
In Vitro Conservation through Slow Growth Storage Technique of Fruit Species: An Overview of the Last 10 Years





Abstract
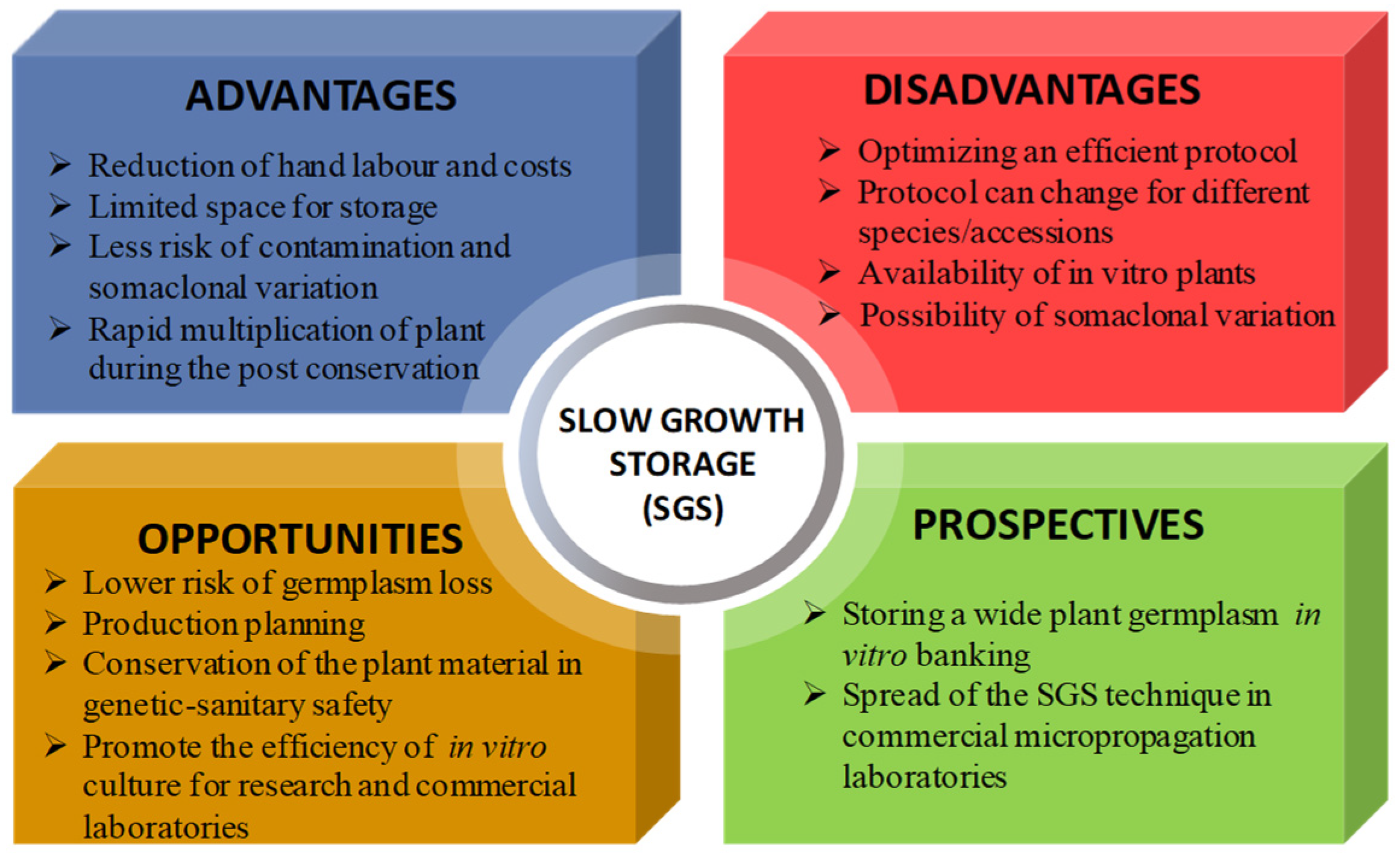
:1. Introduction
2. Factors Affecting SGS of Shoot Cultures of Temperate Fruit Crops
2.1. Temperature and Light Conditions
2.2. Storage Medium Composition
2.3. Containers for SGS
2.4. Genotype Effect
3. Conservation in SGS of Tropical Fruit Species

{kind=link}
{kind=link}
| Species | Medium | Temperature (°C) | Light Condition | Storage Time (months) | Survival (%) | References |
|---|---|---|---|---|---|---|
| Ananas comosus | ½MS, 30 g L−1 sucrose | 21 | 12 h, 20 μmol m−2 s−1 | 24 * | 100 | [70] |
| Carica pubescens | ½MS, 2 ppm BA | 8 | 16 h, 18 watts | 6 | 90 | [65] |
| Colocasia esculenta | MS, 4% mannitol | NR | 16 h | 24 | 80 | [67] |
| Hancornia speciosa | MS, 15 g L−1 sucrose, 5 g L−1 sorbitol | 25 | 16 h, 30 μmol m−2 s−1 | 4 | 95 | [66] |
| Musa spp. | MS, 30 g L−1 sucrose | 25 | 14 h, blue light (450–465 nm) 50 μmol m−2 s−1 | 3.5 | 100 | [72] |
| ½MS or MS, 2.25 mg L−1 BAP, 0.175 mg L−1 IAA, 30 g L−1 sucrose | 18 | NR | 5 | 100 | [76] | |
| MS, 4% mannitol | 23 | 12 h | 16 | NR | [55] | |
| Musa acuminata × balbisiana | MS, 2.25 mg L−1 BAP, 0.175 mg L−1 IAA, 3% sucrose, 2.5 and 5 ppm PBZ | 18-22 | Natural light | 6 | NR | [81] |
| Musa balbisiana | MS, 1% sucrose | 25 | 16 h, 20 μmol m−2 s−1 | 6 | 25 | [75] |
| Phoenix dactylifera | MS, 10 mg L−1 2,4-D, 3 mg L−1 2iP, 6 mg L−1 ABA, 102 g L−1 sucrose, 3 g L−1 AC | 15 | Darkness | 12 | 91.8 | [82] |
| MS | 5 | Darkness | 12 | 70 | [63] | |
| Simmondsia chinensis | MS, 1 mg L−1 BA | 5 | Darkness | 9 | NR | [64] |
| Vanilla planifolia | MS, 2 mg L−1 BA, 3 mg L−1 ABA | 24 | 16 h, 40–50 μmol m−2 s−1 | 6 | 90 | [68] |
| MS, 3 mg L−1 ABA | 22 | 16 h, 50 μmol m−2 s−1 | 4 | 93.3 | [69] |
4. Genetic Stability
5. In Vitro Banking Strategy
6. Conclusions and Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kulak, V.; Longboat, S.; Brunet, N.D.; Shukla, M.; Saxena, P. In Vitro technology in plant conservation: Relevance to biocultural diversity. Plants 2022, 11, 503. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, H. Classification of seed storage types for ex situ conservation in relation to temperature and moisture. In Ex Situ Plant Conservation: Supporting Species Survival in the Wild; Guerrant, E., Havens, K., Maunder, M., Eds.; Island Press: Washington, DC, USA, 2004; pp. 139–161. [Google Scholar]
- Shii, C.T.; Ma, S.S.; Huang, P.L.; Chin, S.W. Current and future development in clonal germplasm conservation of vegetatively propagated crops. In Proceedings of the Plant Germplasm Conservation: Perspectives for the 2000s, Taiwan, China, 10–11 August 1994. [Google Scholar]
- Reed, B.M. The in vitro genebank of temperate fruit and nut crops at the National Clonal Germplasm Repository-Corvallis. In Management of Field and In Vitro Germplasm Collections; Intl. Plant Genetic Resources Institute: Rome, Italy, 1999; pp. 132–135. [Google Scholar]
- Levy, L.; Damsteegt, V.; Welliver, R. First Report of Plum pox virus (Sharka Disease) in Prunus persica in the United States. Plant Dis. 2000, 84, 202. [Google Scholar] [CrossRef] [PubMed]
- García-Ramírez, Y.; Barrera, G.P.; Freire-Seijo, M.; Barbón, R.; Concepción-Hernández, M.; Mendoza-Rodríguez, M.F.; Torres-García, S. Effect of sucrose on physiological and biochemical changes of proliferated shoots of Bambusa vulgaris Schrad. Ex Wendl in temporary immersion. Plant Cell. Tissue Organ Cult. 2019, 137, 239–247. [Google Scholar] [CrossRef]
- Martelli, G.P.; Boscia, D.; Porcelli, F.; Saponari, M. The olive quick decline syndrome in south-east Italy: A threatening phytosanitary emergency. Eur. J. Plant Pathol. 2016, 144, 235–243. [Google Scholar] [CrossRef]
- Engelmann, F. In vitro conservation of tropical plant germplasm—A review. Euphytica 1991, 57, 227–243. [Google Scholar] [CrossRef]
- Lambardi, M.; Ozudogru, E.A. Advances in the safe storage of micropropagated woody plants at low temperature. Acta Hortic. 2013, 988, 29–42. [Google Scholar] [CrossRef]
- Grout, B.W.W. In vitro conservation of germplasm. In Developments in Crop Science; Elsevier: Amsterdam, The Netherlands, 1990; Volume 19, pp. 394–411. [Google Scholar]
- Engelmann, F. Management of field and in vitro germplasm collections. In Proceedings of the Consultation Meeting, Rome, Italy, 5–20 January 1996; IPGRI: Rome, Italy, 1999; p. 165. [Google Scholar]
- Cha-um, S.; Kirdmanee, C. Minimal growth in vitro culture for preservation of plant species. Fruit Veg. Cereal Sci. Biotechnol. 2007, 1, 13–25. [Google Scholar]
- Rajasekharan, P.E.; Sahijram, L. In Vitro conservation of plant germplasm. In Plant Biology and Biotechnology; Bahadur, B., Venkat Rajam, M., Sahijram, L., Krishnamurthy, K.V., Eds.; Springer: New Delhi, India, 2015; Volume II, pp. 417–443. ISBN 978-81-322-2282-8. [Google Scholar]
- Reed, B.M.; Gupta, S.; Uchendu, E.E. In vitro genebanks for preserving tropical biodiversity. In Conservation of Tropical Plant Species; Springer: New York, NY, USA, 2013; pp. 77–106. [Google Scholar]
- Mascarello, C.; Sacco, E.; Di Silvestro, D.; Pamato, M.; Ruffoni, B. Reduction of the subculture frequency in three Mediterranean species during micropropagation. Acta Hortic. 2017, 1155, 461–466. [Google Scholar] [CrossRef]
- Capuana, M.; Di Lonardo, S. In vitro conservation of chestnut (Castanea sativa) by slow growth. Vitr. Cell. Dev. Biol.-Plant 2013, 49, 605–610. [Google Scholar] [CrossRef]
- Carvalho, M.D.J.D.S.D.; Souza, A.D.S.; Santos, E.B.; Filho, W.D.S.S.; Ledo, C.A.D.S.; Costa, E.M.R.; Souza, F.V.D. In vitro conservation of ‘Florida Rough’ lemon plants. Ciência Rural 2022, 52, 1–9. [Google Scholar] [CrossRef]
- Arbeloa, A.; Marín, J.A.; Andreu, P.; García, E.; Lorente, P. In vitro conservation of fruit trees by slow growth storage. Acta Hortic. 2017, 1155, 101–106. [Google Scholar] [CrossRef]
- Sedlak, J.; Zidova, P.; Paprstein, F. Slow growth in vitro conservation of fruit crops. Acta Hortic. 2019, 1234, 119–124. [Google Scholar] [CrossRef]
- Turdiyev, T.T.; Kovalchuk, I.Y.; Kabylbekova, B.Z.; Chukanova, N.I.; Frolov, S.N. In vitro germplasm cold storage of fruit and berry plants of Kazakhstan. Eurasian J. Biosci. 2020, 1219, 1213–1219. [Google Scholar]
- Lukoševičiute, V.; Rugienius, R.; Staniene, G.; Blažyte, A.; Gelvonauskiene, D.; Bendokas, V.; Gelvonauskis, B.; Sasnauskas, A.; Stanys, V.; Bobinas, Č. Low temperature storage of Fragaria sp. and Pyrus sp. genetic resources in vitro. Zemdirbyste 2012, 99, 125–130. [Google Scholar]
- Monticelli, S.; Gentile, A.; Forni, C.; Caboni, E. Slow growth in “Pisana” apricot and in “Cerina” apple, two Italian cultivars. Acta Hortic. 2020, 1285, 117–124. [Google Scholar] [CrossRef]
- Kabylbekova, B.; Kovalchuk, I.; Chukanova, N.; Turdiyev, T. Cold storage of in vitro apple germplasm in Kazakhstan. Acta Hortic. 2021, 1324, 77–82. [Google Scholar] [CrossRef]
- Koç, I.; Akdemir, H.; Onay, A.; Çiftçi, Y.Ö. Cold-induced genetic instability in micropropagated Pistacia lentiscus L. plantlets. Acta Physiol. Plant. 2014, 36, 2373–2384. [Google Scholar] [CrossRef]
- Ozudogru, E.A.; Benelli, C.; Dradi, G.; Lambardi, M. Effect of culture container and carbohydrate content on in vitro slow growth storage of the cherry rootstock ‘Gisela®5. ’ Acta Physiol. Plant. 2017, 39, 2–9. [Google Scholar] [CrossRef]
- Sota, V.; Kongjika, E. The effect of nutrient media in micropropagation and in vitro conservation of wild population of mahaleb cherry (Prunus mahaleb L.). J. Microbiol. Biotech. Food Sci. 2014, 3, 453–456. [Google Scholar]
- Gianní, S.; Sottile, F. In vitro storage of plum germplasm by slow growth. Hortic. Sci. 2015, 42, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Sota, V.; Kongjika, E. Micropropagation and in vitro Conservation via Slow Growth Techniques of Prunus webbii (Spach) Vierh: An Endangered Plant Species in Albania. Int. J. Biol. Life Agric. Sci. 2019, 13, 280–283. [Google Scholar]
- Kovalchuk, I.; Zhumagulova, Z.; Turdiev, T.; Reed, B.M. Growth medium alterations improve in vitro cold storage of pear germplasm. Cryo-Letters 2014, 35, 197–203. [Google Scholar] [PubMed]
- Reed, B.M.; DeNoma, J.; Wada, S.; Postman, J. Micropropagation of pear (Pyrus sp.). In Protocols for Micropropagation of Selected Economically-Important Horticultural Plant; Humana Press: Totowa, NJ, USA, 2013; pp. 3–18. ISBN 978-1-4614-3775-8. [Google Scholar]
- Reed, B.M.; DeNoma, J. Medium-term in vitro storage of pear as a complementary germplasm preservation technique. Acta Hortic. 2016, 1113, 251–256. [Google Scholar] [CrossRef]
- Nin, S.; Benelli, C.; Petrucci, W.A.; Turchi, A.; Pecchioli, S.; Gori, M.; Giordani, E. In vitro propagation and conservation of wild bilberry (Vaccinium myrtillus L.) genotypes collected in the Tuscan Apennines (Italy). J. Berry Res. 2019, 9, 411–430. [Google Scholar] [CrossRef]
- Pan, X.J.; Zhang, W.E.; Li, X. In vitro conservation of native Chinese wild grape (Vitis heyneana Roem. & Schult) by slow growth culture. VITIS-J. Grapevine Res. 2014, 53, 207–214. [Google Scholar]
- AlMousa, R.; Hassan, N. Effect of osmotic regulators on In vitro conservation of grape. J. Genet. Environ. Resour. Conserv. 2022, 10, 60–66. [Google Scholar]
- Hassanen, S.A.; Abido, A.I.A.; Aly, M.A.M.; Rayan, G.A. In vitro preservation of grapevine (Vitis vinifera L.) Muscat of Alexandria and Black Monukka cultivars as genetic resource. Afr. J. Basic Appl. Sci. 2013, 5, 55–63. [Google Scholar] [CrossRef]
- Sota, V.; Kongjika, E. Slow growth in vitro conservation of Zizyphus jujuba Mill. Poljopr. I Sumar. 2014, 60, 27–37. [Google Scholar]
- Benson, E.E.; Harding, K.; Debouck, D.; Dumet, D.; Escobar, R.; Mafla, G.; Panis, B.; Panta, A.; Tay, D.; Van Den, I.; et al. Refinement and Standardization of Storage Procedures for Clonal Crops; Global Public Goods Phase 2. Part 3: Multi-crop guidelines for developing in vitro conservation best practices for clonal crops; System-Wide Genetic Resources Programme (SGRP): Rome, Italy, 2011; p. 144. ISBN 9789290439059. [Google Scholar]
- Lambardi, M.; De Carlo, A. Application of tissue culture to the germplasm conservation of temperate broad-leaf trees. In Micropropagation of Woody Trees and Fruits; Springer: Dordrecht, The Netherlands, 2003; pp. 815–840. [Google Scholar]
- Lambardi, M.; Roncasaglia, R.; Previati, A.; De Carlo, A.; Dradi, G.; Da Re, F.; Calamai, L. In vitro slow growth storage of fruit rootstocks inside gas-tight or gas-permeable containers. Acta Hortic. 2006, 725, 483–488. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Lloyd, G.; McCown, B. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot tip culture. Int. Plant Propagators’ Soc. 1980, 30, 421–427. [Google Scholar]
- Driver, J.A.; Kunijuki, A. In vitro propagation of paradox walnut rootstock. Hortscience 1984, 19, 507–550. [Google Scholar] [CrossRef]
- Rugini, E. In vitro propagation of some olive (Olea europaea sativa L.) cultivars with different root-ability, and medium development using analytical data from developing shoots and embryos. Sci. Hortic. 1984, 24, 123–134. [Google Scholar] [CrossRef]
- Lambardi, M.; Benelli, C.; De Carlo, A.; Fabbri, A.; Grassi, S.; Lynch, P.T. Medium- and long- term in vitro conservation of olive germplasm (Olea europaea L.). Acta Hortic. 2002, 586, 109–112. [Google Scholar] [CrossRef]
- Gardi, T.; Micheli, M.; Piccioni, E.; Sisani, G.; Standardi, A. Cold storage of micropropagated shoots of some fruit species. Italus Hortus 2001, 8, 32–40. [Google Scholar]
- Reski, R.; Abel, W.O. Induction of budding on chloronemata and caulonemata of the moss, Physcomitrella patens, using isopentenyladenine. Planta 1985, 165, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Anjum, M.A.; Ahmed, M.J.; Sajid, G.M.; Yoqub, A.; Shafqat, M. Role of plant growth regulators in preservation of Pyrus germplasm in vitro. Afr. J. Biotechnol. 2011, 10, 14029–14037. [Google Scholar] [CrossRef] [Green Version]
- Selvakumar, G.; Bindu, G.H.; Bhatt, R.M.; Upreti, K.K.; Paul, A.M.; Asha, A.; Shweta, K.; Sharma, M. Osmotolerant cytokinin producing microbes enhance tomato growth in deficit irrigation conditions. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2018, 88, 459–465. [Google Scholar] [CrossRef]
- Rivin, C.J.; Grudt, T. Abscisic acid and the developmental regulation of embryo storage proteins in maize. Plant Physiol. 1991, 95, 358–365. [Google Scholar] [CrossRef]
- Gupta, S.; Parakh, D.B.; Verma, S.K. Ex situ conservation of strawberry (Fragaria spp.) germplasm in ICAR-NBPGR. Acta Hortic. 2020, 1297, 89–98. [Google Scholar] [CrossRef]
- Martínez-Santos, E.; Cruz-Cruz, C.A.; Spinoso-Castillo, J.L.; Bello-Bello, J.J. In vitro response of vanilla (Vanilla planifolia Jacks. ex Andrews) to PEG-induced osmotic stress. Sci. Rep. 2021, 11, 22611. [Google Scholar] [CrossRef]
- Silva, T.d.S.; Nepomuceno, C.F.; Soares, T.L.; de Santana, J.R.F. In vitro conservation of Poincianella pyramidalis (Tul.) L.P. Queiroz under minimal growth conditions. Ciência E Agrotecnologia 2019, 43, 14519. [Google Scholar] [CrossRef] [Green Version]
- Reed, B.M. Application of gas-permeable bags for in vitro cold storage of strawberry germplasm. Plant Cell Rep. 1991, 10, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Kovalchuk, I.; Lyudvikova, Y.; Volgina, M.; Reed, B.M. Medium, container and genotype all influence in vitro cold storage of apple germplasm. Plant Cell. Tissue Organ Cult. 2009, 96, 127–136. [Google Scholar] [CrossRef]
- Irish, B.M.; Goenaga, R.; Reed, B.M. Amending storage vessel and media increase transfer interval of Musa spp. tissue culture plantlets. J. Agric. Univ. Puerto Rico 2013, 97, 1–13. [Google Scholar]
- Jourdan, P. Short- and medium-term in vitro conservation and management of germplasm within the USDA’s National Plant Germplasm System. Acta Hortic. 2018, 1224, 31–38. [Google Scholar] [CrossRef]
- Khan, M.R.; Muhammad, A.; Hussain, I.; Shah, S.H.; Kumar, T.; Inam, S.M.; Zubair, H.U.; Rehman, A.; Sher, N.; Rehman, S.; et al. Rapid in vitro multiplication of sugarcane elite genotypes and detection of sugarcane mosaic virus through two steps RT-PCR. Int. J. Agric. Biol. 2012, 14, 870–878. [Google Scholar]
- Tyagi, R.K.; Goswami, R.; Sanayaima, R.; Singh, R.; Tandon, R.; Agrawal, A. Micropropagation and slow growth conservation of cardamom (Elettaria cardamomum Maton). Vitr. Cell. Dev. Biol.-Plant 2009, 45, 721–729. [Google Scholar] [CrossRef]
- Du, Y.P.; Li, W.Y.; Zhang, M.F.; Bin He, H.; Jia, G.X. The establishment of a slow-growth conservation system in vitro for two wild lily species. Afr. J. Biotechnol. 2012, 11, 1981–1990. [Google Scholar] [CrossRef]
- Jain, S.M. Prospects of in vitro conservation of date palm genetic diversity for sustainable production. Emir. J. Food Agric. 2011, 23, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Lyons, J.M. Chilling injury in plants. Annu. Rev. Plant Physiol. 1973, 24, 445–466. [Google Scholar] [CrossRef]
- Graham, D.; Patterson, B.D. Responses of plants to low, nonfreezing temperatures: Proteins, metabolism, and acclimation. Ann. Rev. Plant Physiol. 1982, 33, 347–374. [Google Scholar] [CrossRef]
- Bekheet, S.A. In vitro conservation of date palm tissue cultures. In Date Palm Biotechnology Protocols Volume II: Germplasm Conservation and Molecular Breeding, Methods in Molecular Biology; Al-Khayri, J., Jain, S., Johnson, D., Eds.; Springer Science+Business Media LLC.: Berlin/Heidelberg, Germany, 2017; Volume 1638, pp. 15–24. [Google Scholar] [CrossRef]
- Bekheet, S.A.; Matter, M.A.H.S.; Taha, E.-A.A.A. In vitro conservation of jojoba (Simmondsia chinensis) shootlet cultures using osmotic stress and low temperature. Middle East J. 2016, 5, 396–402. [Google Scholar]
- Rahayu, E.S.; Habibah, N.A. Optimization of in vitro conservation protocol of Carica pubescens lenne & k. koch through medium concentration, temperature and irradiation duration decrease. Biosaintifika J. Biol. Biol. Educ. 2016, 8, 87–95. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, K.S.; Aloufa, M.A.I. Slow growth in vitro culture for conservation of Hancornia speciosa gomes. Floresta 2022, 52, 7–16. [Google Scholar] [CrossRef]
- Bhuiyan, M.; Hossain, M.; Haque, M. In vitro conservation of taro (Colocasia esculenta var. globulifera) as influenced by mannitol. Bangladesh J. Agric. Res. 2016, 41, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Bello-Bello, J.J.; García-García, G.G.; Iglesias-Andreu, L. Conservación de vainilla (Vanilla planifolia jacks.) bajo condiciones de lento crecimiento in vitro. Rev. Fitotec. Mex. 2015, 38, 165. [Google Scholar] [CrossRef]
- Bautista-Aguilar, J.R.; Iglesias-Andreu, L.G.; Martínez-Castillo, J.; Ramírez-Mosqueda, M.A.; Ortiz-García, M.M. In vitro conservation and genetic stability in Vanilla planifolia Jacks. HortScience 2021, 56, 1494–1498. [Google Scholar] [CrossRef]
- da Silva, R.L.; Ferreira, C.F.; da Silva Ledo, C.A.; de Souza, E.H.; da Silva, P.H.; de Carvalho Costa, M.A.P.; Souza, F.V.D. Viability and genetic stability of pineapple germplasm after 10 years of in vitro conservation. Plant Cell. Tissue Organ Cult. 2016, 127, 123–133. [Google Scholar] [CrossRef]
- Panis, B.; Nagel, M.; Van den Houwe, I. Challenges and prospects for the conservation of crop genetic resources in field genebanks, in in vitro collections and/or in liquid nitrogen. Plants 2020, 9, 1634. [Google Scholar] [CrossRef]
- Rodrigues, P.H.V.; Oliveira, E.; Demetrio, C.; Ambrosano, G.; Piedade, S.M.S. The Effect of slow-grown in vitro storage under different light spectra on banana plantlets Cv. Prata Catarina (AAB). Res. Sq. 2021, 150, 479–485. [Google Scholar]
- Van den houwe, I.; De Smet, K.; du Montcel, H.T.; Swennen, R. Variability in storage potential of banana shoot cultures under medium term storage conditions. Plant Cell. Tissue Organ Cult. 1995, 42, 269–274. [Google Scholar] [CrossRef]
- Van den houwe, I.; Chase, R.; Sardos, J.; Ruas, M.; Kempenaers, E.; Guignon, V.; Massart, S.; Carpentier, S.; Panis, B.; Rouard, M.; et al. Safeguarding and using global banana diversity: A holistic approach. CABI Agric. Biosci. 2020, 1, 15. [Google Scholar] [CrossRef]
- Kanchanapoom, K.; Promssorn, N. Micropropagation and in vitro germplasm conservation of endangered Musa balbisiana ‘Kluai Hin’ (BBB group). Afr. J. Biotechnol. 2012, 11, 6464–6469. [Google Scholar] [CrossRef]
- Zainy, A.H.S.; Erum, S.; Shinwari, Z.K.; Alam, J.; Khan, U.; Jan, S.A. Ex-situ conservation and morpho-biochemical analysis of exotic cultivars of banana. Pak. J. Bot. 2019, 51, 1703–1709. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Trejgell, A.; Kamińska, M.; Tretyn, A. In vitro slow growth storage of Senecio macrophyllus shoots. Acta Physiol. Plant. 2015, 37, 234. [Google Scholar] [CrossRef] [Green Version]
- Aloni, R. The induction of vascular tissues by auxin. In Plant Hormones; Springer: Dordrecht, The Netherlands, 2010; pp. 485–518. ISBN 9781402026867. [Google Scholar]
- Lolaei, A.; Mobasheri, S.; Bemana, R.; Teymori, N. Role of paclobutrazol on vegetative and sexual growth of plants. Int. J. Agric. Crop Sci. 2013, 5, 958–961. [Google Scholar]
- Indrayanti, R.; Putri, R.E.; Sedayu, A.; Adisyahputra. Effect of paclobutrazol for in vitro medium-term storage of banana variant cv. Kepok (Musa acuminata x balbisiana Colla). AIP Conf. Proc. 2018, 2019, 020009. [Google Scholar] [CrossRef]
- El-Dawayati, M.M. In vitro conservation of date palm shoot-tip explants and callus cultures under minimal growth conditions. In Methods in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2017; Volume 1638, pp. 49–59. [Google Scholar]
- Bednarek, P.T.; Orłowska, R. Plant tissue culture environment as a switch-key of (epi)genetic changes. Plant Cell. Tissue Organ Cult. 2020, 140, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Kamińska, M.; Gołębiewski, M.; Tretyn, A.; Trejgell, A. Efficient long-term conservation of Taraxacum pieninicum synthetic seeds in slow growth conditions. Plant Cell, Tissue Organ Cult. 2018, 132, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Leva, A.R.; Petruccelli, R.; Rinaldi, L.M.R. Somaclonal variation in tissue culture: A case study with olive. In Recent Advances in Plant In Vitro Culture; InTech: Croatia, Balkans, 2012. [Google Scholar]
- Cassells, A.C.; Curry, R.F. Oxidative stress and physiological, epigenetic and genetic variability in plant tissue culture: Implications for micropropagators and genetic engineers. Plant Cell. Tissue Organ Cult. 2001, 64, 145–157. [Google Scholar] [CrossRef]
- Tyagi, P.; Khanduja, S.; Kothari, S.L. In vitro culture of Capparis decidua and assessment of clonal fidelity of the regenerated plants. Biol. Plant. 2010, 54, 126–130. [Google Scholar] [CrossRef]
- Kaçar, Y.A.; Dönmez, D.; Biçen, B.; Erol, M.H.; Şimşek, Ö.; Mendi, Y.Y. Micropropagation of Spathiphyllum with temporary immersion bioreactor system. Turkish J. Agric.-Food Sci. Technol. 2020, 8, 1195–1200. [Google Scholar]
- Simsek, O.; Donmez, D.; Saridas, M.A.; Paydas-Kargi, S.; Aka-Kacar, Y. Genetic relationship and polymorphism of turkish myrtles (Myrtus communis L.) as revealed by inter simple sequence repeat (issr). Appl. Ecol. Environ. Res. 2020, 18, 1141–1149. [Google Scholar] [CrossRef]
- Dönmez, D. Regeneration of plants from alginate-encapsulated shoot tips of myrtle (Myrtus communis L.). Erwerbs-Obstbau 2022, 64, 307–314. [Google Scholar] [CrossRef]
- Kamińska, M.; Tretyn, A.; Trejgell, A. Genetic stability assessment of Taraxacum pieninicum plantlets after long-term slow growth storage using ISSR and SCoT markers. Biologia 2020, 75, 599–604. [Google Scholar] [CrossRef]
- Ruta, C.; Lambardi, M.; Ozudogru, E.A. Biobanking of Vegetable Genetic Resources by In Vitro Conservation and Cryopreservation; Springer: Dordrecht, The Netherlands, 2020; Volume 29, ISBN 1053102002051. [Google Scholar]
- Guarino, L.; Ramanatha Rao, V.; Reid, R. Collecting Plant Genetic Diversity: Technical Guidelines; IUCN—International Union for Conservation of Nature: Gland, Switzerland, 1995.
- CBD Progress in Implementation of the targets of the global strategy for plant conservation. Eighteenth meeting of the Subsidiary Body on Scientific, Technical and Technological Advice, Montreal, QC, Canada, 23–28 June 2014.
- Benelli, C. Plant cryopreservation: A look at the present and the future. Plants 2021, 10, 2744. [Google Scholar] [CrossRef]
- Panis, B. Sixty years of plant cryopreservation: From freezing hardy mulberry twigs to establishing reference crop collections for future generations. Acta Hortic. 2019, 1234, 1–8. [Google Scholar] [CrossRef]


| Species | Medium | Temperature (°C) | Light Condition | Storage Time (Months) | Survival (%) | References |
|---|---|---|---|---|---|---|
| Arbutus unedo | MS, 1 mg L−1 zeatin | 18 | 16 h, 30 μE s−1 m−2 | 6 | 80 | [15] |
| Castanea sativa | WPM, 0.44 μM BA, 30 g L−1 sucrose | 8 | 16 h, 30 μM m−2 s−1 | 48 | 82 | [16] |
| Ceratonia siliqua | MS, 0.1 mg L−1 BA | 18 | 16 h, 30 μE s−1 m−2 | 6 | 100 | [15] |
| Citrus jambhiri | WPM, 25 g L−1 sucrose | 22 | 12 h, 20 μmol m−2 s−1 | 12 | NR | [17] |
| Crataegus monogyna | ½MS, 5 mM BA, 0.5 mM IBA, 20 g L−1 sucrose | 4 | Darkness | 7–12 | 99 | [18] |
| Cydonia oblonga | ½MS, 5 mM BA, 0.5 mM IBA, 20 g L−1 sucrose | 4 | Darkness | 7–12 | 99 | [18] |
| Eriobotrya japonica | ½MS, 5 mM BA, 0.5 mM IBA, 20 g L−1 sucrose | 4 | Darkness | 7–12 | 99 | [18] |
| Ficus carica | ½MS, 5 mM BA, 0.5 mM IBA, 20 g L−1 sucrose | 4 | Darkness | 7–12 | 99 | [18] |
| Fragaria x ananassa | MS, 1 mg L−1 BA, 30 g L−1 sucrose | 4 | 16 h, 40 μmol m−2 s−1 | 7 | 32 | [19] |
| Fragaria spp. | MS | 4 | 10 h, 10 μmol m−2 s−1 | 15–18 | NR | [20] |
| Knop medium | 4 | Darkness | 15 | 100 | [21] | |
| Malus domestica | ½MS, 5 mM BA, 0.5 mM IBA, 20 g L−1 sucrose | 4 | Darkness | 7–12 | 99 | [18] |
| MS, 1 mg L−1 BA, 30 g L−1 sucrose | 4 | 16 h, 40 μmol m−2 s−1 | 7 | 90 | [19] | |
| MS, 0.44 μM BA, 130.5 mM sucrose | 4 | Darkness | 6 | NR | [22] | |
| Malus spp. | MS (25–50% NO3), 2% sucrose + 2% mannitol | 4 | 10 h, 10 μmol m−2 s−1 | ≥36 | NR | [20] |
| MS, 3% sucrose | 4 | 10 h, 15 μmol m−2 s−1 | 18–20 | NR | [23] | |
| Pistacia lentiscus | MS, 1 mg L−1 BA, 3% sucrose | 4 | Darkness | 12 | NR | [24] |
| Prunus avium | MS, 1 mg L−1 BA, 30 g L−1 sucrose | 4 | 16 h, 40 μmol m−2 s−1 | 7 | 79.4 | [19] |
| Prunus avium × P. cerasus | MS, 1 mg L−1 BA, 30 g L−1 sucrose | 4 | 16 h, 40 μmol m−2 s−1 | 7 | 92 | [19] |
| DKW, 0.5 mg L−1 BA, 45 or 60 g L−1 sucrose | 4 | Darkness | 16 | NR | [25] | |
| Prunus mahaleb | ½MS media without sucrose | 25 | 16 h, 43.4 μmol m−2 s−1 | 4 | 74.1 | [26] |
| Prunus spp. | ½MS, 5 mM BA, 0.5 mM IBA, 20 g L−1 sucrose | 4 | Darkness | 7–12 | 98.6 | [18] |
| MS, 0.5 mg L−1 BA, 0.1 mg L−1 IBA, 2% sucrose + 2% mannitol | 4 | 10 h, 10 μmol m−2 s−1 | 30 | NR | [20] | |
| MS, 2.2 μM BA, 0.49 μM IBA, 20 g L−1 sucrose | 4 | Darkness | 12 | 100 | [27] | |
| Prunus webbii | MS, 0.7 mg L−1 BA, 0.01 mg L−1 NAA, 0.1 mg L−1 GA3, 3% sucrose | 4 | Darkness | 6 | 42.6 | [28] |
| Punica granatum | ½MS, 5 mM BA, 0.5 mM IBA, 20 g L−1 sucrose | 4 | Darkness | 7–12 | 99 | [18] |
| Pyrus communis | ½MS, 5 mM BA, 0.5 mM IBA, 20 g L−1 sucrose | 4 | Darkness | 7–12 | 99 | [18] |
| MS, 1 mg L−1 BA, 30 g L−1 sucrose | 4 | 16 h, 40 μmol m−2 s−1 | 7 | 91 | [19] | |
| Pyrus spp. | MS, 30 g L−1 sucrose | 4 | Darkness | 6 | 100 | [21] |
| MS, 3% sucrose, 0.5 mg L−1 BA, 0.1 mg L−1 IBA/without PGRs | 4 | 10 h, 7 μmol m−2 s−1 | 18/15 | NR | [29] | |
| MS (25% NO3), 2% sucrose, 2% mannitol | 4 | 10 h, 10 μmol m−2 s−1 | 36 | NR | [20] | |
| ½MS nitrogen | 1-4 | 12 h, (10–20 µE m−2 s−1)/dark | 12–48 | NR | [30] | |
| MS | 4 | 12 h, 10 µE m−2 s−1 | 48 | NR | [31] | |
| Ribes nigrum | MS, 0.5 mg L−1 BA, 0.1 mg L−1 IBA, 2% sucrose + 2% mannitol | 4 | 10 h, 10 μmol m−2 s−1 | 18 | NR | [20] |
| Rubus spp. | MS, 0.5 mg L−1 BA, 0.1 mg L−1 IBA, 3% sucrose | 4 | 10 h, 10 μmol m−2 s−1 | 15 | NR | [20] |
| Vaccinium myrtillus | MS, 30 g L−1 sucrose | 4 | Darkness | 6 | 90 | [32] |
| Vitis heyneana | MS, 0.05 mg L−1 IBA, 0.1 mg L−1 IAA, 0.5 mg L−1 ABA, 10 g L−1 mannitol | 10 | 16 h, 40 μmol m−2 s−1 | 12 | 47.8 | [33] |
| Vitis vinifera | ¾ MS, 5.5% sorbitol | 5 | Darkness | 12 | 88.9 | [34] |
| MS, 300 µM ribose | 15 | 16 h, 3000 Lux | 12 | 73 | [35] | |
| Ziziphus jujuba | MS, 1 mg L−1 BA, 0.05 mg L−1 IBA, 3% sucrose | 4 | Darkness | 10 | 78.6 | [36] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benelli, C.; Tarraf, W.; Izgu, T.; De Carlo, A. In Vitro Conservation through Slow Growth Storage Technique of Fruit Species: An Overview of the Last 10 Years. Plants 2022, 11, 3188. https://doi.org/10.3390/plants11233188
Benelli C, Tarraf W, Izgu T, De Carlo A. In Vitro Conservation through Slow Growth Storage Technique of Fruit Species: An Overview of the Last 10 Years. Plants. 2022; 11(23):3188. https://doi.org/10.3390/plants11233188
Chicago/Turabian StyleBenelli, Carla, Waed Tarraf, Tolga Izgu, and Anna De Carlo. 2022. "In Vitro Conservation through Slow Growth Storage Technique of Fruit Species: An Overview of the Last 10 Years" Plants 11, no. 23: 3188. https://doi.org/10.3390/plants11233188
APA StyleBenelli, C., Tarraf, W., Izgu, T., & De Carlo, A. (2022). In Vitro Conservation through Slow Growth Storage Technique of Fruit Species: An Overview of the Last 10 Years. Plants, 11(23), 3188. https://doi.org/10.3390/plants11233188