
N-glycoproteins in Plant Cell Walls: A Survey

Abstract
:1. Introduction
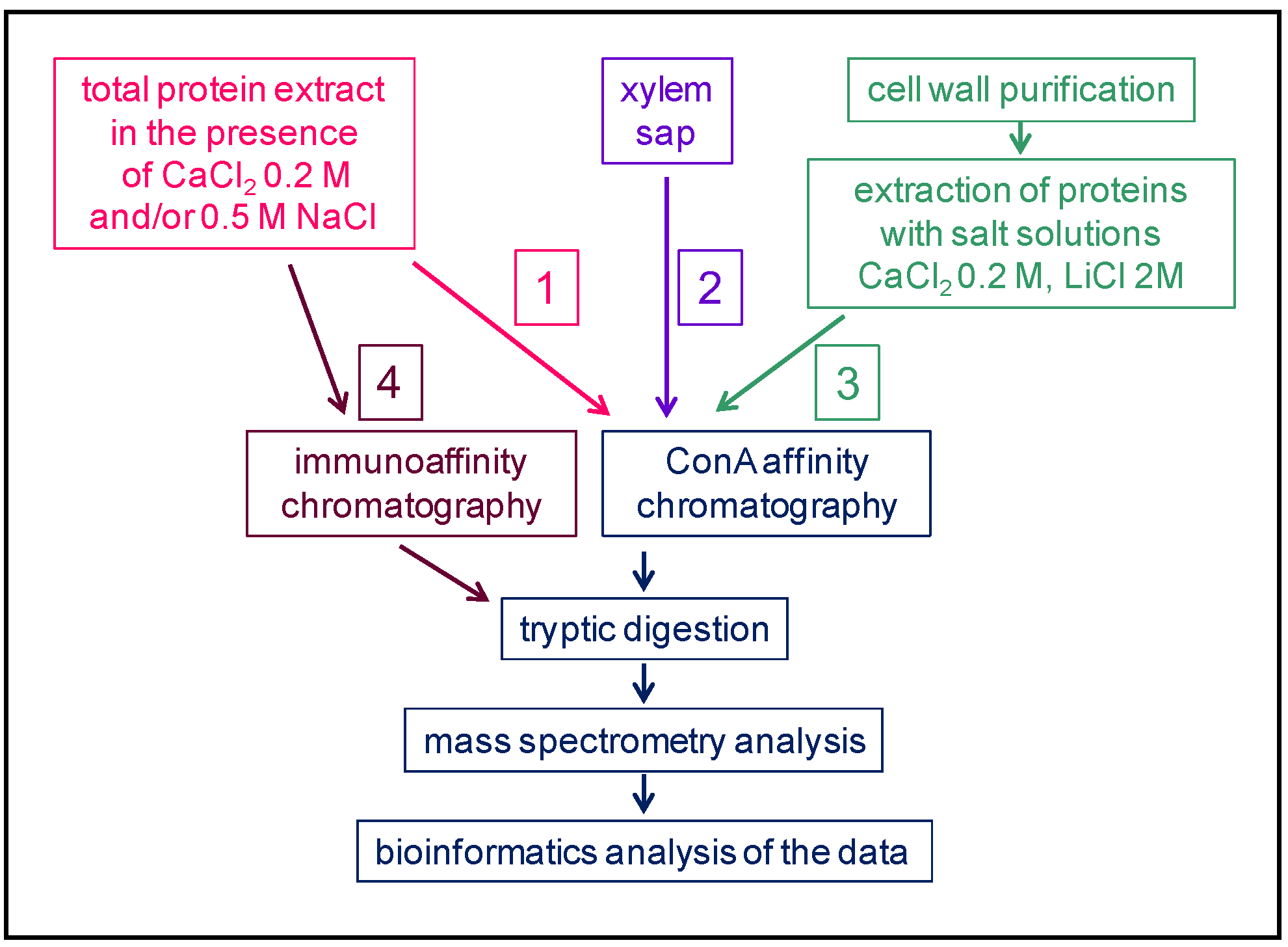
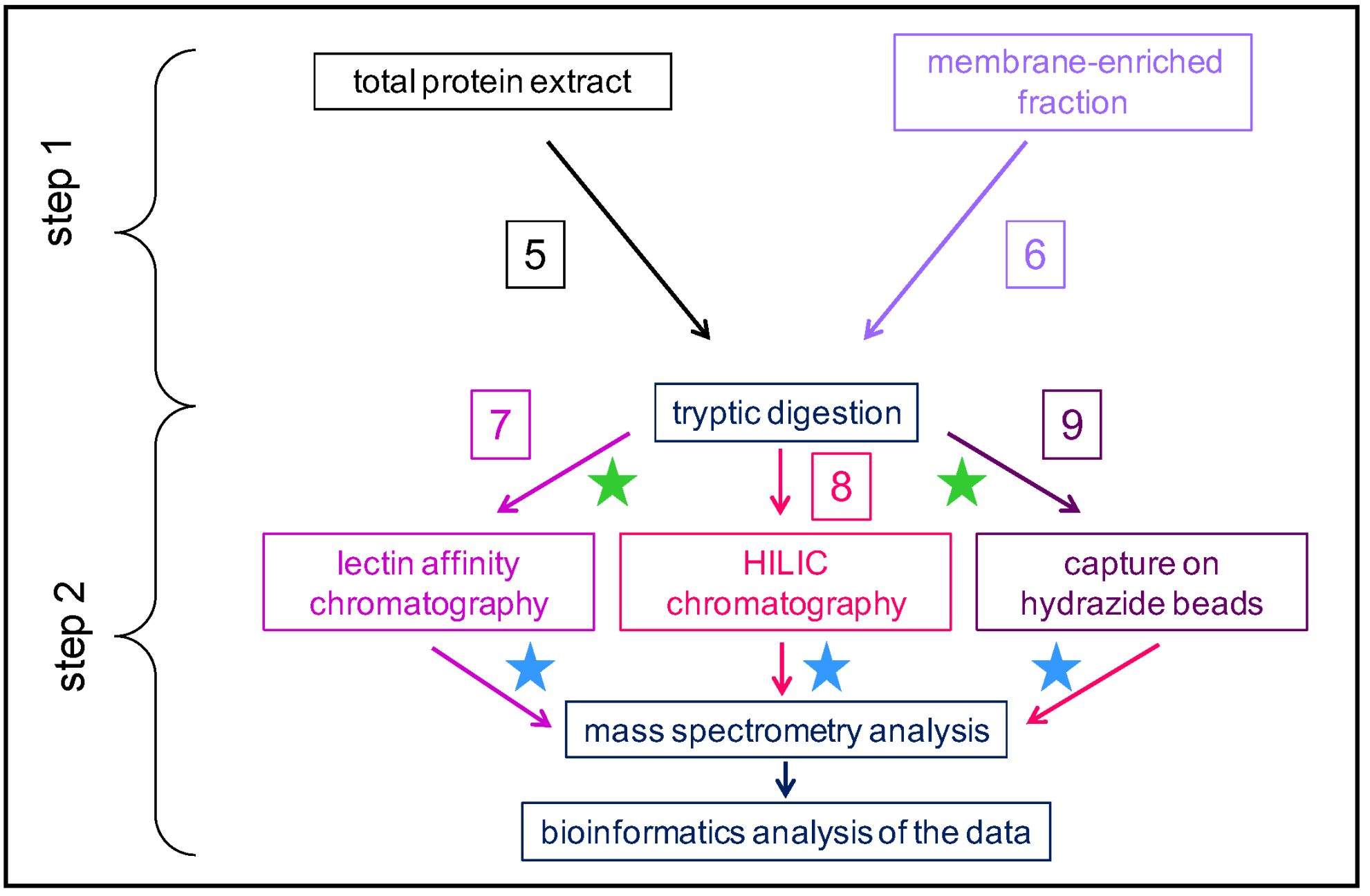
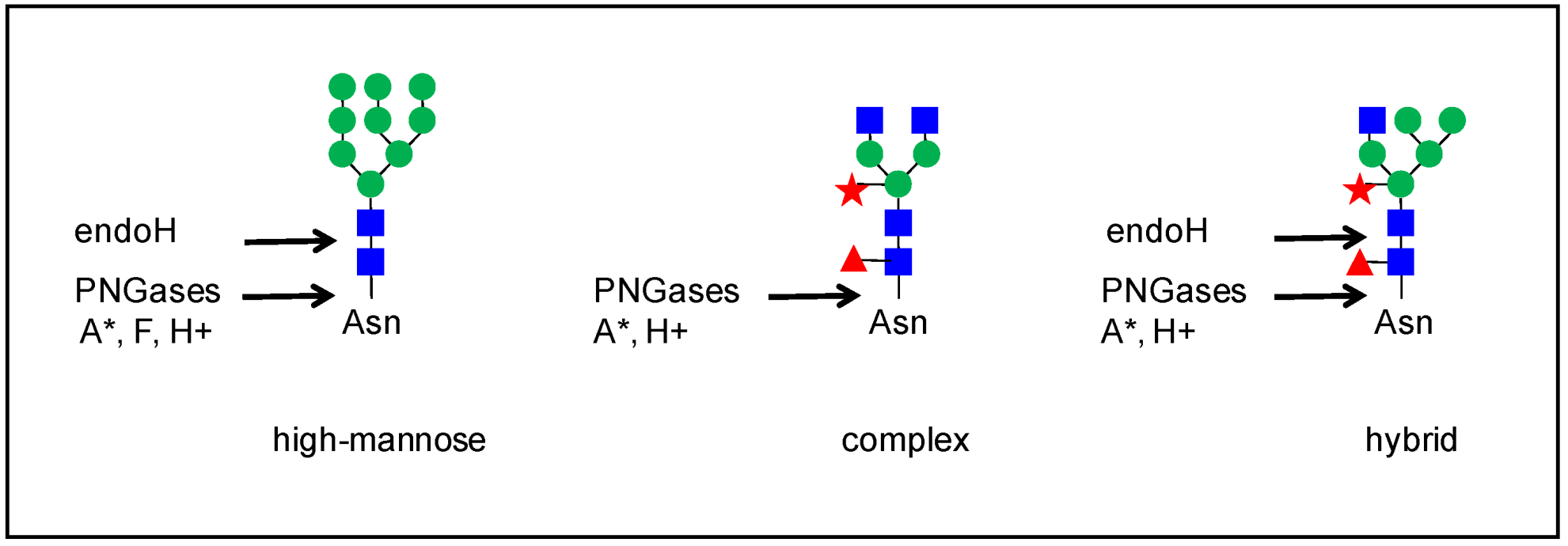
2. An Overview of the Strategies
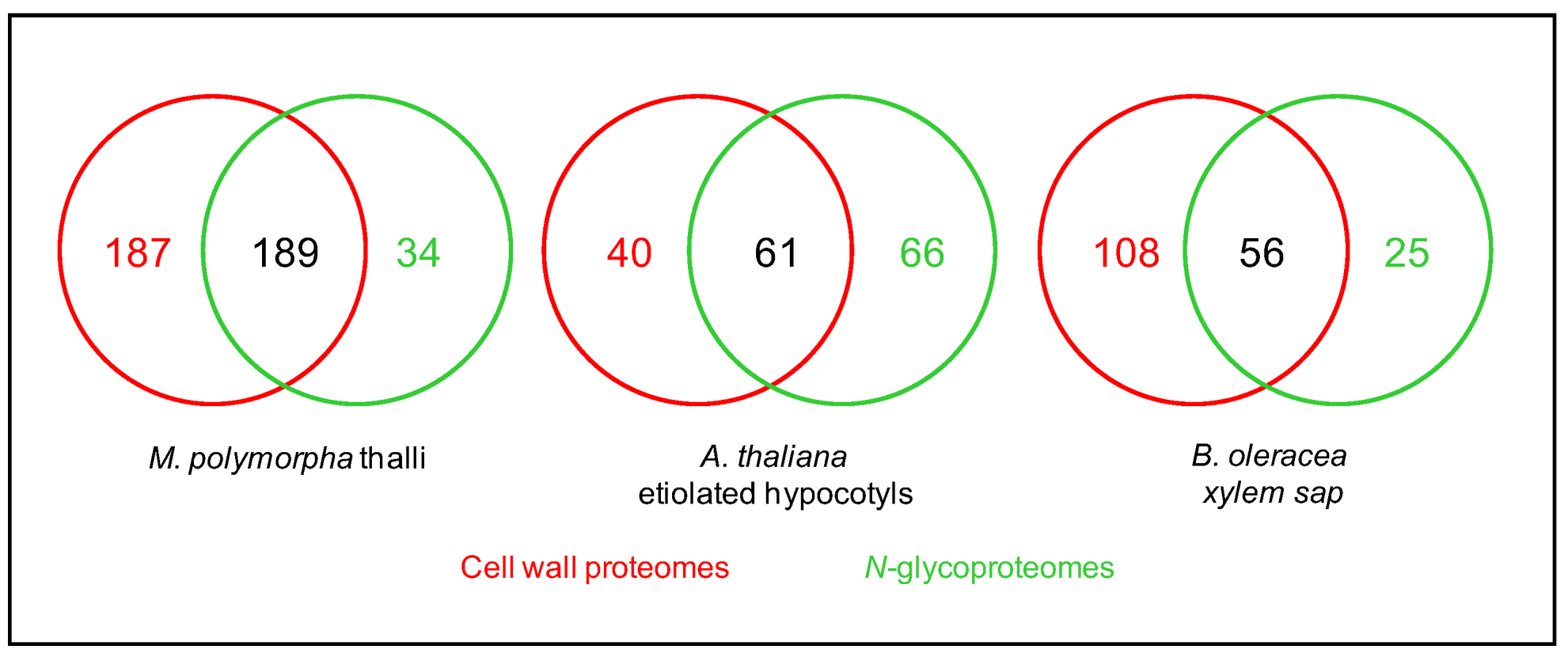
3. An Overview of the Present N-glycoproteomics Studies
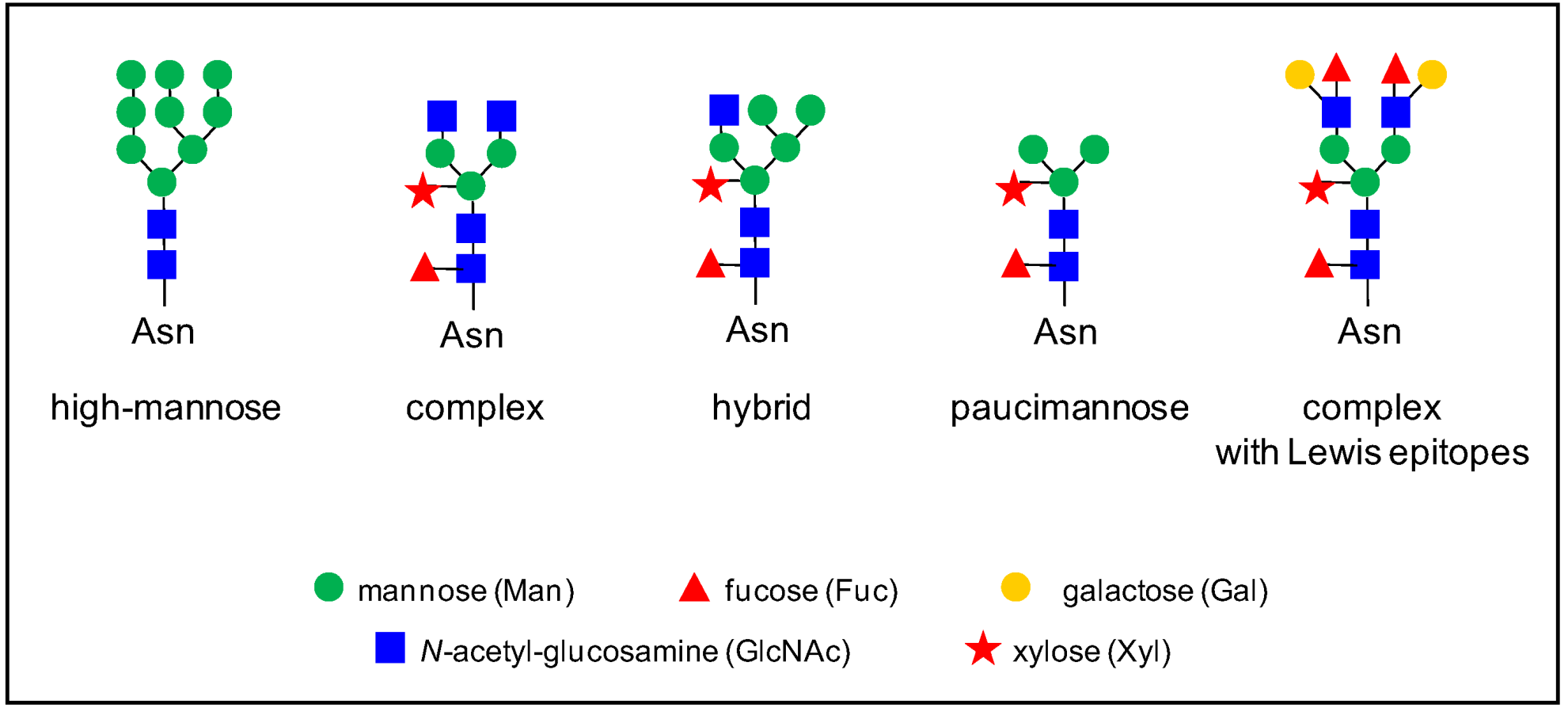
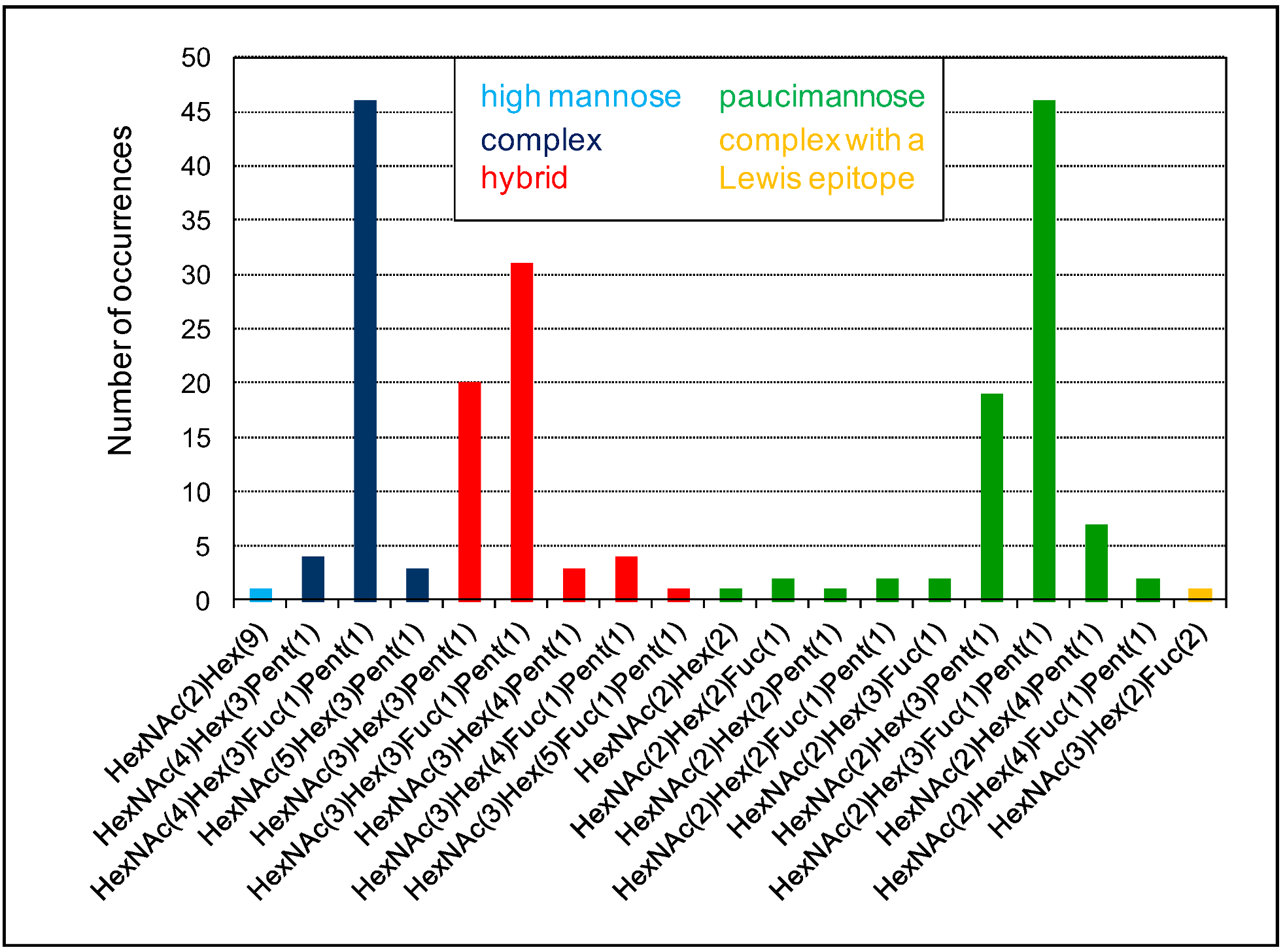
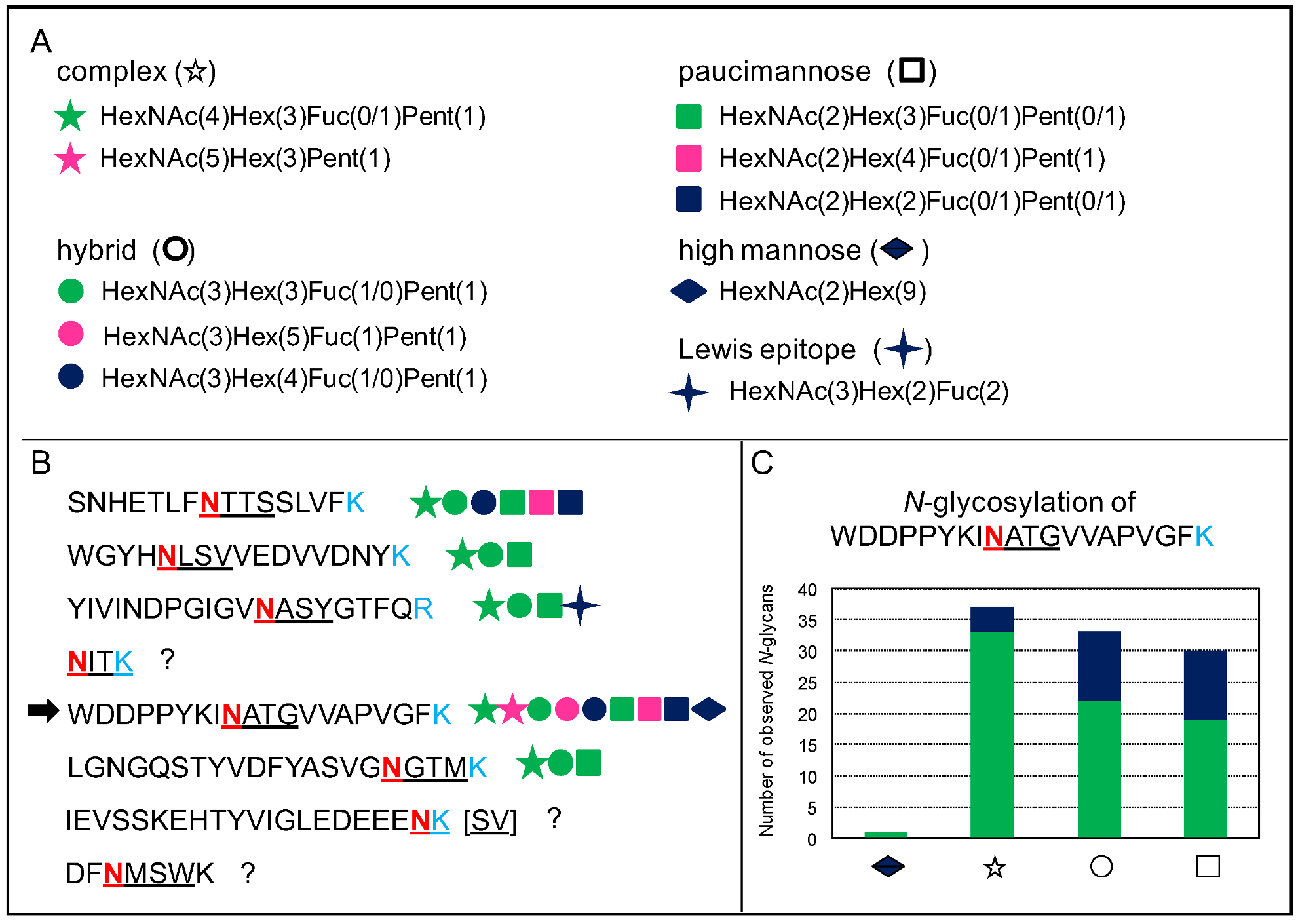
4. The Heterogeneity of Plant N-glycans
5. Roles of N-glycans
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spiro, R.G. Protein glycosylation: Nature, distribution, enzymatic formation, and disease implications of glycopeptide bonds. Glycobiology 2002, 12, 43R–56R. [Google Scholar] [PubMed]
- Sigrist, C.; de Castro, E.; Cerutti, L.; Cuche, B.; Hulo, N.; Bridge, A.; Bougueleret, L.; Xenarios, I. New and continuing developments at PROSITE. Nucleic Acids Res. 2012, 41, D344–D347. [Google Scholar] [CrossRef] [Green Version]
- Nguema-Ona, E.; Vicré-Gibouin, M.; Gotté, M.; Plancot, B.; Lerouge, P.; Bardor, M.; Driouich, A. Cell wall O-glycoproteins and N-glycoproteins: Aspects of biosynthesis and function. Front. Plant Sci. 2014, 5, 499. [Google Scholar] [CrossRef] [Green Version]
- Kieliszewski, M.J. The latest hype on Hyp-O-glycosylation codes. Phytochemistry 2001, 57, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Canut, H.; Albenne, C.; Jamet, E. Post-translational modifications of plant cell wall proteins and peptides: A survey from a proteomics point of view. Biochim. Biophys. Acta 2016, 1864, 983–990. [Google Scholar] [CrossRef]
- Duruflé, H.; Hervé, V.; Balliau, T.; Zivy, M.; Dunand, C.; Jamet, E. Proline hydroxylation in cell wall proteins: Is it yet possible to define rules? Front. Plant Sci. 2017, 8, 1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.; Varnai, P.; Lamport, D.T.; Yuan, C.; Xu, J.; Qiu, F.; Kieliszewski, M.J. Plant O-hydroxyproline arabinogalactans are composed of repeating trigalactosyl subunits with short bifurcated side chains. J. Biol. Chem. 2010, 285, 24575–24583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulick, M.; Bertozzi, C. The glycosylphosphatidylinositol anchor: A complex membrane-anchoring structure for proteins. Biochemistry 2014, 47, 6991–7000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faye, L.; Boulaflous, A.; Benchabane, M.; Gomord, V.; Michaud, D. Protein modifications in the plant secretory pathway: Current status and practical implications in molecular pharming. Vaccine 2005, 23, 1770–1778. [Google Scholar] [CrossRef] [PubMed]
- Gomord, V.; Fitchette, A.; Menu-Bouaouiche, L.; Saint-Jore-Dupas, C.; Plason, C.; Michaud, D.; Faye, L. Plant-specific glycosylation patterns in the context of therapeutic protein production. Plant Biotechnol. J. 2010, 8, 564–587. [Google Scholar] [CrossRef] [PubMed]
- Toustou, C.; Walet-Balieu, M.; Kiefer-Meyer, M.; Houdou, M.; Lerouge, P.; Foulquier, F.; Bardor, M. Towards understanding the extensive diversity of protein N-glycan structures in eucaryotes. Biol. Rev. 2022, 97, 732–748. [Google Scholar] [CrossRef] [PubMed]
- Schoberer, J.; Strasser, R. Plant glyco-biology. Semin. Cell Dev. Biol. 2018, 80, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R. Plant protein glycosylation. Glycobiology 2016, 26, 926–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strasser, R.; Seifert, G.; Doblin, M.; Johnson, K.; Ruprecht, C.; Pfrengle, F.; Bacic, A.; Estevez, J. Cracking the “sugar code”: A snapshot of N- and O-glycosylation pathways and functions in plants cells. Front. Plant Sci. 2021, 12, 640919. [Google Scholar] [CrossRef]
- Mócsai, R.; Figl, R.; Troschl, C.; Strasser, R.; Svehla, E.; Windwarder, M.; Thader, A.; Altmann, F. N-glycans of the microalga Chlorella vulgaris are of the oligomannosidic type but highly methylated. Sci. Rep. 2019, 9, 33. [Google Scholar] [CrossRef] [Green Version]
- Mathieu-Rivet, E.; Scholtz, M.; Arias, C.; Dardelle, F.; Schulze, S.; Le Mauff, F.; Teo, G.; Hochmal, A.; Blanco-Rivero, A.; Loutenier-Bourhiss, C.; et al. Exploring the N-glycosylation pathway in Chlamydomonas reinhardtii unravels novel complex structures. Mol. Cell. Proteom. 2013, 12, 3160–3183. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-May, E.; Sørensen, I.; Fei, Z.; Zhang, S.; Domozych, D.; Rose, J. The secretome and N-glycosylation profiles of the charophycean green alga, Penium margaritaceum, resemble those of embryophytes. Proteomes 2018, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Remco, V.; Loutelier-Bourhis, C.; Fitchette, A.; Margerie, P.; Gonneau, M.; Faye, L.; Lerouge, P. Protein N-glycosylation is similar in the moss Physcomitrella patens and in higher plants. Planta 2003, 218, 269–275. [Google Scholar]
- Pattison, R.; Amtmann, A. N-glycan production in the endoplasmic reticulum of plants. Trends Plant Sci. 2009, 14, 92–99. [Google Scholar] [CrossRef]
- Fanata, W.; Lee, K.; Son, B.; Yoo, J.; Harmoko, R.; Ko, K.; Ramasamy, N.; Kim, K.; Oh, D.; Jung, H.; et al. N-glycan maturation is crucial for cytokinin-mediated development and cellulose synthesis in Oryza sativa. Plant J. 2013, 73, 966–979. [Google Scholar] [CrossRef]
- Strasser, R.; Altmann, F.; Mach, L.; Glössel, J.; Steinkellner, H. Generation of Arabidopsis thaliana plants with complex N-glycans lacking β1,2-linked xylose and core α1,3-linked fucose. FEBS Lett. 2004, 561, 132–136. [Google Scholar] [CrossRef]
- Minic, Z.; Jamet, E.; Negroni, L.; der Garabedian, P.A.; Zivy, M.; Jouanin, L. A sub-proteome of Arabidopsis thaliana trapped on Concanavalin A is enriched in cell wall glycoside hydrolases. J. Exp. Bot. 2007, 58, 2503–2512. [Google Scholar] [CrossRef]
- Ligat, L.; Lauber, E.; Albenne, C.; San Clemente, H.; Valot, B.; Zivy, M.; Pont-Lezica, R.; Arlat, M.; Jamet, E. Analysis of the xylem sap proteome of Brassica oleracea reveals a high content in secreted proteins. Proteomics 2011, 11, 1798–1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Giboulot, A.; Zivy, M.; Valot, B.; Jamet, E.; Albenne, C. Combining various strategies to increase the coverage of the plant cell wall glycoproteome. Phytochemistry 2011, 72, 1109–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brewer, C.; Bhattacharya, L. Concanavalin A interactions with asparagine-linked glycopeptides. The mechanisms of binding of oligomannose, bisected hybrid, and complex type carbohydrates. Glycoconj. J. 1988, 5, 159–173. [Google Scholar] [CrossRef]
- Beilhammer, G.; Maresch, D.; Altmann, F.; Van Damme, E.; Strasser, R. Lewis a glycans are present on proteins involved in cell wall biosynthesis and appear evolutionary conserved among natural Arabidopsis thaliana accessions. Front. Plant Sci. 2021, 12, 630891. [Google Scholar] [CrossRef]
- Etxebarria, J.; Calvo, J.; Martin-Lomas, M.; Reichardt, N. Lectin-array blotting: Profiling protein glycosylation in complex mixtures. ACS Chem. Biol. 2012, 7, 1729–1737. [Google Scholar] [CrossRef]
- Lis, H.; Sharon, N. Lectin-carbohydrate interactions. Curr. Opin. Struct. Biol. 1991, 1, 741–749. [Google Scholar] [CrossRef]
- Shibuya, N.; Goldstein, I.; Van Damme, E.; Peumans, W. Binding properties of a mannose-specific lectin from the snowdrop (Galanthus nivalis) bulb. J. Biol. Chem. 1988, 263, 728–734. [Google Scholar] [CrossRef]
- Van Damme, E.; Smith, D.; Cummings, R.; Peumans, W. Glycan arrays to decipher the specificity of plant lectins. In The Molecular Immunology of Complex Carbohydrates-3. Advances in Experimental Medicine and Biology; Wu, A., Ed.; Springer: Boston, MA, USA, 2011; Volume 705, pp. 757–767. [Google Scholar]
- Cummings, R.; Darwill, A.; Ezler, M.; Hahn, M. Glycan-recognizing probes as tools. In Essentials of Glycobiology [Internet], 4th ed.; Varki, A., Cummings, R., Esko, J., Stanley, P., Hart, G., Aebi, M., Mohnen, D., Kinoshita, T., Packer, N., Prestegard, J., et al., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2022. [Google Scholar]
- Ruiz-May, E.; Hucko, S.; Howe, K.; Zhang, S.; Sherwood, R.; Thannhauser, T.; Rose, J. A comparative study of lectin affinity based plant N-glycoproteome profiling using tomato fruit as a model. Mol. Cell. Proteom. 2014, 13, 566–579. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.; Zhu, D.; Duan, W.; Deng, X.W.; Zhang, J.; Ye, X.; Yan, Y. Plasma membrane N-glycoproteome analysis of wheat seedling leaves under drought stress. Int. J. Biol. Macromol. 2021, 193, 1541–1550. [Google Scholar] [CrossRef]
- Bu, T.; Shen, J.; Chao, Q.; Shen, Z.; Yan, Z.; Zheng, H.; Wang, B. Dynamic N-glycoproteome analysis of maize seedling leaves during de-etiolation using Concanavalin A lectin affinity chromatography and a nano LC-MS/MS-based iTRAQ approach. Plant Cell Rep. 2017, 36, 1943–1958. [Google Scholar] [CrossRef]
- Yoo, J.; Ko, K.; Seo, H.; Park, S.; Fanata, W.; Harmoko, R.; Ramasamy, N.; Thulasinathan, T.; Mengiste, T.; Lim, J.; et al. Limited addition of the 6-arm β1,2-linked N-acetylglucosamine (GlcNAc) residue facilitates the formation of the largest N-glycan in plants. J. Biol. Chem. 2015, 290, 16560–16572. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Mentik, R.; Henquet, M.; Cordewener, J.; van Dijk, A.; Bosch, D.; America, A.; van der Krol, A. N-glycan occupancy of Arabidopsis N-glycoproteins. J. Proteom. 2013, 93, 343–355. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, H.; Du, H.; Liu, Z.; Bao, Z.; Shi, Q. Comparative N-glycoproteome analysis provides novel insight into the regulation mechanism in tomato (Solanum lycopersicum L.) during fruit ripening process. Plant Sci. 2020, 293, 110413. [Google Scholar] [CrossRef]
- Geng, F.; Liu, X.; Wang, J.; He, R.; Zhao, J.; Xiang, D.; Zou, L.; Peng, L.; Zhao, G. In-depth mapping of the seed phosphoproteome and N-glycoproteome of Tartary buckwheat (Fagopyrum tataricum) usinf off-line pH RPLC fractionation and nLC-MS/MS. Int. J. Biol. Macromol. 2019, 137, 688–696. [Google Scholar] [CrossRef]
- Wang, T.; Cai, Z.; Gu, X.; Ma, H.; Du, Y.; Huang, K.; Voglmeir, J.; Liu, L. Discovery and characterization of a novel extremely acidic bacterial N-glycanase with combined advantages of PNGase F and A. Biosci. Rep. 2014, 34, e00149. [Google Scholar] [CrossRef]
- Veličković, D.; Liao, Y.; Thibert, S.; Veličković, M.; Anderton, C.; Voglmeir, J.; Stacey, G.; Zhou, M. Spatial mapping of plant N-glycosylation cellular heterogeneity inside soybean root nodules provided insights into legume-rhizobia symbiosis Front. Plant Sci. 2022, 13, 869281. [Google Scholar]
- Cao, L.; Diedrich, J.; Ma, Y.; Wang, N.; Pauthner, M.; Park, S.; Delahunty, C.; McLellan, J.; Burton, D.; Yates, J.; et al. Global site-specific analysis of glycoprotein N-glycan processing. Nat. Protoc. 2018, 13, 1196–1212. [Google Scholar] [CrossRef]
- Pedersen, C.; Loke, I.; Lorentzen, A.; Wolf, S.; Kamble, M.; Kristensen, S.; Munch, D.; Radutoiu, S.; Spillner, E.; Roepstorff, P.; et al. N-glycan maturation mutants in Lotus japonicus for basic and applied glycoprotein research. Plant J. 2017, 91, 394–407. [Google Scholar] [CrossRef] [Green Version]
- Zeng, W.; Ford, K.; Bacic, A.; Heazlewood, J. N-linked glycan micro-heterogeneity in glycoproteins of Arabidopsis. Mol. Cell. Proteom. 2018, 17, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Albenne, C.; Canut, H.; Boudart, G.; Zhang, Y.; San Clemente, H.; Pont-Lezica, R.; Jamet, E. Plant cell wall proteomics: Mass spectrometry data, a trove for research on protein structure/function relationships. Mol. Plant 2009, 2, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Desaire, H. Glycopeptide analysis, recent developments and applications. Mol. Cell. Proteom. 2013, 12, 893–901. [Google Scholar] [CrossRef] [Green Version]
- Pelham, H. The retention signal for soluble proteins of the endoplasmic reticulum. Trends Plant Sci. 2010, 15, 483–486. [Google Scholar] [CrossRef]
- San Clemente, H.; Jamet, E. WallProtDB, a database resource for plant cell wall proteomics. Plant Methods 2015, 11, 2. [Google Scholar] [CrossRef] [Green Version]
- Albenne, C.; Canut, H.; Jamet, E. Plant cell wall proteomics: The leadership of Arabidopsis thaliana. Front. Plant Sci. 2013, 4, 111. [Google Scholar] [CrossRef] [Green Version]
- Guerra-Guimarães, L.; Pinheiro, C.; Chaves, I.; Barros, D.; Ricardo, C. Protein dynamics in the plant extracellular space. Proteomes 2016, 4, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krause, C.; Richter, S.; Knöll, C.; Jürgens, G. Plant secretome-From cellular process to biological activity. Biochim. Biophys. Acta 2013, 1834, 2429–2441. [Google Scholar] [CrossRef]
- San Clemente, H.; Kolkas, H.; Jamet, E. Plant cell wall proteomes: The core of conserved protein families and the case of non-canonical proteins. Int. J. Biol. Sci. 2022, 23, 4273. [Google Scholar] [CrossRef]
- Kolkas, H.; Balliau, T.; Chourré, J.; Zivy, M.; Canut, H.; Jamet, E. The cell wall proteome of Marchantia polymorpha reveals specificities compared to those of flowering plants. Front. Plant Sci. 2022, 12, 765846. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, G.; Lv, D.; Li, X.; Yan, Y. N-linked glycoproteome profiling of seedling leaf in Brachypodium distachyon L. J. Proteome Res. 2015, 14, 1727–1738. [Google Scholar] [CrossRef]
- Catalá, C.; Howe, K.; Hucko, S.; Rose, J.; Thannhauser, T. Towards characterization of the glycoproteome of tomato (Solanum lycopersicum) fruit using Concanavalin A lectin affinity chromatography and LC-MALDI-MS/MS analysis. Proteomics 2011, 11, 1530–1544. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, K.; Pandey, P.; Rajamani, V.; Padmalatha, K.; Dhandapani, G.; Kanakachari, M.; Leelavathi, S.; Kumar, P.; Reddy, V. Glycoproteome of elongating cotton fiber cells. Mol. Cell. Proteom. 2013, 12, 3777–3789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Ma, L.; Cao, D.; Gong, Z.; Fan, J.; Hu, H.; Jin, X. Investigation of cell wall proteins of C. sinensis leaves by combining cell wall proteomics and N-glycoproteomics. BMC Plant Biol. 2021, 21, 384. [Google Scholar] [CrossRef] [PubMed]
- Sultana, N.; Florance, H.; Johns, A.; Smirnoff, N. Ascorbate deficiency influences the leaf cell wall glycoproteome in Arabidopsis thaliana. Plant Cell Environ. 2015, 38, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Medzihradszky, K.; Wang, Z.; Burlingame, A.; Chalkley, R. N-Glycopeptide profiling in Arabidopsis inflorescence. Mol. Cell. Proteom. 2016, 15, 2048–2054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Wang, D.; She, J.; Li, J.; Zhu, J.; She, Y. Endoplasmic reticulum-associated N-glycan degradation of cold-upregulated glycoproteins in response to chilling stress in Arabidopsis. New Phytol. 2016, 212, 282–296. [Google Scholar] [CrossRef] [Green Version]
- Irshad, M.; Canut, H.; Borderies, G.; Pont-Lezica, R.; Jamet, E. A new picture of cell wall protein dynamics in elongating cells of Arabidopsis thaliana: Confirmed actors and newcomers. BMC Plant Biol. 2008, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Duruflé, H.; Hervé, V.; Ranocha, P.; Balliau, T.; Zivy, M.; Chourré, J.; San Clemente, H.; Burlat, V.; Albenne, C.; Déjean, S.; et al. Cell wall adaptation of two contrasted ecotypes of Arabidopsis thaliana, Col and Sha, to sub-optimal growth conditions: An integrative study. Plant Sci. 2017, 263, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Shigeyama, T.; Watanabe, A.; Tokuchi, K.; Toh, S.; Sakurai, N.; Shibuya, N.; Kawakami, N. α-Xylosidase plays essential roles in xyloglucan remodelling, maintenance of cell wall integrity, and seed germination in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 5615–5629. [Google Scholar] [CrossRef] [Green Version]
- Lannoo, N.; Van Damme, E. Review/N-glycans: The making of a varied toolbox. Plant Sci. 2015, 239, 67–83. [Google Scholar] [CrossRef]
- Strasser, R.; Bondili, J.; Schoberer, J.; Svoboda, B.; Liebminger, E.; Glössl, J.; Altmann, F.; Steinkellner, H.; Mach, L. Enzymatic properties and subcellular localization of Arabidopsis β-N-acetylhexosaminidases. Plant Physiol. 2007, 145, 5–16. [Google Scholar] [CrossRef]
- Dupoiron, S.; Zischeck, C.; Ligat, L.; Carbonne, J.; Boulanger, A.; de Bernonville, T.D.; Lautier, M.; Rival, P.; Jamet, E.; Lauber, E.; et al. The N-glycan cluster from Xanthomonas campestris pv. campestris: A toolbox for sequential plant N-glycan processing. J. Biol. Chem. 2015, 290, 6022–6036. [Google Scholar]
- Renzi, F.; Manfredi, P.; Mally, M.; Moes, S.; Jenö, P.; Cornelis, G. The N-glycan glycoprotein deglycosylation complex (Gpd) from Capnocytophaga canimorsus deglycosylates human IgG. PLoS Pathog. 2011, 7, 1002118. [Google Scholar] [CrossRef] [Green Version]
- Kasturi, L.; Eshleman, J.; Wunner, W.; Shakin-Eshleman, S. The hydroxy amino acid in an Asn-X-Ser/Thr sequon can influence N-linked core glycosylation efficiency and the level of expression of a cell surface glycoprotein. J. Biol. Chem. 1995, 270, 14756–14761. [Google Scholar] [CrossRef] [Green Version]
- Mellquist, J.; Kasturi, L.; Spitalnik, S.; Shakin-Eshleman, S. The amino acid following an Asn-X-Ser/Thr sequon is an important determinant of N-linked core glycosylation efficiency. Biochemistry 1998, 37, 6833–6837. [Google Scholar] [CrossRef] [PubMed]
- Shakin-Eshleman, S.; Spitalnik, S.; Kasturi, L. The amino acid at the X position of an Asn-X-Ser sequon is an important determinant of N-linked core-glycosylation efficiency. J. Biol. Chem. 1996, 271, 6363–6366. [Google Scholar] [CrossRef] [Green Version]
- Häweker, H.; Rips, S.; Koiwa, H.; Salomon, S.; Saijo, Y.; Chinchilla, D.; Robatzek, S.; von Schaewen, A. Pattern recognition receptors require N-glycosylation to mediate plant immunity. J. Biol. Chem. 2010, 285, 4629–4636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tams, J.; Vind, J.; Welinder, K. Adapting protein solubility by glycosylation.: N-Glycosylation mutants of Coprinus cinereus peroxidase in salt and organic solutions. Biochim. Biophys. Acta 1999, 1432, 214–221. [Google Scholar] [CrossRef]
- Francoz, E.; Ranocha, P.; Nguyen-Kim, H.; Jamet, E.; Burlat, V.; Dunand, C. Roles of cell wall peroxidases in plant development. Phytochemistry 2015, 112, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Lige, B.; Shengwu, M.; van Huystee, R. The effects of the site-directed removal of N-glycosylation from cationic peanut peroxidase on its function. Arch. Biochem. Biophys. 2001, 386, 17–24. [Google Scholar] [CrossRef]
- Wan, L.; van Huystee, R. A study on glycosylation of peanut cationic peroxidase. Biochem. Biophys. Res. Commun. 1993, 194, 1398–1405. [Google Scholar] [CrossRef]
- Vain, T.; Faris Crowell, E.; Timpano, H.; Biot, E.; Desprez, T.; Mansoori, N.; Trindade, L.; Pagant, S.; Robert, S.; Höfte, H.; et al. The cellulase KORRIGAN is part of the cellulose synthase complex. Plant Physiol. 2014, 165, 1521–1532. [Google Scholar] [CrossRef]
- Liebminger, E.; Grass, J.; Altmann, F.; Mach, L.; Strasser, R. Characterizing the link between glycosylation state and enzymatic activity of the endo-β1,4-glucanase KORRIGAN1 from Arabidopsis thaliana. J. Biol. Chem. 2013, 288, 22270–22280. [Google Scholar] [CrossRef] [Green Version]
- Rips, S.; Bentley, N.; Jeong, I.; Welch, J.; von Schaewen, A.; Koiwa, H. Multiple N-glycans cooperate in the subcellular targeting and functioning of Arabidopsis KORRIGAN1. Plant Cell 2014, 26, 3792–3808. [Google Scholar] [CrossRef] [Green Version]
- Marín-Menguiano, M.; Moreno-Sánchez, I.; Barrales, R.; Fernández-Álvarez, A.; Ibeas, J. N-glycosylation of the protein disulfide isomerase Pdi1 ensures full Ustilago maydis virulence. PLoS Pathog. 2019, 15, e1007687. [Google Scholar] [CrossRef] [Green Version]
- Strasser, R.; Stadlmann, J.; Schähs, M.; Stiegler, G.; Quendler, H.; Mach, L.; Glössl, J.; Weterings, K.; Pabst, M.; Steinkellner, H. Generation of glyco-engineered Nicotiana benthamiana for the production of monoclonal antibodies with a homogeneous human-like N-glycan structure Plant Biotechnol. J. 2008, 6, 392–402. [Google Scholar]
- Shin, Y.; König-Beihammer, J.; Vavra, U.; Schwestka, J.; Kienzl, N.; Klausberger, M.; Laurent, E.; Grünwald-Gruber, C.; Vierlinger, K.; Hofner, M.; et al. N-glycosylation of the SARS-CoV-2 receptor binding domain is important for functional expression in plants. Front. Plant Sci. 2021, 12, 689104. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lectin | Specificity a | Reference |
|---|---|---|
| Concanavalin A (ConA) | Man | [25,27] |
| branched, terminal α-D-Man | ||
| terminal β-D-Glc terminal N-GlcNAc | ||
| α-1,2-oligoMan | ||
| Wheat Germ Agglutinin (WGA) | GlcNAc polymer of GlcNAc Sia | [27,28] |
| NeuNAc | ||
| Galanthus nivalis Agglutinin (GNA) | terminal α-D-Man | [27,29,30] |
| α-D-Man-1,3-α-D-Man | ||
| high Man N-glycan | ||
| Lens culinaris agglutinin (LCH) | terminal α-D-Man terminal α-D-Glc complex (Man/N- GlcNAc core with α-1,6 Fuc) | [27] |
| Plant/Green Algae Species | Organ | Strategy 1 | Size 2 | Predicted CWPs 3 | Reference |
|---|---|---|---|---|---|
| Marchantia polymorpha | thallus 4 | 3 | 249 (92.0%) | 221 (88.8%) | [52] |
| Zea mays | seedling leaf | 5/7 | 476 (100%) | 307 (64.5%) | [34] |
| Triticum aestivum | seedling leaf | 6/8 | 312 (100%) | 236 (75.6%) | [33] |
| Brachypodium distachyon | seedling leaf | 5/7 | 35 (100%) | 28 (80.0%) | [53] |
| Fagopyrum tataricum | seed | 5/8 | 285 (100%) | nd | [38] |
| Solanum lycopersicum | fruit (pericarp) 4 | 5/8 | 363 (96.7%) | 202 (55.6%) | [37] |
| fruit (pericarp) 4 | 1 | 108 (97.2%) | 101 (93.5%) | [54] | |
| Gossypium hirsutum | seed(fiber cells) 4 | 1 | 199 (91.5%) | 114 (57.3%) | [55] |
| Camellia sinensis | leaf 4 | 5/8 | 382 (97.9%) | 267 (69.9%) | [56] |
| Arabidopsis thaliana | hypocotyl (etiolated) 4 | 3 | 127 (91.3%) | 123 (96.9%) | [24] |
| seedling | 5/8 | 912 (nd) | nd | [59] | |
| seedling/stem/floret | 5/8 | 538 (100%) | 343(64%) | [43] | |
| seedling and leaf4 | 9 | 173 (84.0%) | 135 (78.0%) | [36] | |
| leaf 4 | 3 | 62 (98.4%) | 58 (93.5%) | [57] | |
| stem 4 | 1 | 98 (100%) | 88 (89.8%) | [22] | |
| inflorescence 4 | 5/7 | 265 (96.6%) | 190 (71.7%) | [58] | |
| Brassica oleracea | xylem sap 4 | 2 | 75 (94.7%) | 74 (98.7%) | [23] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
San Clemente, H.; Jamet, E. N-glycoproteins in Plant Cell Walls: A Survey. Plants 2022, 11, 3204. https://doi.org/10.3390/plants11233204
San Clemente H, Jamet E. N-glycoproteins in Plant Cell Walls: A Survey. Plants. 2022; 11(23):3204. https://doi.org/10.3390/plants11233204
Chicago/Turabian StyleSan Clemente, Hélène, and Elisabeth Jamet. 2022. "N-glycoproteins in Plant Cell Walls: A Survey" Plants 11, no. 23: 3204. https://doi.org/10.3390/plants11233204
APA StyleSan Clemente, H., & Jamet, E. (2022). N-glycoproteins in Plant Cell Walls: A Survey. Plants, 11(23), 3204. https://doi.org/10.3390/plants11233204