


Plant Pyranocoumarins: Description, Biosynthesis, Application

 and
and
Abstract
:1. Introduction
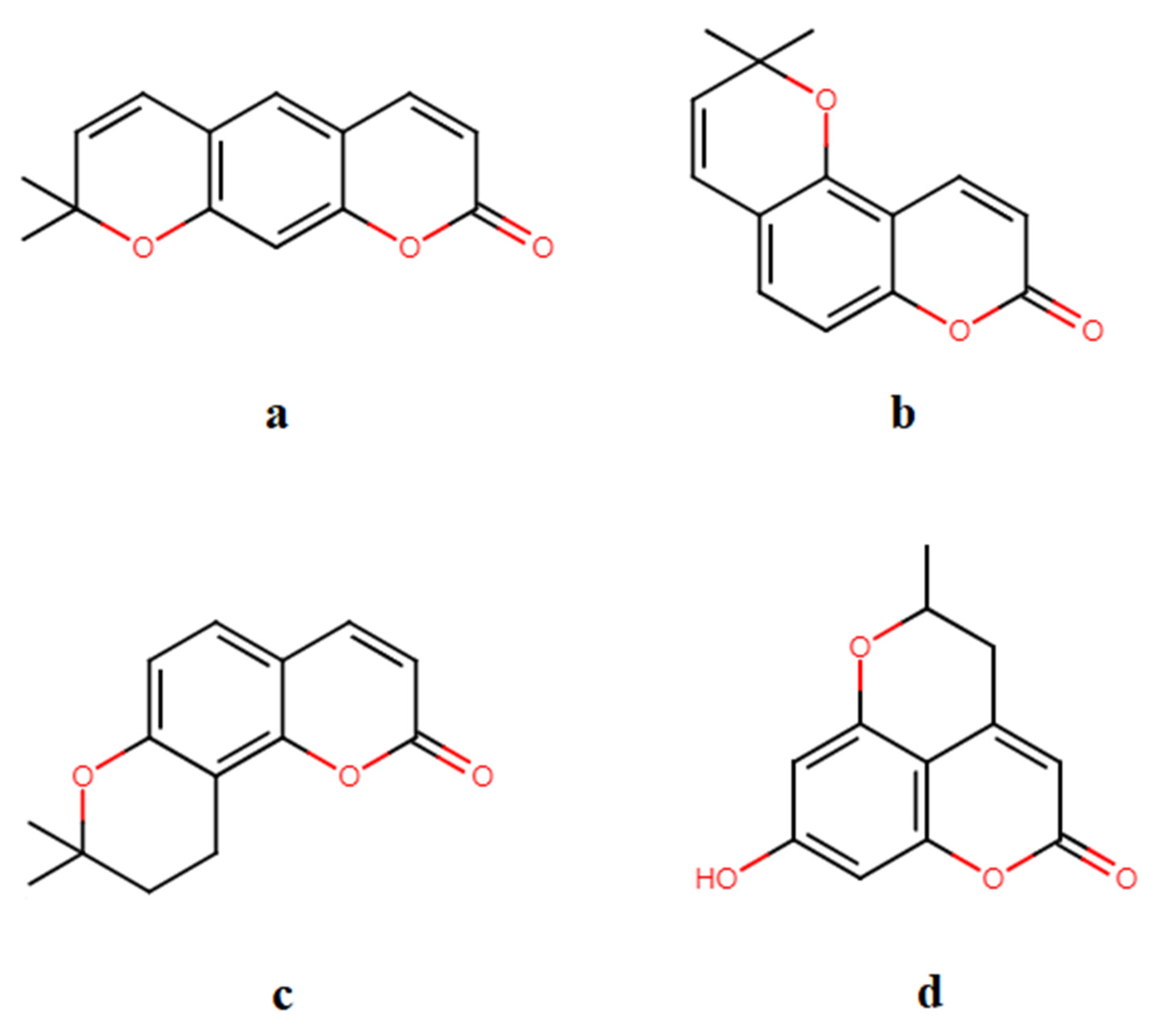
2. Pyranocoumarins Structure and Classification
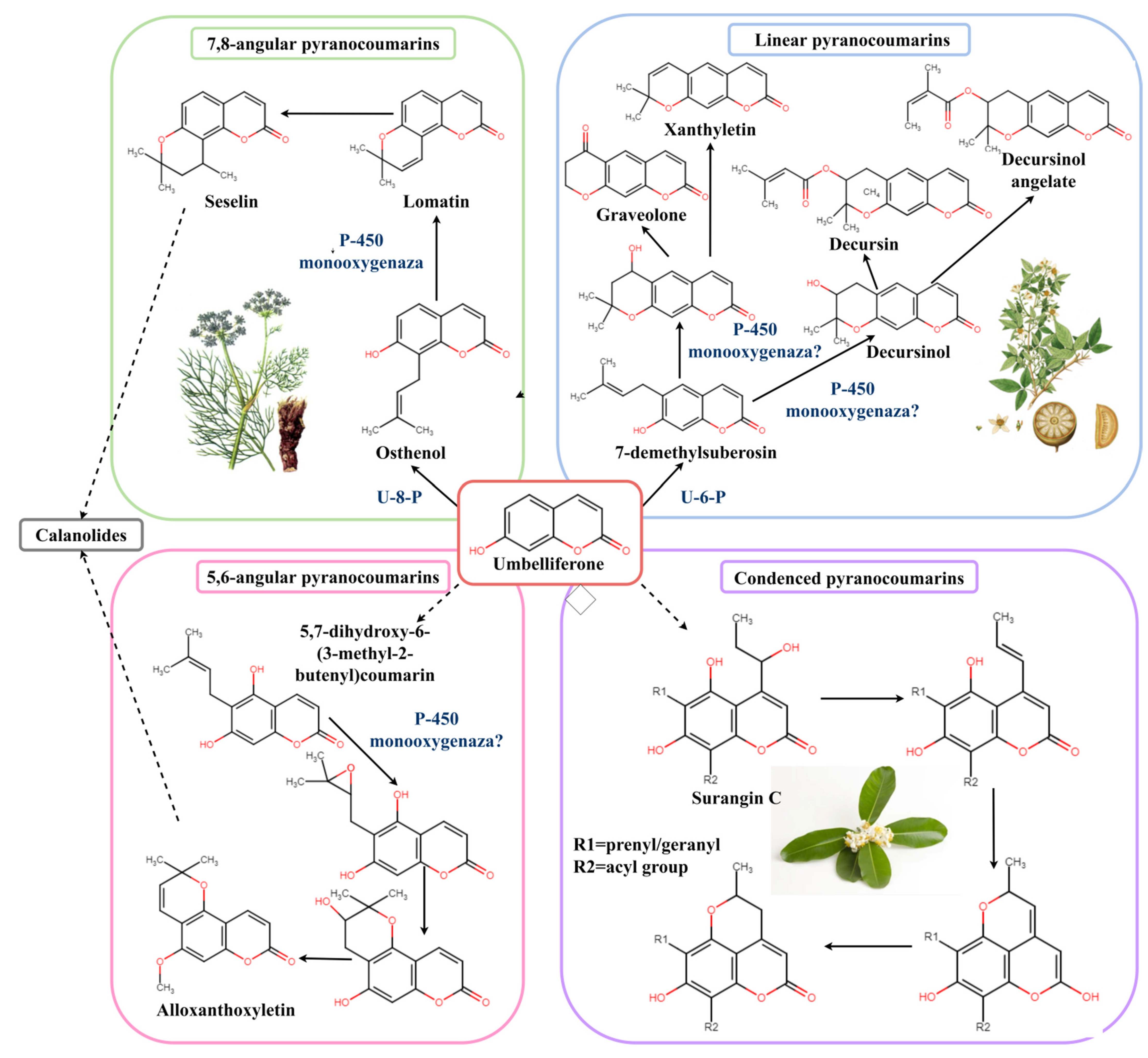
3. Biosynthesis of Pyranocoumarins in Cells
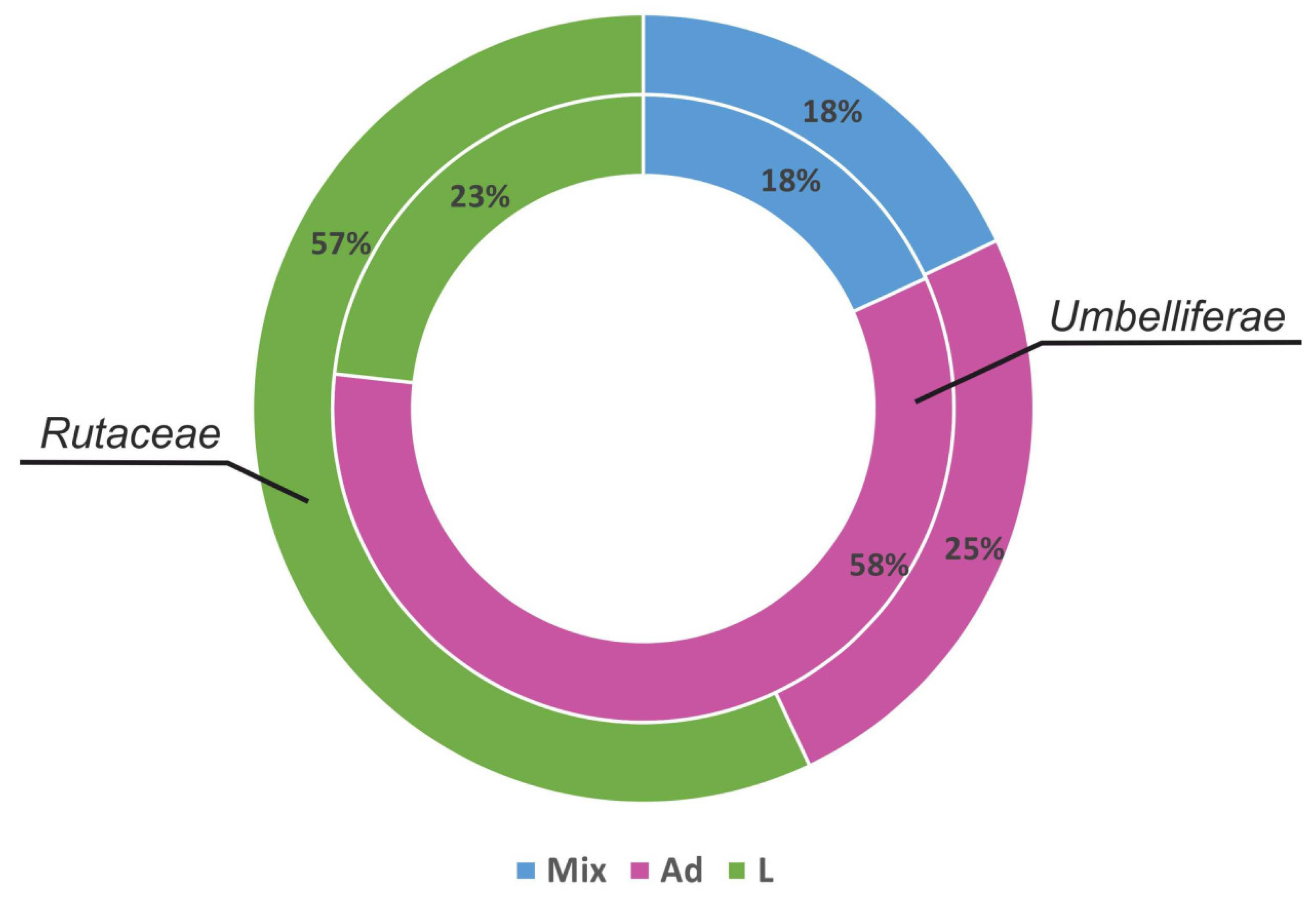
4. Pyranocoumarins Distribution in Plant World
5. Biological Activity of Pyranocoumarins
5.1. Anti-Inflammatory Activity
5.2. Antioxidant Activity
5.3. Antimicrobial Activity
5.4. Anti-Cancer Activity
5.5. Antivirus Activity
5.6. Antihyperglycemic and Antidyslipidemic Activity
5.7. Other Activities
6. Prospects for the Pyranocoumarins Study
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bynum, H.; Bynum, W. Remarkable Plants That Shape Our World; University of Chicago Press: Chicago, IL, USA, 2014; p. 239. ISBN 978-0226204741. [Google Scholar]
- Veeresham, C. Natural products derived from plants as a source of drugs. J. Adv. Pharm. Technol. Res. 2012, 3, 200–201. [Google Scholar] [CrossRef]
- Soriano-Castell, D.; Liang, Z.; Maher, P.; Currais, A. The search for anti-oxytotic/ferroptotic compounds in the plant world. Br. J. Pharmacol. 2021, 178, 3611–3626. [Google Scholar] [CrossRef]
- Ledford, H. World’s largest plant survey reveals alarming extinction rate. Nature 2019, 570, 148–149. [Google Scholar] [CrossRef] [Green Version]
- Zobel, A.M.; Brown, S.A. Coumarins in the interections between the plant and its environment. Allelopath. J. 1995, 2, 9–20. [Google Scholar]
- Babekov, A.U.; Keneshov, B.M.; Mamytov, T.A.; Tynybekova, G.T. Pharmacological activity of some biologically active substances. Izv. OshTU 2008, 2, 118–121. [Google Scholar]
- Beena, K.P.; Sathya Pooja, G. A Review on Pyranocoumarins and its biological importance. Asian J. Res. Chem. 2022, 15, 176–181. [Google Scholar] [CrossRef]
- Vogel, A. Darstellung von Benzoesäure aus der Tonka-Bohne und aus den Meliloten—Oder Steinklee—Blumen. Annal. Phyhk. 1820, 64, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.; Hosansky, N.; Bywater, W.G.; Tamelen, E.E. Constitution of Samidin, Dihydrosamidin and Visnadin. J. Am. Chem. Soc. 1956, 79, 3534–3540. [Google Scholar] [CrossRef]
- Lozhkin, A.V.; Sakanyan, E.I. Natural coumarins: Methods of extraction. Chem. Pharm. J. 2006, 40, 47–56. [Google Scholar] [CrossRef]
- Joshi, P.P. A Review in Biological Activities of Linear Pyranocoumarins. Int. J. Adv. Study Res. Work. 2021, 4, 20–27. [Google Scholar] [CrossRef]
- Reddy, C.S.; Kim, S.; Hur, M.; Kim, Y.; Park, C.; Lee, W.; Jang, J.; Koo, S. Natural Korean Medicine Dang-Gui: Biosynthesis, Effective Extraction and Formulations of Major Active Pyranocoumarins, Their Molecular Action Mechanism in Cancer, and Other Biological Activities. Molecules 2017, 22, 2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Robledo, H.B.; Cruz-Sosa, F.; Bernabé-Antonio, A.; Guerrero-Analco, A.; Olivares-Romero, J.B.; Alexandro Alonso-Sánchez, A.; Emanuel Villafán, E.; Ibarra-Laclette, E. Identification of candidate genes related to calanolide biosynthesis by transcriptome sequencing of Calophyllum brasiliense (Calophyllaceae). BMC Plant Biol. 2016, 16, 177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, L.; Jiang, C.; Xing, C.; Kim, S.-H.; Lu, J. Anti-cancer and Other Bioactivities of Korean Angelica gigas Nakai (AGN) and Its Major Pyranocoumarin Compounds. Anti-Cancer Agents Med. Chem. 2012, 12, 1239–1254. [Google Scholar] [CrossRef] [PubMed]
- Amaral, J.C.; Silva, M.M.; Silva, M.F.G.F.; Alves, T.C.; Ferreira, G.; Forim, M.R.; Fernandes, J.B.; Pina, E.S.; Lopes, A.A.; Pereira, A.M.S.; et al. Advances in the Biosynthesis of Pyranocoumarins: Isolation and 13C-Incorporation Analysis by High-Performance Liquid Chromatography–Ultraviolet–Solid-Phase Extraction–Nuclear Magnetic Resonance Data. J. Nat. Prod. 2020, 83, 1409–1415. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, T.; Luo, J.; Zhang, Q.; Xu, S.; Han, C.; Xu, J.; Chen, M.; Chen, Y.; Kong, L. Integration of a Decrescent Transcriptome and Metabolomics Dataset of Peucedanum praeruptorum to Investigate the CYP450 and MDR Genes Involved in Coumarins Biosynthesis and Transport. Front. Plant Sci. 2015, 6, 996. [Google Scholar] [CrossRef] [Green Version]
- Mahandru, M.M.; Ravindran, V.K. Surangin C, a Coumarin from Mammea longifolia. Phytochemistry 1986, 25, 555–556. [Google Scholar] [CrossRef]
- Prachyawarakorn, V.; Mahidol, C.; Ruchirawat, S. Pyranocoumarins from the twigs of Mammea siamensis. Phytochemistry 2006, 67, 924–928. [Google Scholar] [CrossRef]
- Lupini, A.; Araniti, F.; Sunseri, F.; Abenavoli, M.R. Gravitropic response induced by coumarin: Evidences of ROS distribution involvement. Plant Signal. Behav. 2013, 8, e23156. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Gruber, M.Y.; Hegedus, D.D.; Lydiate, D.J.; Gao, M.-J. Effects of a Coumarin Derivative, 4-Methylumbelliferone, on Seed Germination and Seedling Establishment in Arabidopsis. J. Chem. Ecol. 2011, 37, 880. [Google Scholar] [CrossRef]
- Srinivasan, S.; Sarada, D.V.L. Antifungal Activity of Phenyl Derivative of Pyranocoumarin from Psoralea corylifolia L. Seeds by Inhibition of Acetylation Activity of Trichothecene 3-O-Acetyltransferase (Tri101). BioMed Res. Int. 2012, 2012, 310850. [Google Scholar] [CrossRef] [Green Version]
- Yoon, M.; Kim, Y.; Ryu, S.; Choi, G.; Jang, K.; Han, S.; Kim, J. In vitro and in vivo antifungal activities of decursin and decursinol angelate isolated from Angelica gigas against Magnaporthe oryzae, the causal agent of rice blast. Pestic. Biochem. Physiol. 2011, 101, 118–124. [Google Scholar] [CrossRef]
- Son, C.; Baek, I.; Song, G.; Kang, J.; Kwon, K. Pharmacological Effect of Decursin and Decursinol Angelate from Angelica gigas Nakai. Yakhak Hoeji 2009, 53, 303–313. [Google Scholar]
- Yu, P.; Jin, H.; Zhang, J.; Wang, G.; Li, J.; Zhu, Z.; Tian, Y.; Wu, S.; Xu, W.; Zhang, J.; et al. Pyranocoumarins Isolated from Peucedanum praeruptorum Dunn Suppress Lipopolysaccharide-Induced Inflammatory Response in Murine Macrophages Through Inhibition of NF-kB and STAT3 Activation. Inflammation 2012, 35, 967–977. [Google Scholar] [CrossRef]
- Lee, J.; Lee, Y.; Kim, J.; Bang, O. Pyranocoumarins from Root Extract of Peucedanum praeruptorum Dunn Multidrug Resistance Reversal and Anti-Inflammatory Activities. Molecules 2015, 20, 20967–20978. [Google Scholar] [CrossRef] [Green Version]
- Chun, J.; Tosun, A.; Kim, Y.S. Anti-inflammatory effect of corymbocoumarin from Seseli gummiferum subsp. corymbosum through suppression of NF-κB signaling pathway and induction of HO-1 expression in LPS-stimulated RAW 264.7 cells. Int. Immunopharmacol. 2016, 31, 207–215. [Google Scholar] [CrossRef]
- Ouyang, G.Q.; Li, C.J.; Yang, J.Z.; Li, L.; Song, X.Y.; Jiang, Y.N.; Chen, N.H.; Ma, J.; Zhang, D.M. Bioactive Coumarins from the Stems of Clausena emarginata. Chem. Biodivers. 2016, 13, 1178–1185. [Google Scholar] [CrossRef]
- Menghini, L.; Epifano, F.; Genovese, S.; Marcotullio, M.C.; Sosa, S.; Tubaro, A. Antiinflammatory activity of coumarins from Ligusticum lucidum Mill. subsp. cuneifolium (Guss.) Tammaro (Apiaceae). Phytother. Res. 2010, 24, 1697–1699. [Google Scholar] [CrossRef]
- Alshibl, H.M.; Al-Abdullah, E.S.; Haiba, M.E.; Alkahtani, H.M.; Awad, G.E.A.; Mahmoud, A.H.; Ibrahim, B.M.M.; Bari, A.; Villinger, A. Synthesis and Evaluation of New Couamrin as Antioxidant, Antimicrobal, and Anti-Inflammatory Agents. Molecules 2020, 25, 3251. [Google Scholar] [CrossRef]
- Munteanu, I.G.; Apetrei, C. Analytical Methods Used in Determining Antioxidant Activity: A Review. Int. J. Mol. Sci. 2021, 22, 73380. [Google Scholar] [CrossRef]
- Domínguez-Mendoza, E.A.; Cornejo-Garrido, J.; Burgueno-Tapia, E.; Ordaz-Pichardo, C. Antidiabetic effect, antioxidant activity, and toxicity of 3′,4′-Di-O-acetyl-cis-khellactone in Streptozotocin-induced diabetic rats. Bioorganic Med. Chem. Lett. 2016, 26, 4086–4091. [Google Scholar] [CrossRef]
- Lee, S.; Shin, D.S.; Kim, J.S.; Oh, K.B.; Kang, S.S. Antibacterial coumarins from Angelica gigas roots. Arch. Pharm. Res. 2003, 26, 449–452. [Google Scholar] [CrossRef] [PubMed]
- Basile, A.; Sorbo, S.; Spadaro, V.; Bruno, M.; Maggio, A.; Faraone, N.; Rosselli, S. Antimicrobial and Antioxidant Activities of Coumarins from the Roots of Ferulago campestris (Apiaceae). Molecules 2009, 14, 939–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, P.P.; Zhao, J.; Liu, Z.L.; Duan, Y.B.; Hou, Y.P.; Zhao, C.Q.; Wu, M.; Wei, M.; Wang, N.H.; Lv, Y.; et al. Evaluation of antifungal activities and structure-activity relationships of coumarin derivatives. Pest. Manag. Sci. 2017, 73, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, Y.S.; Jung, S.H.; Shin, K.H.; Kim, B.K.; Kang, S.S. Anti-tumor activities of decursinol angelate and decursin from Angelica gigas. Arch. Pharm. Res. 2003, 26, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Yim, D.; Singh, R.P.; Agarwal, C.; Lee, S.; Chi, H.; Agarwal, R. A Novel Anticancer Agent, Decursin, Induces G1 Arrest and Apoptosis in Human Prostate Carcinoma Cells. Cancer Res. 2005, 65, 1035–1044. [Google Scholar] [CrossRef]
- Jiang, C.; Lee, H.; Li, G.; Guo, J.; Malewicz, B.; Zhao, Y.; Lee, E.; Lee, H.; Lee, J.; Lim, M.; et al. Potent Antiandrogen and Androgen Receptor Activities of an Angelica gigas—Containing Herbal Formulation: Identification of Decursin as a Novel and Active Compound with Implications for Prevention and Treatment of Prostate Cancer. Cancer Res. 2006, 66, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Guo, J.; Wang, Z.; Xiao, B.; Lee, H.; Lee, E.; Kim, S.; Lu, J. Decursin and decursinol angelate inhibit estrogen-stimulated and estrogen-independent growth and survival of breast cancer cells. Breast Cancer Res. 2007, 9, R77. [Google Scholar] [CrossRef] [Green Version]
- Song, G.-Y.; Lee, J.-H.; Cho, M.; Park, B.-S.; Kim, D.-E.; Oh, S. Decursin Suppresses Human Androgen-Independent PC3 Prostate Cancer Cell Proliferation by Promoting the Degradation of β-Catenin. Mol. Pharmacol. 2007, 72, 1599–1606. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.R.; Lee, J.H.; Kim, J.Y.; Park, K.W.; Jeong, I.Y.; Shim, K.H.; Lee, M.K.; Seo, K.I. Decursin from Angelica gigas Nakai induces apoptosis in RC-58T/h/SA#4 primary human prostate cancer cells via a mitochondria-related caspase pathway. Food Chem. Toxicol. 2011, 49, 2517–2523. [Google Scholar] [CrossRef]
- Kim, S.A.; Oh, H.K.; Kim, J.Y.; Hong, J.W.; Cho, S.I. A Review of Pharmacological Effects of Angelica gigas, Angelica sinensis, Angelica acutiloba and their Bioactive Compounds. J. Korean Med. Sci. 2011, 32, 1–24. [Google Scholar]
- Yim, N.; Lee, J.; Cho, W.; Yang, M.; Kwak, D.; Ma, J. Decursin and decursinol angelate from Angelica gigas Nakai induce apoptosis via induction of TRAIL expression in cervical cancer cells. Eur. J. Integr. Med. 2011, 3, 299–307. [Google Scholar] [CrossRef]
- Urbagarova, B.M.; Shults, E.E.; Taraskin, V.V.; Radnaeva, L.D.; Petrova, T.N.; Rybalova, T.V.; Frolova, T.S.; Pokrovskii, A.G.; Ganbaatar, J. Chromones and coumarins from Saposhnikovia divaricata (Turcz.) Schischk. Growing in Buryatia and Mongolia and their cytotoxicity. J. Ethnopharmacol. 2020, 261, 112517. [Google Scholar] [CrossRef]
- Su, C.R.; Yeh, S.F.; Liu, C.M.; Damu, A.G.; Kuo, T.H.; Chiang, P.C.; Bastow, K.F.; Lee, K.H.; Wu, T.S. Anti-HBV and cytotoxic activities of pyranocoumarin derivatives. Bioorg. Med. Chem. 2009, 17, 6137–6143. [Google Scholar] [CrossRef]
- Liang, T.; Yue, W.; Li, Q. Chemopreventive Effects of Peucedanum praeruptorum DUNN and Its Major Constituents on SGC7901 Gastric Cancer Cells. Molecules 2010, 15, 8060–8071. [Google Scholar] [CrossRef]
- Gheibi, S.; Shokohinia, Y.; Kiani, A.; Sadrjavadi, K.; Nowroozi, A.; Shahlaei, M. Molecular insight into the Grandivitin- matrix metalloproteinase 9 interactions. J. Photochem. Photobiol. B Biol. 2016, 162, 493–499. [Google Scholar] [CrossRef]
- Waziri, P.M.; Abdullah, R.; Rosli, R.; Omar, A.R.; Abdul, A.B.; Kassim, N.K.; Malami, I.; Etti, I.C.; Sani, J.M.; Lila, M.A.M.; et al. Clausenidin Induces Caspase 8-Dependent Apoptosis and Suppresses Production of VEGF in Liver Cancer Cells. Asian Pac. J. Cancer Prev. 2018, 19, 917–922. [Google Scholar] [CrossRef]
- Al-Abboodi, A.S.; Al-Sheikh, W.M.; Eid, E.E.M.; Azam, F.; Al-Qubaisi, M.S. Inclusion complex of clausenidin with hydroxypropyl-β-cyclodextrin: Improved physicochemical properties and anti-colon cancer activity. Saudi Pharm. J. 2021, 29, 223–235. [Google Scholar] [CrossRef]
- Barnard, D.L.; Xu, Z.-Q.; Stowell, V.D.; Yuan, H.; Smee, D.F.; Samy, R.; Sidwell, R.W.; Nielsen, M.K.; Sun, L.; Cao, H.; et al. Coumarins and pyranocoumarins, potential novel pharmacophores for inhibition of measles virus replication. Antivir. Chem. Chemother. 2002, 13, 39–59. [Google Scholar] [CrossRef]
- Taira, N.; Nugara, R.N.; Inafuku, M.; Takara, K.; Ogi, T.; Ichiba, T.; Iwasaki, H.; Okabe, T.; Oku, H. In vivo and in vitro anti-obesity activities of dihydropyranocoumarins derivatives from Peucedanum japonicum Thunb. J. Funct. Foods 2017, 29, 19–28. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Fedorov, I.A.; Kaschenko, N.I.; Chirikova, N.K.; Vennos, C. Khellactone Derivatives and Other Phenolics of Phlojodicarpus sibiricus (Apiaceae): HPLC-DAD-ESI-QQQ-MS/MS and HPLC-UV Profile, and Antiobesity Potential of Dihydrosamidin. Molecules 2019, 24, 2286. [Google Scholar] [CrossRef] [Green Version]
- Hossin, A.Y.; Inafuku, M.; Oku, H. Dihydropyranocoumarins Exerted Anti-Obesity Activity In Vivo and its Activity Was Enhanced by Nanoparticulation with Polylactic-Co-Glycolic Acid. Nutrients 2019, 11, 3053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauwald, H.W.; Brehm, O.; Odenthal, K.P. The involvement of a Ca2+ channel blocking mode of action in the pharmacology of Ammi visnaga fruits. Planta Med. 1994, 60, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Gulyaev, S.M. Zashhitnoe dejstvie Phlojodicarpus sibiricus pri ishemii golovnogo mozga u krys [Protective effect of Phlojodicarpus sibiricus on cerebral ischemia in rats]. Acta Biomed. Sci. 2009, 67, 172–174. [Google Scholar]
- Gulyaev, S.M. Protivotrevozhnoe dejstvie Phlojodicarpus sibiricus [Anti-anxiety effect of Phlojodicarpus sibiricus]. Acta Biomed. Sci. 2009, 67, 175. [Google Scholar]
- Urbanova, E.Z.; Guljaev, S.M.; Nikolaev, S.M.; Turtueva, T.A. Membranostabilizirujushhij jeffekt jekstrakta Phlojodicarpus sibiricus [Membrane stabilizing effect of Phlojodicarpus sibiricus extract]. Sib. Med. Zhurnal 2013, 123, 104–105. [Google Scholar]
- Urbanova, E.Z.; Guljaev, S.M.; Nikolaev, S.M.; Turtueva, T.A. Nejrofarmakologicheskie jeffekty Phlojodicarpus sibiricus (Steph. ex Spreng.) K.-Pol. [Neuropharmacological effects of Phlojodicarpus sibiricus (Steph. ex Spreng.) K.-Pol.]. Vestn. Buryatskogo Gos. Univ. 2013, 12, 125–128. [Google Scholar]
- Xiong, Y.; Wang, J.; Wu, F.; Li, J.; Zhou, L.; Jong, L. Effects of (±)-praeruptorin A on airway inflammation, airway hyperresponsiveness and NF-κB signaling pathway in a mouse model of allergic airway disease. Eur. J. Pharmacol. 2012, 683, 316–324. [Google Scholar] [CrossRef]
- Xia, H.; Li, C.; Yang, J.; Ma, J.; Li, Y.; Li, L.; Zhang, D. Hepatoprotective pyranocoumarins from the stems of Clausena emarginata. Phytochemistry 2016, 130, 238–243. [Google Scholar] [CrossRef]
- Widelski, J.; Luca, S.V.; Skiba, A.; Maciąg, M.; Budzyńska, B.; Marcourt, L.; Wolfender, J.-L.; Skalicka-Woźniak, K. Coumarins from Seseli devenyense Simonk.: Isolation by Liquid–Liquid Chromatography and Potential Anxiolytic Activity Using an In Vivo Zebrafish Larvae Model. Int. J. Mol. Sci. 2021, 22, 41829. [Google Scholar] [CrossRef]
- Onder, A.; Cinar, A.S.; Baran, M.Y.; Kuruüzüm-Uz, A.; Trendafilova, A. Coumarins from Seseli petraeum M. Bieb. (Apiaceae) and their α-glucosidase inhibitory activity. S. Afr. J. Bot. 2022, 144, 458–463. [Google Scholar] [CrossRef]
- Ferrero-Miliani, L.; Seiden, J.B.; Nielsen, O.H. Regulation of cytokine production in inflammatory bowel disease. Ugeskr. Laeger 2006, 168, 1487–1850. [Google Scholar]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Morrison, P.W.J.; Connon, C.J.; Khutoryanskiy, V.V. Cyclodextrin-Mediated Enhancement of Riboflavin Solubility and Corneal Permeability. Mol. Pharm. 2013, 10, 756–762. [Google Scholar] [CrossRef]
- Hasani-Ranjbar, S.; Jouyandeh, Z.; Abdollahi, M. A systematic review of anti-obesity medicinal plants—An update. J. Diabetes Metab. Disord. 2013, 12, 28. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.Y.; Jannat, S.; Jung, H.; Choi, J. Insulin-Mimetic Dihydroxanthyletin-Type Coumarins from Angelica decursiva with Protein Tyrosine Phosphatase 1B and α-Glucosidase Inhibitory Activities and Docking Studies of Their Molecular Mechanisms. Molecules 2021, 10, 292. [Google Scholar] [CrossRef]
- Morikawa, T.; Luo, F.; Manse, Y.; Sugita, H.; Saeki, S.; Chaipech, S.; Pongpiriyadacha, Y.; Muraoka, O.; Nonomiya, K. Geranylated Coumarins from Thai Medicinal Plant Mammea siamensis with Testosterone 5α-Reductase Inhibitory Activity. Front. Chem. 2020, 8, 199. [Google Scholar] [CrossRef] [Green Version]
- Khandy, M.T.; Kochkin, D.V.; Tomilova, S.V.; Galishev, B.A.; Klyushin, A.G.; Nosov, M.K. Obtainment and phytochemical screening of callus and suspension cell cultures of Phlojodicarpus sibiricus (Steph. Ex Spreng.) K.-Pol. Biotechnology 2020, 36, 54–61. [Google Scholar] [CrossRef]
- Khandy, M.T.; Kochkin, D.V.; Tomilova, S.V.; Nosov, A.M.; Klyushin, A.G.; Galishev, B.A. Growth and Biosynthetic Characteristics of Phlojodicarpus sibiricus Cell Suspension Cultures. Russ. J. Plant. Physiol. 2021, 68, 569–578. [Google Scholar] [CrossRef]
- Murray, R.D.H. The Naturally Occurring Coumarins. In Progress in the Chemistry of Organic Natural Products; Herz, W., Falk, H., Kirby, G.W., Moore, R.E., Eds.; Springer: Vienna, Austria, 2002; Volume 83, pp. 1–619. [Google Scholar] [CrossRef]
- Wei, S.-P.; Luan, J.-Y.; Lu, L.-N.; Wu, W.-J.; Ji, Z.-Q. A new benzofuran glucoside from Ficus tikoua Bur. Int. J. Mol. Sci. 2011, 12, 84946. [Google Scholar] [CrossRef] [Green Version]
- Sarker, S.D.; Nahar, L. Progress in the Chemistry of Naturally Occurring Coumarins. In Progress in the Chemistry of Organic Natural Products; Kinghorn, A.D., Falk, H., Gibbons, S., Kobayashi, J.I., Eds.; Springer: Berlin, Germany, 2017; Volume 106, pp. 1–241. [Google Scholar] [CrossRef]
- He, R.; Zhang, Y.; Wu, L.; Nie, H.; Huang, Y.; Liu, B.; Deng, S.; Yang, R.; Huang, S.; Nong, Z.; et al. Benzofuran glycosides and coumarins from the bark of Streblus indicus (Bur.) Corner. Phytochemistry 2017, 138, 170–177. [Google Scholar] [CrossRef]
- Mi, J.; Peng, Y.; Zhang, H.; Wang, X.; Huo, Y.; Wang, Z.; Liu, Y.; Gao, Y.; Zhang, H. A new benzofuran derivative glycoside and a new coumarin glycoside from roots of Heracleum dissectum Ledeb. Med. Chem. Res. 2018, 27, 470–475. [Google Scholar] [CrossRef]
- Matsuo, Y.; Yamaguchi, E.; Hakamata, R.; Ootomo, K.; Takatori, K.; Fukaya, H.; Mimaki, Y. Benzofuran and coumarin derivatives from the root of Angelica dahurica and their PPAR-γ ligand-binding activity. Phytochemistry 2020, 173, e112301. [Google Scholar] [CrossRef] [PubMed]
- Nosov, A.M.; Popova, E.V.; Kochkin, D.V. Isoprenoid Production via Plant Cell Cultures: Biosynthesis, Accumulation and Scaling-Up to Bioreactors. In Production of Biomass and Bioactive Compounds Using Bioreactor Technology; Paek, K.Y., Murthy, H., Zhong, J.J., Eds.; Springer: Berlin, Germany, 2014; pp. 563–623. [Google Scholar] [CrossRef]
- Khandy, M.T.; Kochkin, D.V.; Tomilova, S.V.; Galishev, B.A.; Sukhanova, E.S.; Klyushin, A.G.; Ivanov, I.M.; Nosov, A.M. Obtaining and study of callus and suspension plant cell cultures of Tribulus terrestris L., a producer of steroidal glycosides. Appl. Biochem. Microbiol. 2017, 53, 800–806. [Google Scholar] [CrossRef]
- Xu, H.; Kim, Y.; Suh, S.; Uddin, M.R.; Lee, S.; Park, S. Decursin Production from Hairy Root Culture of Angelica gigas. J. Korean Soc. App. Biolo. Chem. 2008, 51, 349–351. [Google Scholar] [CrossRef]
- Kaul, B.; Staba, E.J. Ammi visnaga (L.) Lam. tissue cultures. Planta Med. 1967, 15, 145–156. [Google Scholar] [CrossRef] [PubMed]
- El-Fiky, F.K.; Remmel, R.P.; Staba, E.J. Ammi visnaga: Somatic Embryo and Furanochromone Production in Embryos, Seedlings, and Plants. Planta Med. 1989, 55, 446–451. [Google Scholar] [CrossRef]
- Krolicka, A.; Staniszewska, I.; Malinski, E.; Lojkowska, E. Stimulation of furanochromone accumulation in callus cultures of Ammi visnaga L. by addition of elicitors. Pharm. Int. J. Pharm. Sci. 2019, 58, 590–592. [Google Scholar]
- Al-Saleh, M.M.; Shibli, R.A.; Al-Qadiri, H.M.; Tahtamouni, R.W.; Darwish, M.M.; Al-Qudah, T.S. Investigating the Antimicrobial Potential of in-vitro Grown Microshoots and Callus Cultures of Ammi visnaga (L.) Lam. Jordan J. Biol. Sci. 2019, 12, 43–48. [Google Scholar]
- Ekiert, H.; Gomolka, E. Coumarin compounds in Ammi majus L. callus cultures. Pharmazie 2000, 55, 684–687. [Google Scholar]
- Krolicka, A.; Staniszewska, I.; Bielawski, K.; Malinski, E.; Safranek, J.; Lojkowska, E. Establishment of hairy root cultures of Ammi majus. Plant Sci. 2001, 160, 259–264. [Google Scholar] [CrossRef]
- Krolicka, A.; Lojkowska, E.; Staniszewska, I.; Malinski, E.; Safranek, J. Identification of secondary metabolites in in vitro culture of Ammi majus treated with elicitors. Acta Hortic. 2001, 560, 255–258. [Google Scholar] [CrossRef]
- Staniszewska, I.; Krolicka, A.; Lojkowska, E.; Malinski, E.; Safranek, J. Elicitation od secondary metabolites in in vitro cultures of Ammi majus L. Enzym. Microb. Technol. 2003, 33, 565–568. [Google Scholar] [CrossRef]
- Krolicka, A.; Kartanowicz, R.; Wosinski, S.A.; Spitter, A.; Kaminski, M.; Lojkowska, E. Induction of secondary metabolite production in transformed callus of Ammi majus L. grown after electromagnetic treatment of the culture medium. Enzym. Microb. Technol. 2006, 39, 1386–1391. [Google Scholar] [CrossRef]
- Magdum, S.; Kumar, S. Organogenesis of Ammi majus via Callus Culture from leaf. Plant Tissue Cult. Biotechnol. 2013, 23, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Eeva, M.; Ojala, T.; Tammela, P.; Galambosi, B.; Vuorela, H.; Hiltunen, R.; Fagerstedt, K.; Vuorela, P. Propagation of Angelica archangelica Plants in an Air-Spaged Bioreactor from a Novel Embryogenic Cell Line, and their Production of Coumarins. Biol. Plant. 2003, 46, 343–347. [Google Scholar] [CrossRef]
- Eeva, M. Plant secondary metabolites in Peucedanum palustre and Angelica archangelica and their plant cell cultures. Ph.D. Thesis, University of Helsinki, Helsinki, Findland, 2010; p. 77. [Google Scholar]
- Ahn, K.; Sim, W.; Kim, H.; Kim, I. Immunostimulating polysaccharide from cell culture of Angelica gigas Nakai. Biotechnol. Lett. 1998, 20, 5–7. [Google Scholar] [CrossRef]
- Lee, S.; Cho, W.; Lee, S.; Park, K.; Choi, K.; Hang, H. Effects of Plant Growth Regulators in In Vitro Germination and Organ Formation of Wild Angelica gigas N. Korean J. Plant Res. 2012, 25, 456–464. [Google Scholar] [CrossRef] [Green Version]
- Tsay, H.S.; Huang, H.L. Somatic embryo formation and germination from immature embryo-derived suspension-cultured cells of Angelica sinensis (Oliv.) Diels. Plant Cell Rep. 1998, 17, 670–674. [Google Scholar] [CrossRef]
- Chen, C.; Agrawal, D.C.; Lee, M.; Lee, R.; Kuo, C.; Wu, C.; Tsay, S. Influence of LED light spectra on in vitro somatic embryogenesis and LC-MC analysis of chlorogenic and rutin Peucedanum japonicum Thumb.: A medicinal herb. Bot. Stud. 2016, 57, 9. [Google Scholar] [CrossRef]
- Mitrofanova, I.V.; Mitrofanova, O.V.; Rudenko, M.I.; Lesnikova-Sedoshenko, N.P.; Kuzmina, T.N. In vitro morphogenesis in endangered plant Seseli lehmanii degen. In I International Symposium on Botanical Gardens and Landscapes 1298; Acta Horticulturae: Leuven, Belgium, 2020; pp. 257–264. [Google Scholar] [CrossRef]
- Hazeena, M.S.; Sulekha, G.R. Callus induction and plantlet regeneration in Aegle marmelos (L.) Corr. using cotyledon explants. J. Trop. Agric. 2008, 46, 79–84. [Google Scholar]
- Mishra, M.R.; Srivastava, R.K.; Akhtar, N. Enhanced Alkaloid Production from Cell Culture System of Catharanthus reseus in Combined effect of Nutrient Salts, Sucrose and Plant Growth Regulators. J. Biotechnol. Biomed. Sci. 2018, 1, 14–34. [Google Scholar] [CrossRef] [Green Version]
- Cappelletti, E.M.; Innocenti, G.; Caniato, R.; Filippini, R.; Povan, A. Haplophyllum patavinum (L.) G. Don fil. (Paduan rue): In Vitro Regeneration, and the Production of Coumarin Compounds. Biotechnol. Agric. Foresty 1998, 41, 238–260. [Google Scholar] [CrossRef]
- Hussain, M.A.; Nathar, V.N.; Mir, J.I. Gas chromatography-mass spectrometry (GC-MS) analysis in callus extracts of Ruta graveolens L. World J. Pharm. Res. 2017, 6, 1195–1210. [Google Scholar] [CrossRef]
- Rout, G.R.; Samantaray, S.; Das, P. In vitro rooting of Psoralea corylifilia Linn: Peroxidase activity as a marker. Plant Growth regul. 2000, 30, 215–219. [Google Scholar] [CrossRef]
- Nosov, A.M. Ispol’zovanie kletochnyh tehnologij dlja promyshlennogo poluchenija biologicheski aktivnyh veshhestv rastitel’nogo proishozhdenija [The use of cellular technologies for the industrial production of biologically active substances of plant origin]. Biotehnologija 2010, 5, 8–28. [Google Scholar]
- Paek, K.-Y.; Murthy, H.N.; Zhong, J.-J. (Eds.) Production of Biomass and Bioactive Compounds Using Bioreactor Technology; Springer: Dordrecht, The Netherlands, 2014; p. 709. [Google Scholar]
- Smanski, M.J.; Zhou, H.; Claesen, J.; Shen, B.; Fischbach, M.A.; Voigt, C.A. Synthetic biology to access and expand nature’s chemical diversity. Nat. Rev. Microbiol. 2016, 14, 135–149. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.; Hwang, S.; Kim, W.; Lee, Y.; Kim, J.H.; Cho, S.; Kim, H.U.; Yoon, Y.J.; Oh, M.K.; Palsson, B.O.; et al. Systems and synthetic biology to elucidate secondary metabolite biosynthetic gene clusters encoded in Streptomyces genomes. Nat. Prod. Rep. 2021, 38, 1330–1361. [Google Scholar] [CrossRef]
- Leonard, E.; Ajikumar, P.K.; Thayer, K.; Xiao, W.H.; Mo, J.D.; Tidor, B.; Stephanopoulos, G.; Prather, K.L.J. Combining Metabolic and Protein Engineering of a Terpenoid Biosynthetic Pathway for Overproduction and Selectivity Control. Proc. Natl. Acad. Sci. USA 2010, 107, 13654–13659. [Google Scholar] [CrossRef] [Green Version]
- Ro, D.K.; Paradise, E.M.; Ouellet, M.; Fisher, K.J.; Newman, K.L.; Ndungu, J.M.; Ho, K.A.; Eachus, R.A.; Ham, T.S.; Kirby, J.; et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature 2006, 440, 940–943. [Google Scholar] [CrossRef]
- Park, D.; Yang, D.; Ha, D.; Lee, S. Biosynthesis of dihydroquercetin in Escherichia coli from glycerol. bioRxiv 2020. [Google Scholar] [CrossRef]
- Luo, X.Z.; Reiter, M.A.; d’Espaux, L.; Wong, J.; Denby, C.M.; Lechner, A.; Zhang, Y.F.; Grzybowski, A.T.; Harth, S.; Lin, W.Y.; et al. Complete Biosynthesis of Cannabinoids and Their Unnatural Analogues in yeast. Nature 2019, 567, 123–126. [Google Scholar] [CrossRef]
- Ajikumar, P.K.; Xiao, W.H.; Tyo, K.E.J.; Wang, Y.; Simeon, F.; Leonard, E.; Mucha, O.; Phon, T.H.; Pfeifer, B.; Stephanopoulos, G. Isoprenoid Pathway Optimization for Taxol Precursor Overproduction in Escherichia coli. Science 2010, 330, 70–74. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.R.; Smolke, C.D. Engineering Biosynthesis of The Anticancer Alkaloid Noscapine in yeast. Nat. Commun. 2016, 7, 12137. [Google Scholar] [CrossRef] [Green Version]
- Galanie, S.; Thodey, K.; Trenchard, I.J.; Filsinger Interrante, M.; Smolke, C.D. Complete Biosynthesis of Opioids in yeast. Science 2015, 349, 1095–1100. [Google Scholar] [CrossRef] [Green Version]
- Bu, X.-L.; He, B.-B.; Weng, J.-Y.; Jiang, C.-C.; Zhao, Y.-L.; Li, S.-M.; Xu, J.; Xu, M.-J. Constructing Microbial Hosts for the Production of Benzoheterocyclic Derivatives. ACS Synth. Biol. 2020, 9, 2282–2290. [Google Scholar] [CrossRef]
- Malikov, V.M.; Saidkhodzhaev, A.I. Coumarins. Plants, structure, properties. Chem. Nat. Compd. 1998, 34, 345–409. [Google Scholar] [CrossRef]
- Sonnenberg, H.; Kaloga, M.; Eisenbach, N.; Froemming, K.K. Isolation and characterization of an angular-type dihydropyranocoumarine glycoside from the fruits of Ammi visnaga (L.) Lam. (Apiaceae). Z. Fur Nat. 1995, 50, 729–731. [Google Scholar] [CrossRef]
- Zgorka, G.; Dragan, T.; Glowniak, K.; Basiura, E. Determination of furanochromones and pyranocoumarins in drugs and Ammi visnaga fruits by combined solid-phase extraction-high-performance liquid chromatography and thin-layer chromatography-high-performance liquid chromatography. J. Chromatogr. A 1998, 797, 305–309. [Google Scholar] [CrossRef]
- Sellami, H.K.; Napolitano, A.; Masullo, M.; Smiti, S.; Piacente, S.; Pizza, C. Influence of growing conditions on metabolite profile of Ammi visnaga umbels with special reference to bioactive furanochromones and pyranocoumarins. Phytochemistry 2013, 95, 197–206. [Google Scholar] [CrossRef]
- Winderl, B.; Schwaiger, S.; Ganzera, M. Fast and improved separation of major coumarins in Ammi visnaga (L.) Lam. by supercritical fluid chromatography. J. Sep. Sci. 2016, 39, 4042–4048. [Google Scholar] [CrossRef]
- Ahmed, S.; Fahim, J.; Abdelmohsen, U. Chemical and biological potential of Ammi visnaga (L.) Lam. and Apium graveolens L.: A review (1963–2020). J. Adv. Biomed. Pharm. Sci. 2021, 4, 160–176. [Google Scholar] [CrossRef]
- Imanly, G.A.; Serkerov, S.V. Chemical components of roots and aerial parts of the Ammi visnaga (L.) lam. Khimija Rastit. Syr’ja 2017, 3, 237–240. [Google Scholar] [CrossRef] [Green Version]
- Avramenko, L.G.; Nikonov, G.K.; Pimenov, M.G. Andelin-A new dihydropyranocoumarin from the roots of Angelica decursiva. Chem. Nat. Compd. 1970, 6, 186–189. [Google Scholar] [CrossRef]
- Ali, M.Y.; Seong, S.H.; Jannat, S.; Jung, H.A.; Choi, J.S. Ethnobotany, Phytochemistry, and Pharmacology of Angelica decursiva Fr. et Sav. Nat. Prod. Sci. 2019, 25, 1135168. [Google Scholar] [CrossRef] [Green Version]
- Jung, D.J.; Porzel, A.; Huneck, S. Gigasol and other coumarins from Angelica gigas. Phytochemistry 1991, 30, 710–712. [Google Scholar] [CrossRef]
- Kang, S.Y.; Lee, K.Y.; Sung, S.H.; Park, M.J.; Kim, Y.C. Coumarins isolated from Angelica gigas inhibit acetylcholinesterase: Structure-activity relationships. J. Nat. Prod. 2001, 64, 683–685. [Google Scholar] [CrossRef]
- Park, Y.; Jeong, D.; Sim, S.; Kim, N.; Park, H.; Jeon, G. The Characteristics of Growth and Active Compounds of Angelica gigas Nakai Population in Mt. Jeombong. Korean J. Plant Res. 2019, 32, 9–18. [Google Scholar] [CrossRef]
- Geidarov, I.; Serkerov, S. Coumarins from roots of Angelica purpurascens. Chem. Nat. Compd. 2017, 53, 149–150. [Google Scholar] [CrossRef]
- Erdelmeier, C.A.J.; Sticher, O. Coumarin Derivatives from Eryngium campestre1. Planta Med. 1985, 51, 407–409. [Google Scholar] [CrossRef]
- Ahmadi, F.; Valadbeigi, S.; Sajjadi, S.E.; Shokoohinia, Y.; Azizian, H.; Taheripak, G. Grandivittin as a natural minor groove binder extracted from Ferulago macrocarpa to ct-DNA, experimental and in silico analysis. Chem. Biol. Interact. 2016, 258, 89–101. [Google Scholar] [CrossRef]
- Lee, J.W.; Lee, C.; Jin, Q.; Yeon, E.T.; Lee, D.; Kim, S.-Y.; Han, S.B.; Hong, J.T.; Lee, M.K.; Hwang, B.Y. Pyranocoumarins from Glehnia littoralis inhibit the LPS-induced NO production in macrophage RAW 264.7 cells. Bioorganic Med. Chem. Lett. 2014, 24, 2717–2719. [Google Scholar] [CrossRef]
- Swagen, T.M.; Cardellina, J.H. Coumarins from Musineon divaricatum☆. Phytochemistry 1985, 24, 805–813. [Google Scholar] [CrossRef]
- Sarkhail, P. Traditional uses, phytochemistry and pharmacological properties of the genus Peucedanum: A review. J. Ethnopharmacol. 2014, 156, 235–270. [Google Scholar] [CrossRef]
- Liu, R.; Sun, Q.; Shi, Y.; Kong, L. Isolation and purification of coumarin compounds from the root of Peucedanum decursivum (Miq.) Maxim by high-speed counter-current chromatography. J. Chromatogr. A 2005, 1076, 127–132. [Google Scholar] [CrossRef]
- Duh, C.-Y.; Wang, S.-K.; Wu, Y.-C. Cytotoxic pyranocoumarins from the aerial parts of Peucedanum japonicum. Phytochemistry 1991, 30, 2812–2814. [Google Scholar] [CrossRef]
- Ikeshiro, Y.; Mase, I.; Tomita, Y. Dihydropyranocoumarins from roots of Peucedanum japonicum. Phytochemistry 1992, 31, 4303–4306. [Google Scholar] [CrossRef]
- Ikeshiro, Y.; Mase, I.; Tomita, Y. Dihydropyranocoumarins from Peucedanum japonicum. Phytochemistry 1993, 33, 1543–1545. [Google Scholar] [CrossRef]
- Lu, M.; Nicoletti, M.; Battinelli, L.; Mazzanti, G. Isolation of praeruptorins A and B from Peucedanum praeruptorum Dunn. and their general pharmacological evaluation in comparison with extracts of the drug. Il Farmaco 2001, 56, 417–420. [Google Scholar] [CrossRef]
- Song, Y.; Jing, W.; Yan, R.; Wang, Y. Research progress of the studies on the roots of Peucedanum praeruptorum Dunn (Peucedani radix). Pak. J. Pharm. Sci. 2015, 28, 71–81. [Google Scholar]
- Kong, L.-Y.; Wu, X.; Niwa, M. Two Dihydropyranocoumarins from Peucedanum wawrii. Heterocycles 2003, 60, 599–606. [Google Scholar] [CrossRef]
- Babilev, F.V.; Nikonov, G.K. Coumarins of the roots of Phloiodicarpus villosus Turcz. Chem. Nat. Compd. 1965, 1, 383–386. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Chirikova, N.K. New compounds from flowers of Phlojodicarpus sibiricus. Chem. Nat. Compd. 2020, 56, 628–632. [Google Scholar] [CrossRef]
- Valencia-Islas, N.; Abbas, H.; Bye, R.; Toscano, R.; Mata, R. Phytotoxic Compounds from Prionosciadium watsoni. Nat. Prod. 2002, 65, 828–834. [Google Scholar] [CrossRef] [PubMed]
- Shagova, M.E.; Florya, V.N.; Kuznetsova, G.A.; Perel’son, M.E. Diesters of khellactone and rutarin (campesenin) from Seseli campestre growing in Moldavia. Chem. Nat. Compd. 1973, 9, 665. [Google Scholar] [CrossRef]
- Abyshev, A.Z.; Sidorova, I.P.; Abyshev, D.Z.; Florya, V.I.; Zmeikov, V.P.; Kerimov Yu, B. Comparative characterization of the coumarin composition ofSeseli campestregrowing in Moldavia and in the Caucasus. Chem. Nat. Compd. 1982, 18, 400–407. [Google Scholar] [CrossRef]
- Malikov, V.M.; Saidkhodzhaev, A.I. Coumarins: Plants, structures, properties. Chem. Nat. Compd. 1998, 34, 517–548. [Google Scholar] [CrossRef]
- Dukhovlinova, L.I.; Sklyar, Y.E.; Pimenov, M.G. Coumarins of the roots of Seseli coronatum. Chem. Nat. Compd. 1974, 10, 802–803. [Google Scholar] [CrossRef]
- Bagirov, V.; Yu; Belyi, M.B. An investigation of Seseli peucedanoides. Chem. Nat. Compd. 1981, 17, 585–586. [Google Scholar] [CrossRef]
- Razavi, S.M.; Imanzadeh, G.; Jahed, F.S.; Zarrini, G. Pyranocoumarins from Zosima absinthifolia (Vent) link roots. Russ. J. Bioorg. Chem. 2013, 39, 215–217. [Google Scholar] [CrossRef]
- Sklyar, Y.E.; Avramenko, L.G.; Pimenov, M.G.; Avetisyan, R.N. Coumarins of Zosima korovinii. Chem. Nat. Compd. 1982, 18, 744. [Google Scholar] [CrossRef] [Green Version]
- Gray, A.I.; Waterman, P.G. Coumarins in the Rutaceae. Phytochemistry 1978, 17, 845–864. [Google Scholar] [CrossRef]
- Panthong, K.; Srisud, Y.; Rukachaisirikul, V.; Hutadilok-Towatana, N.; Voravuthikunchai, S.P.; Tewtrakul, S. Benzene, coumarin and quinolinone derivatives from roots of Citrus hystrix. Phytochemistry 2013, 88, 79–84. [Google Scholar] [CrossRef]
- Ribeiro, A.B.; Abdelnur, P.V.; Garcia, C.F.; Belini, A.; Severino, V.G.P.; da Silva, M.F.G.F.; Fernandes, J.B.; Vieira, P.C.; de Souza, A.A.; Machado, M.A. Chemical Characterization of Citrus sinensis Grafted on C. limonia and the Effect of Some Isolated Compounds on the Growth of Xylella fastidiosa. J. Argic. Food Chem. 2008, 56, 7815–7822. [Google Scholar] [CrossRef]
- Waziri, P.M.; Abdullah, R.; Yeap, S.K.; Omar, A.R.; Abdul, A.B.; Kassim, N.K.; Malami, I.; Karunakaran, T.; Imam, M.U. Clausenidin from Clausena excavata induces apoptosis in hepG2 cells via the mitochondrial pathway. J. Ethnopharmacol. 2016, 194, 549–558. [Google Scholar] [CrossRef]
- Joshi, B.S.; Kamat, V.N.; Saksena, A.K. Structures of clausenin and clausenidin two new pyranocoumarins from the roots of clausena heptaphylla Wt. & Arn. Tetrahedron 1967, 23, 4785–4789. [Google Scholar] [CrossRef]
- Mohammadhosseini, M.; Venditti, A.; Frezza, C.; Serafini, M.; Bianco, A.; Mahdavi, B. The Genus Haplophyllum Juss.: Phytochemistry and Bioactivities—A Review. Molecules 2021, 26, 4664. [Google Scholar] [CrossRef]
- Ulubelen, A.; Mericli, A.H.; Mericli, F.; Sonmez, U.; Ilarslan, R. Alkaloids and Coumarins from Haplophyllum Thesioides. Nat. Prod. Lett. 1993, 1, 269–272. [Google Scholar] [CrossRef]
- Nikonov, G.K.; Manaeva, Z.A.; Pek, G.Y. Ostruthol from Xanthogallum purpurascens. Khimiya Prir. Soedin. 1966, 2, 360–361. [Google Scholar] [CrossRef]
- Chang, Y.; Bai, M.; He, H.; Huang, S.-W.; Song, S.-J. Phytochemical investigation on the fruits of Camptotheca acuminata and their chemotaxonomic significance. Biochem. Syst. Ecol. 2020, 93, 104169. [Google Scholar] [CrossRef]
- Erst, A.S.; Chernonosov, A.A.; Petrova, N.V.; Kulikovskiy, M.S.; Maltseva, S.Y.; Wang, W.; Kostikova, V.A. Investigation of Chemical Constituents of Eranthis longistipitata (Ranunculaceae): Coumarins and Furochromones. Int. J. Mol. Sci. 2022, 23, 10406. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Activity | Pure Substance/Extract | Pyranocoumarins Form | Model | Mechanism of Action | References |
|---|---|---|---|---|---|
| The anti-inflammatory activity | Decursin from Angelica gigas | L * | In vitro in macrophages | Decursin suppressed the expression of proteins matrix metalloproteinase 9, monocytic chemoattractant protein 1, interleukin 8, tumor necrosis factor α and IL-1β due to the fact that it inhibited the activity of the NF-kB translocation signaling pathway. | [23] |
| Praeruptorin A from Peucedanum praeruptorum | Ad * | In vitro in murine macrophage of RAW264.7 cell line | Pyranocoumarin suppressed the cytoplasmic loss of inhibitor κB-α protein and inhibited the translocation of NF-κB from cytoplasm to nucleus. | [24] | |
| (+)-Praeruptorin A from P. praeruptorum roots | Ad | In vitro in murine macrophage of RAW264.7 cell line | The presence of (+)-praeruptorin A in medium reduced nitric oxide production in cells. | [25] | |
| Corymbocoumarin from Seseli gummiferum subsp. corymbosum | Ad | In vitro in murine macrophage of RAW264.7 cell line | The presence of corimbocoumarin in medium suppressed the nuclear factor κB (NF-κB) signaling pathway activation and heme oxygenase (HO)-1 expression induction in cells. | [26] | |
| 10-(3,7-Dimethylocta-1,6-dien-3-yl)-5-methoxy-8,8- dimethylpyranocoumarin, nordentain from Clausena emarginata | L | In vitro in murine microglial of BV2 cell line | The presence of pyranocoumarin in medium reduced the nitric oxide production in cells. | [27] | |
| (+)-Praeruptorin A, visnadin from Ligusticum lucidum subsp. cuneifolium | Ad | In vivo in mice | (+)-Praeruptorin A reduced local edema by 22%, visnadin, by 43%. The anti-inflammatory activity of these compounds appears to be modulated by the substituents on their aromatic ring. | [28] | |
| Artificially synthesized 3,4-angular pyranocoumarins | A * | Against proteinase enzymes | Proteinase activity inhibition. The best effect was shown by pyranocoumarins with amino groups in the pyran ring | [29] | |
| In vivo in rats | Rat paw edema decreased to 29.2%. | ||||
| Against COX-1 and COX-2 | Most of the pyranocoumarin derivatives showed themselves against COX-2 (IC50 ≤ 10 µM). | ||||
| Antioxidant activity | Grandivittin, agasyllin, aegelinol from Ferulago campestris | L | In vitro in human polymorphonuclear neutrophils (PMN) respiratory burst cells and on human whole blood leukocytes (WB) | It is suggested that pyranocoumarins scavenge reactive oxygen species or interfere with cellular activation mechanisms. | [30] |
| Artificially synthesized 3′,4′-Di-O-acetyl-cis-khellactone (DOAcK) | Ad | In vivo in rats | DOAcK administration increased the activities of Catalase, Glutathione Peroxidase and Super Oxide Dismutase. | [31] | |
| Antimicrobial activity | Decursin and decursinol angelate from Angelica gigas | L | In vitro in Bacillus subtilis | The six-membered ring and senecioylic acid type side chain closely related to the enhanced antibacterial activities of coumarins against B. subtilis. Although a precise mechanism has not yet been clarified, these phenomena might be considered to occur due to the differences in the binding affinities of both compounds on the active sites of the enzymes or receptors from the differences in the position of the side chain moiety. | [32] |
| Agasyllin, aegelinol from Ferulago campestris | L | In vitro in bacteria | At a concentration of 16 to 125 µg/mL, the growth of Staphylococcus aureus, Salmonella thypii, Enterobacter cloacae and Enterobacter earogenes was suppressed, from 5 to 25 µg/mL—Helicobacter pylori. | [33] | |
| Pd-D-V and Disenecioyl Khellactone from P. decursivum | L | In vitro against S. sclerotiorum, B. cinerea, F. graminearum and C. capsici | Pd-DV at a concentration of 30 μg/mL in medium inhibited the growth of S. sclerotiorum by 86.4 ± 0.7, and also inhibited the growth of B. cinerea, F. graminearum and C. capsici by almost 50%. | [34] | |
| Ad | In vitro against S. sclerotiorum, T. cucumeris, B. cinerea and F. graminearum | Disenecioyl Khellactone at a concentration of 30 μg/mL inhibited the growth of S. sclerotiorum and B. cinerea by almost 50%, and the growth of F. framinearum, T. cucumeris and C. capsica by almost 30%. | |||
| Agasyllin, aegelinol from Ferulago campestris | L | Magnaporthe oryzae in vitro and in vivo | Pyranocoumarins inhibited M. oryzae spore germination and upressoria at concentrations of 50 and 200 µg/mL and prevented rice disease by more than 80% at 100 and 300 µg/mL. | [22] | |
| Phenyl derivative of pyranocoumarin (PDP) from Psoralea corylifolia L. | L | In vitro in Fusarium oxysporum, F. graminearum, and F. moniliforme | The ligand PDP showed bifurcated hydrogen bond interaction with active site residues at TYR 413 and a single hydrogen bond interaction at ARG 402 with a docking score −7.19 and glide energy of −45.78 kcal/mol. This indicated a strong binding of the ligand with the trichothecene 3-O-acetyltransferase, preventing as a result the acetylation of the trichothecene mycotoxin and destruction of the “self-defense mechanism” of the Fusarium sp. | [21] | |
| Artificially synthesized 3,4-angular pyranocoumarins | A | In vitro in bacteria and fungi | Most pyranocoumarins exhibited inhibitory activity in zones with diameters in the range of 15–19 mm or more. It is assumed that activity is associated with lipophilicity of molecules. | [29] | |
| Anticancer activity | Decursin and decursinol from A. gigas | L | In vivo in mice | Tumor size decreased by 40.6% and 45.6% with introduction of decursin and decursinol angelate | [35] |
| In vitro in prostate cancer DU145 and LNCaP cells | Decursin inhibited cell growth by stopping the G1, G2 and S phases at different dosages (25–100 μM) | [36] | |||
| In vitro in LNCaP cells | Decursin inhibits androgen-stimulated nuclear translocation of the androgen receptor and reduces the amount of androgen receptor protein. | [37] | |||
| In vitro in MCF-7 cells | Decursin and decursinol angelate exerted growth inhibitory effects on MCF-7 cells through G1 arrest and caspase-mediated apoptosis. | [38] | |||
| In vitro in prostate cancer PC-3 cells | Decursin suppressed cell proliferation by suppressing Wnt/β-catenin pathway. | [39] | |||
| In vitro in RC cells | Decursin inhibits cell proliferation by inducing apoptosis, which is mediated by both caspase-dependent and caspase-independent apoptosis pathways. | [40] | |||
| In vitro in U266, MM.1S, ARH77 cells | Decursin showed a synergistic effect with bortezomib due to inhibition of STAT3 (activation of signal transducers and the activator of transcription 3)-induced proliferative and angiogenic effect in multiple myeloma. | [41] | |||
| In vitro in HeLa cells | Extract increases expression of TRAIL, which stimulates extrinsic and intrinsic pathways of apoptosis through the activation of caspase-8 and caspase-9, respectively. | [42] | |||
| (+)-Decursinol from Saposhnikovia divaricata | L | In vitro in MEL-8, U-937, DU-145, MDA-MB-231 and BT-474 cell lines | It inhibited growth and proliferation of DU-154 prostate cancer cells and MEL-8 melanoma cells. The mechanism has not been considered. | [43] | |
| Clausarin from Clausena excavata | L | In vitro in the multi-drug resistant cell line KB-VIN cells | The mechanism(s) of action should be further investigated. | [44] | |
| Artificially synthesized linear pyranocoumarins | L | ||||
| (±)-Praeruptorin A & (±)-Praeruptorin B from Peucedanum praeruptorum | Ad | In vitro in SGC7901 cells | (±)-Praeruptorin A increases cancer cells drug sensitivity, presumably by inhibiting the expression of P-glycoprotein, which is responsible for the efflux of drugs from cancer cells through ATP-dependent pumps. | [45] | |
| Angular pyranocoumarins from Peucedanum praeruptorum roots | Ad | In vitro in MES-SA/Dx5 | Pyranocoumarins inhibited drug efflux via MDR protein, which prevents multidrug-resistant cancer formation. | [25] | |
| Grandivitin from Ferulago macropara | L | Molecular modeling analysis against MMP9 | By binding to Matrix metalloproteinase 9 (MMP9), it affected secondary structure and modified tertiary structure of this protein. | [46] | |
| Clausenidin from Clausena excavata | L | In vitro in HepG2 cells | Clausenidin increased the activity of caspase-8 and expression of protein components of the death inducing signaling complex (DISC) in HepG2 cells. | [47] | |
| L | In vitro in HT29 and CCD-18Co | HT29 colorectal adenocarcinoma cells treated with clausenidin/hydroxypropyl-β-cyclodextrin complex showed cell cycle arrest and death by apoptosis associated with caspase activation. | [48] | ||
| Antiviral activity | Causenidin and nordentatin from C. excavata | L | In vitro in HepA2 cells | The pyranocoumarins suppressed hepatitis. | [44] |
| Artificially synthesized linear pyranocoumarins | L | B virus surface antigen in HepA2 cells and had anti-HBV values. Their EC50 values were 1.14, 1.34, 1.64 and 1.63 µM. | |||
| Artificially synthesized 5,6-angular pyranocoumarins | A | African green monkey (CV-1) kidney cell lines infected with measles virus | The compounds inhibited nine strains of measles virus, and in virucidal tests, drugs did not physically destroy virion to inhibit virus replication. | [49] | |
| Antihyperglycemic and antidyslipidemic activity | Pyranocoumarins isolated from P. japonicum | Ad | In vitro on 3T3-L1 adipocytes | Inhibited lipid accumulation and lipogenic gene expression in 3T3-L1 adipocytes. | [50] |
| Root and herb extracts of P. sibiricus, separately isolated fractions of dihydrosamidine, kelactone esters | Ad | In vitro in 3T3-L1 cells | Root extracts and esters of kellacton inhibited triacylglycerol accumulation, while activity depended on acyl groups type. | [51] | |
| Dihydropyranocoumarins isolated from P. japonicum | Ad | In vivo in mice | Consumption of dihydropyranocumarins significantly reduced average size of adipocytes and increased mRNA levels of genes associated with thermogenesis. Nanoparticulation of DPCs with polylactic-co-glycolic acid (PLGA) dramatically increased its activity almost 100-fold over that of a non-nanoparticulated form. | [52] | |
| (+)-trans-decursidinol, Pd-C-I and Pd-C-II, Pd-C-III isolated from Angelica decursiva | L | In vitro in HepG2 cells | Pyranocoumarins increased glucose uptake and decreased PTP1B expression in insulin resistant HepG2 cells; inhibited albumin nitration mediated by ONOO-, removed peroxynitrite, ROS. | [30] | |
| Other activities/effects | Visnadin from Ammi visnaga | Ad | In vitro in aortic segments of guinea-pigs | Visnadin predominantly inhibits contractile responses mediated by penetration of Ca2+ through Ca2+. | [53] |
| Tincture from Phlojodicarpus sibiricus | Ad | In vivo in rats | Tincture showed anxiolytic properties, presumably due to GABAA benzodiazepine receptors activation. | [54] | |
| Tincture from P. sibiricus | Ad | In vivo in rats | The extract limits the development of emotional and neurological disorders in cerebral ischemia in rats. | [55] | |
| Extract of P. sibiricus | - | In vitro in erythrocites | Extract inhibited hemolysis of erythrocytes, which is presumably due to the membrane-stabilizing properties of coumarins. | [56] | |
| Extract of P. sibiricus | - | In vivo in rats | Extract has a neuromodulatory effect, probably due to activation of GABA (gamma aminobutyric acid)-ergic system by coumarin compounds. | [57] | |
| (±)-Praeruptorin from P. praeruptorum | Ad | In vivo in mice | (±)-Praeruptorin A suppressed airway inflammation, airway hyperreactivity and remodeling, reduced serum IL-4 and IL-13 levels in BALF and IgE, inhibited TGF-β1 and pSmad2/3 expression, increased Smad7 expression in lung tissue, and also increased INF-γ levels in BALF. | [58] | |
| Clauemarmarin C, D & 5-hydroxy-8,8-dimethyl-10- (7-hydroxy-3,7-dimethylocta-1,5-dien-3-yl) pyranocoumarin from C. emarginata | L | In vitro in WB-F344 cells | They showed a hepatoprotective effect, the mechanism was not considered. | [59] | |
| Mammeasins E and F from Mammea siamensis | C * | Against enzyme testosterone 5α-reductase | Inhibited testosterone 5α-reductase. | [21] | |
| cis-Khellactone, d-laserpitin, isolaserpitin and octanoyllomatin, isolated from Seseli devenyense | Ad | In vivo on zebrafish larvae | Coumarins reduced anxiety behavior (anxiolytic activity) in zebrafish larvae. The mechanism was not considered. | [60] | |
| Angular pyranocoumarins from Seseli petraeum | Ad | Against enzymes α-amylase and α-glucosidase | The coumarins exhibited notable inhibitory activity against the α-glucosidase enzyme and low inhibitory potential against α-amylase. | [61] |
| Species | Explant | Growth Conditions, Media | Results | Pyranocoumarins | References |
|---|---|---|---|---|---|
| Umbelliferae (Apiaceae) | |||||
| Ammi visnaga (L.) Lam | Fruits | MS * with 1.0 mg/L 2,4-D * | Callus tissue contained 45 mg visnagin/1100 g dry weight. | Not detected | [79] |
| Fruits |
|
| Not detected | [80] | |
| Hypocotyl of sterile seedlings | MS with 2.5 mg/L NAA * and 1.0 mg/L BA | Callus tissue with visnagin and/or khellin. | Not detected | [81] | |
| Seeds | MS with 1.0 mg/L BA and 2.0 mg/L 2,4-D | Microshoots in vitro and callus culture. | Not detected | [82] | |
| Ammi majus L. | Not specified | Linsmaier-Skoog’s media with NAA and BA | Tissue and organ cultures containing different concentrations of the linear furanocoumarins psoralen, bergapten, xanthotoxin, isopimpinellin, imperatorin and their precursor umbelliferone. | Not detected | [83] |
| Plantlets | MS with 2.0 mg/L NAA, 2.0 mg/L BA | Callus, suspension and hairy roots of A. majus containing umbelliferone were obtained. | Not detected | [84] | |
| Plantlets [84] | Not specified | The effect of various elicitors on the synthesis of coumarins (umbelliferone, marmesinin, scopoletin) in hairy roots of A. majus was shown. | Not detected | [85] | |
| Plantlets [84] | MS with various phytohormons | The effect of various elicitors on the synthesis of secondary metabolites (scopoletin, dehydrogeijerin) of in callus, cell suspension and hairy roots of A. majus by exposing them to elicitors. | Not detected | [86] | |
| Hairy root culture | MS with 2.5 mg/L NAA, 1.0 mg/L BA | Elicitation with ADR-4® (electromagnetic treatment) induced also two times higher accumulation of bergapten. | Not detected | [87] | |
| Leafs | MS with 2 mg/L IAA *, 2 mg/L Kin * | Callus of A. majus were obtained. About 81% of calluses converted to shoot on medium with 50 mg/L glutamine and 40 mg/L adenine. Plantlets with shoot were transferred to 1/2 MS with different concentration of IBA and glutamine. Maximum rooting of 75.96% was observed on 1/2 MS supplemented with 0.2% of IBA and 100 mg/L glutamine. | Not detected | [88] | |
| Angelica archangelica (L.) subsp. archangelica | Embryogenic cell line from seeds | Hormone-free, modified B5 * | Embryogenic cell line of A. archangelica was obtained, after 5 years it did not decrease capacity of embryo formation. | Not detected | [89] |
| Plantlets from embryogenic cell line | - | 14 coumarins were identified in the roots of A. archangelica, oxypeucedanin hydrate and oxypeucedanin being the main compounds. | Not detected | [90] | |
| Angelica gigas Nakai | Young plants | Schenk and Hildebrandt media with 2.0 mg/L 2,4-D, 1.0 mg/L Kin | An immunostimulating polysaccharide was produced extracellularly by suspension cell culture of A. gigas. | Not detected | [91] |
| Leaves and stems | Hormone-free MS | Hairy roots were induced from leaf and stem explants of A. gigas. Biomass growth and decursin production was faster in hairy roots than in wild type of A. gigas. | Decursin | [78] | |
| Stems, roots and hypocotyls In vitro | MS with NAA and 2,4-D, BA, GA * and TDZ * | The highest callus induction rate was obtained from in vitro germinated stem, root and hypocotyl on the MS medium with 1.0 mg/L NAA and 0.5 mg/L BA. | Not detected | [92] | |
| Angelica sinensis (Oliv.) Diets. | Immature embryos | MS, B5, White | Study showed that embryogenic callus growth was more rapid on MS basal medium than on B5 or White medium. Suspension culture and somatic embryos were obtained from this callus. | Not detected | [93] |
| Peucedanum japonicum Thunb. | Root, leaf blade and petiole parts of seedlings In vitro | MS with 0.1–5.0 mg/L 2,4-D and 0.1–5.0 mg/L ABA * | Embryogenesis was induced in media MS with ABA from callus subculture (MS with 2,4-D). Four-month-old tissue culture plants derived from somatic embryos showed significantly of chlorogenic acid (10.5 mg/g dw). | Not detected | [94] |
| Phlojodicarpus sibiricus (Steph. e x Spreng.) K.-Pol. | Seeds, leaves, roots and hypocotyls In vitro | MS with c 1.0 mg/L 2,4-D and 0.5 mg/L BA | Callus was obtained from seeds, leaves, roots and hypocotyls in vitro. Dihydrosamidin, visnadin, khellactone derivatives were identified in callus cultures. | Dihydrosamidin, visnadin, khellactone derivatives | [68] |
| Callus | MS with c 1.0 mg/L 2,4-D and 0.5 mg/L BA | Suspension culture was obtained from callus. | Not detected | [69] | |
| Seseli lehmannii Degen | Callus and somatic embryos | MS with 1.0–2.0 mg/L BA and 0.5–2.0 mg/L NAA | Organogenesis of callus was induced in MS media with 1.0–2.0 mg/L BAP and 0.5–2.0 mg/L NAA. | Not detected | [95] |
| Rutaceae | |||||
| Aegle marmelos L. | Leaves | MS with 2.26 μM 2,4-D and 2.2 μM benzyl adenine | Callus was obtained from leaves on MS with 2,4-D and BA. Derived shoots from callus were rooted in vitro on MS medium supplemented with 12.3 μM indole-3-butyric acid. | Not detected | [96] |
| Catharanthus roseus Linn. | Leaves, roots, axillary buds, shoot tips | MS and B5 with various concentrations of 2,4-D, Kin, NAA | Callus culture was obtained. There was no significant difference in growing parameters between cultures in both types of media formulations on agar (MS or B5 salts). However, the alkaloid content was 2–3 times higher in suspension culture compared to agar medium in similar treatments. | Not detected | [97] |
| Haplophyllum patavinum (L.) G. Don fil. | Plantlets | B5 with 3 mg/L IAA and MS with 3 mg/L IAA | H. patavinum is not rich in coumarin compounds in vivo, but a selected cell strain exhibited in vitro coumarin biogenetic potentialities stronger than in vivo. | Not detected | [98] |
| Ruta graveolens L. | Whole plant | MS with 1.5 mg/L 2,4-D and 1.5 mg/L NAA | MS with 2, 4-D (1.5 mg/L) and NAA (1.5 mg/L) responded well by giving the maximum percentage of callus induction (97.22 ± 2.54). Coumarins were detected by GC-MS in biomass. | Not detected | [99] |
| Fabaceae | |||||
| Psoralea corylifolia L. (Buguchi) | Leaves and stems | MS with IAA | Rooting was induced in microshoots of P. corylifolia. Peroxidase activity increased considerably during root induction indicating a key role of peroxidase in rooting of P. corylifolia microshoots in vitro. | Not detected | [100] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khandy, M.T.; Sofronova, A.K.; Gorpenchenko, T.Y.; Chirikova, N.K. Plant Pyranocoumarins: Description, Biosynthesis, Application. Plants 2022, 11, 3135. https://doi.org/10.3390/plants11223135
Khandy MT, Sofronova AK, Gorpenchenko TY, Chirikova NK. Plant Pyranocoumarins: Description, Biosynthesis, Application. Plants. 2022; 11(22):3135. https://doi.org/10.3390/plants11223135
Chicago/Turabian StyleKhandy, Maria T., Anastasia K. Sofronova, Tatiana Y. Gorpenchenko, and Nadezhda K. Chirikova. 2022. "Plant Pyranocoumarins: Description, Biosynthesis, Application" Plants 11, no. 22: 3135. https://doi.org/10.3390/plants11223135
APA StyleKhandy, M. T., Sofronova, A. K., Gorpenchenko, T. Y., & Chirikova, N. K. (2022). Plant Pyranocoumarins: Description, Biosynthesis, Application. Plants, 11(22), 3135. https://doi.org/10.3390/plants11223135