
Panax notoginseng Suppresses Bone Morphogenetic Protein-2 Expression in EA.hy926 Endothelial Cells by Inhibiting the Noncanonical NF-κB and Wnt/β-Catenin Signaling Pathways



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
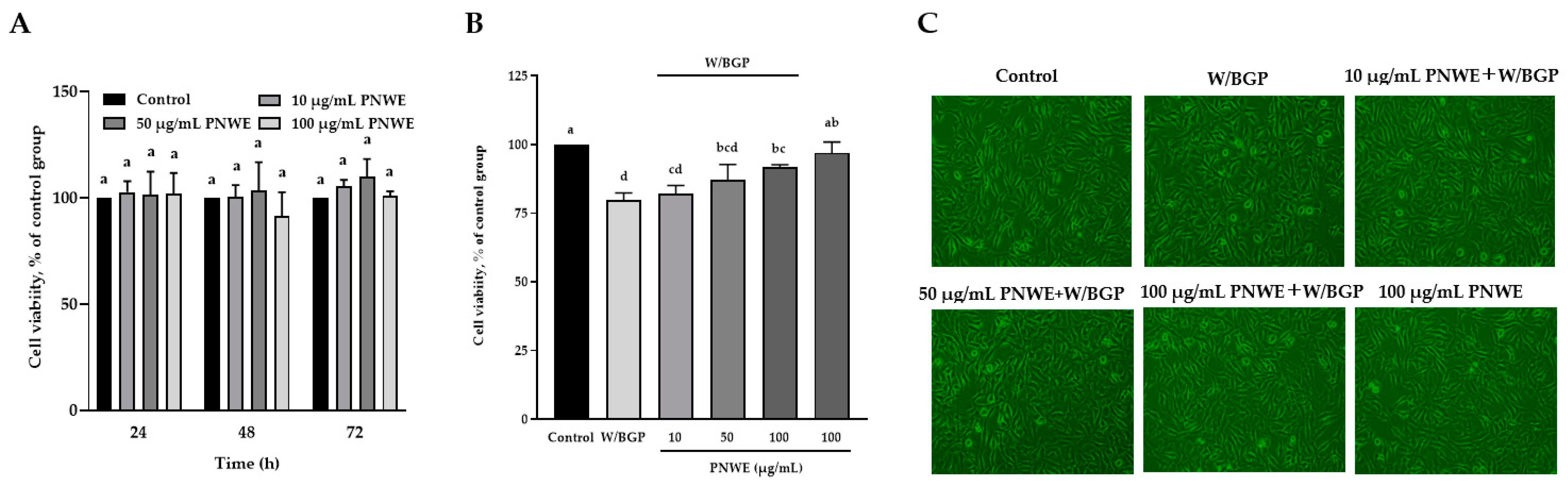
2.1. PNWE Protects EA.hy926 Cells from W/BGP-Induced Damage
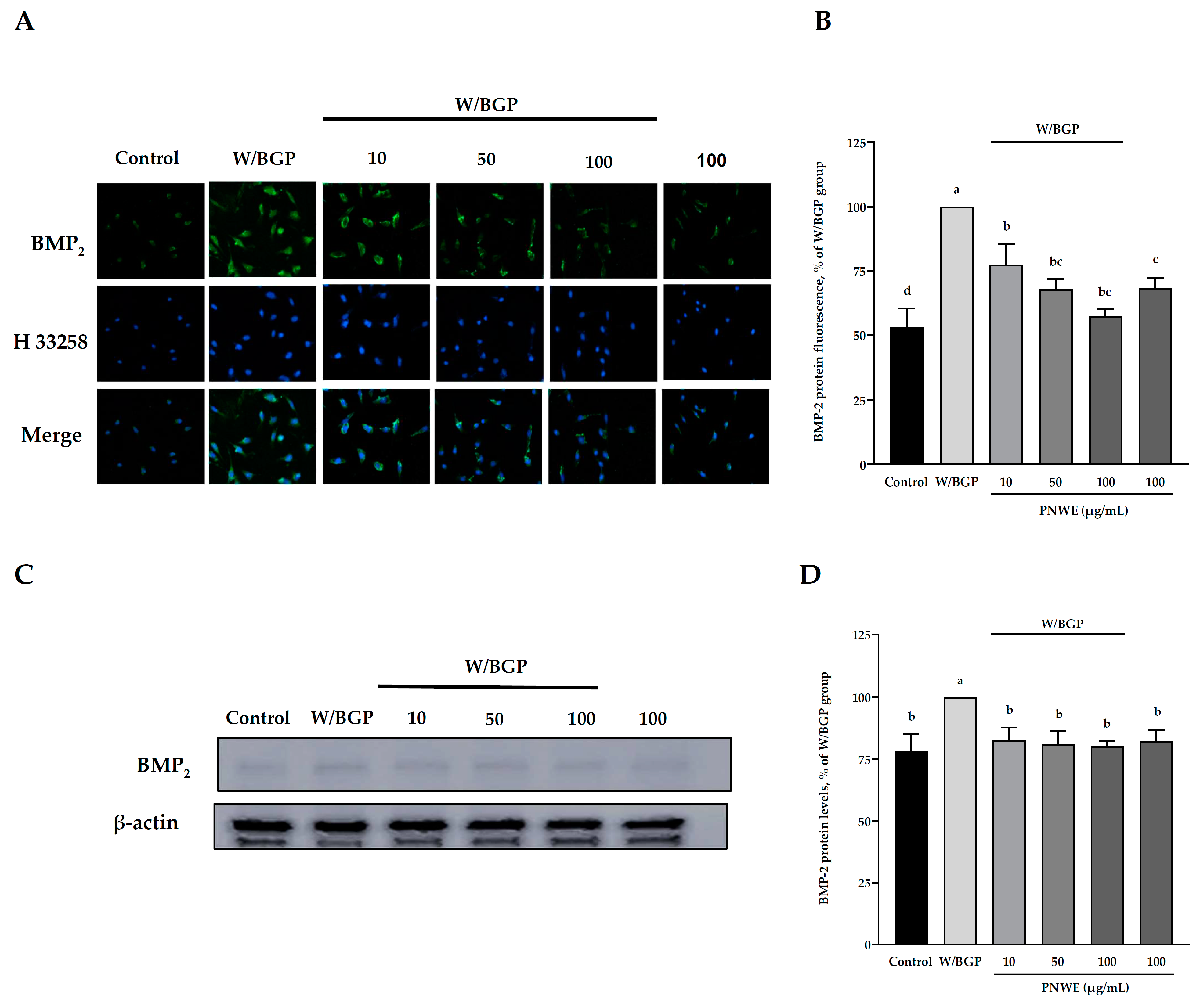
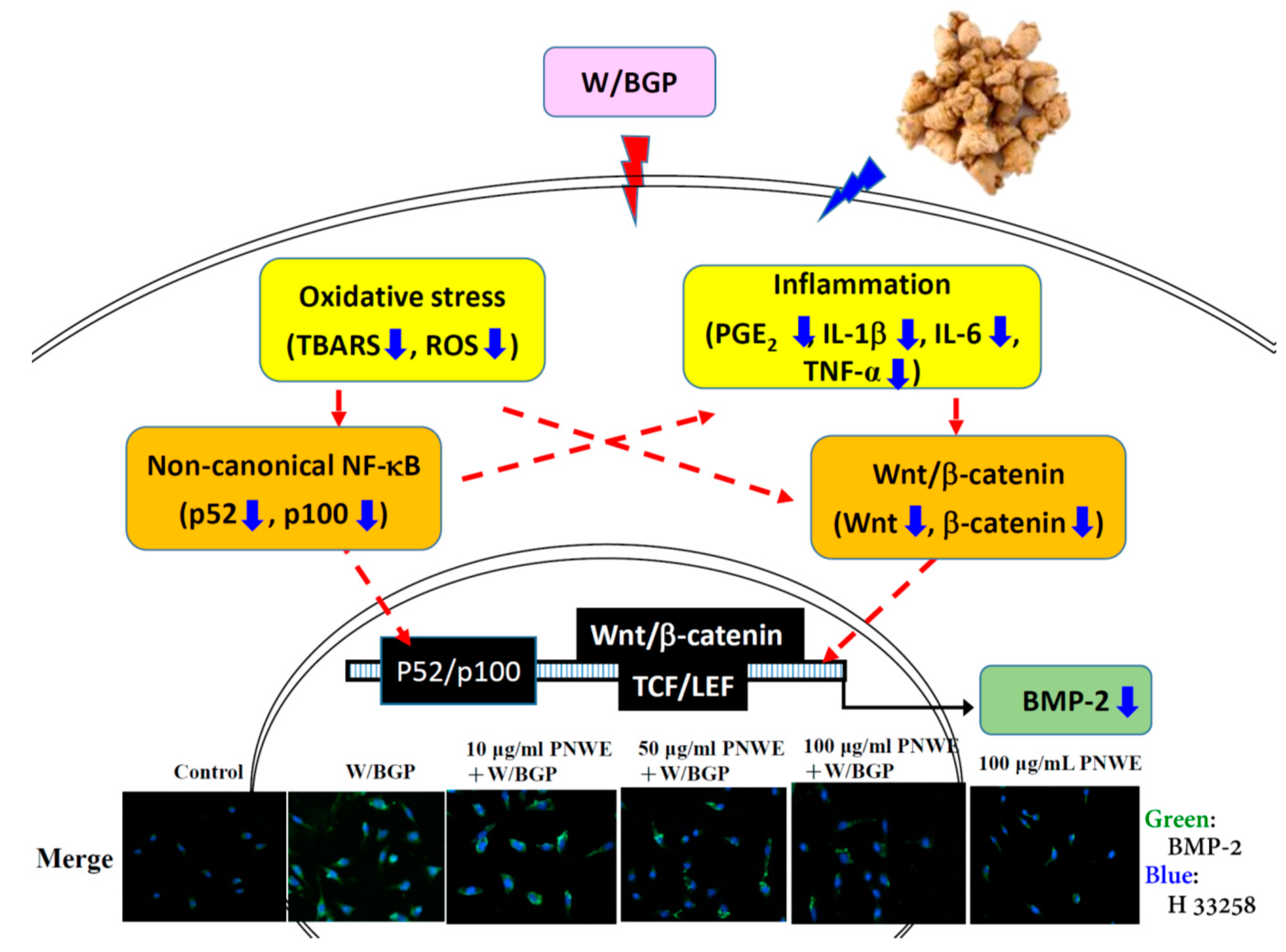
2.2. PNWE Decreased W/BGP-Induced BMP-2 Protein Expression in EA.hy926 Cells
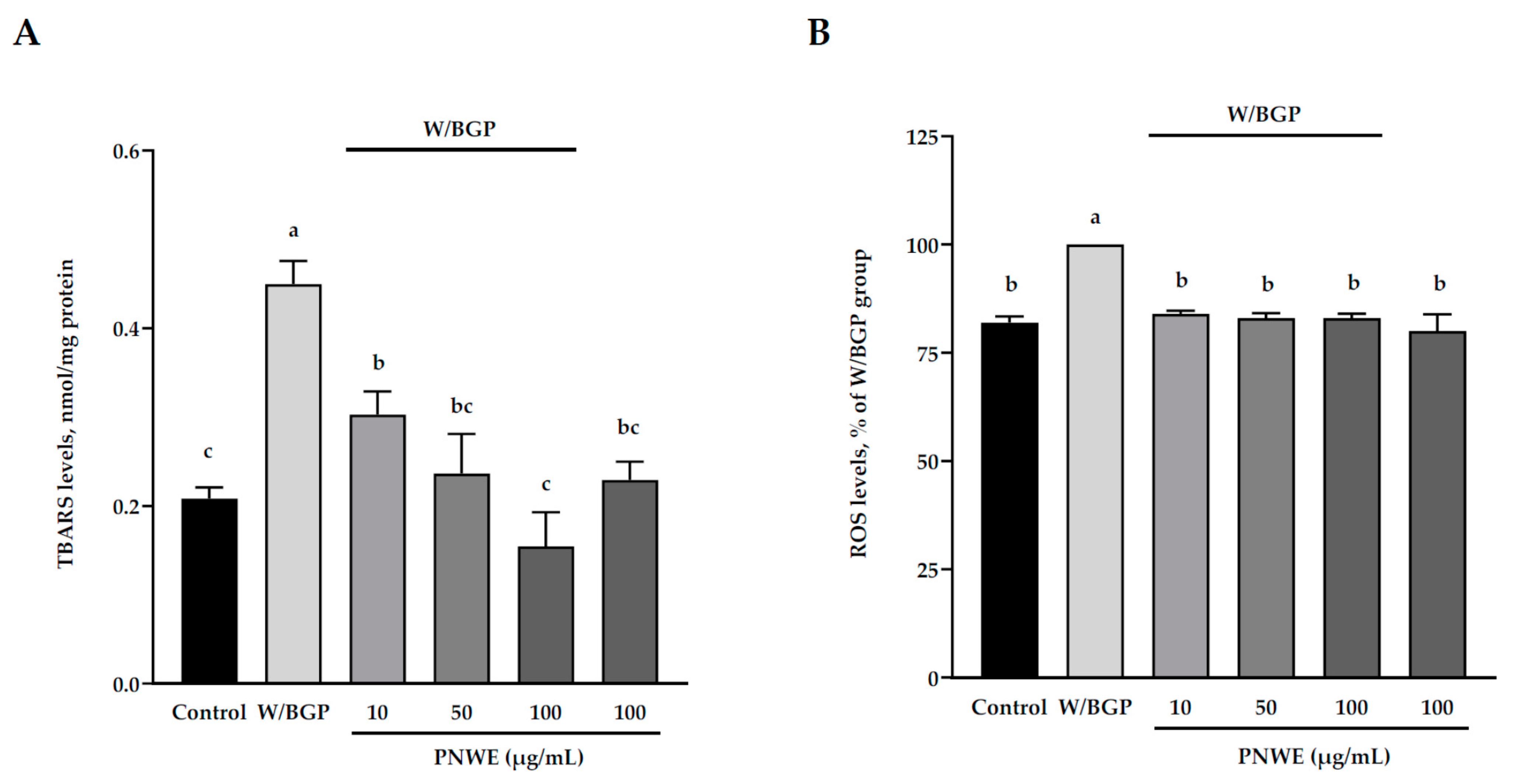
2.3. PNWE Decreased W/BGP-Induced Oxidative Stress in EA.hy926 Cells
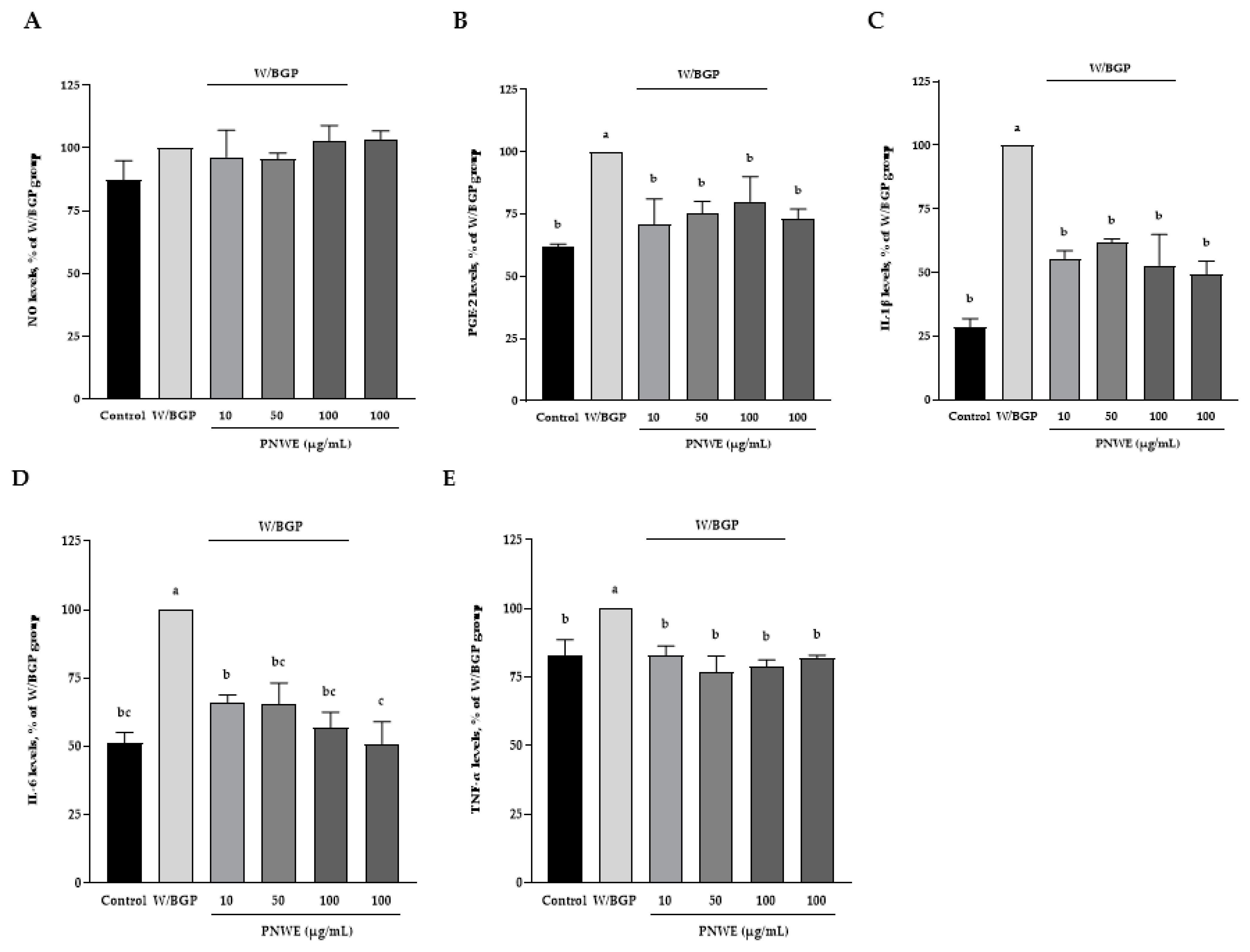
2.4. PNWE Alleviated the W/BGP-Induced Inflammatory Response in EA.hy926 Cells
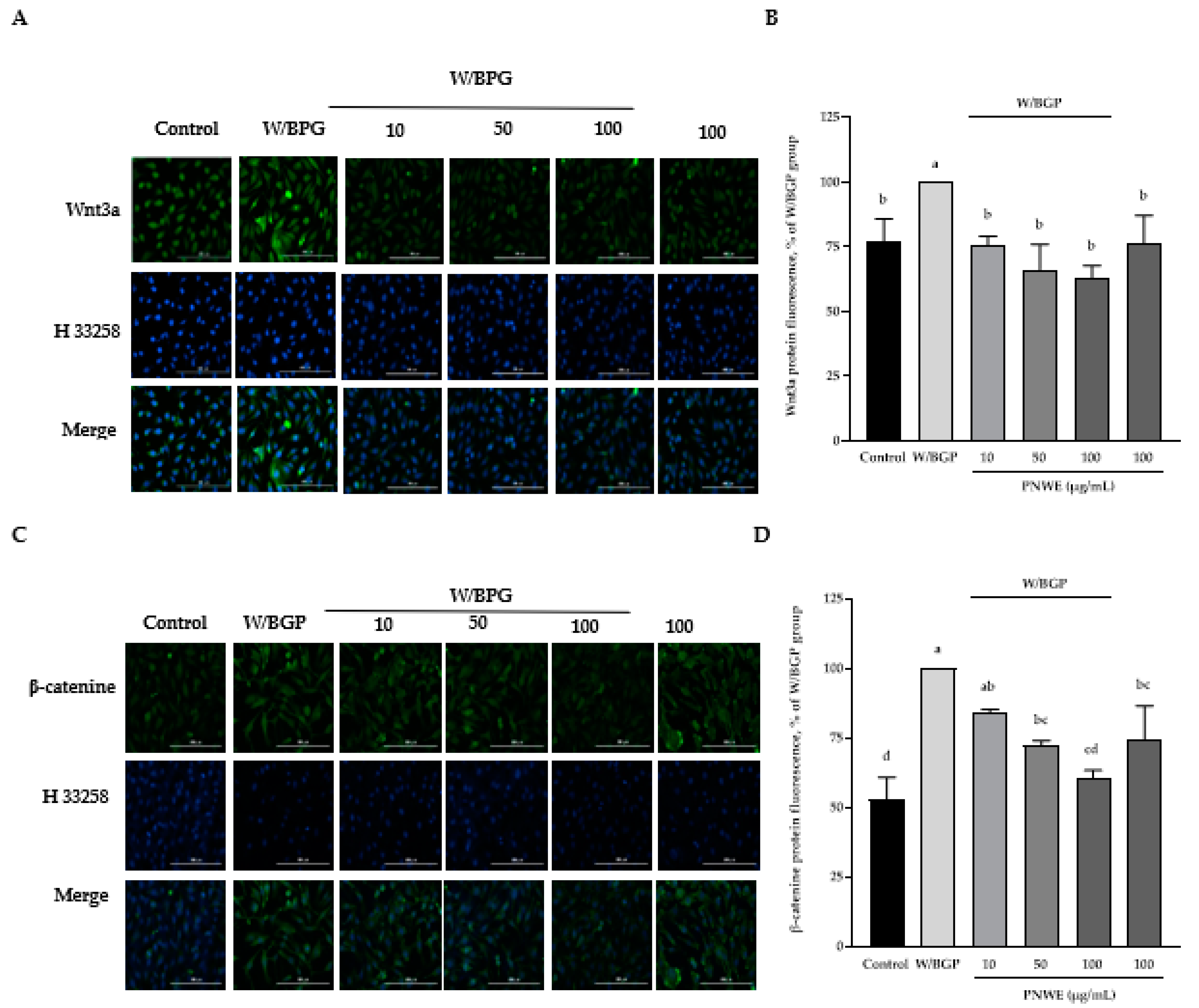
2.5. PNWE Decreased the W/BGP-Induced Activation of the Wnt3a/β-Catenin Signaling Pathway in EA.hy926 Cells
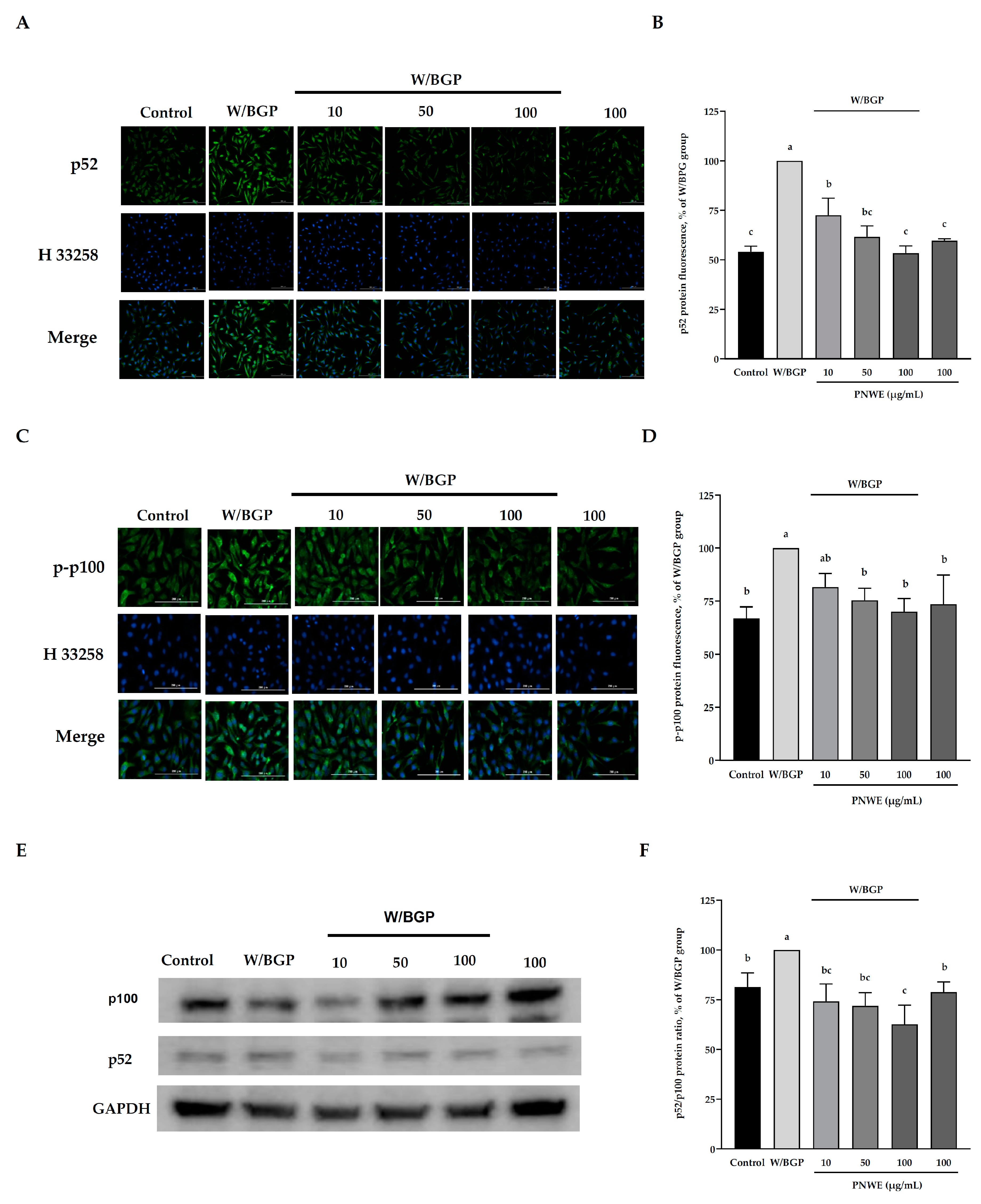
2.6. PNWE Decreases the W/BGP-Induced Activation of the Noncanonical NF-κB Signaling Pathway in EA.hy926 Cells
3. Discussion
4. Materials and Methods
4.1. Preparation of PN Water Extract (PNWE)
4.2. Cell Culture and Treatment
4.3. Cell Viability Analysis
4.4. Intracellular Lipid Peroxidation and Reactive Oxygen Species (ROS) Level Analysis
4.5. Intracellular NO and Prostaglandin E2 (PGE2) Determination
4.6. Intracellular Interleukin-1β (IL-1β), Interleukin-6 (IL-6), and Tumor Necrosis Factor-α (TNF-α) Level Assessment
4.7. Intracellular BMP-2, Wnt, β-Catenin, p52, and p100 Content Analysis by Immunocytochemistry
4.8. Immunoblotting Analysis of Intracellular BMP-2, p52, and p100 Protein Expression
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Yang, H.; Liu, Z.; Hu, X.; Lu, X.; Gui, L.; Cai, Z.; Dai, C. Protective effect of Panax notoginseng saponins on apolipoprotein-E-deficient atherosclerosis-prone Mice. Curr. Pharm. Des. 2022, 28, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, H.; Yang, H.R.; Zeng, Y.J. Hypoglycemic effects of novel Panax notoginseng polysaccharide in mice with diet-induced obesity. Foods 2022, 11, 3101. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Zhang, Y.; Liu, T.; Li, J.; Li, H. The anti-osteosarcoma effect from Panax notoginseng saponins by inhibiting the G0 / G1 phase in the cell cycle and affecting p53-mediated autophagy and mitochondrial apoptosis. J. Cancer 2021, 12, 6383–6392. [Google Scholar] [CrossRef]
- Hsieh, S.L.; Hsieh, S.; Kuo, Y.H.; Wang, J.J.; Wang, J.C.; Wu, C.C. Effects of Panax notoginseng on the metastasis of human colorectal cancer cells. Am. J. Chin. Med. 2016, 44, 851–870. [Google Scholar] [CrossRef]
- Lee, C.Y.; Hsieh, S.L.; Hsieh, S.; Tsai, C.C.; Hsieh, L.C.; Kuo, Y.H.; Wu, C.C. Inhibition of human colorectal cancer metastasis by notoginsenoside R1, an important compound from Panax notoginseng. Oncol. Rep. 2017, 37, 399–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, D.; Tan, R.Z.; Zhao, C.Y.; Li, J.C.; Zhong, X.; Diao, H.; Lin, X.; Duan, D.D.; Fan, J.M.; Xie, X.S.; et al. Astragalus mongholicus Bunge and Panax notoginseng (Burkill) F.H. Chen formula for renal injury in diabetic nephropathy-In Vivo and In Vitro evidence for autophagy regulation. Front. Pharmacol. 2020, 11, 732. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Chao, C.T.; Huang, J.W.; Huang, K.C. Vascular calcification as an underrecognized risk factor for frailty in 1783 community-dwelling elderly individuals. J. Am. Heart Assoc. 2020, 9, e017308. [Google Scholar] [CrossRef]
- Li, Y.; Guo, Q.; Huang, J.; Wang, Z. Antidepressant active ingredients from Chinese traditional herb Panax notoginseng: A pharmacological mechanism review. Front. Pharmacol. 2022, 13, 922337. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, C.; Miao, L.; Tan, Y.; Zhou, Y.; Cheong, M.S.; Huang, Y.; Wang, Y.; Yu, H.; Cheang, W.S. Panax notoginseng protects against diabetes-associated endothelial dysfunction: Comparison between ethanolic extract and total saponin. Oxid. Med. Cell Longev. 2021, 2021, 4722797. [Google Scholar] [CrossRef]
- Pan, C.; Huo, Y.; An, X.; Singh, G.; Chen, M.; Yang, Z.; Pu, J.; Li, J. Panax notoginseng and its components decreased hypertension via stimulation of endothelial-dependent vessel dilatation. Vascul. Pharmacol. 2012, 56, 150–158. [Google Scholar] [CrossRef]
- Lee, S.N.; Moon, D.; Her, S.H.; Jang, W.Y.; Moon, K.W.; Yoo, K.D.; Lee, K.; Lee, J.H.; Lee, J.H.; Lee, S.R.; et al. Impact of diabetes mellitus on periprocedural and 18-month clinical outcomes in Korean patients requiring rotational atherectomy: Results from the ROCK Registry. Ann. Saudi Med. 2022, 42, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.R.; Joo, Y.S.; Kim, H.W.; Park, J.T.; Chang, T.I.; Son, N.H.; Yoo, T.H.; Kang, S.W.; Sung, S.; Lee, K.B.; et al. Korean cohort study for outcomes in patients with chronic kidney disease (KNOW-CKD) investigators. Coronary artery calcification score and the progression of chronic kidney disease. J. Am. Soc. Nephrol. 2022, 33, 1590–1601. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Hao, J.; Wang, X.; Liu, J.; Ni, J.; Hao, L. The role of AIF-1 in the aldosterone-induced vascular calcification related to chronic kidney disease: Evidence from mice model and cell co-culture model. Front. Endocrinol. 2022, 13, 917356. [Google Scholar] [CrossRef] [PubMed]
- Lau, W.L.; Ix, J.H. Clinical detection, risk factors, and cardiovascular consequences of medial arterial calcification: A pattern of vascular injury associated with aberrant mineral metabolism. Semin. Nephrol. 2013, 33, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, V.P.; Pozzan, G.; Castelli, V.; Caffaro, R.V. Arteriosclerosis, atherosclerosis, arteriolosclerosis, and Monckeberg medial calcific sclerosis: What is the difference? J. Vasc. Bras. 2021, 20, e20200211. [Google Scholar] [CrossRef] [PubMed]
- Joensuu, K.; Uusitalo-Kylmälä, L.; Hentunen, T.A.; Heino, T.J. Angiogenic potential of human mesenchymal stromal cell and circulating mononuclear cell cocultures is reflected in the expression profiles of proangiogenic factors leading to endothelial cell and pericyte differentiation. J. Tissue Eng. Regen. Med. 2018, 12, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, C.; Qian, C.; Abela, G.; Sun, W.; Kong, X. Ginkgo Biloba extract EGB761 alleviates warfarin-induced aortic valve calcification through the BMP2/Smad1/5/Runx2 signaling pathway. J. Cardiovasc. Pharmacol. 2021, 78, 411–421. [Google Scholar] [CrossRef]
- Ingwersen, L.C.; Frank, M.; Naujokat, H.; Loger, K.; Bader, R.; Jonitz-Heincke, A. BMP-2 long-term stimulation of human pre-osteoblasts induces osteogenic differentiation and promotes transdifferentiation and bone remodeling processes. Int. J. Mol. Sci. 2022, 23, 3077. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Akiyama, S.; Katagiri, T.; Namiki, M.; Kurokawa, T.; Suda, T. Smad1 and smad5 act downstream of intracellular signalings of BMP-2 that inhibits myogenic differentiation and induces osteoblast differentiation in C2C12 myoblasts. Biochem. Biophys. Res. Commun. 1997, 238, 574–580. [Google Scholar] [CrossRef]
- Nohe, A.; Keating, E.; Knaus, P.; Petersen, N.O. Signal transduction of bone morphogenetic protein receptors. Cell Signal. 2004, 16, 291–299. [Google Scholar] [CrossRef]
- Luo, K. Signaling cross talk between TGF-β/smad and other signaling pathways. Cold Spring Harb. Perspect. Biol. 2017, 9, a022137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, Y.; Fukushima, H.; Maruyama, T.; Kuroishi, K.N.; Osawa, K.; Nagano, K.; Aoki, K.; Weih, F.; Doi, T.; Zhang, M.; et al. Accumulation of p100, a precursor of NF-κB2, enhances osteoblastic differentiation in vitro and bone formation in vivo in aly/aly mice. Mol. Endocrinol. 2012, 26, 414–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hénaut, L.; Sanz, A.B.; Martin-Sanchez, D.; Carrasco, S.; Villa-Bellosta, R.; Aldamiz-Echevarria, G.; Massy, Z.A.; Sanchez-Nino, M.D.; Ortiz, A. TWEAK favors phosphate-induced calcification of vascular smooth muscle cells through canonical and non-canonical activation of NFκB. Cell Death Dis. 2016, 7, e2305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajla, S.; Mondol, A.S.; Nagasawa, A.; Zhang, Y.; Kato, M.; Matsuno, K.; Yabe-Nishimura, C.; Kamata, T. A crucial role for Nox 1 in redox-dependent regulation of Wnt-β-catenin signaling. FASEB J. 2012, 26, 2049–2059. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Troncone, L.; Augur, Z.M.; Kim, S.S.J.; McNeil, M.E.; Yu, P.B. The role of bone morphogenetic protein signaling in vascular calcification. Bone 2020, 141, 115542. [Google Scholar] [CrossRef] [PubMed]
- Sulistyowati, E.; Hsu, J.H.; Lee, S.J.; Huang, S.E.; Sihotang, W.Y.; Wu, B.N.; Dai, Z.K.; Lin, M.C.; Yeh, J.L. Potential actions of baicalein for preventing vascular calcification of smooth muscle cells in vitro and in vivo. Int. J. Mol. Sci. 2022, 23, 5673. [Google Scholar] [CrossRef]
- Hong, O.K.; Lee, S.S.; Yoo, S.J.; Lee, M.K.; Kim, M.K.; Baek, K.H.; Song, K.H.; Kwon, H.S. Gemigliptin inhibits interleukin-1β-Induced endothelial-mesenchymal transition via canonical-bone morphogenetic protein pathway. Endocrinol. Metab. 2020, 35, 384–395. [Google Scholar] [CrossRef]
- Mao, W.; Fan, Y.; Wang, X.; Feng, G.; You, Y.; Li, H.; Chen, Y.; Yang, J.; Weng, H.; Shen, X. Phloretin ameliorates diabetes-induced endothelial injury through AMPK-dependent anti-EndMT pathway. Pharmacol. Res. 2022, 179, 106205. [Google Scholar] [CrossRef]
- Xue, Q.; He, N.; Wang, Z.; Fu, X.; Aung, L.H.H.; Liu, Y.; Li, M.; Cho, J.Y.; Yang, Y.; Yu, T. Functional roles and mechanisms of ginsenosides from Panax ginseng in atherosclerosis. J. Ginseng. Res. 2021, 45, 22–31. [Google Scholar] [CrossRef]
- Su, P.; Du, S.; Li, H.; Li, Z.; Xin, W.; Zhang, W. Notoginsenoside R1 inhibits oxidized low-density lipoprotein induced inflammatory cytokines production in human endothelial EA.hy926 cells. Eur. J. Pharmacol. 2016, 770, 9–15. [Google Scholar] [CrossRef]
- Li, X.Q.; Huang, T.Y. Notoginsenoside R1 alleviates high glucose-induced inflammation and oxidative stress in HUVECs via upregulating miR-147a. Kaohsiung J. Med. Sci. 2021, 37, 1101–1112. [Google Scholar] [CrossRef]
- Jimi, E.; Katagiri, T. Critical roles of NF-κB signaling molecules in bone metabolism revealed by genetic mutations in osteopetrosis. Int. J. Mol. Sci. 2022, 23, 7995. [Google Scholar] [CrossRef]
- Jimi, E.; Fei, H.; Nakatomi, C. NF-κB signaling regulates physiological and pathological chondrogenesis. Int. J. Mol. Sci. 2019, 20, 6275. [Google Scholar] [CrossRef] [Green Version]
- Kunnumakkara, A.B.; Shabnam, B.; Girisa, S.; Harsha, C.; Banik, K.; Devi, T.B.; Choudhury, R.; Sahu, H.; Parama, D.; Sailo, B.L.; et al. Inflammation, NF-κB, and chronic diseases: How are they linked? Crit. Rev. Immunol. 2020, 40, 1–39. [Google Scholar] [CrossRef]
- Toker, H.; Görgün, E.P.; Korkmaz, E.M. Analysis of IL-6, IL-10 and NF-kappaB gene polymorphisms in aggressive and chronic periodontitis. Cent. Eur. J. Public Health 2017, 25, 157–162. [Google Scholar] [CrossRef] [Green Version]
- Infantino, V.; Pierri, C.L.; Iacobazzi, V. Metabolic routes in inflammation: The citrate pathway and its potential as therapeutic target. Curr. Med. Chem. 2020, 26, 7104–7116. [Google Scholar] [CrossRef]
- Jimi, E.; Hirata, S.; Shin, M.; Yamazaki, M.; Fukushima, H. Molecular mechanisms of BMP-induced bone formation: Cross-talk between BMP and NF-κB signaling pathways in osteoblastogenesis. Jpn. Dent. Sci. Rev. 2010, 46, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Csiszar, A.; Smith, K.E.; Koller, A.; Kaley, G.; Edwards, J.G.; Ungvari, Z. Regulation of bone morphogenetic protein-2 expression in endothelial cells: Role of nuclear factor-kappaB activation by tumor necrosis factor-alpha, H2O2, and high intravascular pressure. Circulation 2005, 111, 2364–2372. [Google Scholar] [CrossRef] [Green Version]
- Csiszar, A.; Smith, K.; Labinskyy, N.; Orosz, Z.; Rivera, A.; Ungvari, Z. Resveratrol attenuates TNF-alpha-induced activation of coronary arterial endothelial cells: Role of NF-kappaB inhibition. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H1694–H1699. [Google Scholar] [CrossRef]
- Zhang, M.; Zhou, S.H.; Zhao, S.; Li, X.P.; Liu, L.P.; Shen, X.Q. Pioglitazone can downregulate bone morphogenetic protein-2 expression induced by high glucose in human umbilical vein endothelial cells. Pharmacology 2008, 81, 312–316. [Google Scholar] [CrossRef]
- Rong, S.; Zhao, X.; Jin, X.; Zhang, Z.; Chen, L.; Zhu, Y.; Yuan, W. Vascular calcification in chronic kidney is induced by bone morphogenetic protein-2 via a mechanism involving the Wnt/β-catenin pathway. Cell Physiol. Biochem. 2014, 34, 2049–2060. [Google Scholar] [CrossRef]
- Gao, M.; Chen, T.; Wu, L.; Zhao, X.; Mao, H.; Xing, C. Effect of pioglitazone on the calcification of rat vascular smooth muscle cells through the downregulation of the Wnt/β-catenin signaling pathway. Mol. Med. Rep. 2017, 16, 6208–6213. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Ma, L.; Dai, L.; Zuo, D.; Li, X.; Zhu, H.; Xu, F. TNF-α promotes the malignant transformation of intestinal stem cells through the NF-κB and Wnt/β-catenin signaling pathways. Oncol. Rep. 2020, 44, 577–588. [Google Scholar] [CrossRef]
- Beazley, K.E.; Eghtesad, S.; Nurminskaya, M.V. Quercetin attenuates warfarin-induced vascular calcification in vitro independently from matrix Gla protein. J. Biol. Chem. 2013, 288, 2632–2640. [Google Scholar] [CrossRef] [Green Version]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival. Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods. 1986, 89, 271–277. [Google Scholar] [CrossRef]
- Hsieh, S.L.; Shih, Y.W.; Chiu, Y.M.; Tseng, S.F.; Li, C.C.; Wu, C.C. By-products of the black soybean sauce manufacturing process as potential antioxidant and anti-inflammatory materials for use as functional foods. Plants 2021, 10, 2579. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ping, T.-N.; Hsieh, S.-L.; Wang, J.-J.; Chen, J.-B.; Wu, C.-C. Panax notoginseng Suppresses Bone Morphogenetic Protein-2 Expression in EA.hy926 Endothelial Cells by Inhibiting the Noncanonical NF-κB and Wnt/β-Catenin Signaling Pathways. Plants 2022, 11, 3265. https://doi.org/10.3390/plants11233265
Ping T-N, Hsieh S-L, Wang J-J, Chen J-B, Wu C-C. Panax notoginseng Suppresses Bone Morphogenetic Protein-2 Expression in EA.hy926 Endothelial Cells by Inhibiting the Noncanonical NF-κB and Wnt/β-Catenin Signaling Pathways. Plants. 2022; 11(23):3265. https://doi.org/10.3390/plants11233265
Chicago/Turabian StylePing, Tsu-Ni, Shu-Ling Hsieh, Jyh-Jye Wang, Jin-Bor Chen, and Chih-Chung Wu. 2022. "Panax notoginseng Suppresses Bone Morphogenetic Protein-2 Expression in EA.hy926 Endothelial Cells by Inhibiting the Noncanonical NF-κB and Wnt/β-Catenin Signaling Pathways" Plants 11, no. 23: 3265. https://doi.org/10.3390/plants11233265
APA StylePing, T.-N., Hsieh, S.-L., Wang, J.-J., Chen, J.-B., & Wu, C.-C. (2022). Panax notoginseng Suppresses Bone Morphogenetic Protein-2 Expression in EA.hy926 Endothelial Cells by Inhibiting the Noncanonical NF-κB and Wnt/β-Catenin Signaling Pathways. Plants, 11(23), 3265. https://doi.org/10.3390/plants11233265