
The Streptomyces scabiei Pathogenicity Factor Thaxtomin A Induces the Production of Phenolic Compounds in Potato Tubers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
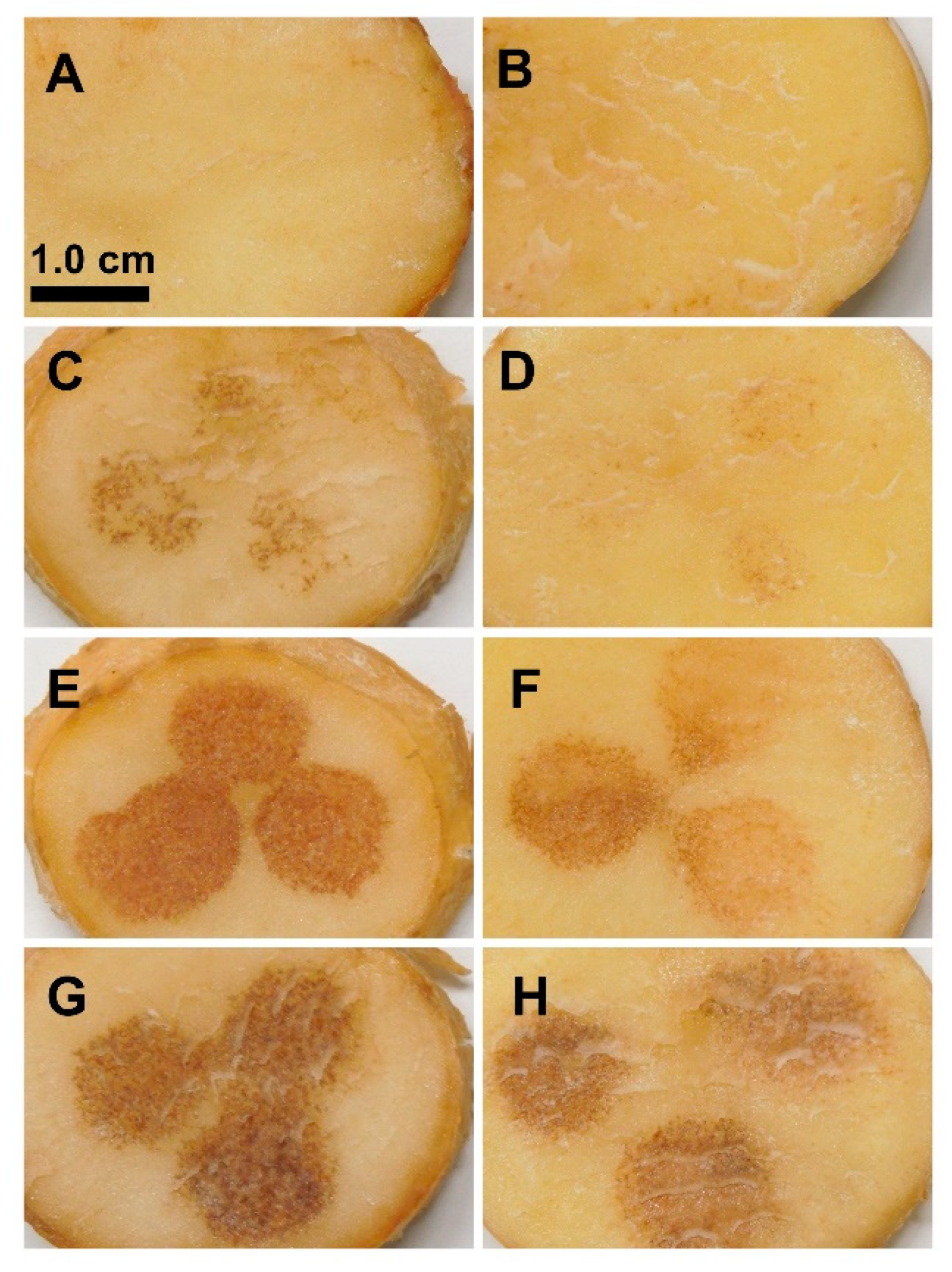
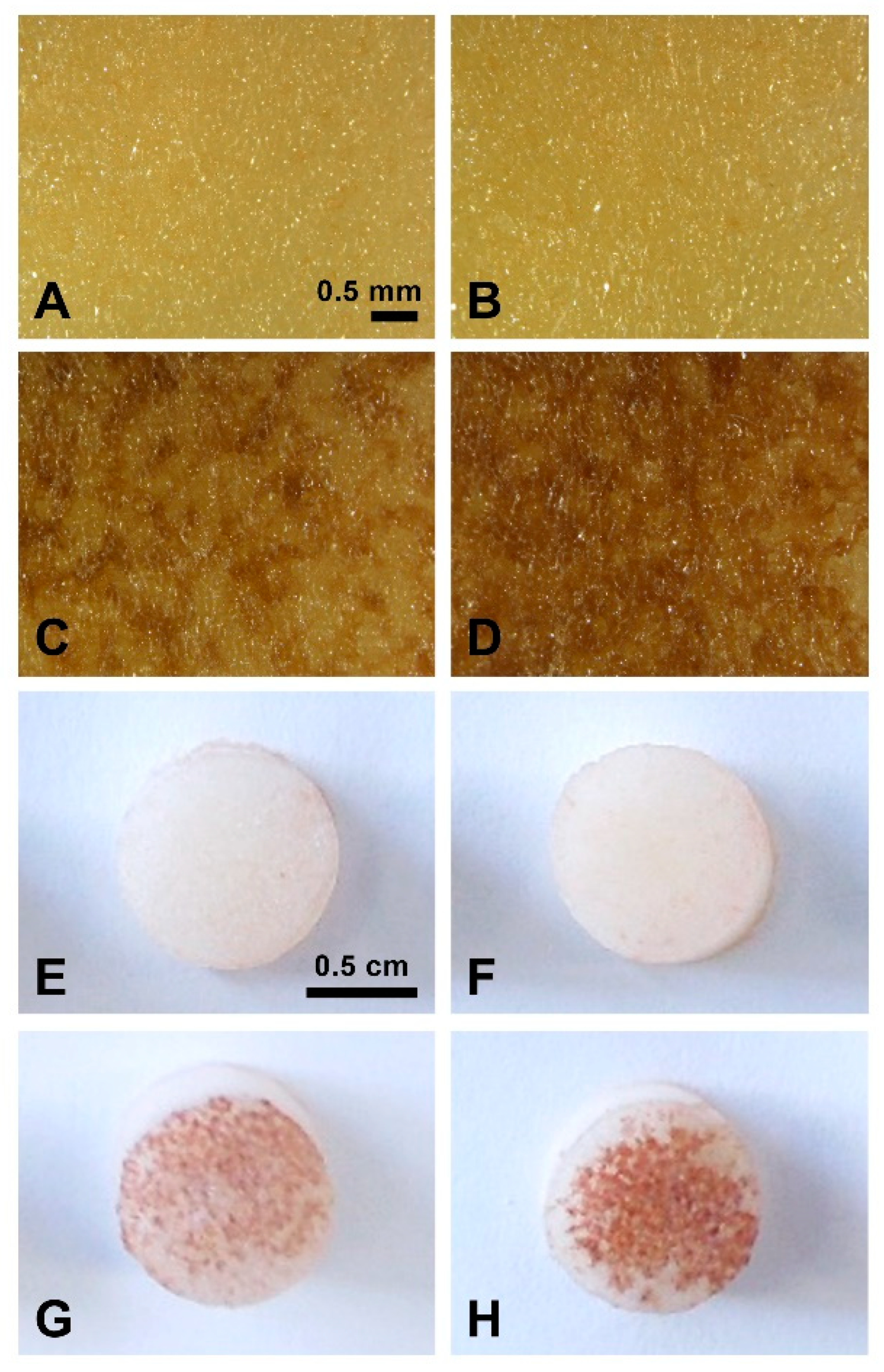
2.1. TA Induces Browning on Potato Tuber Slices
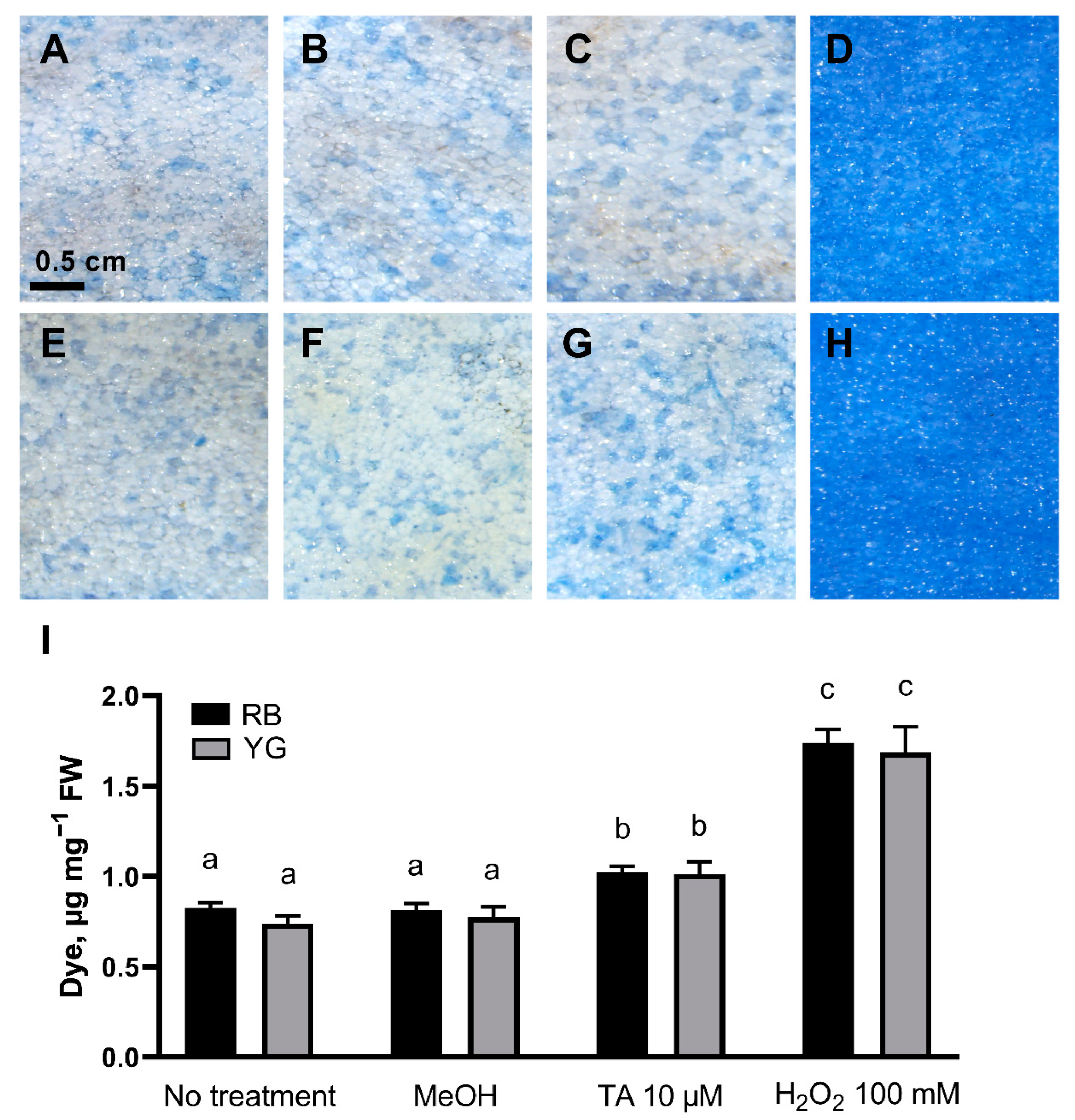
2.2. TA Induces Cell Death in Potato Tuber Parenchyma
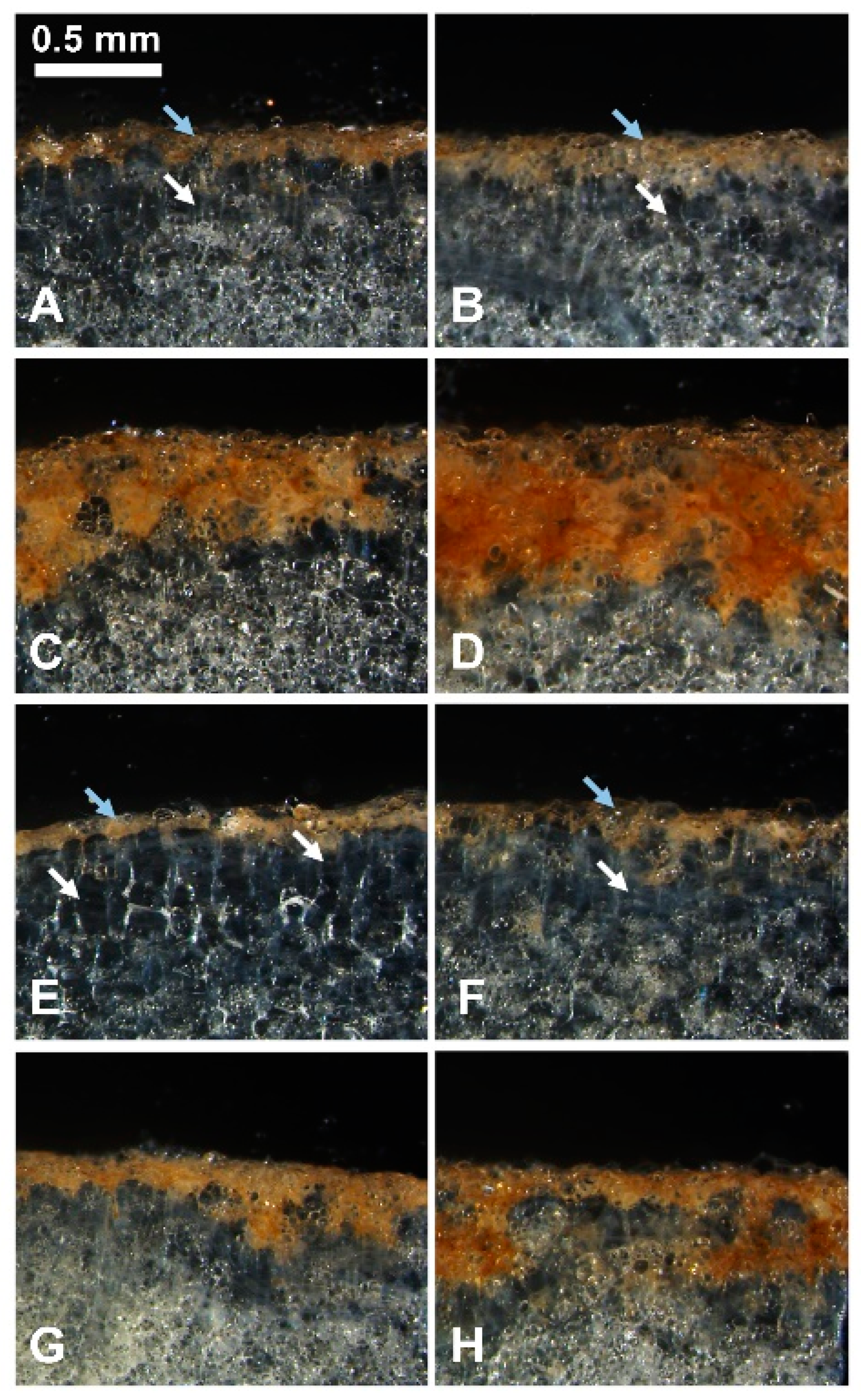
2.3. TA Affects the Formation of the Wound Periderm
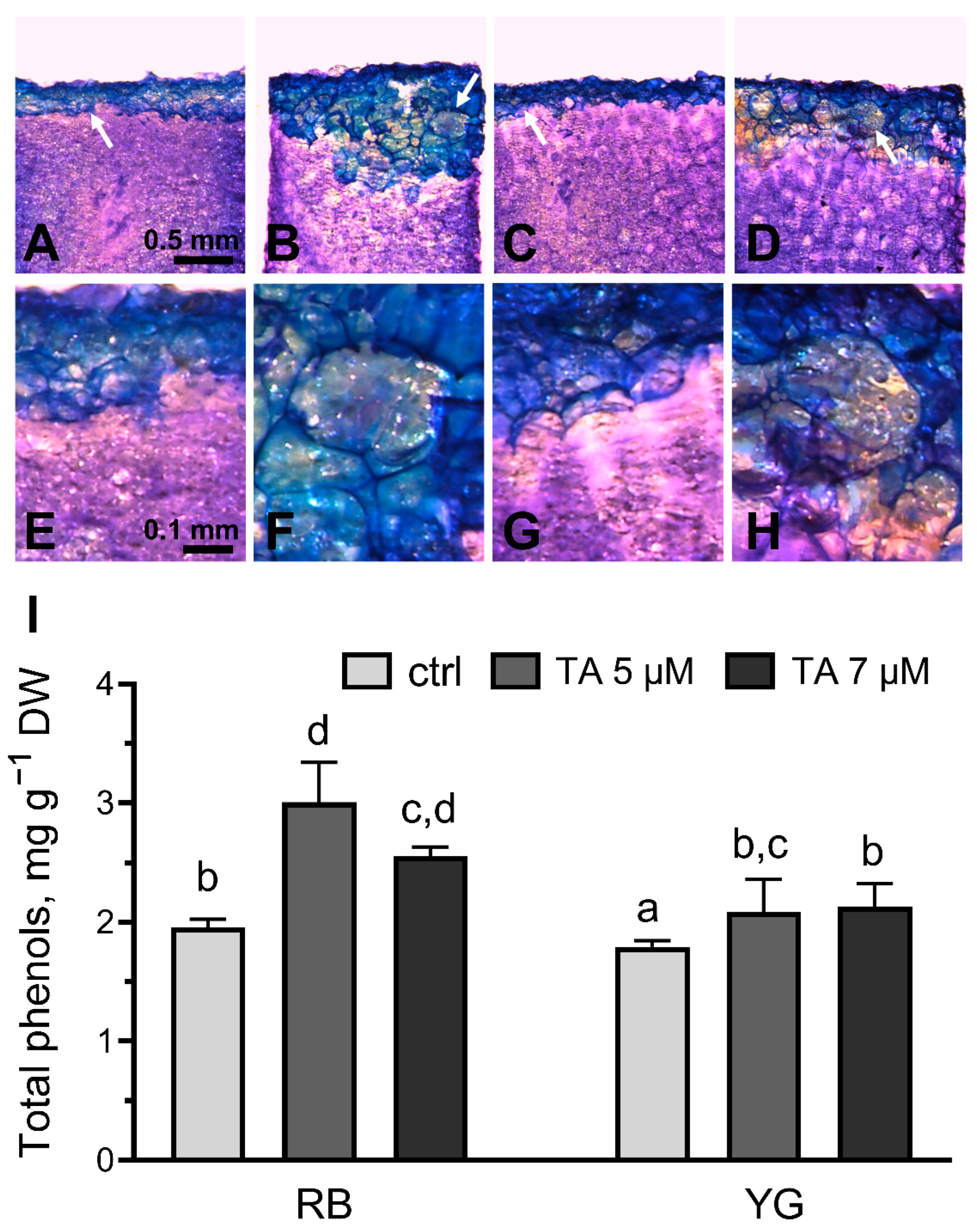
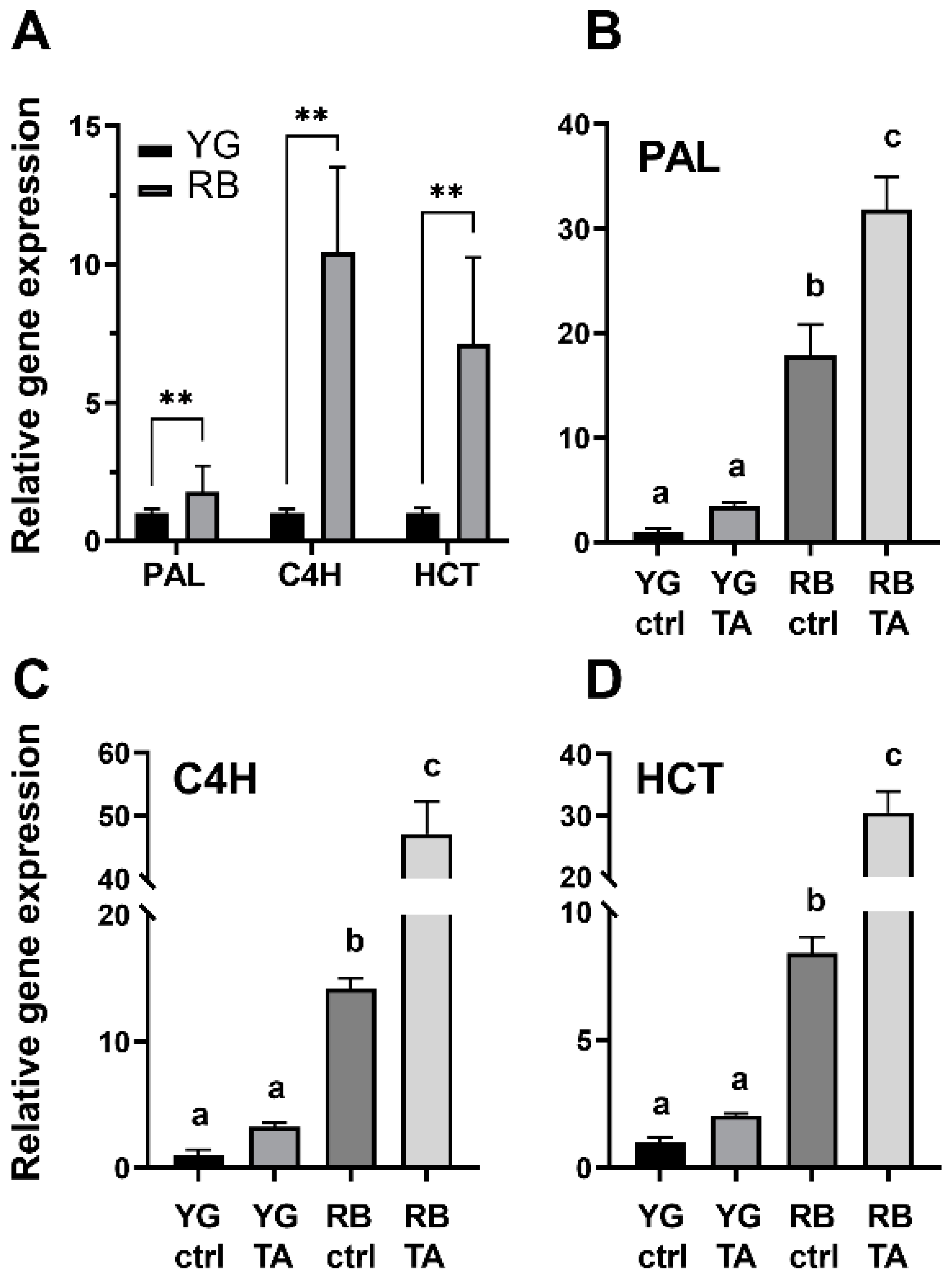
2.4. Phenolic Compounds Deposition and Synthesis in Response to TA
2.5. Inhibition of PAL Activity Reduces TA-Induced Browning
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material
4.3. TA Extraction and Purification
4.4. TA Test on Tuber Slices
4.5. Microscopy Analysis of TA-Induced Symptoms on Potato Parenchyma Cells
4.6. Toluidine Blue O Staining of Phenolics Deposited in Potato Tuber Parenchyma Cells
4.7. TA-Induced Cell Death Determination
4.8. Total Phenols Extraction and Determination
4.9. RNA Extraction and Gene Expression Analysis
4.10. Inhibition of Phenylalanine Ammonia-Lyase Activity
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dees, M.W.; Wanner, L.A. In search of better management of potato common scab. Potato Res. 2012, 55, 249–268. [Google Scholar] [CrossRef]
- Goyer, C.; Vachon, J.; Beaulieu, C. Pathogenicity of Streptomyces scabies mutants altered in thaxtomin A production. Phytopathology 1998, 88, 442–445. [Google Scholar] [CrossRef] [Green Version]
- Joshi, M.V.; Bignell, D.R.D.; Johnson, E.G.; Sparks, J.P.; Gibson, D.M.; Loria, R. The AraC/XylS regulator TxtR modulates thaxtomin biosynthesis and virulence in Streptomyces scabies. Mol. Microbiol. 2007, 66, 633–642. [Google Scholar] [CrossRef]
- Francis, I.M.; Jourdan, S.; Fanara, S.; Loria, R.; Rigali, S. The cellobiose sensor CebR is the gatekeeper of Streptomyces scabies pathogenicity. mBio 2015, 6, e02018. [Google Scholar] [CrossRef] [Green Version]
- King, R.R.; Lawrence, C.H.; Gray, J.A. Herbicidal properties of the thaxtomin group of phytotoxins. J. Agric. Food Chem. 2001, 49, 2298–2301. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.A.; Loria, R. Thaxtomin A: Evidence for a plant cell wall target. Physiol. Mol. 2002, 60, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Scheible, W.-R.; Fry, B.; Kochevenko, A.; Schindelasch, D.; Zimmerli, L.; Somerville, S.; Loria, R.; Somerville, C.R. An Arabidopsis mutant resistant to thaxtomin A, a cellulose synthesis inhibitor from Streptomyces species. Plant Cell 2003, 15, 1781–1794. [Google Scholar] [CrossRef] [Green Version]
- Duval, I.; Brochu, V.; Simard, M.; Beaulieu, C.; Beaudoin, N. Thaxtomin A induces programmed cell death in Arabidopsis thaliana suspension-cultured cells. Planta 2005, 222, 820–831. [Google Scholar] [CrossRef]
- Tegg, R.S.; Melian, L.; Wilson, C.R.; Shabala, S. Plant cell growth and ion flux responses to the Streptomycete phytotoxin thaxtomin A: Calcium and hydrogen flux patterns revealed by the non-invasive MIFE technique. Plant Cell. Physiol. 2005, 46, 638–648. [Google Scholar] [CrossRef]
- Errakhi, R.; Dauphin, A.; Meimoun, P.; Lehner, A.; Reboutier, D.; Vatsa, P.; Briand, J.; Madiona, K.; Rona, J.P.; Barakate, M.; et al. An early Ca2+ influx is a prerequisite to thaxtomin A-induced cell death in Arabidopsis thaliana cells. J. Exp. Bot. 2008, 59, 4259–4270. [Google Scholar] [CrossRef] [PubMed]
- Joglekar, S.; Suliman, M.; Bartsch, M.; Halder, V.; Maintz, J.; Bautor, J.; Zeier, J.; Parker, J.E.; Kombrink, E. Chemical activation of EDS1/PAD4 signaling leading to pathogen resistance in Arabidopsis. Plant Cell Physiol. 2018, 59, 1592–1607. [Google Scholar] [CrossRef] [Green Version]
- Bischoff, V.; Cookson, S.J.; Wu, S.; Scheible, W.-R. Thaxtomin A affects CESA-complex density, expression of cell wall genes, cell wall composition, and causes ectopic lignification in Arabidopsis thaliana seedlings. J. Exp. Bot. 2009, 60, 955–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerat, S.; Babana, A.H.; El Oirdi, M.; El Hadrami, A.; Daayf, F.; Beaudoin, N.; Bouarab, K.; Beaulieu, C. Streptomyces scabiei and its toxin thaxtomin A induce scopoletin biosynthesis in tobacco and Arabidopsis thaliana. Plant Cell Rep. 2009, 28, 1895–1903. [Google Scholar] [CrossRef]
- Wilson, C.R.; Tegg, R.S.; Wilson, A.J.; Luckman, G.A.; Eyles, A.; Yuan, Z.Q.; Hingston, L.H.; Conner, A.J. Stable and extreme resistance to common scab of potato obtained through somatic cell selection. Phytopathology 2010, 100, 460–467. [Google Scholar] [CrossRef] [Green Version]
- Hiltunen, L.H.; Alanen, M.; Laakso, I.; Kangas, A.; Virtanen, E.; Valkonen, J.P.T. Elimination of common scab sensitive progeny from a potato breeding population using thaxtomin A as a selective agent. Plant Pathol 2011, 60, 426–435. [Google Scholar] [CrossRef]
- Beaudoin, N.; Isayenka, I.; Ducharme, A.; Massie, S.; Gagnon, A.; Hogue, R.; Beaulieu, C.; Michaud, D. Habituation to thaxtomin A increases resistance to common scab in ‘Russet Burbank’ potato. PLoS ONE 2021, 16, e0253414. [Google Scholar] [CrossRef]
- Tegg, R.S.; Gill, W.M.; Thompson, H.K.; Davies, N.W.; Ross, J.J.; Wilson, C.R. Auxin-induced resistance to common scab disease of potato linked to inhibition of thaxtomin A toxicity. Plant Dis. 2008, 92, 1321–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, C.R.; Luckman, G.A.; Tegg, R.S.; Yuan, Z.Q.; Wilson, A.J.; Eyles, A.; Conner, A.J. Enhanced resistance to common scab of potato through somatic cell selection in cv. Iwa with the phytotoxin thaxtomin A. Plant Pathol. 2009, 58, 137–144. [Google Scholar] [CrossRef]
- Hiltunen, L.H.; Laakso, I.; Chobot, V.; Hakala, K.S.; Weckman, A.; Valkonen, J.P.T. Influence of thaxtomins in different combinations and concentrations on growth of micropropagated potato shoot cultures. J. Agric. Food Chem. 2006, 54, 3372–3379. [Google Scholar] [CrossRef]
- Tegg, R.S.; Wilson, C.R. Relationship of resistance to common scab disease and tolerance to thaxtomin A toxicity within potato cultivars. Eur. J. Plant Pathol. 2010, 128, 143–148. [Google Scholar] [CrossRef]
- Loria, R.; Bukhalid, R.A.; Creath, R.A.; Leiner, R.H.; Olivier, M.; Steffens, J.C. Differential production of thaxtomins by pathogenic Streptomyces species in vitro. Phytopathology 1995, 85, 537–541. [Google Scholar] [CrossRef]
- Lawrence, H.C.; Clark, M.C.; King, R.R. Induction of common scab symptoms in aseptically cultured potato tubers by the vivotoxin, thaxtomin. Phytopathology 1990, 80, 606–608. [Google Scholar] [CrossRef]
- Ampomah, Y.A.; Friend, J. Insoluble phenolic compounds and resistance of potato tuber disc to Phytophthora and Phoma. Phytochemistry 1988, 27, 2533–2541. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; Espín, J.C. Phenolic compounds and related enzymes as determinants of quality in fruits and vegetables. J. Sci. Food. Agric. 2001, 81, 853–876. [Google Scholar] [CrossRef]
- Yadav, V.; Wang, Z.; Wei, C.; Amo, A.; Ahmed, B.; Yang, X.; Zhang, X. Phenylpropanoid pathway engineering: An emerging approach towards plant defense. Pathogens 2020, 9, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Zhang, A.; Zhao, J.; Shang, J.; Zhu, Z.; Wu, X.; Zha, D. Transcriptome profiling reveals potential genes involved in browning of fresh-cut eggplant (Solanum melongena L.). Sci. Rep. 2021, 11, 16081. [Google Scholar] [CrossRef]
- Brochu, V.; Girard-Martel, M.; Duval, I.; Lerat, S.; Grondin, G.; Domingue, O.; Beaulieu, C.; Beaudoin, N. Habituation to thaxtomin A in hybrid poplar cell suspensions provides enhanced and durable resistance to inhibitors of cellulose synthesis. BMC Plant Biol. 2010, 10, 272. [Google Scholar] [CrossRef] [Green Version]
- Thangavel, T.; Tegg, R.S.; Wilson, C.R. Toughing it out—Disease-resistant potato mutants have enhanced tuber skin defenses. Phytopathology 2016, 106, 474–483. [Google Scholar] [CrossRef] [Green Version]
- Jacyn Baker, C.; Mock, N.M. An improved method for monitoring cell death in cell suspension and leaf disc assays using Evans blue. Plant Cell Tiss. Organ Cult. 1994, 39, 7–12. [Google Scholar] [CrossRef]
- Vijayaraghavareddy, P.; Adhinarayanreddy, V.; Vemanna, R.S.; Sreeman, S.; Makarla, U. Quantification of membrane damage/cell death using Evan’s blue staining technique. Bio Protoc. 2017, 7, e2519. [Google Scholar] [CrossRef]
- Lulai, E.C.; Campbell, L.G.; Fugate, K.K.; McCue, K.F. Biological differences that distinguish the 2 major stages of wound healing in potato tubers. Plant Signal. Behav. 2016, 11, e1256531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, L.; Cai, Q.; Huang, W.; Dastmalchi, K.; Rigau, J.; Molinas, M.; Figueras, M.; Serra, O.; Stark, R.E. Potato native and wound periderms are differently affected by down-regulation of FHT, a suberin feruloyl transferase. Phytochemistry 2018, 147, 30–48. [Google Scholar] [CrossRef] [PubMed]
- Schoenwaelder, M.E.A.; Clayton, M.N. The presence of phenolic compounds in isolated cell walls of brown algae. Phycologia 1999, 38, 161–166. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valiñas, M.A.; Lanteri, M.L.; ten Have, A.; Andreu, A.B. Chlorogenic acid biosynthesis appears linked with suberin production in potato tuber (Solanum tuberosum). J. Agric. Food Chem. 2015, 63, 4902–4913. [Google Scholar] [CrossRef]
- Liu, S.; Qi, Q.; Chao, N.; Hou, J.; Rao, G.; Xie, J.; Lu, H.; Jiang, X.; Gai, Y. Overexpression of artificially fused bifunctional enzyme 4CL1–CCR: A method for production of secreted 4-hydroxycinnamaldehydes in Escherichia coli. Microb. Cell Fac. 2015, 14, 118. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.M.P.; Saxena, P.K. Inhibition of phenylpropanoid biosynthesis in Artemisia annua L.: A novel approach to reduce oxidative browning in plant tissue culture. PLoS ONE 2013, 8, e76802. [Google Scholar] [CrossRef] [Green Version]
- Loria, R.; Bukhalid, R.A.; Fry, B.A.; King, R.R. Plant pathogenicity in the genus Streptomyces. Plant Dis. 1997, 81, 836–846. [Google Scholar] [CrossRef] [Green Version]
- Sabba, R.P.; Lulai, E.C. Histological analysis of the maturation of native and wound periderm in potato (Solanum tuberosum L.) tuber. Ann. Bot. 2002, 90, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Vitti, M.C.D.; Sasaki, F.F.; Miguel, P.; Kluge, R.A.; Moretti, C.L. Activity of enzymes associated with the enzymatic browning of minimally processed potatoes. Braz. Arch. Biol. Technol. 2011, 54, 983–990. [Google Scholar] [CrossRef]
- Teoh, L.S.; Lasekan, O.; Adzahan, N.M.; Hashim, N. The effect of ultraviolet treatment on enzymatic activity and total phenolic content of minimally processed potato slices. J. Food Sci. Technol. 2016, 53, 3035–3042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernards, M.A.; Razem, F.A. The poly(phenolic) domain of potato suberin: A non-lignin cell wall bio-polymer. Phytochemistry 2001, 57, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- Graça, J. Suberin: The biopolyester at the frontier of plants. Front. Chem. 2015, 3, 62. [Google Scholar] [CrossRef]
- Akyol, H.; Riciputi, Y.; Capanoglu, E.; Caboni, M.F.; Verardo, V. Phenolic compounds in the potato and its byproducts: An overview. Int. J. Mol. Sci. 2016, 17, 835. [Google Scholar] [CrossRef] [PubMed]
- Hammerschmidt, R. Rapid deposition of lignin in potato tuber tissue as a response to fungi non-pathogenic on potato. Physiol. Plant Pathol. 1984, 24, 33–42. [Google Scholar] [CrossRef]
- Lattanzio, V.; Cardinali, A.; Palmieri, S. The role of phenolics in the postharvest physiology of fruits and vegetables: Browning reactions and fungal diseases. Ital. J. Food Sci. 1994, 6, 3–22. [Google Scholar]
- Johnson, G.; Schaal, L.A. Accumulation of phenolic substances and ascorbic acid in potato tuber tissue upon injury and their possible role in disease resistance. Am. Potato J. 1957, 34, 200. [Google Scholar] [CrossRef]
- Singhai, P.K.; Sarma, B.K.; Srivastava, J.S. Phenolic acid content in potato peel determines natural infection of common scab caused by Streptomyces spp. World J. Microbiol. Biotechnol. 2011, 27, 1559–1567. [Google Scholar] [CrossRef]
- Dixon, R.; Paiva, N. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Dixon, R.A.; Lamb, C.J.; Masoud, S.; Sewalt, V.J.H.; Paiva, N.L. Metabolic engineering: Prospects for crop improvement through the genetic manipulation of phenylpropanoid biosynthesis and defense responses—A review. Gene 1996, 179, 61–71. [Google Scholar] [CrossRef]
- Awwad, F.; Bertrand, G.; Grandbois, M.; Beaudoin, N. Auxin protects Arabidopsis thaliana cell suspension cultures from programmed cell death induced by the cellulose biosynthesis inhibitors thaxtomin A and isoxaben. BMC Plant Biol. 2019, 19, 512. [Google Scholar] [CrossRef]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef]
- Steenackers, W.; Klíma, P.; Quareshy, M.; Cesarino, I.; Kumpf, R.P.; Corneillie, S.; Araújo, P.; Viaene, T.; Goeminne, G.; Nowack, M.K.; et al. cis-cinnamic acid is a novel, natural auxin efflux inhibitor that promotes lateral root formation. Plant Physiol. 2017, 173, 552–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faucher, E.; Savard, T.; Beaulieu, C. Characterization of actinomycetes isolated from common scab lesions on potato tubers. Can. J. Plant Pathol. 1992, 14, 197–202. [Google Scholar] [CrossRef]
- Beisson, F.; Li, Y.; Bonaventure, G.; Pollard, M.; Ohlrogge, J.B. The acyltransferase GPAT5 is required for the synthesis of suberin in seed coat and root of Arabidopsis. Plant Cell 2007, 19, 351–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, G.M.; Iyer, S.; Knowles, N.R. Extraction of RNA from fresh, frozen, and lyophilized tuber and root tissues. J. Agric. Food Chem. 2007, 55, 1674–1678. [Google Scholar] [CrossRef]
- André, C.M.; Schafleitner, R.; Legay, S.; Lefèvre, I.; Aliaga, C.A.A.; Nomberto, G.; Hoffmann, L.; Hausman, J.-F.; Larondelle, Y.; Evers, D. Gene expression changes related to the production of phenolic compounds in potato tubers grown under drought stress. Phytochemistry 2009, 70, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Navarre, D.A.; Payyavula, R.S.; Shakya, R.; Knowles, N.R.; Pillai, S.S. Changes in potato phenylpropanoid metabolism during tuber development. Plant Physiol. Biochem. 2013, 65, 89–101. [Google Scholar] [CrossRef]
- Nicot, N.; Hausman, J.F.; Hoffmann, L.; Evers, D. Housekeeping gene selection for real-time RT-PCR normalization in potato during biotic and abiotic stress. J. Exp. Bot. 2005, 56, 2907–2914. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isayenka, I.; Beaudoin, N. The Streptomyces scabiei Pathogenicity Factor Thaxtomin A Induces the Production of Phenolic Compounds in Potato Tubers. Plants 2022, 11, 3216. https://doi.org/10.3390/plants11233216
Isayenka I, Beaudoin N. The Streptomyces scabiei Pathogenicity Factor Thaxtomin A Induces the Production of Phenolic Compounds in Potato Tubers. Plants. 2022; 11(23):3216. https://doi.org/10.3390/plants11233216
Chicago/Turabian StyleIsayenka, Iauhenia, and Nathalie Beaudoin. 2022. "The Streptomyces scabiei Pathogenicity Factor Thaxtomin A Induces the Production of Phenolic Compounds in Potato Tubers" Plants 11, no. 23: 3216. https://doi.org/10.3390/plants11233216
APA StyleIsayenka, I., & Beaudoin, N. (2022). The Streptomyces scabiei Pathogenicity Factor Thaxtomin A Induces the Production of Phenolic Compounds in Potato Tubers. Plants, 11(23), 3216. https://doi.org/10.3390/plants11233216