ab | 0.151 | b | 11.7 | 8.1 | ab | 16.2 | bcd | 12.8 |
| Genotype | SGR (cm day−1) | LAR (n day−1) | LMA (g m−2) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2020 | 2021 | 2020 | 2021 | 2020 | 2021 | |||||||||||||
| WW | WD | WW | WD | WW | WD | WW | WD | WW | WD | WW | WD | |||||||
| ARA-24 | 1.7 | 1.9 | bc | 3.0 | ab | 1.9 | 0.7 | 0.44 | ab | 0.42 | 0.19 | a | 79.8 | b | 75.4 | 62.4 | b | 75.4 |
| EVENA-11 | 2.4 | 1.7 | bc | 3.3 | b | 2.3 | 0.64 | 0.40 | ab | 0.43 | 0.22 | ab | 80.8 | b | 77.5 | 63.0 | b | 77.2 |
| EVENA-13 | 1.5 | 0.9 | a | 2.7 | a | 2.2 | 0.67 | 0.31 | a | 0.40 | 0.22 | ab | 70.5 | ab | 76.6 | 60.2 | ab | 76.8 |
| EVENA-14 | 2.1 | 1.2 | ab | 3.0 | ab | 2.3 | 0.73 | 0.31 | a | 0.45 | 0.24 | b | 76.6 | ab | 73.9 | 54.4 | a | 70.9 |
| EVENA-15 | 2.1 | 2.0 | c | 2.9 | ab | 2.0 | 0.73 | 0.45 | b | 0.43 | 0.21 | ab | 67.1 | a | 75.5 | 65.3 | b | 78.4 |
| ENTAV-136 | 1.8 | 1.7 | bc | 3.1 | ab | 2.1 | 0.63 | 0.42 | ab | 0.40 | 0.20 | a | 77.9 | ab | 78.0 | 67.2 | b | 75.3 |
| ENTAV-435 | 2.3 | 1.2 | ab | 3.1 | ab | 2.2 | 0.7 | 0.30 | a | 0.40 | 0.21 | ab | 75.3 | ab | 76.4 | 64.0 | b | 76.4 |
| RJ21 | 2.2 | 1.5 | abc | 3.0 | ab | 2.3 | 0.79 | 0.30 | a | 0.44 | 0.21 | ab | 76.9 | ab | 78.4 | 59.6 | ab | 75.6 |
| VNQ | 2.0 | 1.5 | abc | 2.7 | a | 2.2 | 0.74 | 0.38 | ab | 0.40 | 0.22 | ab | 67.3 | a | 81.4 | 59.6 | ab | 74.3 |
| Genotype | Leaf Mass | Shoot Mass | Yield | Total Biomass (g dw) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| (g dw) | (g dw) | (g fw) | |||||||||
| 2020 | 2021 | 2020 | 2021 | 2021 | 2020 | 2021 | |||||
| ARA-24 | 71.4 | 172.4 | b | 44.4 | 131.4 | a | 428.8 | b | 115.8 | 304 | a |
| EVENA-11 | 75.8 | 198.8 | b | 49.0 | 173.7 | b | 302.8 | ab | 124.8 | 372.3 | bc |
| EVENA-13 | 69.9 | 178.2 | ab | 43.2 | 157.3 | ab | 260.5 | ab | 124.2 | 335.7 | ab |
| EVENA-14 | 70.0 | 247.7 | c | 46.0 | 174.1 | b | 173.5 | a | 113.8 | 421.7 | d |
| EVENA-15 | 69.2 | 139.1 | a | 42.5 | 188.8 | b | 181.2 | a | 113.1 | 327.8 | ab |
| ENTAV-136 | 60.8 | 188.4 | ab | 42.9 | 175.4 | b | 402.8 | b | 116.3 | 364.0 | bc |
| ENTAV-435 | 72.6 | 237.5 | c | 50.2 | 185.9 | b | 330.2 | ab | 120.2 | 423.3 | d |
| RJ21 | 72.5 | 233.1 | c | 51.6 | 161.4 | ab | 352.0 | ab | 119.4 | 394.6 | cd |
| VNQ | 74.5 | 195.7 | ab | 44.9 | 170.0 | b | 325.8 | ab | 107.8 | 365.8 | bc |
| Genotype | WUEi (µmol CO2 mol H2O−1) | WUEWP (g L−1) | δ13C (‰) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2020 | 2021 | 2020 | 2021 | 2021 | |||||||||
| WW | WD | WW | WD | ||||||||||
| ARA-24 | 28.4 | ab | 102.3 | ab | 42.1 | 115.6 | ab | 2.9 | ab | 3.1 | a | −23.1 | a |
| EVENA-11 | 35.7 | b | 103.8 | ab | 39.7 | 117.4 | ab | 3.1 | b | 3.8 | bc | −22.9 | ab |
| EVENA-13 | 26.4 | ab | 94.7 | a | 42.7 | 102.3 | a | 2.8 | ab | 3.4 | ab | −23.0 | ab |
| EVENA-14 | 26.2 | ab | 108.4 | b | 45.9 | 110.8 | ab | 2.9 | ab | 4.3 | d | −23.3 | a |
| EVENA-15 | 30.5 | ab | 99.3 | ab | 46.9 | 112.9 | ab | 2.8 | ab | 3.3 | ab | −22.8 | ab |
| ENTAV-136 | 27.7 | ab | 100.0 | ab | 43.8 | 121 | b | 2.6 | a | 3.7 | bc | −22.3 | b |
| ENTAV-435 | 27.7 | ab | 105.2 | ab | 46.5 | 119.1 | ab | 3.1 | b | 4.3 | d | −23.0 | ab |
| RJ21 | 23.7 | a | 105.5 | ab | 45.7 | 111.7 | ab | 3.1 | b | 4.0 | cd | −22.8 | ab |
| VNQ | 23.4 | a | 100.9 | ab | 43.5 | 107.8 | ab | 3.0 | ab | 3.7 | bc | −23.5 | a |
| Genotype | WUEi (µmol CO2 mol H2O−1) | WUEWP (g dw L−1) | δ13C (‰) | Mean Ranking | ||||
|---|---|---|---|---|---|---|---|---|
| 2020 | 2021 | Avg. | 2020 | 2021 | Avg. | 2021 | ||
| ARA-24 | 2 | 2 | 2 | 2 | 3 | 3 | 1 | 2 |
| EVENA-11 | 1 | 2 | 1 | 1 | 2 | 2 | 2 | 1 |
| EVENA-13 | 3 | 3 | 3 | 2 | 3 | 3 | 2 | 3 |
| EVENA-14 | 2 | 2 | 2 | 2 | 1 | 2 | 1 | 1 |
| EVENA-15 | 2 | 2 | 2 | 2 | 3 | 3 | 2 | 3 |
| ENTAV-136 | 2 | 1 | 1 | 3 | 2 | 3 | 1 | 1 |
| ENTAV-435 | 2 | 2 | 2 | 1 | 1 | 1 | 2 | 1 |
| RJ21 | 2 | 2 | 2 | 1 | 1 | 1 | 2 | 1 |
| VNQ | 3 | 2 | 3 | 2 | 2 | 2 | 3 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buesa, I.; Hernández-Montes, E.; Tortosa, I.; Baraldi, G.; Rosselló, M.; Medrano, H.; Escalona, J.M. Unraveling the Physiological Mechanisms Underlying the Intracultivar Variability of Water Use Efficiency in Vitis vinifera “Grenache”. Plants 2022, 11, 3008. https://doi.org/10.3390/plants11213008
Buesa I, Hernández-Montes E, Tortosa I, Baraldi G, Rosselló M, Medrano H, Escalona JM. Unraveling the Physiological Mechanisms Underlying the Intracultivar Variability of Water Use Efficiency in Vitis vinifera “Grenache”. Plants. 2022; 11(21):3008. https://doi.org/10.3390/plants11213008
Chicago/Turabian StyleBuesa, Ignacio, Esther Hernández-Montes, Ignacio Tortosa, Gabriele Baraldi, Miquel Rosselló, Hipólito Medrano, and Jose Mariano Escalona. 2022. "Unraveling the Physiological Mechanisms Underlying the Intracultivar Variability of Water Use Efficiency in Vitis vinifera “Grenache”" Plants 11, no. 21: 3008. https://doi.org/10.3390/plants11213008
APA StyleBuesa, I., Hernández-Montes, E., Tortosa, I., Baraldi, G., Rosselló, M., Medrano, H., & Escalona, J. M. (2022). Unraveling the Physiological Mechanisms Underlying the Intracultivar Variability of Water Use Efficiency in Vitis vinifera “Grenache”. Plants, 11(21), 3008. https://doi.org/10.3390/plants11213008