
Bioformulation Containing Cohorts of Ensifer adhaerens MSN12 and Bacillus cereus MEN8 for the Nutrient Enhancement of Cicer arietinum L.

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Characterization of Root-Nodulating and Non-Nodulating Endophytic Bacteria
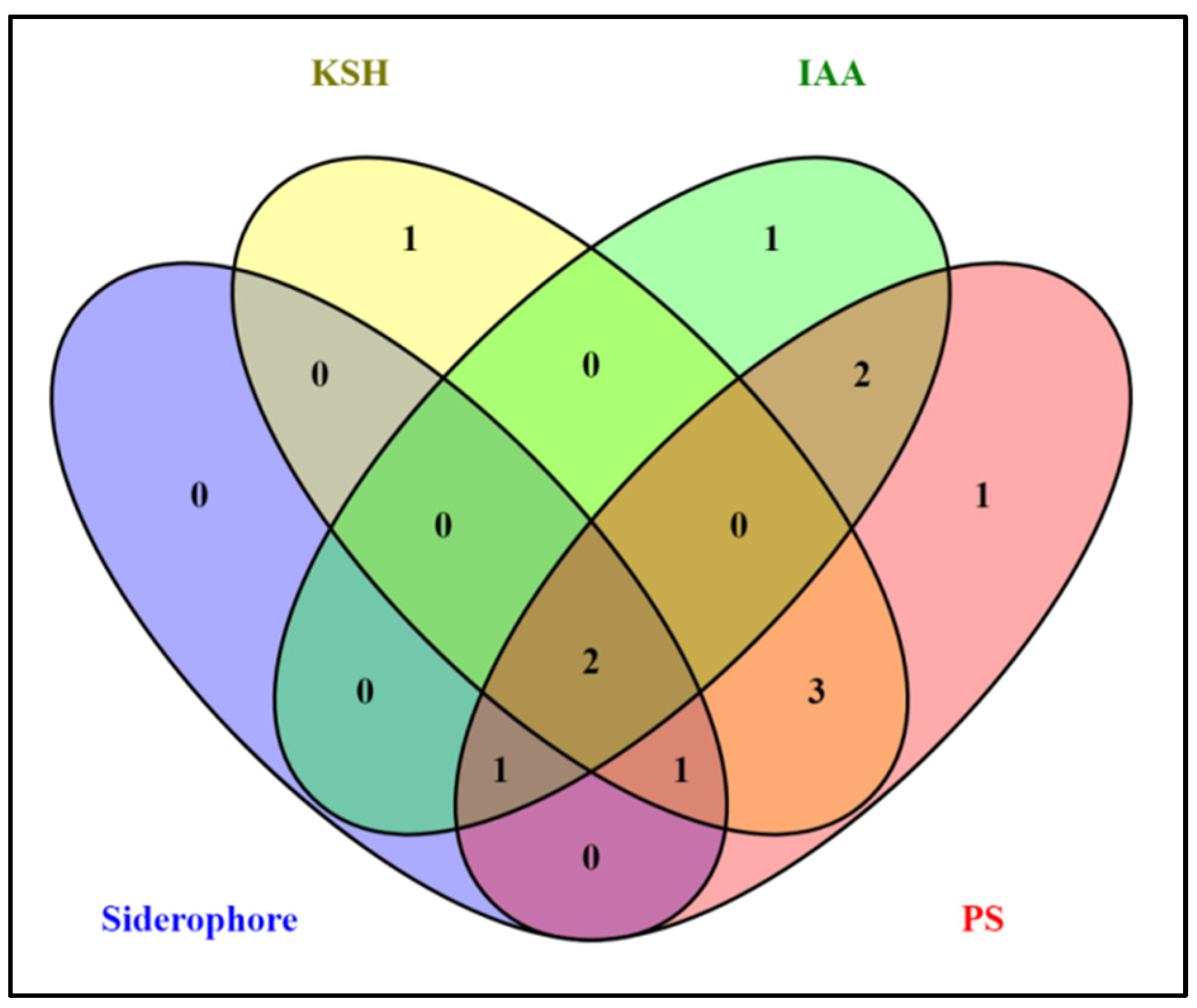
2.2. PGP Attributes of Beneficial Isolates and Venn Analysis
2.3. Synergistic Interaction and Bacterial Consortium
2.4. Molecular Characterization of Bacterial Isolates
2.5. Selection and Physico-Chemical Analysis of Supporting Materials
2.6. Preparation of Bioformulation
2.7. The Viability and Shelf Life of Bacterial Strains
2.8. Field Experiments
2.9. Physico-Chemical Properties of Pre-Harvesting and Post-Harvesting Soil
2.10. Proximate Composition Analysis
2.11. Statistical Analyses
3. Results
3.1. PGP Characterization, and the Identification of Endophytism
3.2. Supporting Materials
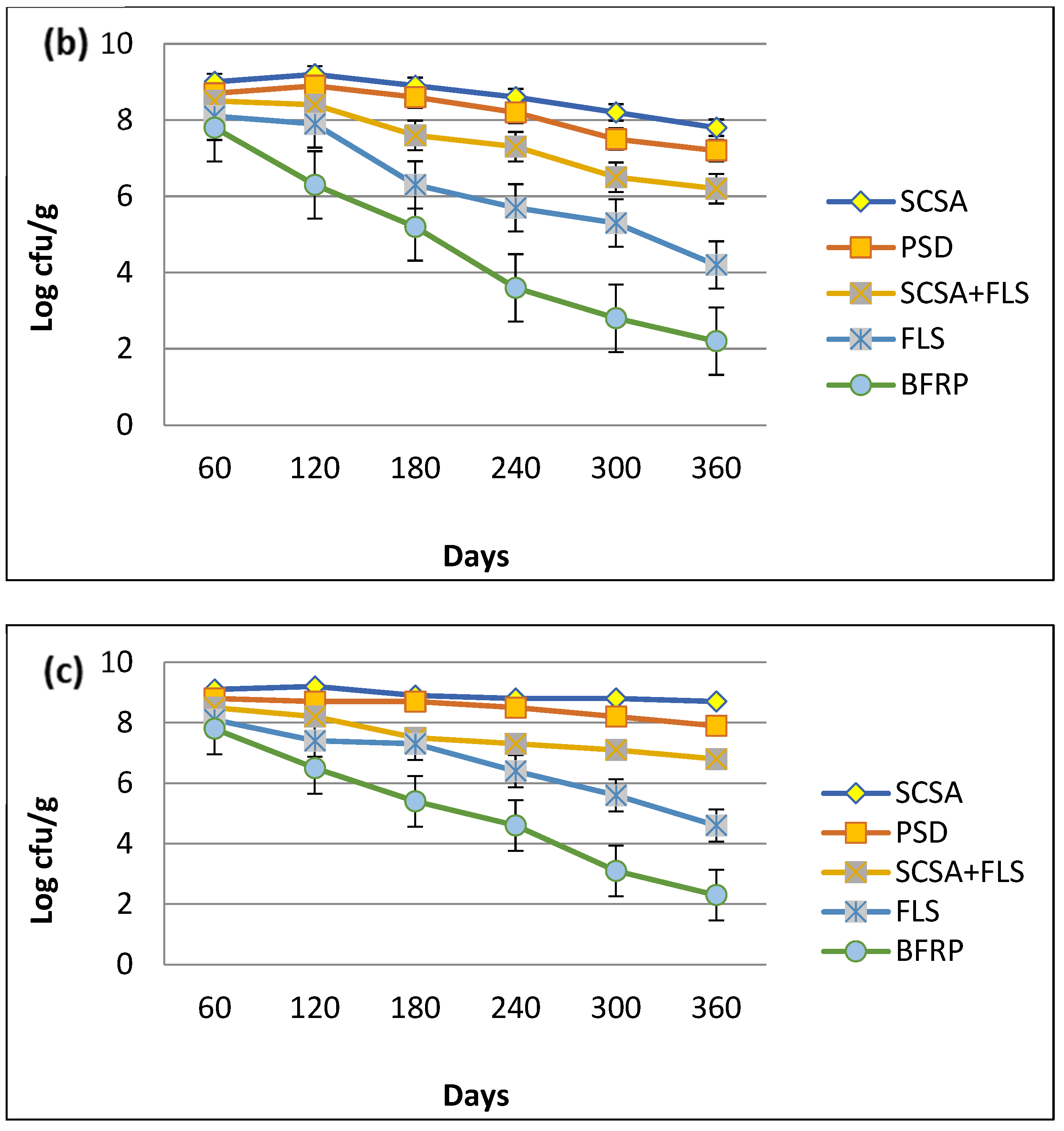
3.3. Determination of Shelf-Life of MSN12 and MEN8 Strains on Different Supporting Materials
3.4. Field Studies
3.4.1. Soil Analysis before Sowing
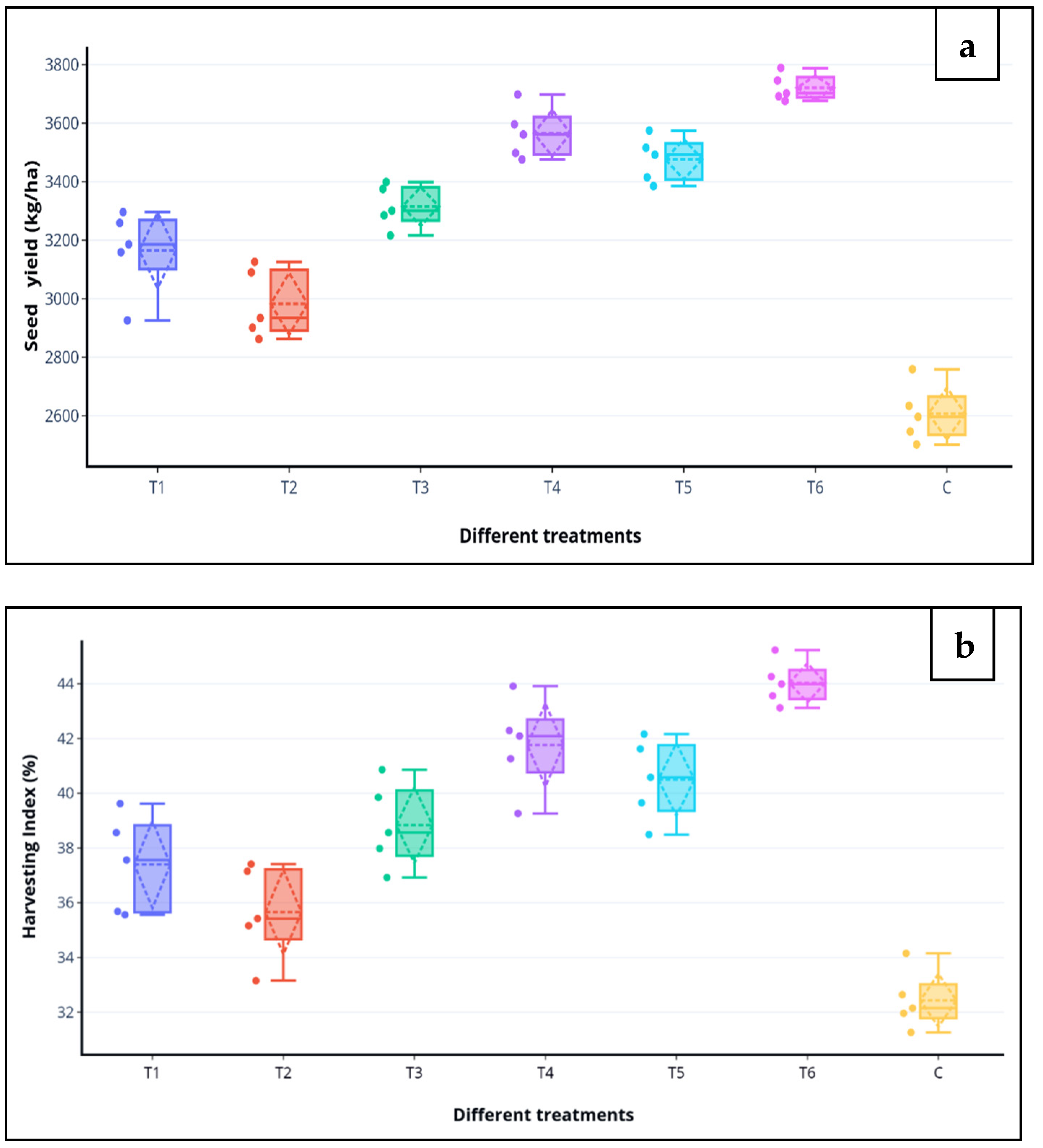
3.4.2. Effects on Seed Germination, Growth and Yield of Chickpea
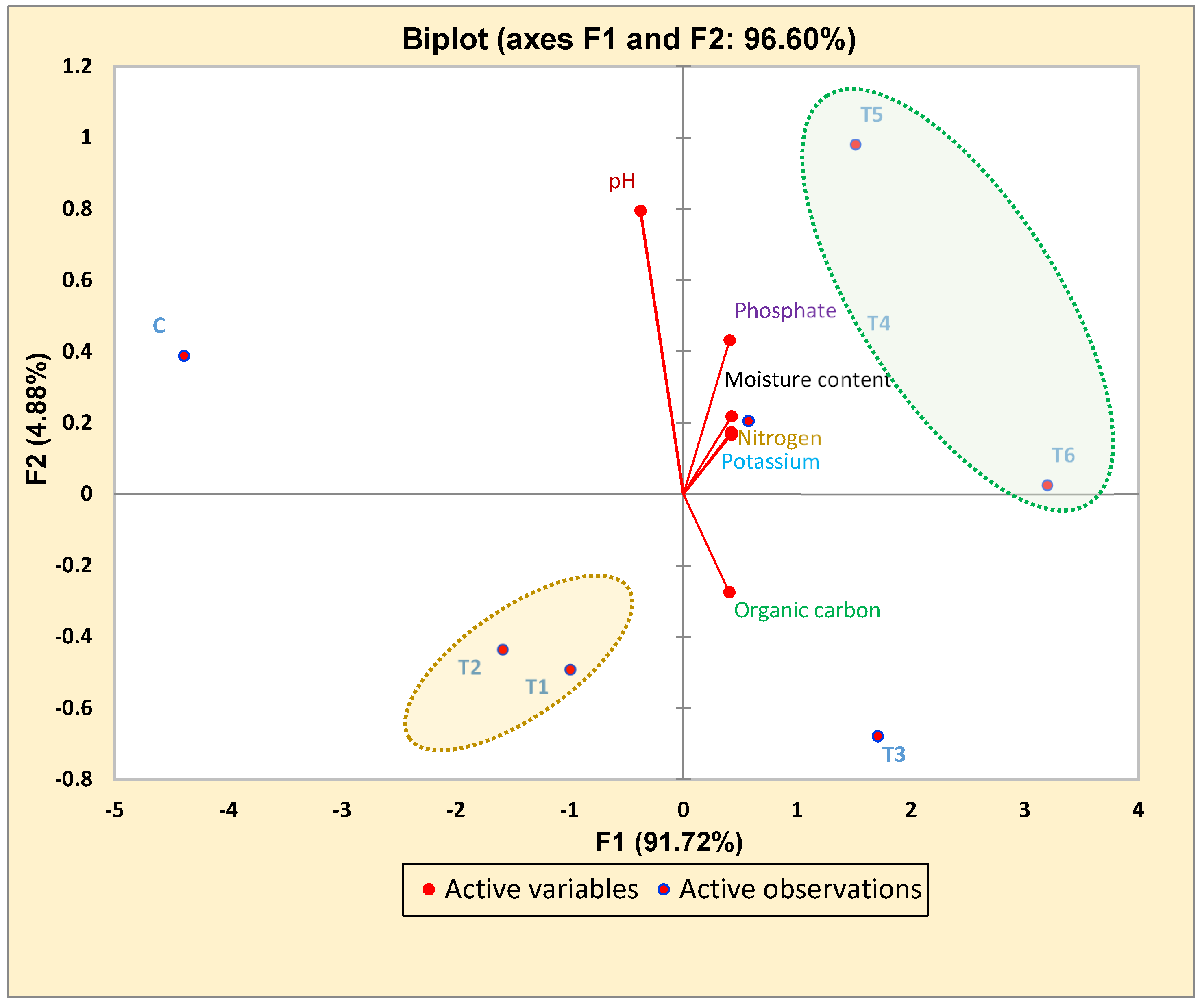
3.5. Correlated Study of Pre- and Post-Harvested Soil via Principal Component Analysis (PCA)
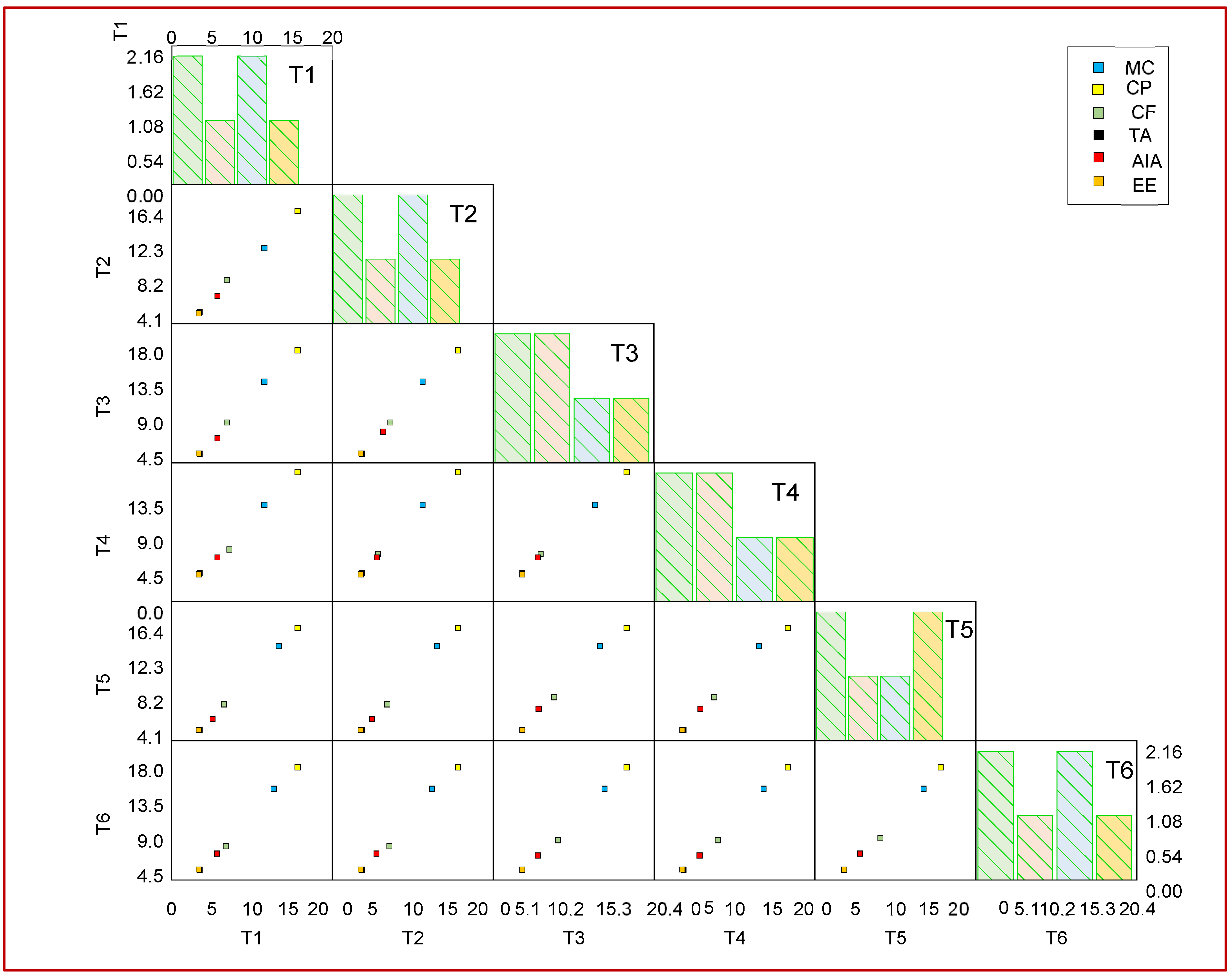
3.6. Correlation Study of Proximate Components via Scatter Matrix Visualization
4. Discussion
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, A. Microbial Biocontrol: Sustainable Agriculture and Phytopathogen Management; Springer Nature Chem: Cham, Switzerland, 2022; Volume 1, pp. 1–369. [Google Scholar]
- Arora, N.K.; Mehnaz, S.; Balestrini, R. Bioformulations: For Sustainable Agriculture; Springer: New Delhi, India, 2016; pp. 1–283. [Google Scholar]
- Pandey, C.; Negi, Y.K.; Maheshwari, D.K.; Rawat, D.; Prabha, D. Potential of native cold tolerant plant growth promoting bacilli to enhance nutrient use efficiency and yield of Amaranthus hypochondriacus. Plant Soil 2018, 428, 307–320. [Google Scholar] [CrossRef]
- Dhiman, S.; Dubey, R.C.; Maheshwari, D.K.; Kumar, S. Sulfur-oxidizing buffalo dung bacteria enhance growth and yield of Foeniculum vulgare Mill. Can. J. Microbiol. 2019, 65, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, D.K.; Dubey, R.C.; Agarwal, M.; Dheeman, S.; Aeron, A.; Bajpai, V.K. Carrier based formulations of biocoenotic consortia of disease suppressive Pseudomonas aeruginosa KRP1 and Bacillus licheniformis KRB1. Ecol. Eng. 2015, 81, 272–277. [Google Scholar] [CrossRef]
- Wang, H.-Y.; Liu, S.; Zhai, L.-M.; Zhang, J.-Z.; Ren, T.-Z.; Fan, B.-Q.; Liu, H.-B. Preparation and utilization of phosphate biofertilizers using agricultural waste. J. Integr. Agric. 2015, 14, 158–167. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Sathya, A.; Vijayabharathi, R.; Srinivas, V. Formulations of Plant Growth-Promoting Microbes for Field Applications. In Microbial Inocul Sustain Agricul Prod; Springer: New Delhi, India, 2016; pp. 239–251. [Google Scholar]
- Sharma, M.; Ghosh, R. An update on genetic resistance of chickpea to Ascochyta blight. Agronomy 2016, 6, 18. [Google Scholar] [CrossRef] [Green Version]
- FAOSTAT. Available online: http://faostat.fao.org/site/567/DesktopDefault.aspx (accessed on 15 June 2022).
- Mishra, R. Ecology Work-Book; Oxford and IBH publishing Co: New Delhi, India, 1968. [Google Scholar]
- Thomas, P.; Sekhar, A.C.; Upreti, R.; Mujawar, M.M.; Pasha, S.S. Optimization of single plate-serial dilution spotting (SP-SDS) with sample anchoring as an assured method for bacterial and yeast cfu enumeration and single colony isolation from diverse samples. Biotechnol. Rep. 2015, 8, 45–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baliyan, N.; Dheeman, S.; Maheshwari, D.K.; Dubey, R.C.; Vishnoi, V.K. Rhizobacteria isolated under field first strategy improved chickpea growth and productivity. Environ. Sustain. 2018, 4, 461–469. [Google Scholar] [CrossRef]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indole acetic acid. Plant Physiol. 1951, 26, 192–198. [Google Scholar] [CrossRef] [Green Version]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Hu, X.; Chen, J.; Guo, J. Two phosphate-and potassium-solubilizing bacteria isolated from Tianmu Mountain, Zhejiang, China. World J. Microbiol. Biotechnol. 2006, 22, 983–990. [Google Scholar] [CrossRef]
- Pikovskaya, R.I. Mobilization of phosphorus and soil in connection with the vital activity of some microbial species. Microbiology 1948, 17, 362–370. [Google Scholar]
- Pierson, E.A.; Weller, D.M. Use of mixtures of fluorescent pseudomonads to suppress take all and improve the growth of wheat. Phytopathology 1994, 84, 940–947. [Google Scholar] [CrossRef]
- Pandey, P.; Maheshwari, D.K. Bioformulation of Burkholderia sp. MSSP with a multispecies consortium for growth promotion of Cajanus cajan. Can. J. Microbiol. 2007, 53, 213–222. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Rodrigues, K.M.; Rodrigues, B.F. Development of carrier based in vitro produced arbuscular mycorrhizal (AM) fungal inocula for organic agriculture. Ann. Adv. Agric. Sci. 2017, 1, 26–37. [Google Scholar]
- Namsena, P.; Buscsaman, P.; Rattanasena, P. Bioformulation of Xenorhabdusstockiae PB09 for controlling mushroom mite, Luciaphorusperniciosus Rack. Bioresour. Bioprocess. 2016, 3, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Bureau of Indian Standards (BIS). Specification for Inoculants; Indian Standards Specification: New Delhi, India, 2000. [Google Scholar]
- Biradar, B.P.; Santhosh, G.P. Cell Protectants, Adjuvants, Surfactant and Preservative and their Role in Increasing the Shelf Life of Liquid Inoculant Formulations of Pseudomonas fluorescens. Int. J. Pure Appl. Biosci. 2018, 6, 116–122. [Google Scholar] [CrossRef]
- RBDC India. Regional Biofertilizers Development Centre, Department of Agriculture and Co-op. Govt of India.. Available online: http://rbdcbangalore.dacnet.nic.in/bfahelp.htm (accessed on 15 June 2022).
- Kumar, A.; Maiti, S.K.; Prasad, M.N.V.; Singh, R.S. Grasses and legumes facilitate phytoremediation of metalliferous soils in the vicinity of an abandoned chromite–asbestos mine. J. Soils Sediments 2017, 17, 1358–1368. [Google Scholar] [CrossRef]
- Subbiah, B.V.; Asija, G.L. A rapid procedure for the estimation of available N in the soils. Curr. Sci. 1956, 25, 259. [Google Scholar]
- Olsen, S.R.; Sommers, L.E. Phosphorus. In Methods of Soil Analysis, 2nd ed.; Page, A.L., Ed.; ASA and SSSA: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Warncke, D.; Brown, J.R. Potassium and Other Basic Cations. Recommended Chemical Soil Test Procedures for the North Central Region; NCR Publication: Columbia, MO, USA, 1998; Volume 1001, pp. 31–45. [Google Scholar]
- Walkley, A.E.; Black, J.A. An examination of the Degtjareff method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Marioli Nobile, C.; Carreras, J.; Grosso, R.; Inga, C.M.; Silva, M.P.; Aguilar, R.; Allende, M.J.; Badini, R.; Martinez, M.J. Proximate composition and seed lipid components of “kabuli”-type chickpea (Cicer arietinum L.) from Argentina. J. Sci. Food Agric. 2013, 4, 729–735. [Google Scholar]
- Baliyan, N.; Dhiman, S.; Dheeman, S.; Kumar SArora, N.K.; Maheshwari, D.K. Optimization of gibberellic acid production in endophytic Bacillus cereus using response surface methodology and its use as plant growth regulator in chickpea. J. Plant Growth Regul. 2021, 41, 3019–3029. [Google Scholar] [CrossRef]
- Koskey, G.; Mburu, S.W.; Awino, R.; Njeru, E.M.; Maingi, J.M. Potential use of beneficial microorganisms for soil amelioration, phytopathogen biocontrol, and sustainable crop production in smallholder agroecosystems. Front. Sustain. Food Syst. 2021, 5, 130. [Google Scholar] [CrossRef]
- Trivedi, P.; Pandey, A.; Palni, L.M.S. Bacterial inoculants for field applications under mountain ecosystem: Present initiatives and future prospects. In Bacteria Agrobiology: Plant Probiotics; Springer: Berlin/Heidelberg, Germany, 2012; pp. 15–44. [Google Scholar]
- Thirumal, G. Study of the Different Factors Influencing the Shelf Life of Carrier Based Biofertilizers. Ph.D. Thesis, Professor Jayashankartelangana State Agricultural University, Hyderabad, India, 2016. [Google Scholar]
- Baliyan, N.; Dhiman, S.; Dheeman, S.; Kumar, S.; Maheshwari, D.K. Optimization of indole-3-acetic acid using response surface methodology and its effect on vegetative growth of chickpea. Rhizosphere 2021, 17, 100321. [Google Scholar] [CrossRef]
- Gontia-Mishra, I.; Sapre, S.; Kachare, S.; Tiwar, S. Molecular diversity of 1-aminocyclopropane-1-carboxylate (ACC) deaminase producing PGPR from wheat (Triticum aestivum L.) rhizosphere. Plant Soil 2017, 414, 213–227. [Google Scholar] [CrossRef]
- Meena, V.S.; Maurya, B.R.; Verma, J.P.; Aeron, A.; Kumar, A.; Kim, K.; Bajpai, V.K. Potassium solubilizing rhizobacteria (KSR): Isolation, identification, and K-release dynamics from waste mica. Ecol. Eng. 2015, 81, 340–347. [Google Scholar] [CrossRef]
- Arora, N.K.; Kang, S.C.; Maheshwari, D.K. Isolation of siderophore-producing strains of Rhizobium meliloti and their biocontrol potential against Macrophomina phaseolina that causes charcoal rot of groundnut. Curr. Sci. 2001, 81, 673–677. [Google Scholar]
- Sahu, P.K.; Gupta, A.; Singh, M.; Mehrotra, P.; Brahmaprakash, G.P. Bioformulation and Fluid Bed Drying: A New Approach Towards an Improved Biofertilizer Formulation. In Eco-Friendly Agro-Biological Techniques for Enhancing Crop Productivity; Springer Gateway East: Singapore, 2018; pp. 47–62. [Google Scholar]
- Van Antwerpen, R.; Meyer, J.H. The effect of burning and trashing on sugarcane leaf analysis. Proc. S. Afr. Sug. Technol. Ass. 2002, 76, 586–595. [Google Scholar]
- Brockwell, J.; Bottomley, P.J. Recent advances in inoculant technology and prospects for the future. Soil Biol. Biochem. 1996, 27, 683–697. [Google Scholar] [CrossRef]
- Reddy, T.S.; Prasad, D.S.V. Stabilization of Soil Using Sugarcane Straw Ash and Polypropylene Fibres. Int. J. Eng. Appl. Sci. 2017, 4, 6–12. [Google Scholar]
- Paliya, S.; Mandpe, A.; Kumar, S.; Kumar, M.S. Enhanced nodulation and higher germination using sludge ash as a carrier for biofertilizer production. J. Environ. Manag. 2019, 250, 109523. [Google Scholar] [CrossRef] [PubMed]
- Suman, A.; Verma, P.; Yadav, A.N.; Srinivasamurthy, R.; Singh, A.; Prasanna, R. Development of hydrogel based bio-inoculant formulations and their impact on plant biometric parameters of wheat (Triticum aestivum L.). Int. J. Curr. Microbiol. Appl. Sci. 2016, 5, 890–901. [Google Scholar] [CrossRef]
- Kushwaha, D.S. Comparison of effect of biofertilizers on seedling growth and available nutrients in soil of sesame (Sesamum indicum L.) varieties. J. Environ. Res. Dev. 2011, 5, 631–637. [Google Scholar]
- Chinnusamy, M.; Kaushik, B.D.; Radha, P. Growth, nutritional and yield parameters of wetland rice as influenced by microbial consortia under controlled conditions. Food Sci. Nutr. 2006, 29, 857–871. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physical Parameters | PSD | SCSA | FLS | SCSA + FLS | BFRP |
|---|---|---|---|---|---|
| Moisture content (%) | 11.6 ± 0.01 | 12.5 ± 0.01 | 3.7 ± 0.01 | 8.3 ± 0.01 | 0.9 ± 0.01 |
| % Water holding capacity (WHC) | 76 ± 1.46 | 89 ± 1.51 | 68 ± 0.45 | 72 ± 0.93 | 42 ± 0.35 |
| pH [1:2.5 (w/v)] | 6.2 ± 0.01 | 6.8 ± 0.01 | 6.9 ± 0.01 | 7.2 ± 0.01 | 6.0 ± 0.01 |
| Chemical oxygen demand (COD) (mg/L) | 25.4 ± 0.92 | 32.4 ± 0.98 | 16.5 ± 0.92 | 21.3 ± 0.91 | 11.5 ± 0.90 |
| Biological oxygen demand (BOD) (mg/L) | 20.6 ± 0.85 | 28.4 ± 0.86 | 22.1 ± 0.72 | 19.6 ± 0.71 | 14.4 ± 0.68 |
| Total carbon (%) | 18.5 ± 0.68 | 24.8 ± 0.72 | 17.3 ± 0.65 | 15.4 ± 0.67 | 12.6 ± 0.61 |
| Total nitrogen (%) | 1.4 ± 0.01 | 1.5 ± 0.01 | 1.1 ± 0.01 | 1.1 ± 0.01 | 0.06 ± 0.00 |
| Different Treatments | SG (%) a | Plant Weight (g) b | Plant Length (cm) b | ||
|---|---|---|---|---|---|
| Fresh | Dry | Root | Shoot | ||
| T1 (MEN8) | 90.9 ** | 3.37 * | 0.89 ** | 11.2 ** | 23.4 ** |
| T2 (MSN12) | 88.1 ** | 3.14 * | 0.81 ** | 10.2 ** | 19.3 ** |
| T3 (MSN12 + MEN8) | 91.4 ** | 4.24 * | 1.17 ** | 13.7 ** | 25.3 ** |
| T4 (MEN8 + SCSA) | 94.6 ** | 4.79 * | 1.35 ** | 14.6 ** | 24.9 ** |
| T5 (MSN12 + SCSA) | 92.2 ** | 4.38 * | 1.23 ** | 13.5 ** | 23.2 ** |
| T6 (MSN12 + MEN8 + SCSA) | 97.8 ** | 5.42 * | 1.58 ** | 16.2 ** | 29.6 ** |
| C (without bio-inoculant and carrier) | 56.5 | 2.83 | 0.59 | 9.8 | 16.5 |
| SEM | 1.7 | 0.63 | 0.12 | 0.61 | 0.72 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baliyan, N.; Qureshi, K.A.; Jaremko, M.; Rajput, M.; Singh, M.; Dhiman, S.; Maheshwari, D.K.; Kant, C.; Kumar, A. Bioformulation Containing Cohorts of Ensifer adhaerens MSN12 and Bacillus cereus MEN8 for the Nutrient Enhancement of Cicer arietinum L. Plants 2022, 11, 3123. https://doi.org/10.3390/plants11223123
Baliyan N, Qureshi KA, Jaremko M, Rajput M, Singh M, Dhiman S, Maheshwari DK, Kant C, Kumar A. Bioformulation Containing Cohorts of Ensifer adhaerens MSN12 and Bacillus cereus MEN8 for the Nutrient Enhancement of Cicer arietinum L. Plants. 2022; 11(22):3123. https://doi.org/10.3390/plants11223123
Chicago/Turabian StyleBaliyan, Nitin, Kamal A. Qureshi, Mariusz Jaremko, Minakshi Rajput, Monika Singh, Sandhya Dhiman, Dinesh Kumar Maheshwari, Chandra Kant, and Ajay Kumar. 2022. "Bioformulation Containing Cohorts of Ensifer adhaerens MSN12 and Bacillus cereus MEN8 for the Nutrient Enhancement of Cicer arietinum L." Plants 11, no. 22: 3123. https://doi.org/10.3390/plants11223123
APA StyleBaliyan, N., Qureshi, K. A., Jaremko, M., Rajput, M., Singh, M., Dhiman, S., Maheshwari, D. K., Kant, C., & Kumar, A. (2022). Bioformulation Containing Cohorts of Ensifer adhaerens MSN12 and Bacillus cereus MEN8 for the Nutrient Enhancement of Cicer arietinum L. Plants, 11(22), 3123. https://doi.org/10.3390/plants11223123