

Stipagrostis pennata (Trin.) De Winter Artificial Seed Production and Seedlings Multiplication in Temporary Immersion Bioreactors

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
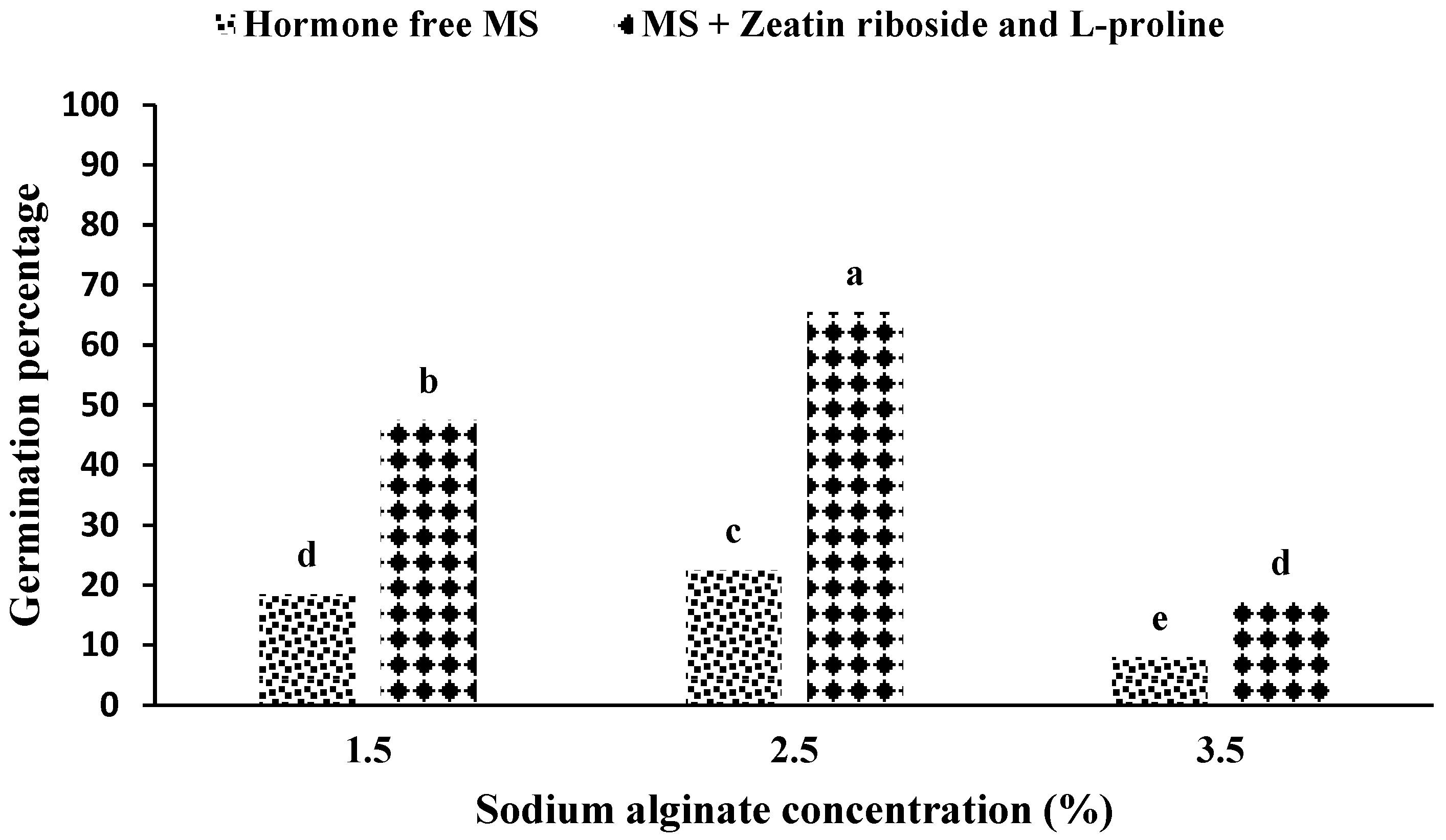
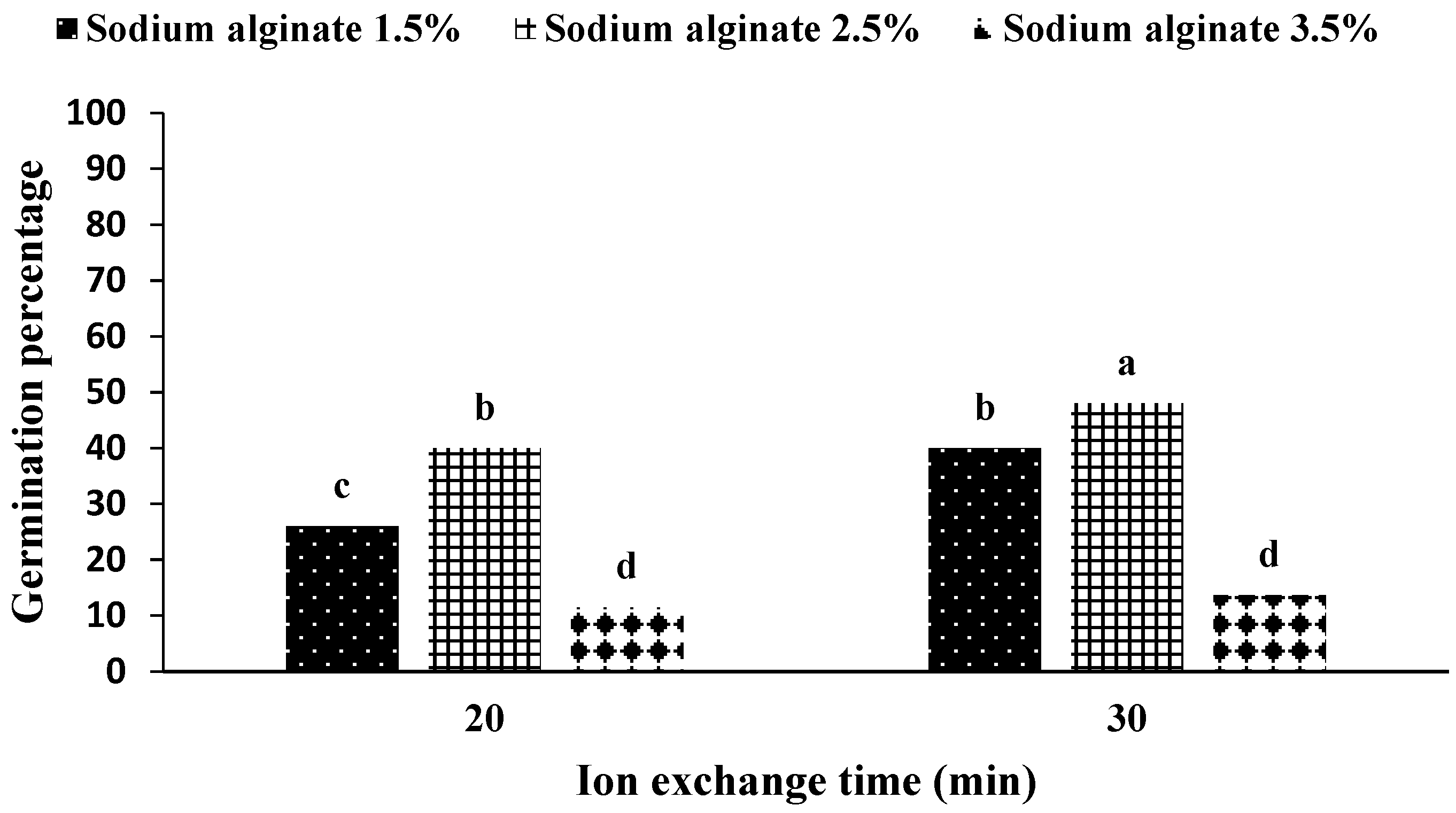
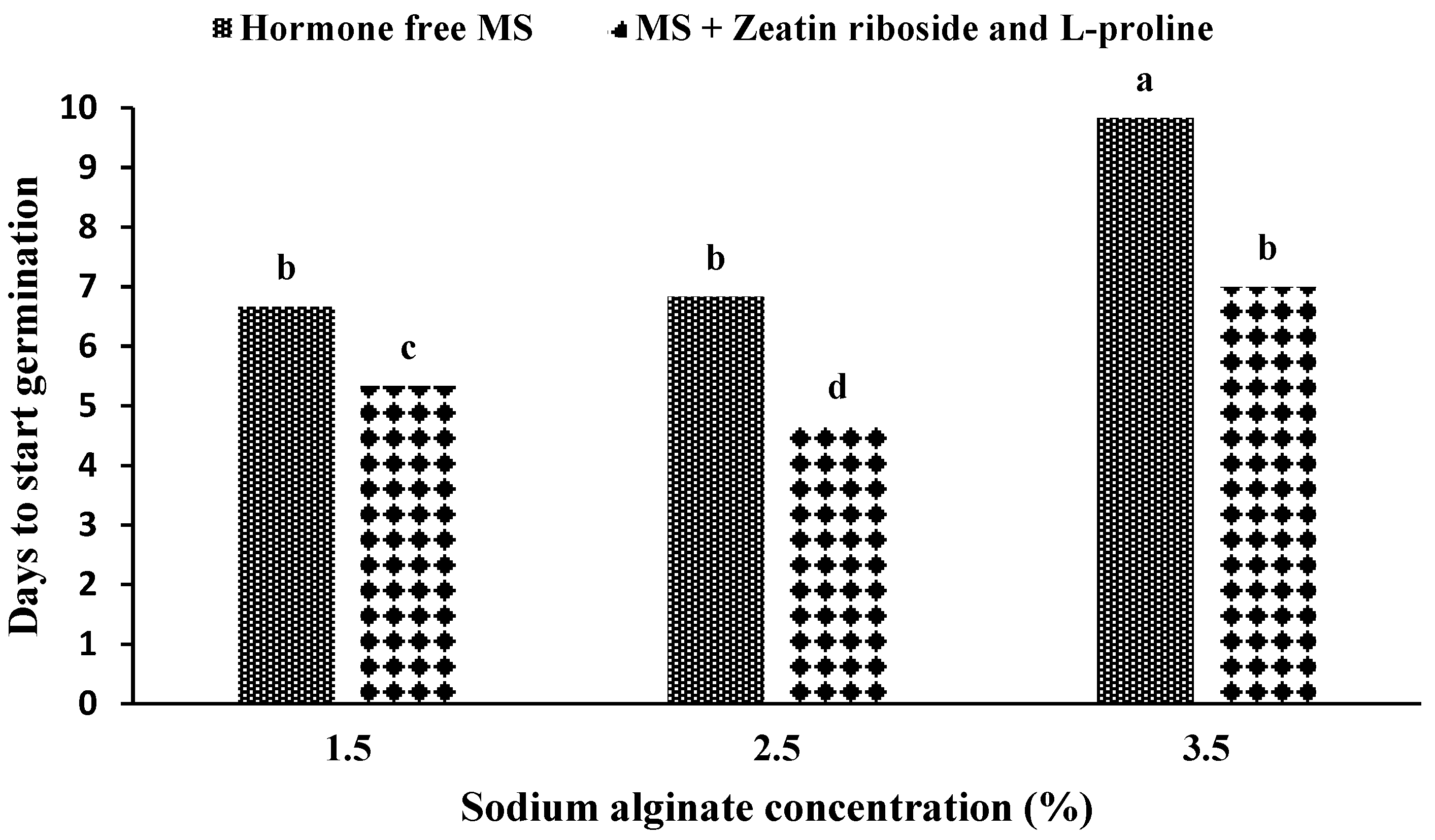
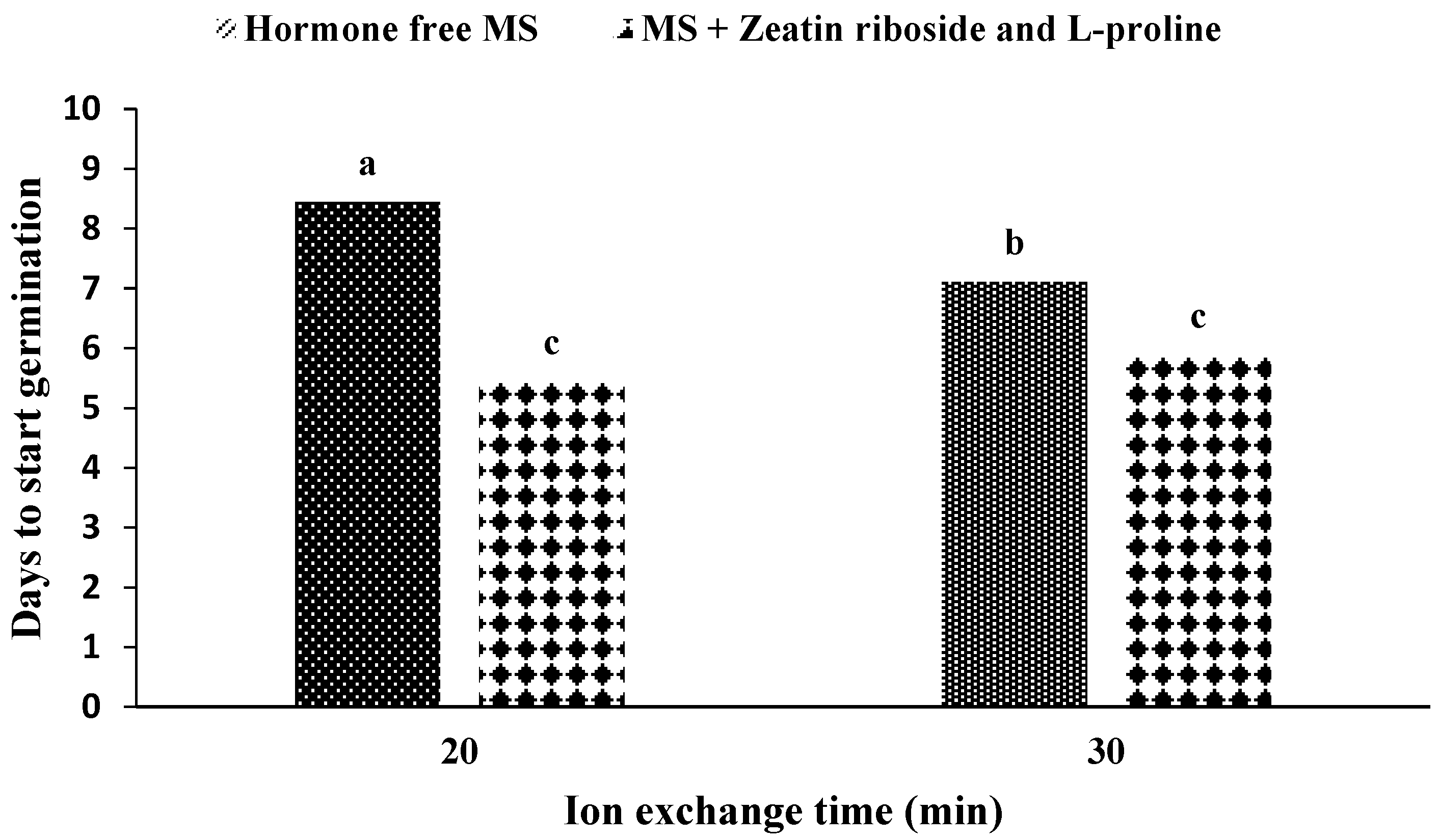
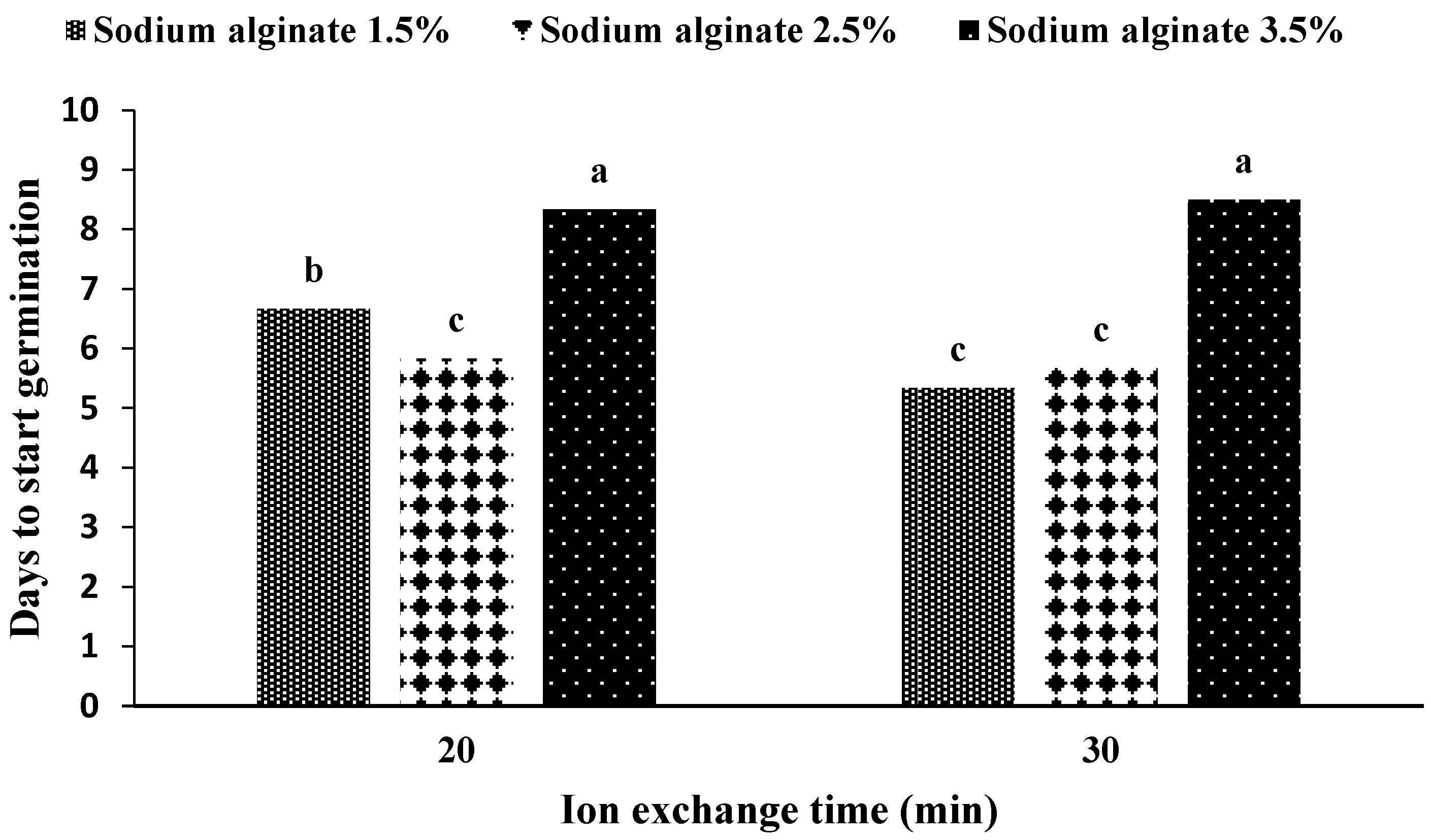
2.1. Production and Germination of Artificial Seeds

2.2. Germination and Seedling Growth on Solid Media and in Temporary Immersion Bioreactors (TIB)
2.2.1. Experiments with Solid Media
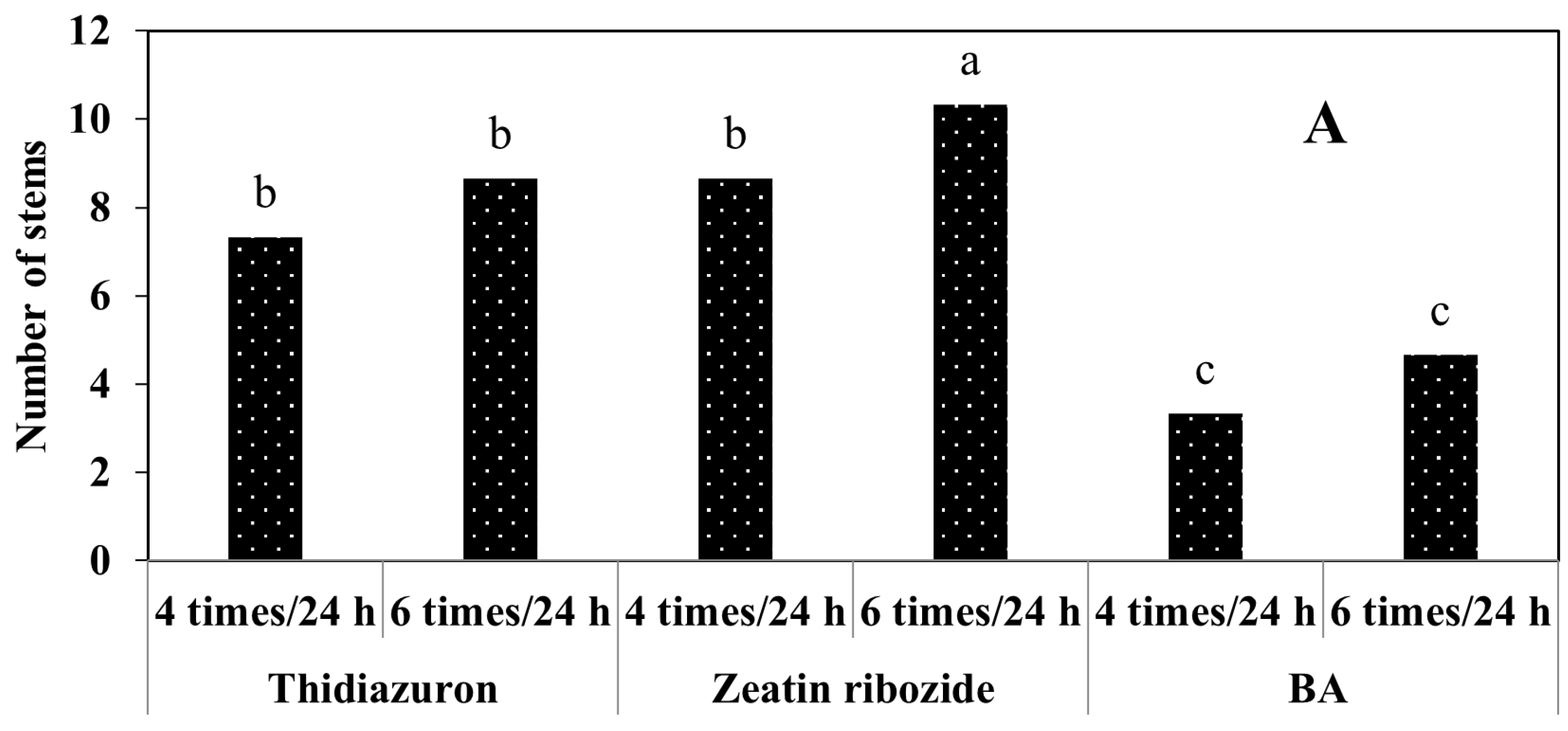
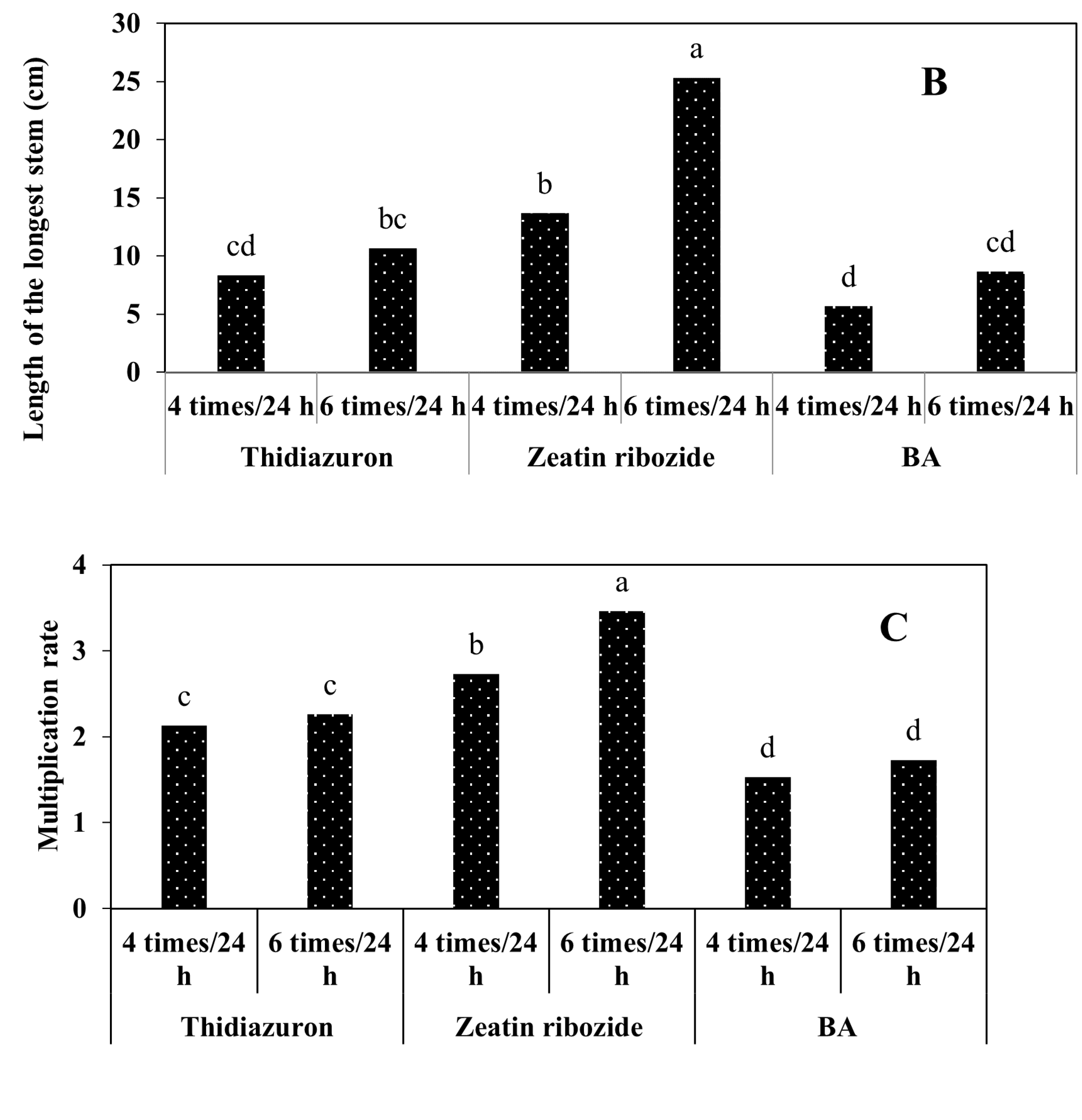
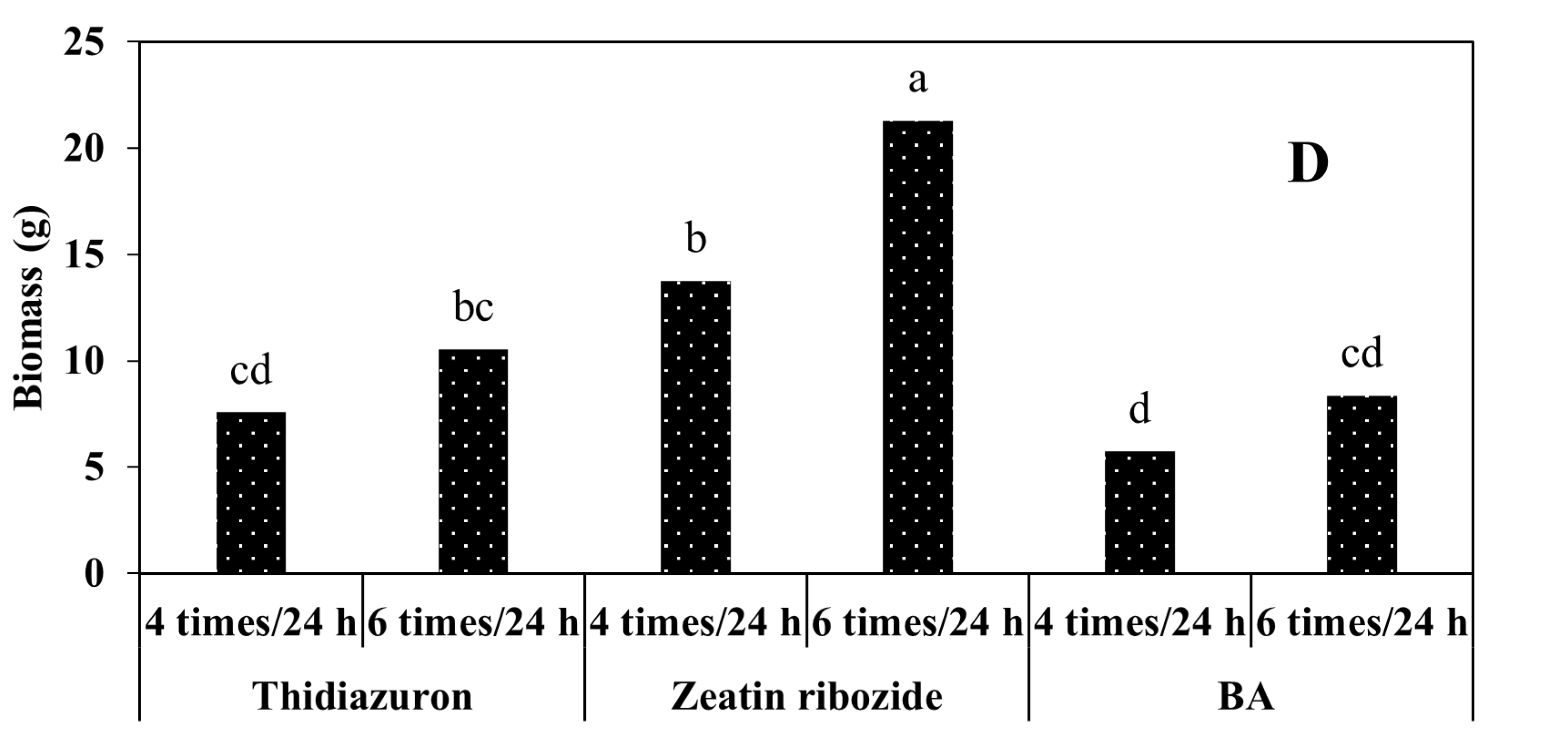
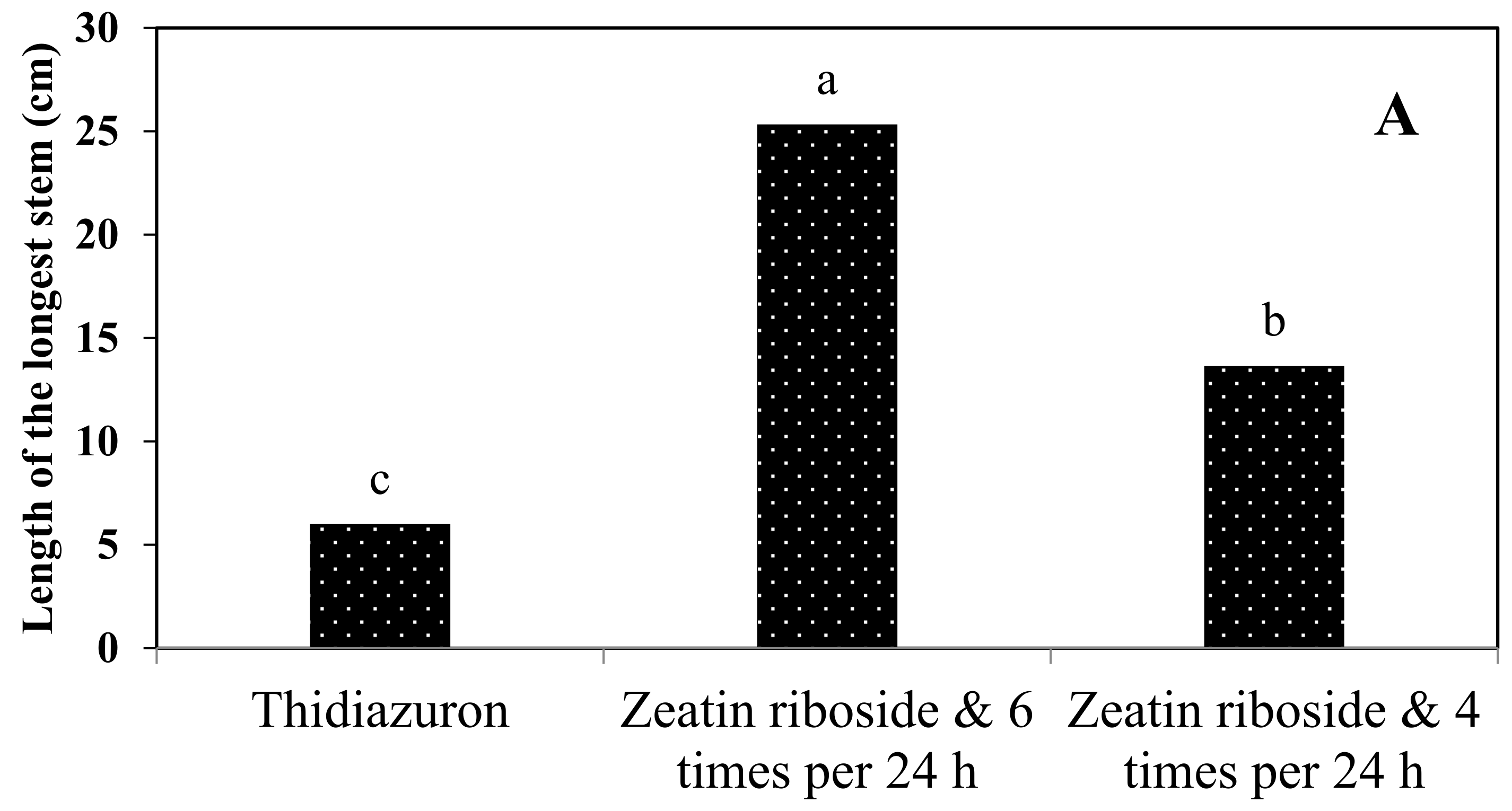
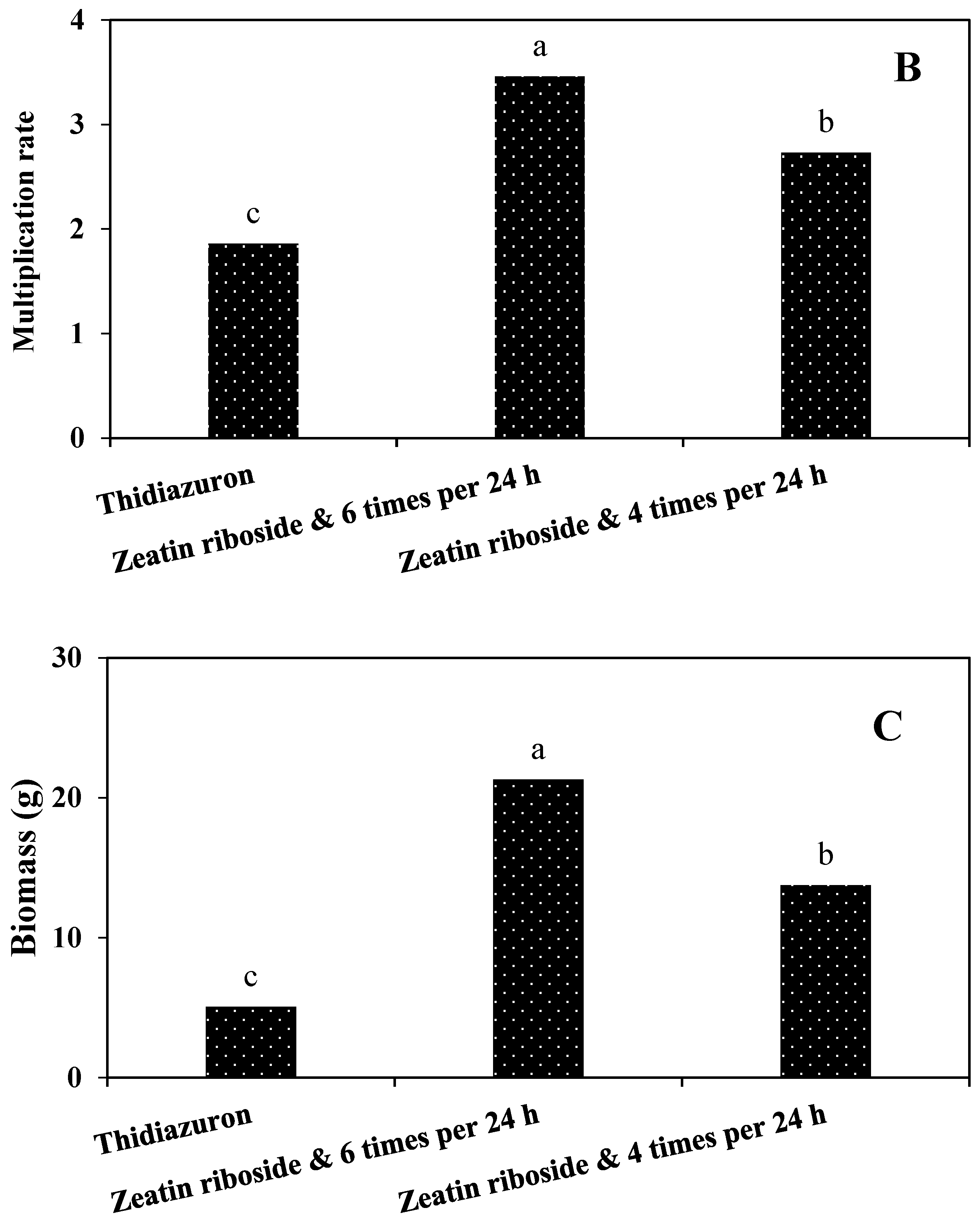
2.2.2. Experiments in TIB
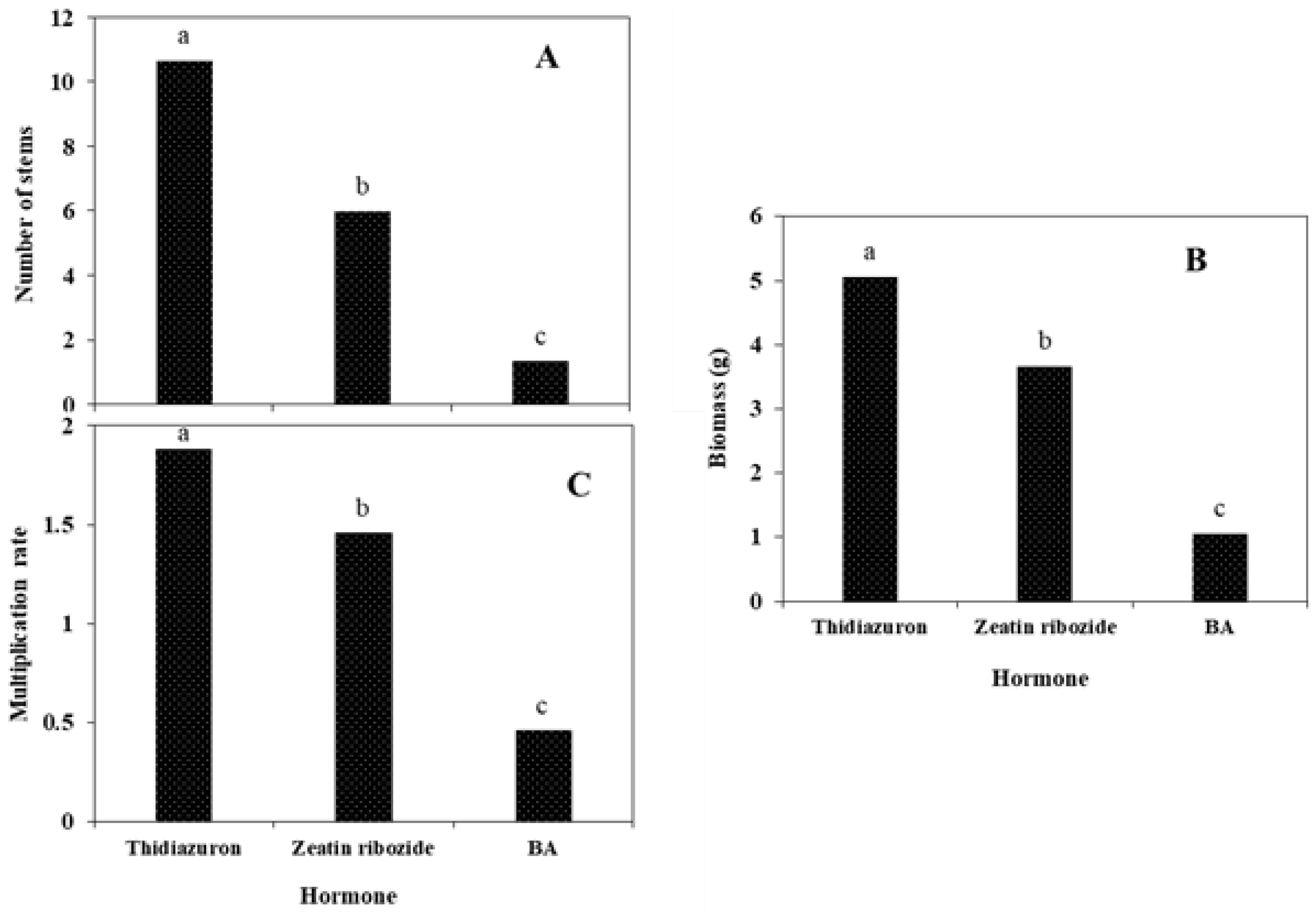
2.2.3. Shoots Micropropagation on Solid Media and in TIB
2.2.4. Soluble Protein Analyses
3. Materials and Methods
3.1. Plant Material
3.2. Encapsulation of Embryogenic Callus
3.3. Effect of Growth Regulators on Artificial Seed Germination and Plant Growth in Solid Cultures
3.4. Temporary Immersion Bioreactors (TIB) System
3.5. Protein Analyses
3.6. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Tian, Y.; Ma, X.; Li, Y.; Cheng, C.; Ge, F.; An, D. Relationship between microbial diversity and nitrogenase activity of Stipagrostis pennata rhizosheath. J. Cell. Biochem. 2019, 120, 13501–13508. [Google Scholar] [PubMed]
- Bahrani, M.J.; Khartegh, M.A. Seed germination of tall three-awn grass (Stipagrostis pennata T. De Winter) is affected by dormancy-breaking treatments, salinity, and harvest time. J. New Seeds 2006, 8, 83–90. [Google Scholar]
- Leonard, J. Altitude de la flore et de la vegetation des deserts de Iran. Fusicule 1982, 1, 2. [Google Scholar]
- Rechinger, K.H. Fifty years of botanical research in the Flora Iranica Area. In Plant Taxonomy, Phytogeography, and Related Subjects; Tan, K., Ed.; The Davis & Hedge Festschrift: Edinburgh, UK, 1989; pp. 301–349. [Google Scholar]
- Gong, B.; Zhu, J.; Zhao, H.; Zhang, Y. Preliminary study on the mechanism for adaptation of Stipagrostis pennata to desert. Int. J. Biol. 2009, 1, 94–100. [Google Scholar]
- Capuano, G.; Piccioni, E.; Standardi, A. Effect of different treatments on the conversion of M. 26 apple rootstock synthetic seeds obtained from encapsulated apical and axillary micro propagated buds. J. Hortic. Sci. Biotechnol. 1998, 73, 289–297. [Google Scholar]
- Ara, H.; Jaiswal, U.; Jaiswal, V.S. Synthetic seed: Prospectus and limitations. Curr. Sci. 2000, 78, 1438–1444. [Google Scholar]
- Danso, K.E.; Ford-Lloyd, B.V. Encapsulation of nodal cuttings and shoot tips for storage and exchange of cassava germplasm. Plant Cell Rep. 2003, 21, 718–725. [Google Scholar]
- Panis, B.J. Cryopreservation of Banana (Musa spp.) Germplasm. Ph.D. Thesis, Catholic University, Leuven, Belgium, 1996. [Google Scholar]
- Rai, M.K.; Asthana, P.; Singh, S.K.; Jaiswal, V.S.; Jaiswal, U. The encapsulation technology in fruit plants. Biotechnol. Adv. 2009, 27, 671–679. [Google Scholar]
- Rai, M.K.; Jaiswal, V.S.; Jaiswal, U. Encapsulation of shoot tips of guava (Psidium guajava L.) for short-term storage and germplasm exchange. Sci. Hortic. 2008, 118, 33–38. [Google Scholar]
- Molle, F. Carrot somatic embryogenesis and its application to synthetic seed. In Synseeds: Applications of Synthetic Seeds to Crop Improvement; Redenbaugh, K., Ed.; CRC Press: Boca Raton, FL, USA; London, UK, 1993; pp. 257–387. [Google Scholar]
- Salimi, Z. Investigation of the Possibility of Somatic Embryogenesis and Synthetic Seed Production of Hyparrhenia hirta. Master’s Thesis, University of Tehran, Tehran, Iran, 2011. (In Persian). [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar]
- Kazemi, Z. Somatic Embryogenesis and Synthetic Seed Production of Hyparrhenia hirta. Master’s Thesis, University of Tehran, Tehran, Iran, 2013. (In Persian). [Google Scholar]
- Hung, C.; Trueman, S. Alginate encapsulation of shoot tips and nodal segments for short-term storage and distribution of the eucalypt Corymbia torelliana × C. citriodora. Acta Physiol. Plant. 2011, 34, 117–128. [Google Scholar]
- Moradi, S.; Azimi, M.; Pourdad, S.; Habibi, F. Encapsulation of two Iranian sunflower hybrid shoot tips and compared conversion and growth of synthetic seeds produced on different substrates. Appl. Crop Breed. 2019, 4, 49–59. (In Persian) [Google Scholar]
- Moradi, S.; Pourdad, S.; Azimi, M.R.; Habibi, F. Effect of alginate matrix composition on growth of synthetic seeds derived from the encapsulation of somatic embryos in two Iranian sunflowers (Helianthus annuus L.) hybrids. Seed Plant J. 2015, 31, 57–74. (In Persian) [Google Scholar]
- Tabassum, B.; Nasir, I.A.; Farooq, A.M.; Rehman, Z.; Latif, Z.; Husnain, T. Viability assessment of in vitro produced synthetic seeds of cucumber. Afr. J. Biotechnol. 2010, 9, 7026–7032. [Google Scholar]
- Klimaszewska, K.; Hargreaves, C.; Lelu-Walter, M.A.; Trontin, J.F. Advances in conifer somatic embryogenesis since the year 2000. In In Vitro Embryogenesis in Higher Plants; Methods in Molecular Biology; Humana Press Inc.: New York, NY, USA, 2016; pp. 131–166. [Google Scholar]
- Vidal, N.; Sánchez, C. Use of bioreactor systems in the propagation of forest trees. Eng. Life Sci. 2019, 19, 896–915. [Google Scholar]
- Redenbaugh, K. (Ed.) Synseeds: Applications of Synthetic Seeds to Crop Improvement; CRC Press: Boca Raton, FL, USA; London, UK; Tokyo, Japan, 1993; p. 471. [Google Scholar]
- Redenbaugh, K.; Slade, D.; Viss, P.; Fujii, J.A. Encapsulation of somatic embryos in synthetic seed coats. HortScience 1987, 22, 803–809. [Google Scholar]
- Haque, S.M.; Ghosh, B. High-frequency somatic embryogenesis and artificial seeds for mass production of true-to-type plants in Ledebouria revoluta: An important cardioprotective plant. Plant Cell Tissue Organ Cult. 2016, 127, 71–83. [Google Scholar]
- Gantait, S.; Kundu, S.; Ali, N.; Sahu, N.C. Synthetic seed production of medicinal plants: A review on the influence of explants, encapsulation agent and matrix. Acta Physiol. Plant. 2015, 37, 98. [Google Scholar]
- Inpuay, K.; Te-chato, S. Primary and secondary somatic embryos as a tool for the tools propagation and artificial seed production of oil palm. J. Agric. Technol. 2012, 8, 597–609. [Google Scholar]
- Ghosh, B.; Sen, S. Plant regeneration from alginate encapsulated somatic embryos of Asparagus cooperi Baker. Plant Cell Rep. 1994, 13, 381–385. [Google Scholar]
- Mujib, A.; Ali, M.; Isah, T. Somatic embryo mediated mass production of Catharanthus roseus in culture vessel (bioreactor). A comparative study. Saudi J. Biol. Sci. 2014, 21, 442–449. [Google Scholar] [PubMed] [Green Version]
- Rizkalla, A.A.; Badr-Elden, A.M.; Ottai, M.E.S.; Nasr, M.I.; Esmail, M.N. Development of artificial seed technology and preservation in sugar beet. Sugar Tech 2012, 14, 312–320. [Google Scholar]
- Huda, A.K.M.N.; Rahman, M.; Bari, M.A. Effect of carbon source in alginate bead on synthetic seed germination in eggplant (Solanum melongena L.). J. Plant Sci. 2007, 2, 538–544. [Google Scholar]
- Gantait, S.; Vijayan, J.; Majee, A. Artificial seed production of Tylophor indicaca indicateerim storing and swapping of germplasm. Hortic. Plant J. 2017, 3, 41–46. [Google Scholar]
- Kadota, M.; Niimi, Y. Effects of cytokines types and their concentrations on shoot proliferation and hyperhydricity in in vitro pear shoot cultivar. Plant Cell Tissue Organ Cult. 2003, 72, 261–265. [Google Scholar]
- Ahmadian, M.; Babaei, A.; Shokri, S.; Hessami, S. Micropropagation of carnation (Dianthus caryophyllus L.) in liquid medium by temporary immersion bioreactor in comparison with solid culture. J. Genet. Eng. Biotechnol. 2017, 15, 309–315. [Google Scholar] [PubMed]
- Ghasemiomran, V.; Movahedi, M.; Alizadeh, F.; Khalvati, L.; Pourahmadi, D. Effects of explant type and plant growth regulator combinations on in vitro direct and indirect regeneration of Stevia rebaudiana. J. Plant Prod. Technol. 2018, 18, 137–147. (In Persian) [Google Scholar]
- Kruger, N.J. The Bradford Method for protein quantitation. In The Protein Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 17–24. [Google Scholar]
- Ehsanpour, A.; Nejati, Z. Effect of nanosilver on chlorophyll, gibberellic acid content and electrophoresis pattern of proteins of potato (Solanum tubersum L.) under in vitro culture. Appl. Biol. 2013, 25, 13–26. [Google Scholar]
- Fathi Rezaei, P.; Nezhad, M.; Aghaee, A. Evaluation of the effect of iron sulfate on growth and some biochemical parameters of garlic plantlets under in vitro conditions. J. Plant Res. (Iran. J. Biol.) 2020, 32, 898–909. [Google Scholar]
- Entisari, M.; Davoodi, D.; Haghnazari, A.; Bagheri, S.; Majidi, E.; Habashi, A.A. Effect of alternative bioreactor on propagation and microtuberization parameters of Potato (Solanum tuberosum L.). J. Crop Breed. 2012, 4, 53–67. (In Persian) [Google Scholar]
- Asadi Aghbolaghi, M.; Sharifzadeh, F.; Omidi, M. Effect of explants and concentrations of plant growth regulators on callus induction in Stipagrostis pennata. Iran. J. Field Crop Sci. 2020, 51, 111–120. (In Persian) [Google Scholar]
- Asadi Aghbolaghi, M.; Dedicova, B.; Ranade, S.S.; Le, K.C.; Sharifzadeh, F.; Omidi, M.; Egertsdotter, U. Protocol development for somatic embryogenesis, SSR markers, and genetic modification of Stipagrostis pennata (Trin.) De Winter. Plant Methods 2021, 17, 1–14. [Google Scholar]
- Le, K.-L.; Dedicova, B.; Johansson, S.; Lelu-Walter, M.-A.; Egertsdotter, U. Temporary immersion bioreactor system for propagation by somatic embryogenesis of hybrid Larch (Larix × eurolepis Henry). Biotechnol. Rep. 2021, 32, 1–8. [Google Scholar]
- Bradford, M. A rapid and sensitive method for the quantification of microgram quantities in utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 254–284. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SOV | df | MS | |
|---|---|---|---|
| Germination (%) | Days to Start Germination | ||
| Germination Medium (M) | 1 | 6588.02 ** | 40.11 ** |
| Sodium alginate (S) | 2 | 3049.69 ** | 26.02 ** |
| Ion exchange time (T) | 1 | 584.02 ** | 1.77 * |
| M × S | 2 | 867.02 ** | 1.69 ** |
| M × T | 1 | 38.02 ns | 7.11 ** |
| S × T | 2 | 105.02 ** | 1.86 ** |
| M × S × T | 2 | 3.02 ns | 0.52 ns |
| Error | 24 | 9.94 | 0.25 |
| CV (%) | - | 10.5 | 7.4 |
| SOV | df | MS | |||||
|---|---|---|---|---|---|---|---|
| Number of Stems | Length of the Longest Stem (cm) | Rooted Samples (%) | Length of the Longest Root (cm) | Biomass | Multiplication Rate | ||
| Hormone | 2 | 3.443 ** | 0.115 ns | 0.012 ns | 0.02 ns | 1.21 ** | 0.388 ** |
| Error | 6 | 0.042 | 0.146 | 0.025 | 0.027 | 0.016 | 0.003 |
| CV (%) | 9.05 | 14.47 | 14.73 | 15.59 | 7.49 | 5.41 | |
| SOV | df | MS | |||||
|---|---|---|---|---|---|---|---|
| Number of Stems | Length of the Longest Stem (cm) | Rooted Samples (%) | Length of the Longest Root (cm) | Biomass | Multiplication Rate | ||
| Hormone (H) | 2 | 1.938 ** | 4.692 ** | 0.023 ns | 0.155 ns | 3.773 ** | 0.350 ** |
| Feeding cycle (F) | 1 | 0.361 ** | 2.584 ** | 0.035 ns | 0.133 ns | 1.773 ** | 0.056 ** |
| (H) × (F) | 2 | 0.004 ns | 0.385 ** | 0.012 ns | 0.033 ns | 0.081 ns | 0.011 ns |
| Error | 12 | 0.023 | 0.055 | 0.01 | 0.052 | 0.056 | 0.003 |
| CV (%) | 5.81 | 6.99 | 9.64 | 19.91 | 7.29 | 4.03 | |
| Treatments | Number of Stems | Length of the Longest Stem (cm) | The Number of Rooted Samples (%) | Length of the Longest Root (cm) | Biomass (g) | Multiplication Rate |
|---|---|---|---|---|---|---|
| Hormone | ||||||
| Thidiazuron | 8 b | 9.5 b | 3.33 a | 0.31 ab | 9.05 b | 2.2 b |
| Zeatin riboside | 9.5 a | 19.5 a | 4 a | 0.85 a | 17.53 a | 3.1 a |
| Benzylaminopurine | 4 c | 7.16 b | 0 a | 0 b | 7.05 b | 1.63 c |
| Feeding Cycle | ||||||
| 4× in 24 h | 6.44 b | 9.22 b | 0.77 a | 0.16 a | 9.02 b | 2.13 b |
| 6× in 24 h | 7.88 a | 14.18 a | 4.11 a | 0.61 a | 13.4 a | 2.48 a |
| SOV | df | MS | |||||
|---|---|---|---|---|---|---|---|
| Number of Stems | Length of the Longest Stem (cm) | Rooted Samples (%) | Length of the Longest Root (cm) | Biomass | Multiplication Rate | ||
| Hormone (H) | 2 | 1.938 ** | 4.692 ** | 0.023 ns | 0.155 ns | 3.773 ** | 0.350 ** |
| Feeding Cycle (F) | 1 | 0.361 ** | 2.584 ** | 0.035 ns | 0.133 ns | 1.773 ** | 0.056 ** |
| (H) × (F) | 2 | 0.004 ns | 0.385 ** | 0.012 ns | 0.033 ns | 0.081 ns | 0.011 ns |
| Error | 12 | 0.023 | 0.055 | 0.01 | 0.052 | 0.056 | 0.003 |
| CV (%) | 5.81 | 6.99 | 9.64 | 19.91 | 7.29 | 4.03 | |
| SOV | df | MS | |||||
|---|---|---|---|---|---|---|---|
| Number of Stems | Length of the Longest Stem (cm) | Rooted Samples (%) | Length of the Longest Root (cm) | Biomass | Multiplication Rate | ||
| Treatment | 2 | 0.091 ns | 5.03 ** | 0.005 ns | 0.001 ns | 4.23 ** | 0.186 ** |
| Error | 6 | 0.035 | 0.117 | 0.027 | 0.003 | 0.066 | 0.003 |
| CV (%) | 6 | 9.22 | 14.83 | 5.29 | 7.33 | 3.76 | |
| SOV | df | MS |
|---|---|---|
| Source of seedling | 4 | 0.057 ns |
| Error | 10 | 0.026 |
| CV (%) | - | 9.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aghbolaghi, M.A.; Dedicova, B.; Sharifzadeh, F.; Omidi, M.; Egertsdotter, U. Stipagrostis pennata (Trin.) De Winter Artificial Seed Production and Seedlings Multiplication in Temporary Immersion Bioreactors. Plants 2022, 11, 3122. https://doi.org/10.3390/plants11223122
Aghbolaghi MA, Dedicova B, Sharifzadeh F, Omidi M, Egertsdotter U. Stipagrostis pennata (Trin.) De Winter Artificial Seed Production and Seedlings Multiplication in Temporary Immersion Bioreactors. Plants. 2022; 11(22):3122. https://doi.org/10.3390/plants11223122
Chicago/Turabian StyleAghbolaghi, Masoumeh Asadi, Beata Dedicova, Farzad Sharifzadeh, Mansoor Omidi, and Ulrika Egertsdotter. 2022. "Stipagrostis pennata (Trin.) De Winter Artificial Seed Production and Seedlings Multiplication in Temporary Immersion Bioreactors" Plants 11, no. 22: 3122. https://doi.org/10.3390/plants11223122
APA StyleAghbolaghi, M. A., Dedicova, B., Sharifzadeh, F., Omidi, M., & Egertsdotter, U. (2022). Stipagrostis pennata (Trin.) De Winter Artificial Seed Production and Seedlings Multiplication in Temporary Immersion Bioreactors. Plants, 11(22), 3122. https://doi.org/10.3390/plants11223122