1. Introduction
Nitrogen fertilization is important for optimizing short-term crop yield [
1], as nitrogen availability is a key factor controlling soil carbon cycling and storage [
2]. The organic carbon storage rate of soil in maize fields under continuous cropping increases with the increase in the nitrogen application amount, reaching a maximum value at the optimal nitrogen application level [
1]. However, excessive nitrogen application has been shown to affect the composition and quantity of organic carbon and nitrogen in the drylands of temperate continental climate, thereby altering soil nitrogen availability [
3]. Maize yield response to nitrogen is related to the amount of irrigation [
4]. Under a water deficit, the amount of nitrogen required for maximum yield decreases [
4].
Water is essential for agriculture in many world regions and for achieving sustainability in production systems. Water scarcity is seriously affecting agricultural production, especially in arid and semi-arid areas. Compared with the traditional method of rough broadcast fertilizer under strip-tillage and hop irrigation by flooding the inter-row, drip irrigation can deliver fertilizers and water to the roots of crops more accurately and reduce the amount of fertilizer required, minimizing environmental risks due to deep leakage of soil moisture and nutrients [
5]. The drip irrigation method is known to have better water use as well as fertilizer use efficiency as compared to other methods of irrigation, particularly if poor quality water is to be used. Shallow buried droppers are a kind of subsurface drip irrigation technology in which the drip pipe is buried 3–5 cm above the surface of the ground without a film covering. Shallow burial drip irrigation with low irrigation frequency increases the moisture volume of soil; however, it does not significantly increase the amount of water seepage under plant roots [
6]. Nevertheless, it is a new water-saving and high-yield technology. It was selected in 2021 as the principal agricultural technology in China, with a cumulative promotion area of more than 2 M ha. It can also reduce water consumption and improve the utilization efficiency of irrigation water [
7]. Additionally, it can compensate for the lack of drainage facilities and solve the problems faced by salt-affected farmlands [
8] as it does not add to soil salinity [
6]. Maize yields show no negative effects at drip irrigation levels of up to 75% compared with full irrigation [
9].
Further comparison with full irrigation has revealed that drip irrigation saves up to 25% of water [
9], whereas fractional nitrogen application improves the efficiency of nitrogen fertilizer and irrigation water usage [
10,
11]. The full irrigation with the subsurface drip irrigation system maximizes potato yield but decreases irrigation water use efficiency, whereas integration of the subsurface drip irrigation system with deficit irrigation is effective in improving water productivity due to less water being consumed, allowing these practices to be used under scarce water conditions [
12]. Maximum nitrogen use efficiency was obtained with good quality irrigation treatment [
13]. Deficit drip irrigation based on crop evapotranspiration and precipitation forecast is beneficial to improve crop water use efficiency and maintain high grain yield in semi-arid and semi-humid regions [
14]. While the effects of irrigation treatments on average dry matter contents of the lines were not found to be significant, significant differences were observed in water use efficiency and kernel yield of dent corn lines [
15]. Calcium chloride activated carbonized biogas fermentation residues can adsorb phosphorus. Phosphorus captured by this sorbent is readily available for plant nutrition [
16].Wood biochar shows excellent results in increasing the amount of plant-available water content in soil and appears to be an excellent tool for recycling nutrients (especially into plant-available forms of phosphorus and nitrogen) [
17].Therefore, calcium chloride activated carbonization of fermentation residues and biochar [
18] were used as novel fertilizers in drip irrigation. Earlier studies have focused on water and nutrient usage efficiency through microbial diversity indicators to better understand the future direction of soil function [
19]. Soil microorganisms are an important part of farmland ecosystems as well as the driving force for the decomposition, transformation, and circulation of organic matter. Soil nitrogen availability may be mediated by changes in microbial community composition to modulate the microbial response to precipitation changes [
20], affecting microbial biomass dynamics [
2]. Furthermore, the microbial and biochemical properties of soil differ under different nitrogen concentrations and water conditions [
21]. Excessive nitrogen application has been shown to significantly increase the nitrogen content of soil microbial biomass in the 0–20 cm and 20–40 cm soil layers of northwest drylands [
3].
Furthermore, excessive application of chemical fertilizers can deteriorate the microbial properties and biochemical functions of the soil [
22]. Moreover, microbial communities have a higher alpha diversity under nitrogen deficiency stress [
23]. N has significant, strong effects on bacterial, fungal, and functional community compositions; the relative abundance of most bacterial nitrogen cycling genes is increased or unaffected by N. In contrast, N decreases or does not change the expression of most bacterial carbon degradation genes [
24]. Bacteria and fungi respond differently to organic and inorganic fertilizers [
25]. Actinomycetes and Gram-positive bacteria can utilize soluble organic nitrogen more efficiently, whereas nitrogen fertilizers are more efficiently utilized by Gram-negative bacteria [
26]. Fertilization changes, especially long-term, can considerably affect the community structure, population, and function. Similarly, soil moisture conditions affect organic matter composition, aeration status, and microbial activity, influencing the mineralization and organic carbon mineralization rate [
21]. Soil moisture correlates with the C and N metabolic potential of the bacterial and archaeal community. Moderate deficit irrigation increases water productivity without affecting microbial communities [
27]. In addition, deficit irrigation increases the extracellular polysaccharide to microbial biomass ratios [
28]. Soil microorganisms provide essential nutrients for crop growth. Their biomass and diversity are sensitive to changes in soil nutrients, organic matter, and pH and are potential indicators of soil quality. Neither excessive nor insufficient nitrogen application and irrigation amounts are conducive to the reproduction of aerobic voluntary nitrogen-fixing bacteria [
29]. The increased precipitation in typical semi-arid areas of inner Mongolia grassland plays an important role in enhancing microbial activity. Increasing precipitation can alleviate the effect of nitrogen on microbial community composition [
30]. Water replenishment and nitrogen application significantly affect the activity of soil microbial communities in artificial grasslands [
31]. Watering alters the stress tolerance of desert scrub soil microorganisms, and fertilization alters the nutrient/oligotrophic properties of the microbial community [
32]. Carbon source and nitrogen source have the same biodegradability [
33]. Fertilization changes the availability of nutrients in agricultural raw materials. The combined effects of irrigation and fertilization on farmland soil bacterial and fungal communities are less studied.
However, the effect of water–nitrogen coupling on soil microbial diversity and activity is complex. Although knowledge regarding the effects of a single factor on soil microbial communities is growing rapidly, not much is known about the interactive effects of these two environmental change factors. In this study, high-throughput sequencing was performed using the Illumina Mi Seq platform to understand the composition and species richness of bacterial and fungal communities. We aimed to reveal the composition characteristics and changes in functional microbial flora in soil under the combined effect of different water conditions and nitrogen concentrations during shallow burial drip irrigation. Understanding the relationship between environmental factors might help clarify the coupling effect of the microbial and chemical properties of soil, providing a theoretical basis for constructing a high-yield and high-quality ecological environment.
This paper utilizes lots of acronym names, all the abbreviations used and their full names to create a list (
Table 1).
3. Discussion
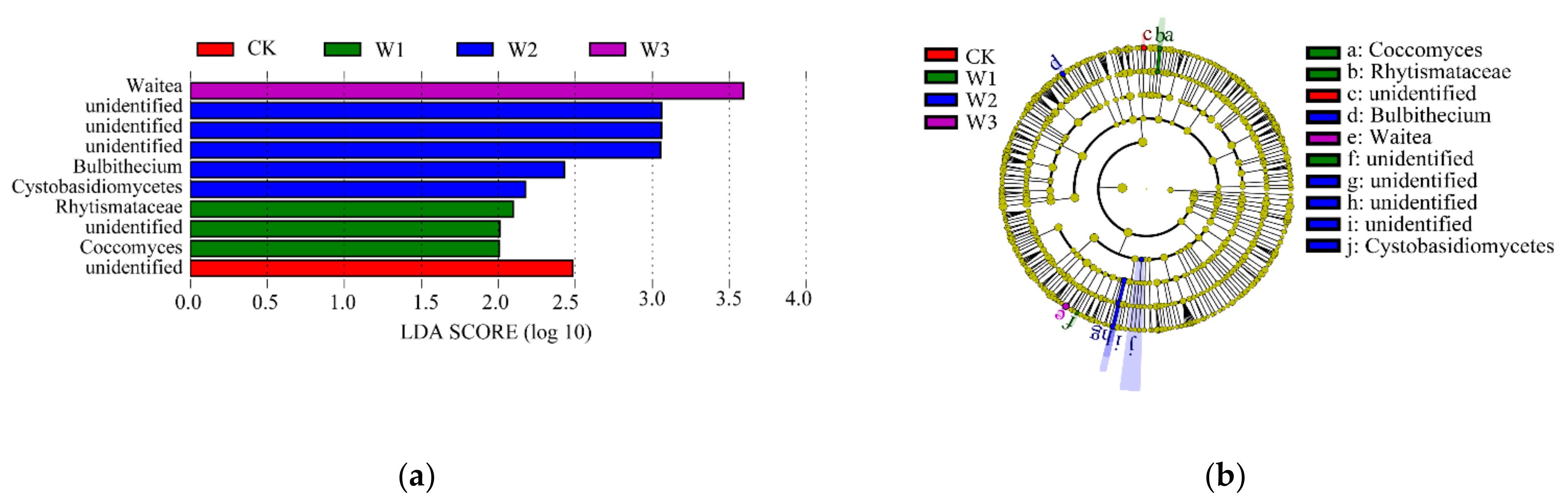
Soil microbial characteristics are regulated by irrigation and are closely related to the cyclic transformation of soil C and N nutrients [
35]. An irrigation quota of 900 m
3·ha
−1 in the Hetao Irrigation area was found to be the most conducive to the reproduction of bacteria and actinomycetes in saline soil, whereas a quota of 1125 m
3·ha
−1 was the best suited for fungal reproduction [
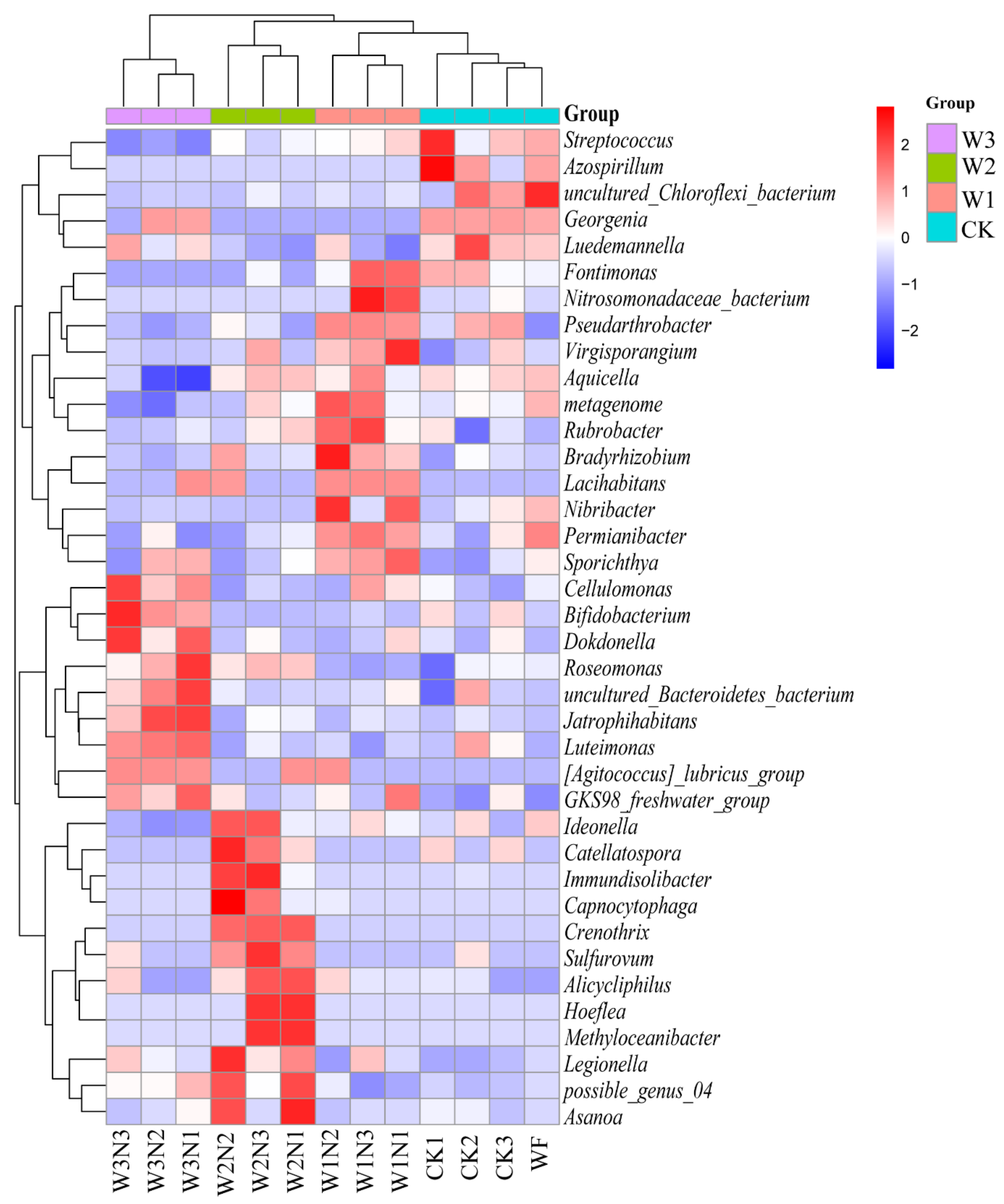
36]. Thus, under high water content conditions, the Shannon, Ace, and Chao1 indices of the bacterial community in the vegetable fields increased significantly. With the increase in soil water content, the relative abundance of
Aeromonas and
Flavobacterium also increases [
37]. In the present study, under high nitrogen conditions, the relative abundance of
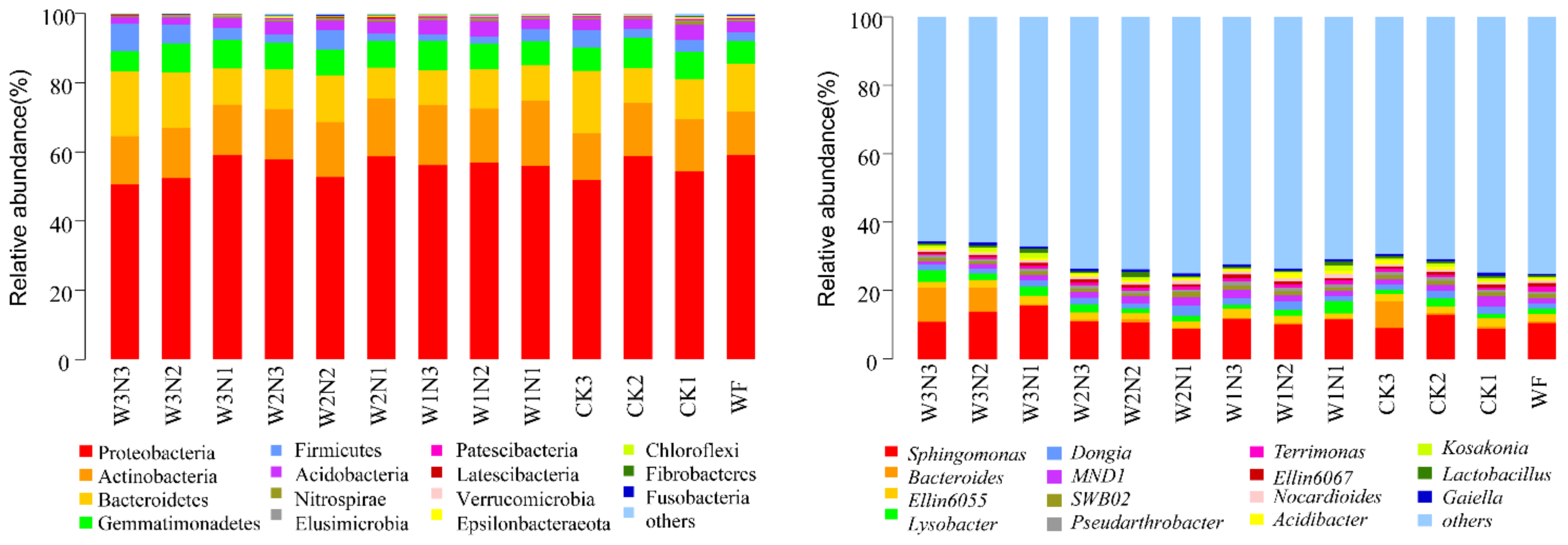
Bacteroides,
Lysobacter, and
Ellin6055 increased with an increase in irrigation amount, and the nitrogen reduction treatment of shallow burial drip irrigation decreased the relative abundance of
Sphingomonas. In the study by Yan et al. [
38], ammonification bacteria significantly increased, and N-fixing bacteria significantly decreased with precipitation reduction. Dinev et al. [
39] showed that fertilization has the strongest multiplication effect on the number of aerobic mesophilic microorganisms in the soil, whereas irrigation has no statistically significant impact. In the study by Jiao et al. [
40], with the increase in soil moisture, the dominant bacterial species shifted from Actinobacteria to Alphaproteobacteria and Acidobacteria. The same pattern was observed in the low nitrogen (N1) group in this study; however, in the high (N3) and middle nitrogen (N2) groups, the trend was different; that is, the relative abundance of Acidobacteria decreased with the increase of irrigation amount in the high nitrogen (N3) group and was more stable in the medium nitrogen (N3) group; low soil moisture content significantly reduced enzyme activity and bacterial richness [
41]. The composition of the bacterial community varied with soil moisture, but the fungal community was more resistant to water stress and acquired labile C more efficiently under low moisture levels [
42]. In the study by Jiao et al. [
40], there was no significant difference in Shannon diversity, relative abundance, and community structure of fungi under different soil water conditions. In addition, bacteria were more affected by stepwise wetting processes than fungi, which is consistent with the results of this study.
Fertilization affects the function and composition of the microbial community [
43]. When the optimum nitrogen application amount for maize in the Hetao Irrigation Area of Inner Mongolia was 280.11 k·ha
−1, soil microorganisms peaked, and the increasing effect was the most significant [
44]. If N is present in conventional bare-field maize fertilizers, the effects of P and K on soil microorganisms are not evident [
45], and a single application of nitrogen and phosphorus in maize fields in black soil areas cannot improve the diversity, uniformity, and soil microbial communities of soil bacteria and fungi. However, the composition of soil bacterial and fungal communities changed, with changes in fungal communities being more significant [
46]. N fertilization increased the relative abundance of the typical copiotrophic bacterial taxa, Alphaproteobacteria, but reduced that of the oligotrophic group, Acidobacteria. N fertilization stimulated most C transformation and N cycling processes [
47]. In this study, the relative abundance of Acidobacteria decreased with the increase of the N application rate under high irrigation (W3) but increased with increasing the N application rate under low irrigation (W1). Yang et al. [
48] selected an eco-friendly nitrogen (N) application level for sugarcane production, showing that soil bacterial richness could be significantly promoted by the medium (482 kg ha
−1) and high nitrogen (964 kg ha
−1) treatments. However, soil bacterial diversity could not be significantly improved. On the contrary, soil bacterial diversity and richness could be improved by low nitrogen (96 kg ha
−1) treatment. N-fertilized soil to reduce subsequent N supply (RCN) did not affect bacterial diversity, whereas RCN altered the community structure by enriching beneficial taxa, such as Actinobacteria (at the phylum level), and Streptomyces, Kribbella, Gaiella, Gemmatimonas (at the genus level), etc. The complexity and connectivity of the bacterial co-occurrence network were enhanced by RCN [
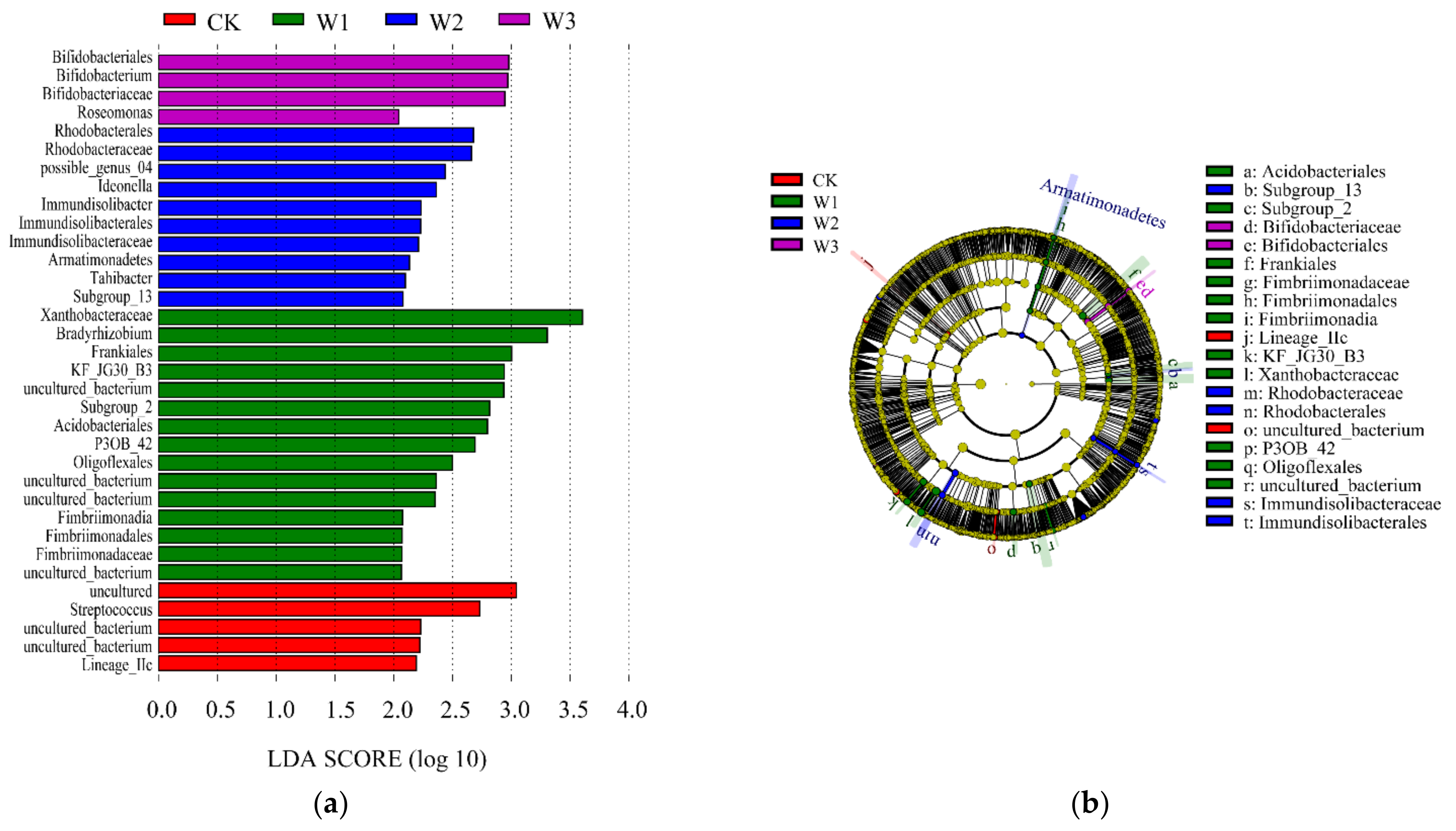
49]. In this study, W2N2 treatment enriched Glomeromycota, nitrogen-fixing bacteria Rhodanobacter and Bradyrhizobium, which make up the arbuscular mycorrhiza of terrestrial plants. In the present study, the effect of nitrogen on fungi also exceeded that on bacteria.
Fertilizer reduction treatment can increase the scale of fungal ecological network and community interaction, with a relatively close synergistic relationship between fungal communities [
50]. A single application of chemical fertilizer can significantly increase soil microbial biomass carbon [
51]. The response of soil microorganisms to water-saving and nitrogen-reducing agricultural measures is related to the complex interaction between water and nitrogen as well as between various groups in the food web [
52]. The application of P and K in maize fields promotes the number of soil bacteria, and the application of inorganic fertilizers changes the community structure of soil microorganisms [
53]. A single application of nitrogen fertilizer, co-application of nitrogen-potassium-phosphorus fertilizer, and co-application of potassium and phosphorus fertilizers increase the relative abundance of soil actinomycetes and fungi during the maize growth period [
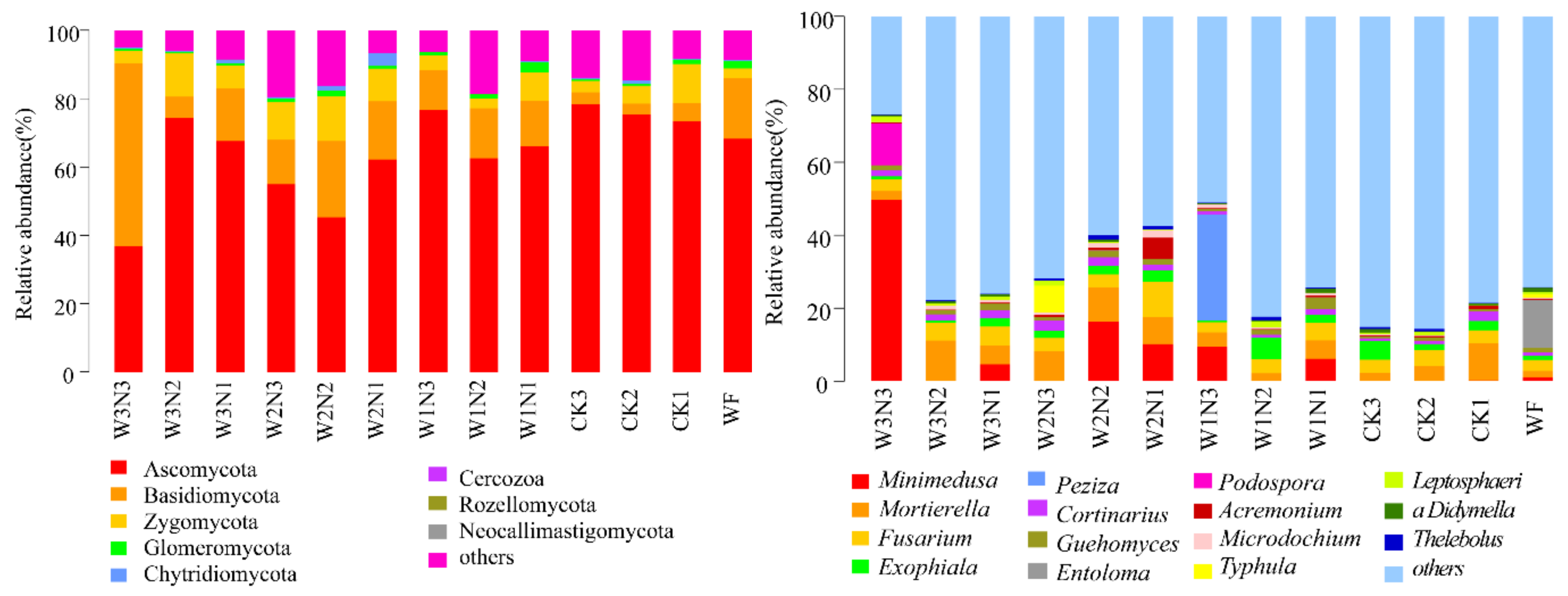
53]. In the present study, the number of bacterial OTUs in shallow burial drip irrigation without fertilizer was the least, and the application of nitrogen, phosphorus, and potassium fertilizers or the application of potassium and phosphorus fertilizers both increased the number of bacterial OTUs. The application of phosphorus and potassium fertilizers increased the relative abundance of Bacteroidetes and decreased the relative abundance of Proteobacteria. The relative abundance of Actinobacteria and Gemmatimonadetes did not change, that of Proteobacteria and Gemmatimonadetes increased, and that of Bacteroidetes decreased after nitrogen application. In this study, soil fungi were mainly distributed in Ascomycota, Basidiomycota, and Zygomycota. The application of phosphorus and potassium fertilizers increased the relative abundance of Ascomycota and decreased that of Basidiomycota but did not affect the relative abundance of Zygomycota. Nitrogen application increased the relative abundance of Basidiomycota and Zygomycota and decreased that of Ascomycota. NPK fertilizers increased the relative abundance of Ascomycota and Zygomycota but decreased that of Basidiomycota. It was observed that the effect of phosphorus and potassium fertilizers on Ascomycota and Basidiomycota was greater than that of nitrogen fertilizers, and the effect of nitrogen fertilizers on the relative abundance of Zygomycota was greater than that of phosphorus and potassium fertilizers. Optimal nitrogen application reduced the fungal richness and diversity, decreasing bacterial richness [
23]. The relative abundances of nitrogen-fixing bacteria
Rhodanobacter and
Bradyrhizobium related to the nitrogen cycle in the optimized nitrogen application treatment were higher than those in conventional nitrogen application treatment [
54]. In the present study, different species in the 50% and 60% conventional irrigation groups and others also belonged to these two genera. Excessive or insufficient nitrogen application and irrigation amounts were not conducive to the reproduction of aerobic autogenic nitrogen-fixing bacteria.
The effect of the nitrogen application amount on the number of anaerobic authigenic nitrogen-fixing bacteria is irregular [
29]. Only one genus,
Luedemannella, has been identified under high water and high nitrogen treatment [
55], and only this genus appeared in the N3 assemblage in the present study. Our previous study showed that optimal nitrogen topdressing (70% constant nitrogen topdressing) under shallow burial drip irrigation reduces carbon and nitrogen emissions from farmland ecosystems in the West Liaohe Plain, improving the carbon efficiency and effective use of nitrogen inputs [
56]. Under natural precipitation, when the irrigation amount is 1958.40–2228.00 m
3·ha
−1, the nitrogen application amount is 209.34–275.70 kg·ha
−1, the density is 67,350–78,150 plants·ha
−1, and the yield can reach 12,000.00–12,716.82 kg·ha
−1 [
57]. There are numerous potential bacterial candidates that could be recruited to assist plants during water-limiting conditions [
58]. In the present study, with a decrease in the nitrogen application amount, compared with the irrigation amounts of 1600 and 2400 m
3·ha
−1, the relative abundance of Actinobacteria and Gemmatimonadetes under the irrigation amount of 2000 m
3·ha
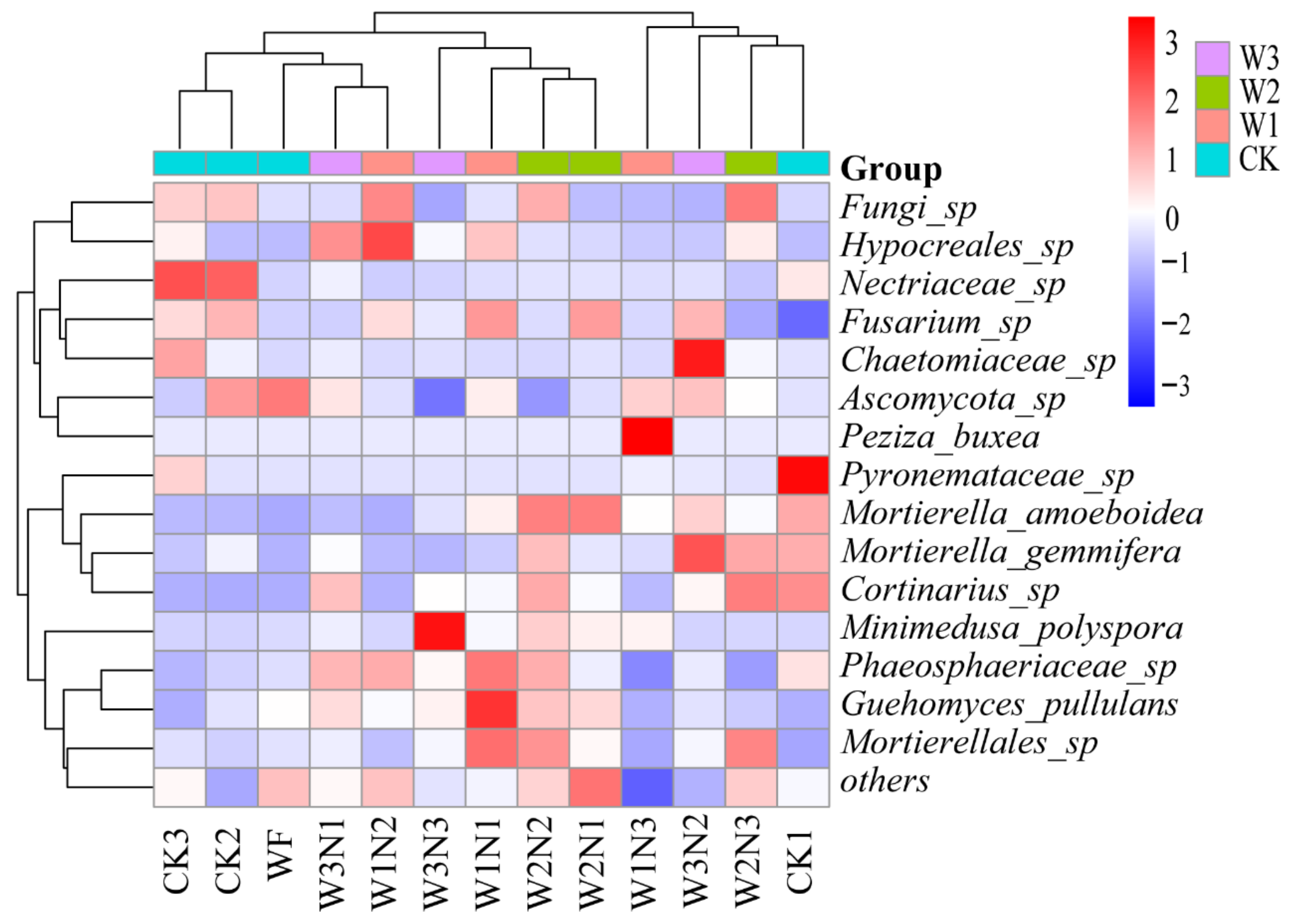
−1 was more stable under high nitrogen conditions. The relative abundances of Proteobacteria, Firmicutes, and Actinobacteria did not change with low-nitrogen treatment, and the relative abundance of Basidiomycota, Zygomycota, and Glomeromycota bacteria was the highest in W2N2 (210 kg·ha
−1 nitrogen application) treatment. The relative abundance of
Minimedusa,
Mortierella,
Guehomyces, and
Thelebolus fungi was the lowest in W2N2; bacteria and fungi had strong antagonistic effects in topsoil. Competition and environmental filtration both affect the abundance, composition, and encoding of gene functions of bacterial and fungal communities. Of note, soil bacteria are more resistant to mineral fertilization interference than fungi [
43]. Moreover, the interactions between organisms can change the microbial community [
59].
Soil water and nitrogen content are the main drivers of microbial community structure [
60]. Previous studies have shown that high water and normal nitrogen treatments render the bacterial community more uniform in greenhouse soil; however, they do not improve diversity [
61]. With increased precipitation and nitrogen addition, the relative abundance of fungi significantly decreases [
30]. Changes in the soil moisture content affect the richness of the soil fungal community in the desert steppe, whereas water and nitrogen treatments exert no notable effect on fungal community diversity [
34]. Nitrogen addition weakened the effect of water addition in terms of soil bacterial diversity and community stability, and did not have an interactive influence [
62]. Research by Guo et al. [
63] shows that N addition, reduced precipitation and their combined effect significantly altered the soil fungal community composition. Soil microbial biomass and composition were more strongly affected by nitrogen fertilization compared with water regime. Nitrogen fertilization increased soil microbial biomass and altered soil microbial community composition, especially under low soil water availability [
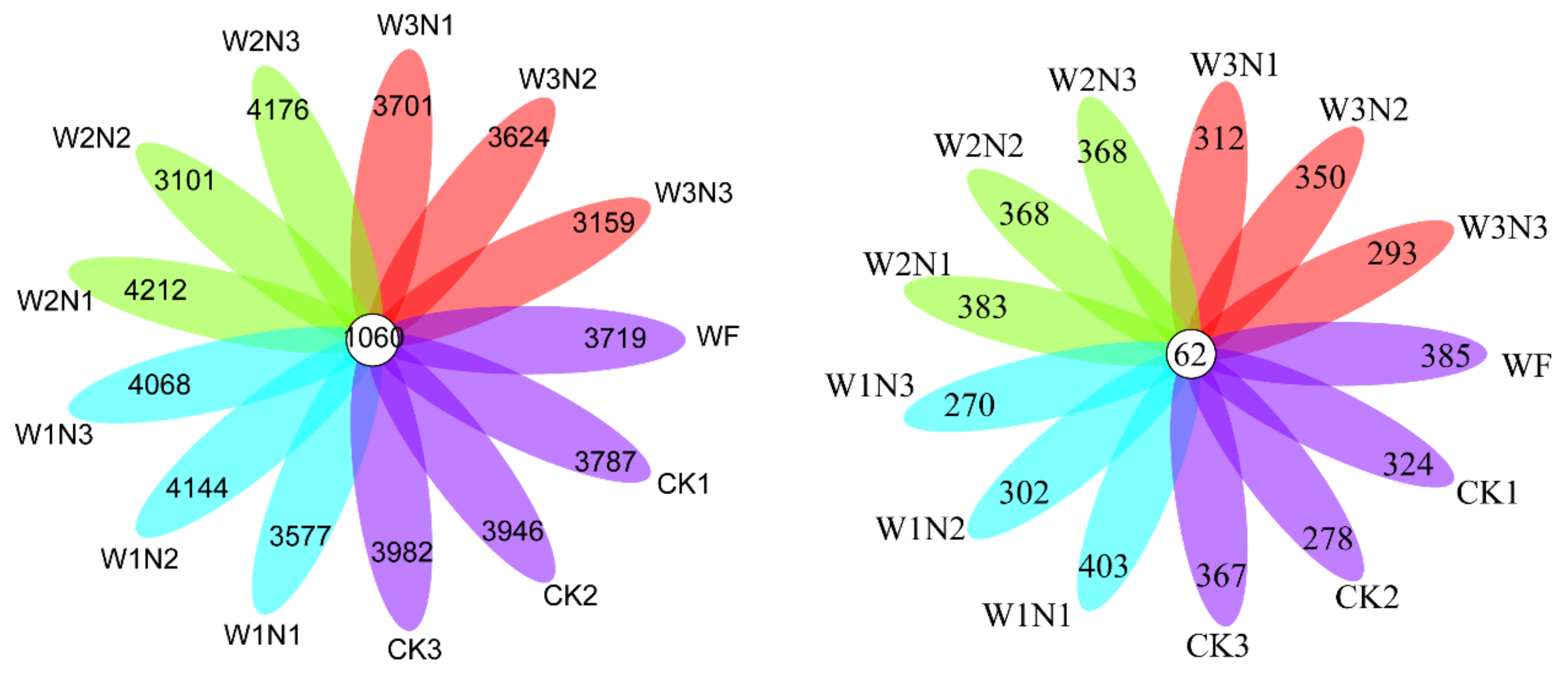
64]. In the present study, higher water volume irrigation in water-saving irrigation decreased soil bacterial and fungal diversity. The drip irrigation quota can affect the response of soil microorganisms to nitrogen. At 50% and 60% conventional irrigation amounts, the relative abundance of Actinobacteria was stable despite the decrease in the nitrogen application amount. In comparison, the relative abundance of Actinobacteria decreased at the 40% conventional irrigation amount. The relative abundance of Gemmatimonadetes increased with the decrease in the nitrogen application amount under the 60% conventional irrigation amount, whereas the relative abundance of Gemmatimonadetes was stable under the 50% conventional irrigation amount but decreased under the 40% conventional irrigation amount. With decrease in the nitrogen application amount, the relative abundance of Proteobacteria increased at 60% but was stable at the 40% conventional irrigation amount. Furthermore, with a decreased nitrogen application amount, the relative abundance of Bacteroidetes was relatively higher. The abundance decreased and exceeded that in N1 and N3 at the 40% and 50% conventional irrigation amounts.
Previous studies have shown that the significant spatial distribution of soil nutrients affects the composition of microbial communities [
65]. The relative abundance of soil bacteria and fungi may be related to changes in soil chemical composition [
66]. Liu et al. [
67] showed that Zygomycota, Glomeromycota, and Chytridiomycota possessed strongly positive associations with available potassium and available phosphate, whereas Ascomycota showed a strong negative association. Plant growth-promoting bacteria, such as the genus
Sphingomonas and families Rhizobiaceae and Micrococcaceae, are potentially associated with soil quality [
68]. The total bacterial and diazotrophic population significantly positively correlated with the available NPK and organic carbon in the soil at each growth stage [
69]. In the present study, the relative abundance of
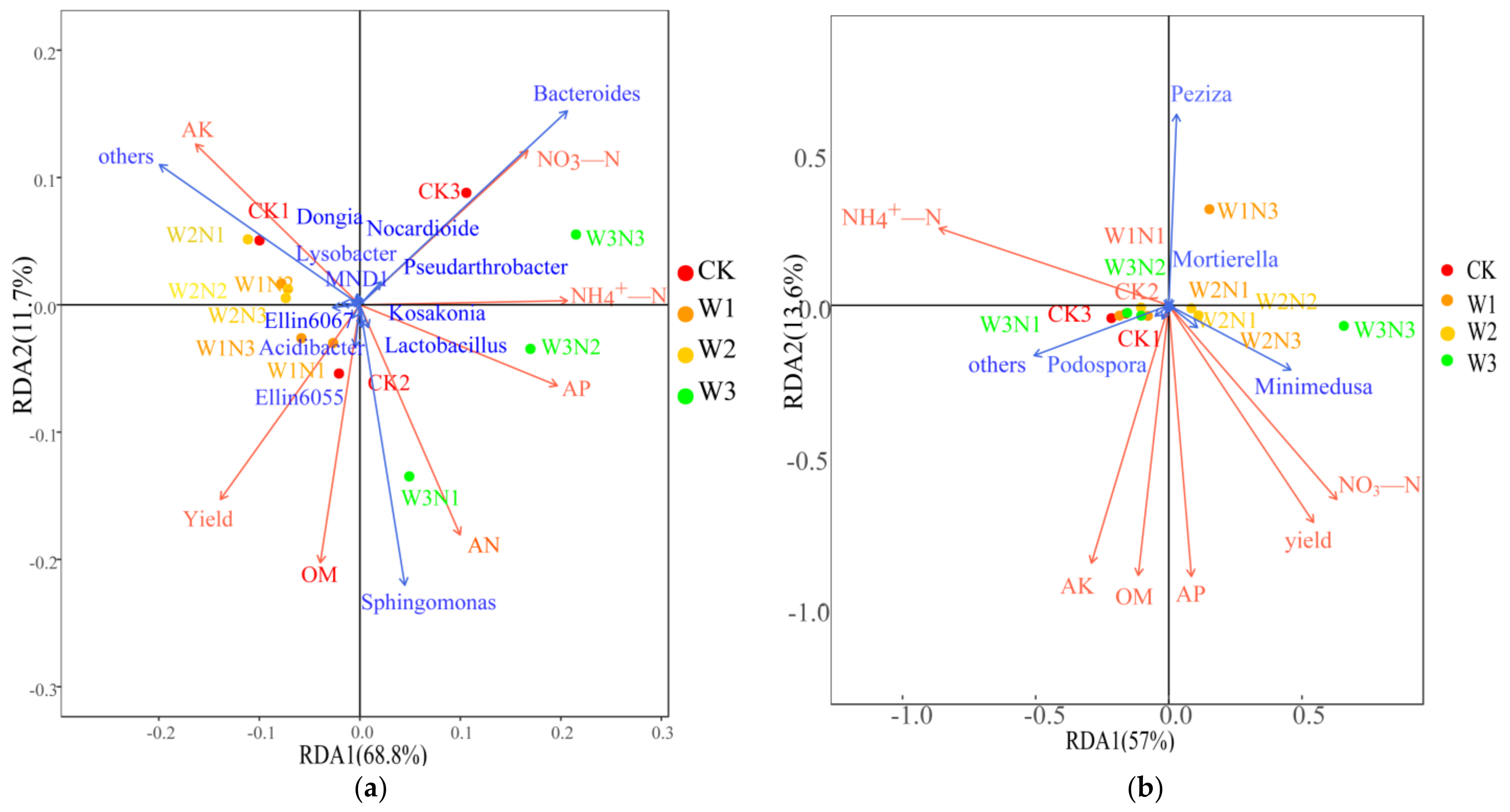
Sphingomonas,
Bacteroides,
Ellin6055, and
Minimedusa was positively correlated with AP. The relative abundance of
Sphingomonas and
Minimedusa positively correlated with yield. The relative abundance of
Sphingomonas, Ellin6055, and
Peziza inversely correlated with NO
3-N. This could be attributed to the complex effects of environmental disturbances and plants on soil microbial communities. In different studies, soil nutrients received varying degrees of disturbance, which in turn, differentially affected the composition of microbial communities; hence, the correlation between microorganisms and soil nutrients was different.
In this study, the effect of irrigation amount on bacterial community composition was greater than that of nitrogen, whereas the effect of nitrogen on fungi exceeded that on bacteria. The relative abundance of Bacteroidetes and Firmicutes decreased in the irrigation volume of 2400 m
3·ha
−1, and the relative abundance of Glomeromycota, which can constitute arbuscular mycorrhizae of terrestrial plants, decreased. Chytridiomycota disappeared in the irrigation volume of 1600 m
3·ha
−1. The irrigation volume of 2000 m
3·ha
−1 and the nitrogen application amount of 210 kg·ha
−1 enhanced the ability to maintain the balance of the soil microbial community. The soil water distribution formed by shallow burial drip irrigation changes the nitrogen cycle and microbial composition, affecting the biomass of nitrifying bacteria and denitrifying bacteria. Thus, the interaction among “root system–soil–microbes” and shallow burial drip irrigation root zone environment is conducive to enhancing the interaction of “roots–soil–microbes” [
70], thereby improving the carbon efficiency and effective use of nitrogen input, ultimately increasing maize yield. Under water-saving and nitrogen-reducing drip irrigation, the relationships between dominant flora and the synergistic mode of soil fungal and bacterial communities warrant further research.
The water-saving and economic benefits of shallow-buried drip irrigation were improved significantly, and the water use efficiency and benefits were increased. In the aspect of ecological benefit, after the implementation of the shallow-buried drip irrigation project, the irrigation water quantity is greatly reduced, the productivity of irrigation water is increased, and the groundwater depth is in a relatively stable state, which ensures the ecological benefit. Considering water-saving, economic and ecological benefits, the comprehensive benefits of shallow-buried drip irrigation are higher than that of traditional border irrigation. The comprehensive performance of shallow-buried drip irrigation planting pattern is better in the aspects of seedling protection effect, seed quality, yield per unit area, cost input, pure benefit, etc., the water-saving planting technology model of shallow-buried drip irrigation is characterized by saving labor, time, water, investment and pollution, which can replace the semi-film drip irrigation widely used at present, it is suitable to be popularized in a semi-arid area (
Table 5).
In the follow-up study, soil metabolomics technology will be used to analyze the different metabolites in soil under the condition of water saving and nitrogen reduction, and combined microbial-metabolite analysis will be used, the relationship between soil microorganism, soil metabolite and plant will be further verified.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}