
Genome-Wide Analysis of Cyclic Nucleotide-Gated Channel Genes Related to Pollen Development in Rice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Motifs Analysis
2.2. Meta-Expression Analysis
2.3. Multiple Sequence Alignment and Phylogenetic Tree Construction
2.4. Cis-Acting Element Analysis
2.5. RNA Extraction, cDNA Construction, and qRT-PCR Analysis
2.6. Subcellular Localization Analysis
2.7. Yeast Two-Hybrid Analysis
2.8. Network Analysis
3. Results
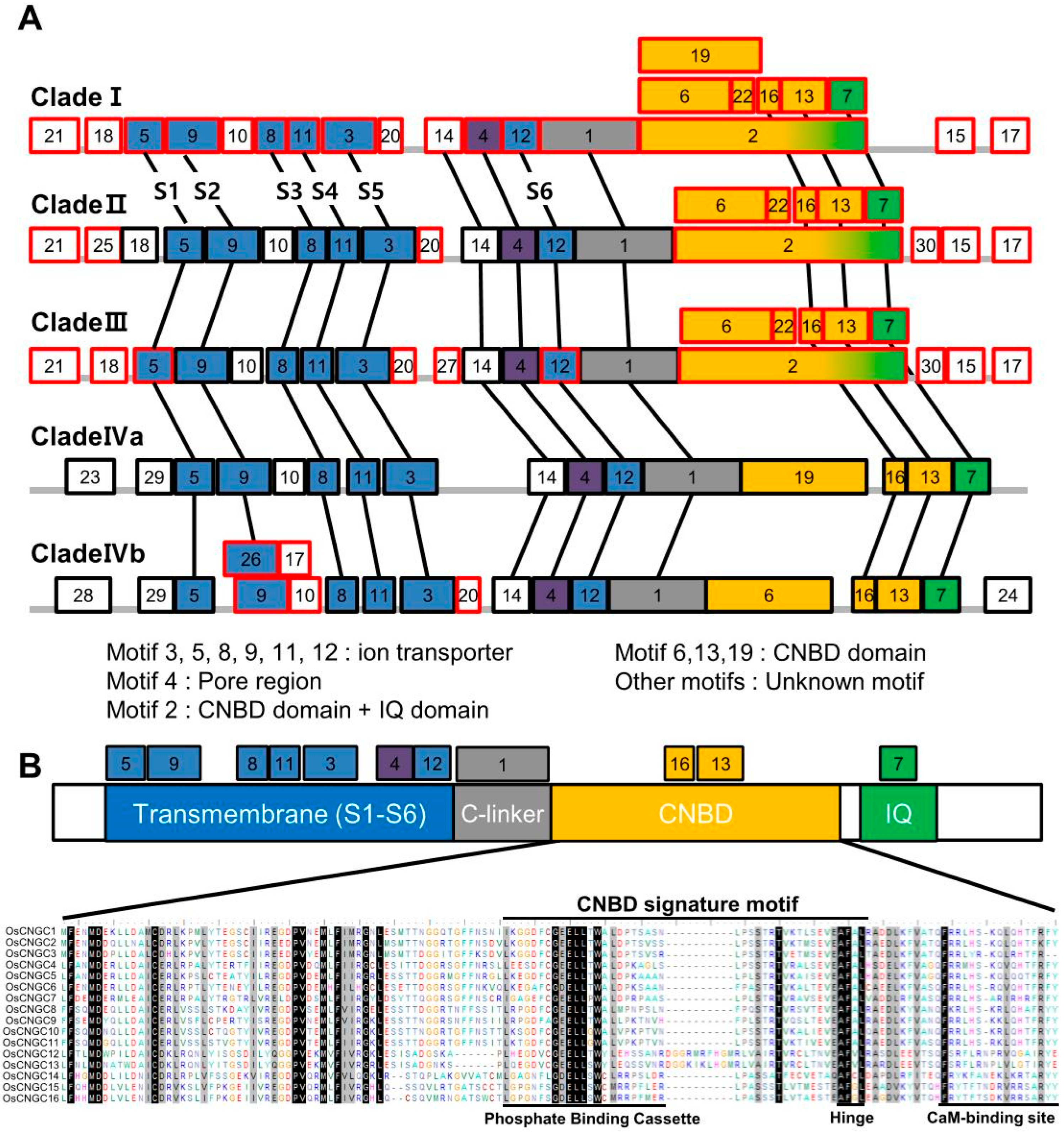
3.1. Phylogenetic Tree and Motif Composition Analyses of OsCNGC Proteins
3.2. Expression Pattern Analysis of OsCNGC Genes
3.3. Cis-Acting Regulatory Element Analysis for Promoter Regions of OsCNGC Family Genes
3.4. Subcellular Localization of OsCNGC4, 5, and 8 Proteins
3.5. Network Analysis of CNGC-Mediated Genes for Pollen Development
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stiess, M.; Bradke, F. Neuronal Polarization: The Cytoskeleton Leads the Way. Dev. Neurobiol. 2011, 71, 430–444. [Google Scholar] [CrossRef]
- Hepler, P.K.; Vidali, L.; Cheung, A.Y. Polarized Cell Growth in Higher Plants. Annu. Rev. Cell Dev. Biol. 2001, 17, 159–187. [Google Scholar] [CrossRef]
- Steinhorst, L.; Kudla, J. Calcium—A Central Regulator of Pollen Germination and Tube Growth. Biochim. Biophys. Acta 2013, 1833, 1573–1581. [Google Scholar] [CrossRef] [Green Version]
- Zheng, R.H.; Su, S.D.; Xiao, H.; Tian, H.Q. Calcium: A Critical Factor in Pollen Germination and Tube Elongation. Int. J. Mol. Sci. 2019, 20, 420. [Google Scholar] [CrossRef] [Green Version]
- Malhó, R.; Read, N.D.; Trewavas, A.J.; Pais, M.S. Calcium Channel Activity during Pollen Tube Growth and Reorientation. Plant Cell 1995, 7, 1173–1184. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-F.; Fan, L.-M.; Zhang, W.-Z.; Zhang, W.; Wu, W.-H. Ca2+-Permeable Channels in the Plasma Membrane of Arabidopsis Pollen Are Regulated by Actin Microfilaments. Plant Physiol. 2004, 136, 3892–3904. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, Z.; Kakar, K.U.; Saand, M.A.; Shu, Q.-Y. Cyclic Nucleotide-Gated Ion Channel Gene Family in Rice, Identification, Characterization and Experimental Analysis of Expression Response to Plant Hormones, Biotic and Abiotic Stresses. BMC Genom. 2014, 15, 853. [Google Scholar] [CrossRef] [Green Version]
- Talke, I.; Blaudez, D.; Maathuis, F.; Sanders, D. CNGCs: Prime Targets of Plant Cyclic Nucleotide Signalling? Trends Plant Sci. 2003, 8, 286–293. [Google Scholar] [CrossRef]
- Ma, W.; Berkowitz, G.A. Ca2+ Conduction by Plant Cyclic Nucleotide Gated Channels and Associated Signaling Components in Pathogen Defense Signal Transduction Cascades. New Phytol. 2011, 190, 566–572. [Google Scholar] [CrossRef]
- Frietsch, S.; Wang, Y.-F.; Sladek, C.; Poulsen, L.R.; Romanowsky, S.M.; Schroeder, J.I.; Harper, J.F. A Cyclic Nucleotide-Gated Channel Is Essential for Polarized Tip Growth of Pollen. Proc. Natl. Acad. Sci. USA 2007, 104, 14531–14536. [Google Scholar] [CrossRef]
- Gao, Q.-F.; Gu, L.-L.; Wang, H.-Q.; Fei, C.-F.; Fang, X.; Hussain, J.; Sun, S.-J.; Dong, J.-Y.; Liu, H.; Wang, Y.-F. Cyclic Nucleotide-Gated Channel 18 Is an Essential Ca2+ Channel in Pollen Tube Tips for Pollen Tube Guidance to Ovules in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 113, 3096–3101. [Google Scholar] [CrossRef] [Green Version]
- Jha, S.K.; Sharma, M.; Pandey, G.K. Role of Cyclic Nucleotide Gated Channels in Stress Management in Plants. Curr. Genom. 2016, 17, 315–329. [Google Scholar] [CrossRef]
- Dietrich, P.; Moeder, W.; Yoshioka, K. Plant Cyclic Nucleotide-Gated Channels: New Insights on Their Functions and Regulation. Plant Physiol. 2020, 184, 27–38. [Google Scholar] [CrossRef]
- Fischer, C.; Kugler, A.; Hoth, S.; Dietrich, P. An IQ Domain Mediates the Interaction with Calmodulin in a Plant Cyclic Nucleotide-Gated Channel. Plant Cell Physiol. 2013, 54, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Lan, W.; Jiang, Y.; Fang, W.; Luan, S. A Calcium-Dependent Protein Kinase Interacts with and Activates a Calcium Channel to Regulate Pollen Tube Growth. Mol. Plant 2014, 7, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Curran, A.; Chang, I.F.; Chang, C.L.; Garg, S.; Miguel, R.M.; Barron, Y.D.; Li, Y.; Romanowsky, S.; Cushman, J.C.; Gribskov, M.; et al. Calcium-Dependent Protein Kinases from Arabidopsis Show Substrate Specificity Differences in an Analysis of 103 Substrates. Front. Plant Sci. 2011, 2, 36. [Google Scholar] [CrossRef] [Green Version]
- Maathuis, F.J.M. CGMP Modulates Gene Transcription and Cation Transport in Arabidopsis Roots. Plant J. 2006, 45, 700–711. [Google Scholar] [CrossRef]
- Rubio, F.; Flores, P.; Navarro, J.; Martínez, V. Effects of Ca2+, K+ and CGMP on Na+ Uptake in Pepper Plants. Plant Sci. 2003, 165, 1043–1049. [Google Scholar] [CrossRef]
- Penson, S.P.; Schuurink, R.C.; Fath, A.; Gubler, F.; Jacobsen, J.V.; Jones, R.L. CGMP Is Required for Gibberellic Acid-Induced Gene Expression in Barley Aleurone. Plant Cell 1996, 8, 2325–2333. [Google Scholar] [CrossRef]
- Mäser, P.; Thomine, S.; Schroeder, J.I.; Ward, J.M.; Hirschi, K.; Sze, H.; Talke, I.N.; Amtmann, A.; Maathuis, F.J.; Sanders, D.; et al. Phylogenetic Relationships within Cation Transporter Families of Arabidopsis. Plant Physiol. 2001, 126, 1646–1667. [Google Scholar] [CrossRef]
- Guo, J.; Islam, M.A.; Lin, H.; Ji, C.; Duan, Y.; Liu, P.; Zeng, Q.; Day, B.; Kang, Z.; Guo, J. Genome-Wide Identification of Cyclic Nucleotide-Gated Ion Channel Gene Family in Wheat and Functional Analyses of TaCNGC14 and TaCNGC16. Front. Plant Sci. 2018, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- Hao, L.; Qiao, X. Genome-Wide Identification and Analysis of the CNGC Gene Family in Maize. PeerJ 2018, 6, e5816. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, Z.; Kakar, K.U.; Ullah, R.; Yu, S.; Zhang, J.; Shu, Q.-Y.; Ren, X. Genome-Wide Identification, Evolution and Expression Analysis of Cyclic Nucleotide-Gated Channels in Tobacco (Nicotiana Tabacum L.). Genomics 2019, 111, 142–158. [Google Scholar] [CrossRef]
- Saand, M.A.; Xu, Y.-P.; Li, W.; Wang, J.-P.; Cai, X.-Z. Cyclic Nucleotide Gated Channel Gene Family in Tomato: Genome-Wide Identification and Functional Analyses in Disease Resistance. Front. Plant Sci. 2015, 6, 303. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Yin, H.; Gu, J.; Li, L.; Liu, Z.; Jiang, X.; Zhou, H.; Wei, S.; Zhang, S.; Wu, J. Genomic Characterization, Phylogenetic Comparison and Differential Expression of the Cyclic Nucleotide-Gated Channels Gene Family in Pear (Pyrus Bretchneideri Rehd.). Genomics 2015, 105, 39–52. [Google Scholar] [CrossRef]
- Kakar, K.U.; Nawaz, Z.; Kakar, K.; Ali, E.; Almoneafy, A.A.; Ullah, R.; Ren, X.; Shu, Q.-Y. Comprehensive Genomic Analysis of the CNGC Gene Family in Brassica Oleracea: Novel Insights into Synteny, Structures, and Transcript Profiles. BMC Genom. 2017, 18, 869. [Google Scholar] [CrossRef] [Green Version]
- Kosuta, S.; Hazledine, S.; Sun, J.; Miwa, H.; Morris, R.J.; Downie, J.A.; Oldroyd, G.E.D. Differential and Chaotic Calcium Signatures in the Symbiosis Signaling Pathway of Legumes. Proc. Natl. Acad. Sci. USA 2008, 105, 9823–9828. [Google Scholar] [CrossRef] [Green Version]
- Tunc-Ozdemir, M.; Tang, C.; Ishka, M.R.; Brown, E.; Groves, N.R.; Myers, C.T.; Rato, C.; Poulsen, L.R.; McDowell, S.; Miller, G.; et al. A Cyclic Nucleotide-Gated Channel (CNGC16) in Pollen Is Critical for Stress Tolerance in Pollen Reproductive Development. Plant Physiol. 2013, 161, 1010–1020. [Google Scholar] [CrossRef] [Green Version]
- Tunc-Ozdemir, M.; Rato, C.; Brown, E.; Rogers, S.; Mooneyham, A.; Frietsch, S.; Myers, C.T.; Poulsen, L.R.; Malhó, R.; Harper, J.F. Cyclic Nucleotide Gated Channels 7 and 8 Are Essential for Male Reproductive Fertility. PLoS ONE 2013, 8, e55277. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.-G.; Liang, L.; Jia, P.-F.; Wang, Y.-C.; Li, H.-J.; Yang, W.-C. Integration of Ovular Signals and Exocytosis of a Ca2+ Channel by MLOs in Pollen Tube Guidance. Nat. Plants 2020, 6, 143–153. [Google Scholar] [CrossRef]
- Xu, Y.; Yang, J.; Wang, Y.; Wang, J.; Yu, Y.; Long, Y.; Wang, Y.; Zhang, H.; Ren, Y.; Chen, J.; et al. OsCNGC13 Promotes Seed-Setting Rate by Facilitating Pollen Tube Growth in Stylar Tissues. PLoS Genet. 2017, 13, e1006906. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.J.; Kim, Y.J.; Hong, W.J.; Lee, C.; Jeon, J.S.; Jung, K.H. Genome-Wide Analysis of Root Hair Preferred Rboh Genes Suggests That Three Rboh Genes Are Associated with Auxin-Mediated Root Hair Development in Rice. J. Plant Biol. 2019, 62, 229–238. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant Cis-Acting Regulatory DNA Elements (PLACE) Database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for Motif Discovery and Searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Moon, S.; Hong, W.-J.; Kim, Y.-J.; Chandran, A.K.N.; Gho, Y.-S.; Yoo, Y.-H.; Nguyen, V.N.T.; An, G.; Park, S.K.; Jung, K.-H. Comparative Transcriptome Analysis Reveals Gene Regulatory Mechanism of UDT1 on Anther Development. J. Plant Biol. 2020, 63, 289–296. [Google Scholar] [CrossRef]
- Vo, K.T.X.; Kim, C.-Y.; Chandran, A.K.N.; Jung, K.-H.; An, G.; Jeon, J.-S. Molecular Insights into the Function of Ankyrin Proteins in Plants. J. Plant Biol. 2015, 58, 271–284. [Google Scholar] [CrossRef]
- Jain, M.; Nijhawan, A.; Tyagi, A.K.; Khurana, J.P. Validation of Housekeeping Genes as Internal Control for Studying Gene Expression in Rice by Quantitative Real-Time PCR. Biochem. Biophys. Res. Commun. 2006, 345, 646–651. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of Housekeeping Genes for Gene Expression Studies in Human Reticulocytes Using Real-Time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- You, M.K.; Lim, S.-H.; Kim, M.-J.; Jeong, Y.S.; Lee, M.-G.; Ha, S.-H. Improvement of the Fluorescence Intensity during a Flow Cytometric Analysis for Rice Protoplasts by Localization of a Green Fluorescent Protein into Chloroplasts. Int. J. Mol. Sci. 2015, 16, 788–804. [Google Scholar] [CrossRef]
- Lee, S.K.; Hong, W.-J.; Silva, J.; Kim, E.-J.; Park, S.K.; Jung, K.-H.; Kim, Y.-J. Global Identification of ANTH Genes Involved in Rice Pollen Germination and Functional Characterization of a Key Member, OsANTH3. Front. Plant Sci. 2021, 12, 609473. [Google Scholar] [CrossRef]
- Diller, T.C.; Madhusudan; Xuong, N.H.; Taylor, S.S. Molecular Basis for Regulatory Subunit Diversity in CAMP-Dependent Protein Kinase: Crystal Structure of the Type II Beta Regulatory Subunit. Structure 2001, 9, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Zagotta, W.N.; Siegelbaum, S.A. Structure and Function of Cyclic Nucleotide-Gated Channels. Annu. Rev. Neurosci. 1996, 19, 235–263. [Google Scholar] [CrossRef]
- Baudier, J.; Deloulme, J.C.; Van Dorsselaer, A.; Black, D.; Matthes, H.W. Purification and Characterization of a Brain-Specific Protein Kinase C Substrate, Neurogranin (P17). Identification of a Consensus Amino Acid Sequence between Neurogranin and Neuromodulin (GAP43) That Corresponds to the Protein Kinase C Phosphorylation Site and the Calmodulin-Binding Domain. J. Biol. Chem. 1991, 266, 229–237. [Google Scholar]
- Chen, S.J.; Klann, E.; Gower, M.C.; Powell, C.M.; Sessoms, J.S.; Sweatt, J.D. Studies with Synthetic Peptide Substrates Derived from the Neuronal Protein Neurogranin Reveal Structural Determinants of Potency and Selectivity for Protein Kinase C. Biochemistry 1993, 32, 1032–1039. [Google Scholar] [CrossRef]
- Chandran, A.K.N.; Hong, W.-J.; Abhijith, B.; Lee, J.; Kim, Y.-J.; Park, S.K.; Jung, K.-H. Rice Male Gamete Expression Database (RMEDB): A Web Resource for Functional Genomic Studies of Rice Male Organ Development. J. Plant Biol. 2020, 63, 421–430. [Google Scholar] [CrossRef]
- Hong, W.-J.; Kim, Y.-J.; Kim, E.-J.; Kumar Nalini Chandran, A.; Moon, S.; Gho, Y.-S.; Yoou, M.-H.; Kim, S.T.; Jung, K.-H. CAFRI-Rice: CRISPR Applicable Functional Redundancy Inspector to Accelerate Functional Genomics in Rice. Plant J. 2020, 104, 532–545. [Google Scholar] [CrossRef]
- Manimaran, P.; Raghurami Reddy, M.; Bhaskar Rao, T.; Mangrauthia, S.K.; Sundaram, R.M.; Balachandran, S.M. Identification of Cis-Elements and Evaluation of Upstream Regulatory Region of a Rice Anther-Specific Gene, OSIPP3, Conferring Pollen-Specific Expression in Oryza sativa (L.) Ssp. Indica. Plant Reprod. 2015, 28, 133–142. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Kim, M.-H.; Hong, W.-J.; Moon, S.; Kim, E.-J.; Silva, J.; Lee, J.; Lee, S.; Kim, S.T.; Park, S.K.; et al. GORI, Encoding the WD40 Domain Protein, Is Required for Pollen Tube Germination and Elongation in Rice. Plant J. 2021, 105, 1645–1664. [Google Scholar] [CrossRef]
- Liu, L.; Zheng, C.; Kuang, B.; Wei, L.; Yan, L.; Wang, T. Receptor-Like Kinase RUPO Interacts with Potassium Transporters to Regulate Pollen Tube Growth and Integrity in Rice. PLoS Genet. 2016, 12, e1006085. [Google Scholar] [CrossRef] [Green Version]
- Duszyn, M.; Świeżawska, B.; Szmidt-Jaworska, A.; Jaworski, K. Cyclic Nucleotide Gated Channels (CNGCs) in Plant Signalling-Current Knowledge and Perspectives. J. Plant Physiol. 2019, 241, 153035. [Google Scholar] [CrossRef]
- Chang, F.; Yan, A.; Zhao, L.-N.; Wu, W.-H.; Yang, Z. A Putative Calcium-Permeable Cyclic Nucleotide-Gated Channel, CNGC18, Regulates Polarized Pollen Tube Growth. J. Integr. Plant Biol. 2007, 49, 1261–1270. [Google Scholar] [CrossRef]
- Bolte, S.; Talbot, C.; Boutte, Y.; Catrice, O.; Read, N.D.; Satiat-Jeunemaitre, B. FM-Dyes as Experimental Probes for Dissecting Vesicle Trafficking in Living Plant Cells. J. Microsc. 2004, 214, 159–173. [Google Scholar] [CrossRef]
- Wang, Q.; Kong, L.; Hao, H.; Wang, X.; Lin, J.; Šamaj, J.; Baluška, F. Effects of Brefeldin A on Pollen Germination and Tube Growth. Antagonistic Effects on Endocytosis and Secretion. Plant Physiol. 2005, 139, 1692–1703. [Google Scholar] [CrossRef] [Green Version]
- Weitz, D.; Ficek, N.; Kremmer, E.; Bauer, P.J.; Kaupp, U.B. Subunit Stoichiometry of the CNG Channel of Rod Photoreceptors. Neuron 2002, 36, 881–889. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Chai, X.; Gao, Q.; Zhou, L.; Zhang, S.; Li, L.; Luan, S. Dynamic Interactions of Plant CNGC Subunits and Calmodulins Drive Oscillatory Ca2+ Channel Activities. Dev. Cell 2019, 48, 710–725.e5. [Google Scholar] [CrossRef] [Green Version]
- Duan, Q.; Liu, M.-C.J.; Kita, D.; Jordan, S.S.; Yeh, F.-L.J.; Yvon, R.; Carpenter, H.; Federico, A.N.; Garcia-Valencia, L.E.; Eyles, S.J.; et al. FERONIA Controls Pectin-and Nitric Oxide-Mediated Male–Female Interaction. Nature 2020, 579, 561–566. [Google Scholar] [CrossRef]
- Escobar-Restrepo, J.-M.; Huck, N.; Kessler, S.; Gagliardini, V.; Gheyselinck, J.; Yang, W.-C.; Grossniklaus, U. The FERONIA Receptor-like Kinase Mediates Male-Female Interactions during Pollen Tube Reception. Science 2007, 317, 656–660. [Google Scholar] [CrossRef]
- Bowler, C.; Neuhaus, G.; Yamagata, H.; Chua, N.-H. Cyclic GMP and Calcium Mediate Phytochrome Phototransduction. Cell 1994, 77, 73–81. [Google Scholar] [CrossRef]
- Mu, Y.; Zou, M.; Sun, X.; He, B.; Xu, X.; Liu, Y.; Zhang, L.; Chi, W. BASIC PENTACYSTEINE Proteins Repress ABSCISIC ACID INSENSITIVE4 Expression via Direct Recruitment of the Polycomb-Repressive Complex 2 in Arabidopsis Root Development. Plant Cell Physiol. 2017, 58, 607–621. [Google Scholar] [CrossRef] [Green Version]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF Family Transcription Factors in Plant Abiotic Stress Responses. Biochim. Et Biophys. Acta (BBA) Gene Regul. Mech. 2012, 1819, 86–96. [Google Scholar] [CrossRef]
- Moeder, W.; Yoshioka, K. CNGCs Break Through-A Rice Cyclic Nucleotide-Gated Channel Paves the Way for Pollen Tube Growth. PLoS Genet. 2017, 13, e1007066. [Google Scholar] [CrossRef] [Green Version]
- Biel, M.; Michalakis, S. Cyclic Nucleotide-Gated Channels. In cGMP: Generators, Effectors and Therapeutic Implications; Schmidt, H.H.H.W., Hofmann, F., Stasch, J.-P., Eds.; Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2009; pp. 111–136. ISBN 978-3-540-68964-5. [Google Scholar]
- Kaupp, U.B.; Seifert, R. Cyclic Nucleotide-Gated Ion Channels. Physiol. Rev. 2002, 82, 769–824. [Google Scholar] [CrossRef] [Green Version]
- Köhler, B.; Blatt, M.R. Protein Phosphorylation Activates the Guard Cell Ca2+ Channel and Is a Prerequisite for Gating by Abscisic Acid. Plant J. 2002, 32, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Stoelzle, S.; Kagawa, T.; Wada, M.; Hedrich, R.; Dietrich, P. Blue Light Activates Calcium-Permeable Channels in Arabidopsis Mesophyll Cells via the Phototropin Signaling Pathway. Proc. Natl. Acad. Sci. USA 2003, 100, 1456–1461. [Google Scholar] [CrossRef] [Green Version]
- Solis-Miranda, J.; Quinto, C. The CrRLK1L Subfamily: One of the Keys to Versatility in Plants. Plant Physiol. Biochem. 2021, 166, 88–102. [Google Scholar] [CrossRef]
- Nissen, K.S.; Willats, W.G.T.; Malinovsky, F.G. Understanding CrRLK1L Function: Cell Walls and Growth Control. Trends Plant Sci. 2016, 21, 516–527. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-K.; Lee, S.-M.; Kim, M.-H.; Park, S.-K.; Jung, K.-H. Genome-Wide Analysis of Cyclic Nucleotide-Gated Channel Genes Related to Pollen Development in Rice. Plants 2022, 11, 3145. https://doi.org/10.3390/plants11223145
Lee S-K, Lee S-M, Kim M-H, Park S-K, Jung K-H. Genome-Wide Analysis of Cyclic Nucleotide-Gated Channel Genes Related to Pollen Development in Rice. Plants. 2022; 11(22):3145. https://doi.org/10.3390/plants11223145
Chicago/Turabian StyleLee, Su-Kyoung, Soo-Min Lee, Myung-Hee Kim, Soon-Ki Park, and Ki-Hong Jung. 2022. "Genome-Wide Analysis of Cyclic Nucleotide-Gated Channel Genes Related to Pollen Development in Rice" Plants 11, no. 22: 3145. https://doi.org/10.3390/plants11223145
APA StyleLee, S. -K., Lee, S. -M., Kim, M. -H., Park, S. -K., & Jung, K. -H. (2022). Genome-Wide Analysis of Cyclic Nucleotide-Gated Channel Genes Related to Pollen Development in Rice. Plants, 11(22), 3145. https://doi.org/10.3390/plants11223145