


Response Mechanisms of Adventitious Root Architectural Characteristics of Nitraria tangutorum Shrubs to Soil Nutrients in Nabkha

Abstract
:1. Introduction
2. Results
2.1. Morphological Characteristics of N. tangutorum Shrub
2.2. Spatial Heterogeneity of Soil Nutrients in N. tangutorum Nabkha
2.3. Morphological Characteristics of Adventitious Root System of N. tangutorum Nabkha
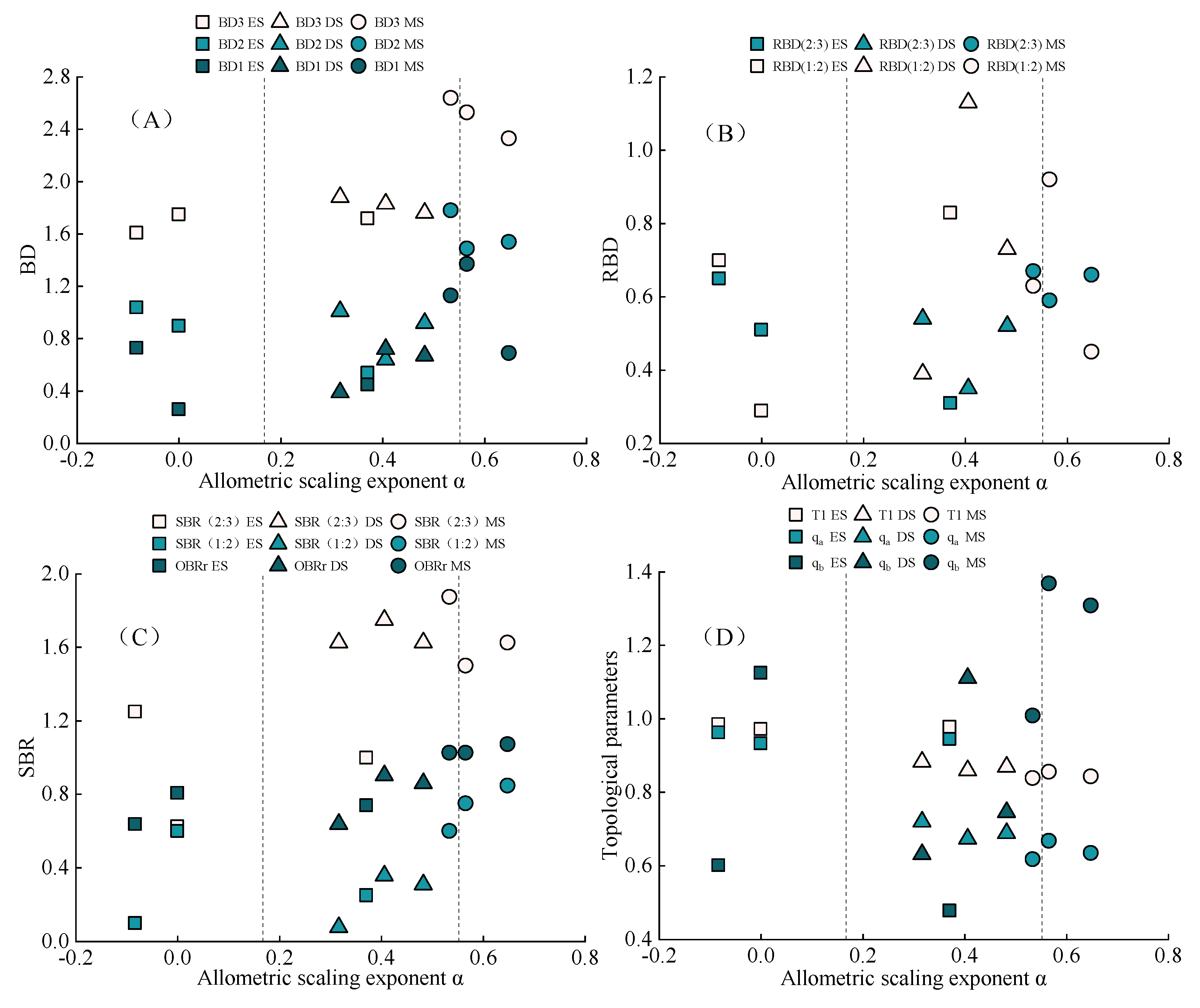
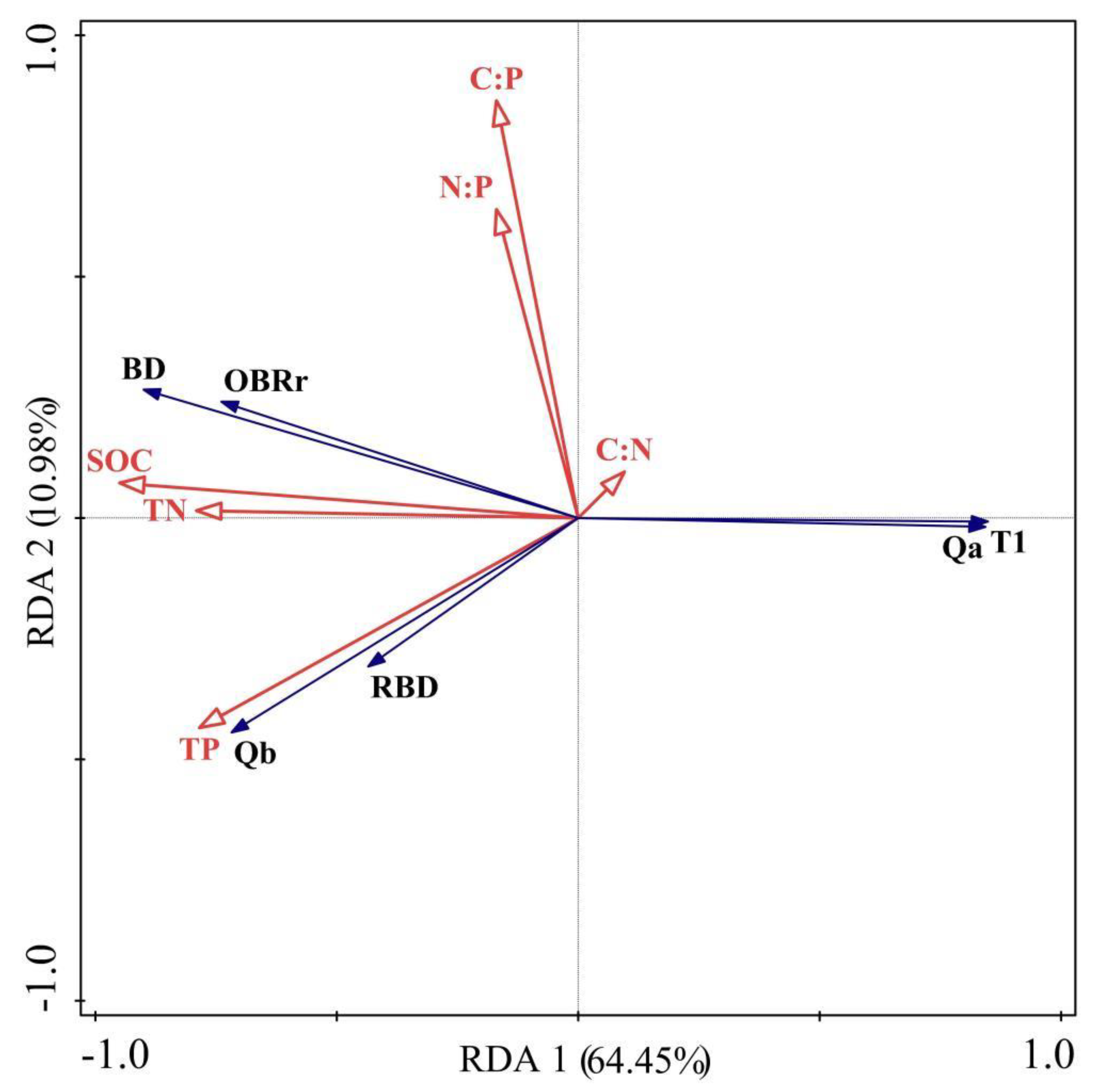
2.4. Response Mechanism of Adventitious Root Architecture of N. tangutorum to Soil Nutrients
3. Discussion
3.1. Growth Relationship between N. tangutorum Shrub and Nabkha in West Ordos
3.2. Spatial Heterogeneity of Soil Nutrients in N. tangutorum Nabkha
3.3. Response Mechanism of Adventitious Root Architecture to Soil Nutrients in Nabkha in Western Ordos
4. Materials and Methods
4.1. Study Area
4.2. Experimental Design and Methods
4.3. Parameter Calculation
4.3.1. Morphological Characteristic Parameters of Nabkha
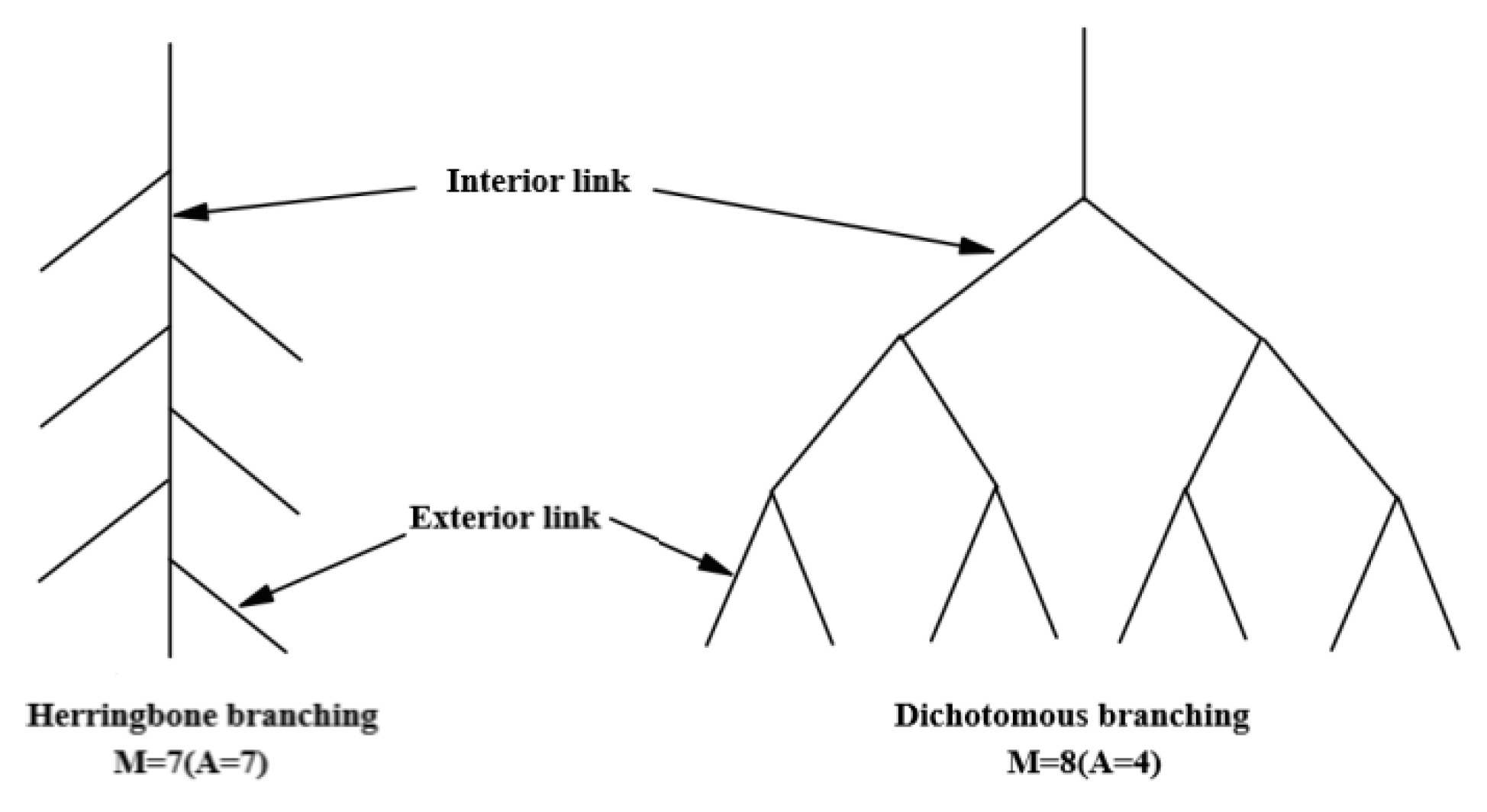
4.3.2. Adventitious Root Branching Rate
4.3.3. Root Diameter Ratio of Adventitious Roots
4.3.4. Topological Index
4.3.5. Allometric Mechanism Model
4.3.6. Soil Nutrient Accumulation Characteristics Calculation
4.4. Sample Determination
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yue, X.L.; Hasi; Zhuang, Y.M.; Zhang, J. Studies on Sandy Grassland Nebkhas-A Review. J. Desert Res. 2005, 25, 738–743. [Google Scholar]
- Li, J.; Zhao, Y.; Liu, H.; Su, Z. Sandy desertification cycles in the southwestern Mu Us Desert in China over the past 80 years recorded based on nebkha sediments. Aeolian Res. 2016, 20, 100–107. [Google Scholar] [CrossRef]
- Valiente-Banuet, A.; Bolongaro-Crevenna, A.; Briones, O.; Ezcurra, E.; Rosas, M.; Nuez, H.; Barnard, G.; Vazquez, E. Spatial relationships between cacti and nurse shrubs in a semi-arid environment in Central Mexico. J. Veg. Sci. 1991, 2, 15–20. [Google Scholar] [CrossRef]
- Ma, X.Z.; Wang, X.P. Root architecture and adaptive strategy of two desert plants in the Alxa Plateau. Acta Ecol. Sin. 2020, 40, 6001–6008. [Google Scholar]
- Wang, X.; Zhang, C.; Zhang, J.; Hua, T.; Lang, L.; Zhang, X.; Wang, L. Nebkha formation: Implications for reconstructing environmental changes over the past several centuries in the Ala Shan Plateau, China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 297, 697–706. [Google Scholar] [CrossRef]
- Wang, L.J.; Sun, D.Y.; Zhao, C.Y.; Li, J.Y.; Sheng, Y. Plant architecture characteristics of Haloxylon ammodendron and Haloxylon persicum in Zhungar Basin. Acta Ecol. Sin. 2011, 31, 4952–4960. [Google Scholar]
- Zhang, L.L.; Zhao, X.Y.; Yuan, H. Advances in the effects of wind on plants. Adv. Earth Sci. 2013, 28, 1349–1353. [Google Scholar]
- Wu, S.L.; Li, Z.Z.; Xiao, C.X.; Sun, Q.M.; Liu, L.M. Research Progress on Nabkhas and Research Significance. J. Desert Res. 2006, 26, 734–738. [Google Scholar]
- El-Bana, M.I.; Nijs, I.; Kockelbergh, F. Microenvironmental and vegetational heterogeneity induced by phytogenic nebkhas in an arid coastal ecosystem. Plant Soil. 2002, 247, 283–293. [Google Scholar] [CrossRef]
- Chang, Z.F.; Zhang, J.H.; Shi, X.G.; Wang, Q.; Zhang, D.K.; Duan, X.F. Initial research on the relationship between sand-mound formation and the layered silhouette of desert plants. Acta Ecol. Sin. 2017, 37, 7351–7358. [Google Scholar]
- Margitta, D.; Arthur, B. Fractal geometry and root system structures of heterogeneous plant communities. Plant Soil 2005, 272, 61–76. [Google Scholar]
- Jonathan, L. Root Architecture and Plant Productivity. Plant Physiol. 1995, 109, 7–13. [Google Scholar]
- Shan, L.S.; Li, Y.; Dong, Q.L.; Geng, D.M. Ecological adaptation of Reaumuria soongorica root system architecture to arid environment. J. Desert Res. 2012, 32, 1283–1290. [Google Scholar]
- Savinov, I.A. Architectural Analysis of Representatives of the Celastrales Order: Structure and Rhythm of Shoot Development in Connection with Adaptations of Species to Different Environmental Conditions. Contemp. Probl. Ecol. 2020, 13, 300–308. [Google Scholar] [CrossRef]
- Yang, X.L.; Zhang, X.M.; Li, Y.L.; Li, S.C.; Sun, H.L. Analysis of root architecture and root adaptive strategy in the Taklimakan desert area of China. Chin. J. Plant Ecol. 2008, 6, 1268–1276. [Google Scholar]
- Xiao, Y.; Tao, Y.; Zhang, Y.M. Biomass allocation and leaf stoichiometric characteristics in four desert herbaceous plants during different growth periods in the Gurbantünggüt Desert, China. J. Desert Res. 2014, 38, 929–940. [Google Scholar]
- Du, J.H.; Yan, P.; Dong, Y.X. The progress and prospects of nebkhas in arid areas. J. Geogr. Sci. 2010, 65, 339–350. [Google Scholar] [CrossRef]
- Albrecht, A.; Serigne, T.K. Carbon sequestration in tropical agroforestry systems. Agric. Ecosyst. Environ. 2003, 99, 15–27. [Google Scholar] [CrossRef]
- Li, X.R. Effects of soil spatial heterogeneity on vegetation restoration in arid sandy areas. Sci. Sin. 2005, 35, 361–370. [Google Scholar]
- Wei, Y.J.; Dang, X.H.; Wang, J.; Zhang, K.H.; Gao, Y.; Li, S.Z. Characterization of the soil ecological stoichiometry of Nitraria tangutorun nebkhas during different succession stages. J. Soil Water Conserv. 2021, 35, 377–384. [Google Scholar]
- Peichl, M.; Thevathasan, N.V.; Gordon, A.M.; Huss, J.; Abohassan, R.A. Carbon sequestration potentials in temperate tree-based inter-cropping systems, southern Ontario, Canada. Agrofor. Syst. 2006, 66, 243–257. [Google Scholar] [CrossRef]
- Li, X.L.; Wei, Y.J.; Dang, X.H.; Dai, Y.Z.; Zhai, B.; Chi, X.; Wu, H.M. Soil mechanical composition and soil nutrient content of Reaumuria soongorica nebkhas. Arid. Zone Res. 2022, 39, 933–942. [Google Scholar]
- Takimoto, A.; Nair, V.D.; Nair, P.K.R. Contribution of trees to soil carbon sequestration under agroforestry systems in the West African Sahel. Agrofor. Syst. 2009, 76, 11–25. [Google Scholar] [CrossRef]
- Hendrick, R.L.; Pregitzer, K.S. Spatial variation in tree root distribution and growth associated with minirhizotrons. Plant Soil 1992, 143, 283–288. [Google Scholar] [CrossRef]
- John, B.; Pandey, H.; Tripathi, R. Decomposition of fine roots of Pinus kesiya and turnover of organic matter, N and P of coarse and fine pine roots and herbaceous roots and rhizomes in subtropical pine forest stands of different ages. Biol. Fertil. Soils 2002, 35, 238–246. [Google Scholar]
- Yang, Y.S.; Chen, G.S.; Lin, P.; Huang, R.Z.; Chen, Y.X.; He, Z.M. Fine root distribution, seasonal pattern and production in a native forest and monoculture plantations in subtropical China. Acta Ecol. Sin. 2003, 23, 1719–1730. [Google Scholar]
- Steinaker, D.F.; Wilson, S.D. Belowground litter contributions to nitrogen cycling at a northern grassland-forest boundary. Ecology 2005, 86, 2825–2833. [Google Scholar] [CrossRef]
- Chen, H.; Zhao, X.; Chen, X.; Lin, Q.; Li, G. Seasonal changes of soil microbial C, N, P and associated nutrient dynamics in a semiarid grassland of north China. Appl. Soil Ecol. 2018, 28, 89–97. [Google Scholar] [CrossRef]
- Su, Y.Z.; Zhao, H.L.; Zhang, T.H. Influencing mechanism of several shrubs and subshrubs on soil fertility in Keerqin sandy land. Chin. J. Appl. Ecol. 2002, 13, 802–806. [Google Scholar]
- Zeng, Q.C.; Xin, L.I.; Dong, Y.H.; Ya-Yun, L.; Cheng, M.; Shao-Shan, A. Ecological Stoichiometry Characteristics and Physical-chemical Properties of Soils at Different Latitudes on the Loess Plateau. J. Nat. Resour. 2015, 30, 870–879. [Google Scholar]
- Yusup, M.; Mansur, S.; Nasima, N. Analysis on the Soil Physicochemical Properties of Nitraria Shrub Dune at Different Evolution Stages in Yanqi Basin of Xinjiang, China. Res. Soil Water Conserv. 2015, 22, 91–94+99. [Google Scholar]
- Catalán, N.; Marcé, R.; Kothawala, D.N.; Tranvik, L. Organic carbon decomposition rates controlled by water retention time across inland waters. Nat. Geosci. 2016, 9, 501–504. [Google Scholar] [CrossRef]
- Zhang, J.H.; Cai, D.X.; Lu, L.H.; Li, Y.X.; Li, H.; Min, H.L.; Yang, B.G.; Nong, Y.; Huang, B. Soil ecological stoichiometry of different aged Teak (Tectona grandis) plantations. Acta Ecol. Sin. 2020, 40, 5718–5728. [Google Scholar]
- Gong, X.W.; Lv, G.H.; Ma, Y.; Zhang, X.N.; He, X.M.; Guo, Z.J. Ecological Stoichiometry Characteristics in the Soil under Crown and Leaves of Two Desert Halophytes with Soil Salinity Gradients in Ebinur Lake Basin. Sci. Silvae Sin. 2017, 53, 28–36. [Google Scholar]
- Angela, H.; Graziella, B.; Claude, D.; Francisco, M.; Martin, C. Plant root growth, architecture and function. Plant Soil 2009, 321, 153–187. [Google Scholar]
- Oppelt, A.L.; Kurth, W.; Godbold, D.L. Topology, scaling relations and Leonardo’s rule in root systems from African tree species. Tree Physiol. 2001, 21, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.F.; Song, F.B.; Zhang, J.Z. Advances of research of roots responses to environmental stress on soil. Chin. Agric. Sci. Bull. 2011, 27, 44–48. [Google Scholar]
- Shan, L.S.; Li, Y.; Ren, W.; Su, S.P.; Dong, Q.L.; Geng, D.M. Root architecture of two desert plants in central Hexi Corridor of Northwest China. Chin. J. Appl. Ecol. 2013, 24, 25–31. [Google Scholar]
- Ban, Z.H.; Wang, Q. Responses of the competition between Alternanthera philoxeroides and Sambucus chinensis to simulated warming. Chin. J. Plant Ecol. 2015, 39, 43–51. [Google Scholar]
- Fitter, A.H. An Architectural Approach to the Comparative Ecology of Plant Root Systems. New Phytol. 1987, 106, 61–77. [Google Scholar] [CrossRef]
- Zhang, P.; Kang, J.L.; Yan, Z.; Tang, J.; Hao, L.; Jin, L. Similarities and differences of the plant communities on two vegetation-dunes and their responses to dune morphology. Acta Ecol. Sin. 2017, 37, 7920–7927. [Google Scholar]
- Mubareke, A.; Chen, Y.N.; Li, W.H.; Hao, M.X.; Ma, J.X.; Su, R. Fine root distribution of populus euphratica Oliv. and its relations with soil factors under extremely arid environment. J. Desert Res. 2011, 31, 1449–1458. [Google Scholar]
- Rivas-Martínez, S.; Rivas-Sáenz, S.; Penas, A. Worldwide Bioclimatic Classifification System; Global Geobotany: Kerkwerve, The Netherlands, 2011; Volume 1, p. 1–634+4 maps. [Google Scholar]
- Dang, X.H.; Meng, Z.J.; Gao, Y.; Wang, J.; Zhang, B.; Liu, B.; Wang, Z.Y.; Zhai, B. Photosynthetic carbon fixation capacity of five natural desert shrubs in west Ordos region. J. Arid. Land Resour. Environ. 2017, 31, 128–135. [Google Scholar]
- Jin, H.J.; Ma, Q.L.; He, M.Z.; Jia, X.H.; Liu, Y.J.; Zhang, Y.J.; Li, F.H. Analysis on community structure and quantitative characteristics of Nitraria tangutorum nebkhas at different succession stage in lower reaches of Shiyang River. Acta Ecol. Sin. 2013, 33, 2248–2259. [Google Scholar]
- Steingraeber, D.A.; Waller, D.M. Non-Stationarity of Tree Branching Patterns and Bifurcation Ratios. Proceedings of the Royal Society of London. Ser. B Biol. Sci. 1986, 228, 187–194. [Google Scholar]
- Berntson, G.M. The Characterization of Topology: A Comparison of Four topological Indices for Rooted Binary Trees. J. Theor. Biol. 1995, 177, 271–281. [Google Scholar] [CrossRef]
- Wang, W.D.; Bai, Z.Q.; Alimu, M.M.T.; Liu, D.; Guo, Z.J. Biomass comparison and estimation models for six dominant species of woody shrubs in the forest zones of the Tianshan Mountains. Acta Ecol. Sin. 2016, 36, 2695–2704. [Google Scholar]
- Gao, G.L.; Ding, G.D.; Zhao, Y.Y.; Feng, W.; Bai, Y.F.; Liu, Z.W. Effects of biological soil crusts on soil particle size characteristics in Mu Us sandland. Trans. Chin. Soc. Agric. Mach. 2014, 45, 115–120. [Google Scholar]
- Jackson, S. Rooting Depths, Lateral Root Spreads and Below-Ground/Above-Ground Allometries of Plants in Water-Limited Ecosystems. J. Ecol. 2002, 90, 480–494. [Google Scholar]
- Niklas, K.J. Modelling below- and above-ground biomass for non-woody and woody plants. Ann. Bot. 2005, 95, 315–321. [Google Scholar] [CrossRef] [Green Version]
- Nquist, B.J.; Niklas, K.J. Global allocation rules for patterns of biomass partitioning in seed plants. Science 2002, 295, 1517–1520. [Google Scholar] [CrossRef] [PubMed]
- Cristina, A.; Ramón, O.; Francisco, I.P. Measuring Plant Interactions: A New Comparative Index. Ecology 2004, 85, 2682–2686. [Google Scholar]
- Bao, S.D. Measurement of soil nutrients. In Soil and Agriculture Chemistry Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2010; pp. 25–144. [Google Scholar]
- Zhang, J.T. Sort. Quantitative Ecology, 3rd ed.; China Science Publishing: Beijing, China, 2020; pp. 177–249. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Stage | Ws (m) | Ls (m) | Hs (m) | C (m2) | Wn (m) | Ln (m) | Hn (m) | Sn (m2) | Vn (m3) | |
|---|---|---|---|---|---|---|---|---|---|---|
| Rudimental stage | 1 | 2.41 | 3.19 | 0.99 | 3.43 | 2.61 | 3.23 | 0.49 | 6.62 | 2.16 |
| 2 | 2.32 | 2.87 | 0.91 | 2.81 | 2.28 | 2.74 | 0.46 | 4.91 | 1.50 | |
| 3 | 2.34 | 2.94 | 0.86 | 2.84 | 2.31 | 2.99 | 0.48 | 5.42 | 1.74 | |
| Developing stage | 4 | 3.78 | 4.15 | 0.99 | 5.19 | 3.76 | 4.18 | 0.68 | 12.34 | 5.60 |
| 5 | 3.32 | 3.94 | 1.09 | 4.97 | 3.56 | 3.98 | 0.64 | 11.13 | 4.75 | |
| 6 | 3.24 | 3.27 | 1.06 | 3.68 | 3.36 | 3.88 | 0.56 | 10.24 | 3.82 | |
| Stabilizing stage | 7 | 4.32 | 5.14 | 1.33 | 8.22 | 5.06 | 5.88 | 0.84 | 23.37 | 13.09 |
| 8 | 4.32 | 5.50 | 1.54 | 9.73 | 5.23 | 6.17 | 0.91 | 25.34 | 15.38 | |
| 9 | 4.07 | 4.76 | 1.41 | 7.47 | 5.11 | 5.99 | 0.89 | 24.04 | 14.26 | |
| Growth Stage | Allometric Scaling Exponent | 95% Confidence Interval | Allometric Relationships | ||
|---|---|---|---|---|---|
| Log10CHs/2 | Log10Vn | α | |||
| Rudimental stage | 0.2300 | 0.3350 | 0.3701 | 0.1671–0.5519 | Isokinetic |
| 0.1060 | 0.2428 | −0.0010 | Allometric | ||
| 0.0861 | 0.2395 | −0.0837 | Allometric | ||
| Developing stage | 0.4096 | 0.7479 | 0.4055 | Isokinetic | |
| 0.4326 | 0.6765 | 0.4823 | Isokinetic | ||
| 0.2903 | 0.5824 | 0.3163 | Isokinetic | ||
| Stabilizing stage | 0.7377 | 1.1168 | 0.5653 | Allometric | |
| 0.8747 | 1.1868 | 0.6474 | Allometric | ||
| 0.7218 | 1.1542 | 0.5334 | Isokinetic | ||
| Stoichiometric Ratio | Soil Depth (cm) | Growth Stage | |||
|---|---|---|---|---|---|
| CK | Rudimental Stage | Developing Stage | Stabilizing Stage | ||
| C:N | 0–10 | 19.52 ± 1.44 Aa | 20.29 ± 3.08 Aa | 18.85 ± 1.31 Aa | 20.22 ± 0.36 Aa |
| 10–20 | 21.25 ± 3.55 Aa | 19.46 ± 1.13 Aa | 13.26 ± 1.45 Bb | 19.69 ± 1.68 Aa | |
| 20–30 | 22.21 ± 2.19 Aa | 20.11 ± 1.79 ABa | 17.69 ± 0.80 Ba | 20.09 ± 2.24 ABa | |
| 30–40 | 20.55 ± 4.94 Aa | 19.14 ± 5.03 Aa | 19.24 ± 1.40 Aa | 15.36 ± 1.84 Ab | |
| 0–40 | 20.68 ± 1.51 A | 19.61 ± 0.67 AB | 17.34 ± 0.44 C | 18.76 ± 0.87 BC | |
| C:P | 0–10 | 4.02 ± 0.62 Aab | 4.21 ± 0.35 Aa | 3.52 ± 0.31 Aab | 4.06 ± 0.48 Aab |
| 10–20 | 5.02 ± 0.08 Aa | 4.12 ± 0.23 Ba | 2.78 ± 0.53 Cb | 4.03 ± 0.23 Bab | |
| 20–30 | 5.07 ± 0.96 Aa | 3.60 ± 1.11 Aa | 4.36 ± 0.71 Aab | 4.58 ± 0.59 Aa | |
| 30–40 | 3.86 ± 0.27 Ab | 3.37 ± 0.41 Aa | 4.79 ± 1.32 Aa | 3.41 ± 0.63 Ab | |
| 0–40 | 4.42 ± 0.15 A | 3.81 ± 0.31 B | 3.78 ± 0.37 B | 4.00 ± 0.33 AB | |
| N:P | 0–10 | 0.21 ± 0.05 Aa | 0.21 ± 0.03 Aa | 0.19 ± 0.01 Aa | 0.20 ± 0.02 Aa |
| 10–20 | 0.24 ± 0.04 Aa | 0.21 ± 0.02 Aa | 0.21 ± 0.04 Aa | 0.21 ± 0.03 Aa | |
| 20–30 | 0.23 ± 0.07 Aa | 0.18 ± 0.06 Aa | 0.25 ± 0.05 Aa | 0.23 ± 0.01 Aa | |
| 30–40 | 0.20 ± 0.05 Aa | 0.19 ± 0.06 Aa | 0.25 ± 0.09 Aa | 0.23 ± 0.06 Aa | |
| 0–40 | 0.21 ± 0.02 A | 0.19 ± 0.02 A | 0.22 ± 0.03 A | 0.21 ± 0.01 A | |
| Name | Explains % | Contribution % | Pseudo-F | p | Importance Sequencing |
|---|---|---|---|---|---|
| SOC | 58.4 | 71.7 | 9.8 | 0.006 | 1 |
| C:P | 8.5 | 10.4 | 1.5 | 0.206 | 2 |
| TP | 5.3 | 6.5 | 1.0 | 0.416 | 3 |
| N:P | 3.9 | 4.7 | 0.5 | 0.648 | 4 |
| C:N | 3.0 | 3.7 | 0.3 | 0.748 | 5 |
| TN | 2.3 | 2.9 | 0.4 | 0.782 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Dang, X.; Gao, Y.; Meng, Z.; Chen, X.; Wang, Y. Response Mechanisms of Adventitious Root Architectural Characteristics of Nitraria tangutorum Shrubs to Soil Nutrients in Nabkha. Plants 2022, 11, 3218. https://doi.org/10.3390/plants11233218
Li X, Dang X, Gao Y, Meng Z, Chen X, Wang Y. Response Mechanisms of Adventitious Root Architectural Characteristics of Nitraria tangutorum Shrubs to Soil Nutrients in Nabkha. Plants. 2022; 11(23):3218. https://doi.org/10.3390/plants11233218
Chicago/Turabian StyleLi, Xiaole, Xiaohong Dang, Yong Gao, Zhongju Meng, Xue Chen, and Yanyi Wang. 2022. "Response Mechanisms of Adventitious Root Architectural Characteristics of Nitraria tangutorum Shrubs to Soil Nutrients in Nabkha" Plants 11, no. 23: 3218. https://doi.org/10.3390/plants11233218
APA StyleLi, X., Dang, X., Gao, Y., Meng, Z., Chen, X., & Wang, Y. (2022). Response Mechanisms of Adventitious Root Architectural Characteristics of Nitraria tangutorum Shrubs to Soil Nutrients in Nabkha. Plants, 11(23), 3218. https://doi.org/10.3390/plants11233218