
Nutritional and Pharmaceutical Applications of Under-Explored Knottin Peptide-Rich Phytomedicines

 , , , ,
, , , , 
Abstract
:1. Introduction
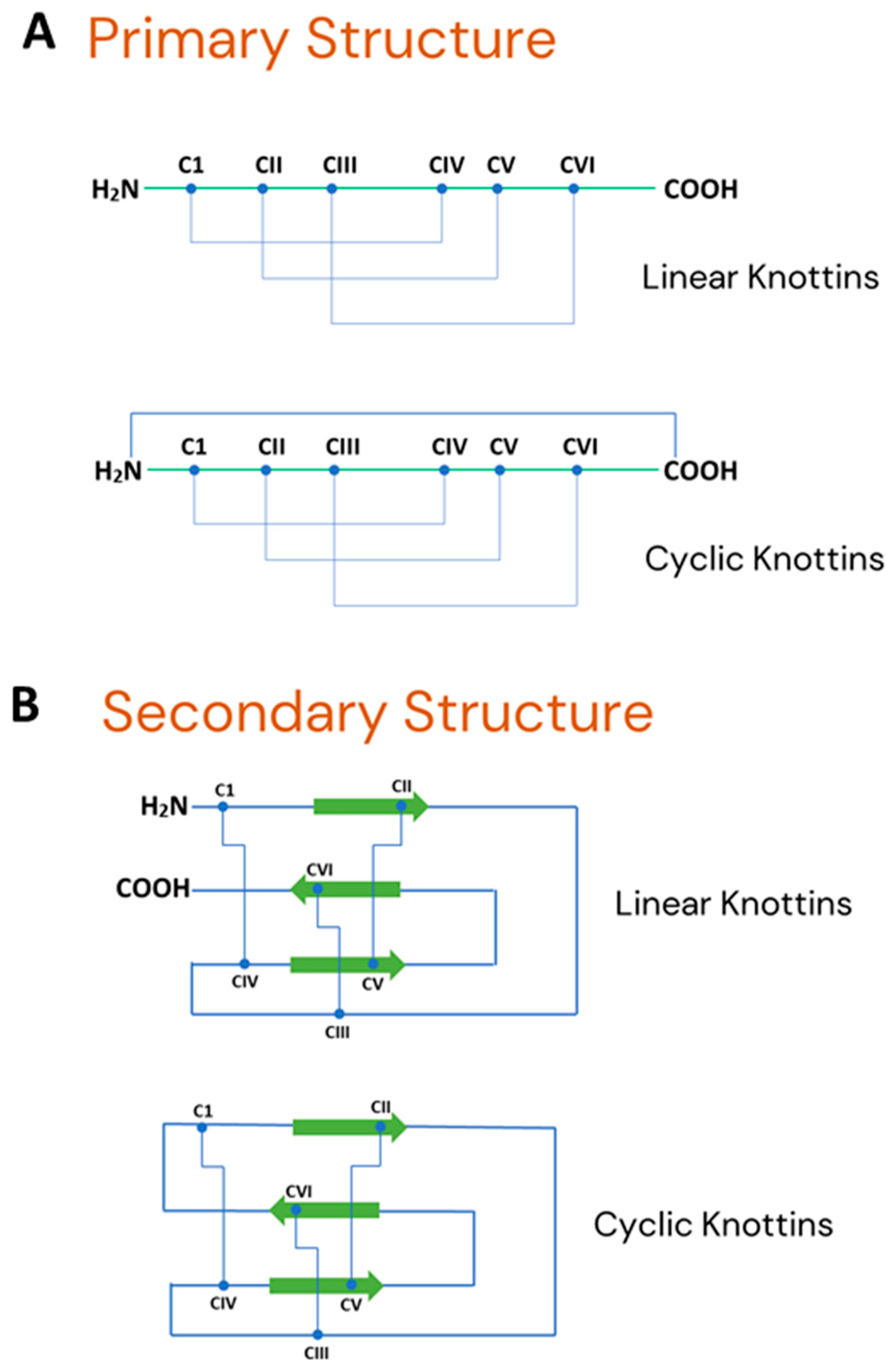
Description, Distribution, Diversity, Abundance and Type of Knottin Peptides
2. Applications of Knottin-Rich Peptides
2.1. Nutritional Applications of Knottin-Rich Phytomedicines

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytomedicine/ Botanical Source/Country | Knottins Present | Nutraceutical Applications | Scientific Evidence | References |
|---|---|---|---|---|
| Ewa/ Clitoria ternatea L. Indonesia | Cyclotides, e.g., Cter M | C. ternatea flowers are used as a natural coloring for food and beverage. It helps to improve food quality by affecting the water and oil retention capacity, colloidal stability, viscosity, and foam generation in the finished product. It possesses anti-osteoporotic, antihypertensive activity and reduces cardiovascular complications. | In silico, in vitro and in vivo | [58,59] |
| Ipecac/ Carapichea ipecacuanha (Brot.) L. Andersson Brazil | Caripe Cyclotides (Caripe 1–13) | It helps to minimize the cooking/frying loss and shrinkage and enhances foaming stability in chicken sausages. Amoebic dysentery, cough, whooping cough and bronchitis can be treated using root extracts and powdered leaf of Carapichea. C. ipecacuanha contains emetine and cephaeline, to induce vomiting. It helps to reduce the risk of certain chronic diseases such as leukemia. The natural action of the bioactive compounds protects the biological attributes such as anti-aging, anti-inflammatory, anti-viral, anti-microbial, and anti-cancer properties. | [60,61,62,63] | |
| Beetroot/ Beta vulgaris (L.) Arcang Mediterranean Europe and North America | bevuTi-I | Beet juices, peels and pomaces is a plausible source of iron and helps in treating malnutrition and iron deficiency anemia. Beetroot extracts have a considerable number of antioxidants which are used in heavy metal toxicity and cancer chemoprevention. Studies have reported the use of beetroot supplementation as an hypoglycaemic and antihypertensive agent. Beetroot is a rich source of calcium, consumption of 100 g of beets powdered leaves meets daily recommendations as prescribed by FDA (1000 mg/day). Beetroot is also a rich source of Vitamin C, helps in immune function and is a great source of folate (vitamin B9) which helps cells and tissues to grow and function. Nitric oxide in beets acts as a vasodilator, thereby reducing blood pressure. It is used as a natural food colorant and labeled as E-162. The peptide present in beetroot is a promising drug against neurodegenerative disorders such as Alzheimer and Multiple Sclerosis. | In silico, in vitro and in vivo (clinical trials) | [13,64,65,66] |
| Oruwo/ Morinda Lucida Benth. Senegal to Sudan and southward to Angola and Zambia | Unknown cysteine stabilized peptides (CSP) | The root, stem bark and leaves of M. Lucida has been used traditionally in the treatment of malaria, sickle cell disease, hypertension, diabetes and other diseases. The antioxidants Vitamin A and E in M. lucida are used in battling degenerative disease like atherosclerosis. Vitamin K can be found in high quantities in the leaves of M. Lucida and helps in building strong bones. Its dietary fiber plays an important role in satisfying human needs for energy The bitter tasting roots are used as flavoring for food and alcohol. The wide range of enzyme and vitamins present in Morinda lucida makes it useful in treatment of different disease conditions. | In vitro, in vivo and in silico | [67] |
| Roselle/ Hibiscus sabdariffa L. Angola | Roseltide | H. sabdariffa is used as food coloring in food industries by incorporating their calyces. Extractions or infusions of dried calyces of zobo flower are used as tea (beverage) to lower body temperature, blood pressure, blood sugar and cholesterol. The leaves and calyces are eaten with pulverized peanuts in Africa as a side dish. It is used a good source of vitamin C and iron It is used with lemon balm and St John’s wort to treat restlessness and poor onset of sleep. Hibiscus tea is used as a mild laxative. | In silico, in vitro and in vivo (clinical trials) | [68] |
| Pea/ Pisum sativum L. Middle East specifically to Turkey and Iraq | PA1b (Pea Albumin 1 Subunit b) | The major components of peas are protein, starch and fiber which help in breaking down carbohydrates, hence are beneficial in the prevention and management of type-2 diabetes. Coumestrol in peas helps to lower the risk of stomach cancer. Addition of peas to food in combination with inulin fiber supplement greatly increases bowel movement frequency. It is a functional ingredient in vegan foods and dried seeds of pea are used as staple foods. Eating green peas regularly helps to lower blood pressure and blood cholesterol level due to the mineral content (Mg, K and Ca) present in them. | In silico, in vitro and in vivo | [39,69] |
| YinXing/ Ginkgo biloba L. China | β-Ginkgotides | Ginkgo-steamed eggs and Ginkgo-fried chicken are popular treats in Southeast Asia. Moreover, G. biloba nuts are used as a side dish in Japan. G. biloba seeds are largely encouraged to be used as nutraceuticals for prevention of neurodegenerative diseases. G. biloba seeds entail a higher percentage of vitamin C, carbohydrates, riboflavin, proteins and other nutrients. G. biloba seeds are incorporated in alcohols, drinks, glazed vegetables, cakes, sweets and other delicacies. | In silico, in vitro and in vivo (clinical trials) | [70] |
| Korean ginseng/ Panax ginseng C.A.Mey China | Ginsenosides | Ginseng tea, the decoction of ginseng roots is a natural home remedy to manage hypertension. It is used in weight loss management. Ginsenosides present in ginseng are chemically similar to female steroidal hormones, hence the tea helps to restore hormonal balance in women. Ginseng is used as a spice to promote skin health due to the presence of free radical fighting antioxidants. Ginseng tea and wine improve sexual health in men. Use of ginseng improves erectile function, sexual desire and intercourse satisfaction. Studies have shown the efficacy of P. ginseng in Chronic Fatigue Syndrome, normal fatigue, cancer related fatigue and postoperative fatigue syndrome. | In silico, in vitro and in vivo (clinical trials) | [71,72,73,74,75,76] |
2.2. Pharmaceutical Applications of Phytomedicines Rich in Knottins
2.3. Antimicrobial
2.4. Anticancer
2.5. Antiviral
2.6. Antifungal
| # Plant Source | Knottins | Pharmaceutical Application | IC50/EC50/Dose | Reference |
|---|---|---|---|---|
| Hedyotis biflora Hornem. Accepted name: Leptopetalum biflorum (L.) Neupane and N.Wikstr. | Hedyotide B1 (HB1), B10 and B11 | Hedyotide B1 possessed strong In vitro antibacterial activity against E. coli and S. salivarius, while Hedyotide B10 and B11 were active against S. aureus and S. epidermidis. | HB1 MIC: 3.4–5.9 μM HB10 and 11: MIC 1.5–2.2 μM | [80] |
| Viola odorata var. variegata DC | Cycloviolacin O2, O3, O19 | Broad spectrum antibacterial activity, CyO19 being the most potent. Cy O2 showed strong in vivo activity in surgical S. aureus wound infection mouse model | CyO19MIC 10 μM against E. coli, P. aeruginosa and S. aureus. S. aureus MIC 25 μM (in vivo) | [82,83] |
| Oldenlandia affinis (Roem. and Schult.) DC. | Kalata B1, B2, B7 | KB1 displayed moderate activity against P. aeruginosa, while KB7 displayed potent activity. KB2 showed mild in vivo activity in the surgical S. aureus wound infection mouse model. A. salmonicida, A. hydrophila, V. anguillarum, V. ordalii, F. psychrophilum | KB1 MIC 40 μM KB7 MIC 1.25 μM KB2 MIC 50 μM | [82,83] |
| Viola arvensis Murray | VarcA | Antimicrobial activity against Gram-negative bacterial infection in aquaculture. (A. salmonicida, A. hydrophila, V. anguillarum, V. ordalii, F. psychrophilum) | MIC 12.5–30.0 μM | [86] |
| Sambucus nigra L. | - | Antimicrobial activity against Gram-negative bacteria affecting aquaculture. | 100 µg/mL gave the strongest antimicrobial activity. | [25,86] |
| Clitoria ternatea L. | Cationic cT15, cT19 and cT20 | Potent activity against Gram-negative bacteria (E. coli), antibacterial activity diminished by high salt concentration, except for cT15. Immunomodulating activity presents an opportunity to be used as an adjuvant in vaccines. | MIC 0.5–0.6 μM (low salt); cT15, cT19 and cT20 MIC 2.5 μM, >10 and >20, respectively, in high salt concentration. | |
| Momordica cochinchinensis (Lour.) Spreng. | MCoTI-II | The linear 9mer peptide grafted into the loop 6 of MCoTI-II showed reduced antibacterial activity but extreme stability in 100% human serum. Antimicrobial activity of the cyclotides was also slightly reduced in 10% human serum. | P. aeruginosa 0.8 and 9 for linear and cyclic peptide, respectively, in 1/5 muller–hilton E. coli 0.4–0.8 and 4.0–9.0 for linear and cyclic peptide, respectively, in 1/5 muller–hilton Klebsiella pneumonia 1.5 and 71 for linear and cyclic peptide, respectively, in 1/5 muller–Hilton. | [116] |
| Hedyotis biflora Hornem. Accepted name: Leptopetalum biflorum (L.) Neupane and N.Wikstr. | Hedyotide B7 | Potent in both in vitro and in vivo experiments against four different pancreatic cell lines. | IC50 of 0.68, 0.45, 0.33 and 0.36 μM against BxPC3, Capan2, MOH-1 and PANC1, respectively. | [91] |
| Psychotria leptothyrsa Accepted name: Psychotria defretesiana W.N.Takeuchi | Psyle A, C and E | Activity on the Human Breast Adenocarcinoma Cell Line (MCF-7) and Its Drug Resistant Subline (MCF-7/ADR) similar to that of CyO2, psyle E is the most potent, and could be combined with conventional anticancer drugs to facilitate drug absorption by cancer cells. | Psyle E IC50 0.64, 1.73 and 0.39 μM against MCF-7, MCF-7/ADR and co-exposure to doxorubicin, respectively. | [90] |
| Hedyotis diffusa Accepted name: Scleromitrion diffusum (Willd.) R.J.Wang | DC1, DC2 and DC3 | Potent cytotoxic activity against human prostate cancer xenografts in both in vivo and in vitro experiments. DC3 is the most potent. | DC3 IC50 of 0.21, 0.76, 0.55 μM against LNcap, PC3, and DU145, respectively. | |
| Melicytus latifolius (Endl.) P.S.Green | Lysine-rich cyclotide: Mela 1–7, Mech 2 and 3. | Strong cytotoxic activity with significantly low hemolytic activity even at high peptide concentration. Their distinct toxicity properties from the usual cyclotide makes them a promising scaffold for drug design. | mela 1 CC50 of 2.09 ± 0.18, 3.07 ± 0.15, 9.83 ± 0.78 and >64 μM against MM96L, HFF-1, HELA and RBCs. | [93] |
| Viola ignobilis Rupr. Accepted name: Viola odorata subsp. odorata | Vigno 5 | Cytotoxic against Hela cells in a dose-dependent manner. | cell viability reduced significantly in a dose dependent manner (2.5, 5.0, 7.5, and 10 μM) to 48%, 35%, 24% and 18% of total, respectively. | [94] |
| Viola odorata L. Accepted name: Viola odorata subsp. odorata | CyO2 | Potent cytotoxic activity against breast cancer cell line MCF-7 and its drug resistance subline MCF-7/ADR. Selectively enhances the uptake of chemotherapeutic drug doxorubicin by cancer cells due to increased pore formation. | CyO2 IC50 3.17, 3.27 and 0.76 μM against MCF-7, MCF-7/ADR and co-exposure to doxorubicin, respectively. | [90] |
| Momordica cochinchinensis (Lour.) Spreng. Momordica cochinchinensis and Spinacia oleracea L. | MCoTI-II MCoTI-Var. 4 SOTI-Var. 1 | MCoTI-II scaffold has been employed in the design of knotting stabilized anticancer drugs because it lacks hemolytic activity against erythrocytes. Have high-affinity inhibitory activity against human matrypase-1 present on the surface of human prostate carcinoma cancer cells (PC-3). | - MCoTI-Var. 4 IC50 0.21 ± 0.11 μM SOTI-Var. 1 IC50 >10 μM | [117,118,119] |
| Clitoria ternatea L. | Cliotide T1 and T4 | Potent cytotoxic activity against HeLa cell line However, hemolytic activity occurs at lower doses compared to Cliotide T2 and T3. | IC50 0.6 μM for Cliotide T1 and T4 HD50 7.1, >100, 13.1 and 8.4 μM for Cliotide T1, T2, T3 and T4, respectively. | [80] |
| Chassalia Parvifolia K.Schum | Circulin A and B | First reported antiviral activity against HIV. | IC50 500 nM and EC50 about 40 to 260 nM in all assays. | [99] |
| Oldenlandia affinis (Roem. and Schult.) DC. | Kalata B1, kalata B1-inspired peptide | KB1 had strong antiviral activity against HIV through disruption of viral membrane envelope. Anti-HIV activity 25 times lower than its cytotoxic concentration, very low chance of emergence of resistance. In contrast, kalata B1-inspired peptide produced from the amino acid modification of KB1 displayed significant activity against Dengue Virus. | VC50 of 2.04 μm for HIV-1 NL4.3 and 4.54 μm for HIV-1 Clade A No free access to the article. | [104,105,106] |
| Alstonia scholaris (L.) R.Br. | Altoids As1 and As3 | Significant membrane permeability and early phase antiviral activity against infection of bronchitis virus (IBV) and Dengue infection (DENV2). Highly stable in the metabolic environment, target specificity and non-cytotoxic even at high peptide concentration made them a good candidate for drug design. | Pretreatment with As1 and As3 inhibits plaque formation in a dose-dependent manner with estimated EC50 of 35 and 55 μm, respectively, against IBV. EC50 of ∼90 μm for As1 against DENV2 NS3 protein expression in the infected cultures. | [107] |
| Viola odorata L. | CyO2 | CyO2 alone decreased HIV-1 infected HuT78 cells and co-exposure increased the anti-HIV activity of Saquinavir and Nelfinavir. | Short term exposure to CyO2 (0.5 μM) alone also showed a 70–80% reduction in HIV p24 levels; while continuous co-exposure to SQV (0.004 μM) and CyO2 (0.025 μM) resulted in significant suppression (~70–80%) in HIV-1 p24 levels. | [90] |
| Viola odorata L. | CyO2, CyO3, CyO19 | Potent activity against C. albicans | 99% activity seen at Minimum Fungicidal Concentration (MFC) of 10 µM. | [82] |
| Viola odorata L. | CyO2, CyO3, CyO19, CyO8 | Potent activity against fungal plant pathogens. Currently being investigated as a potential bio-fungicide against Fusarium head blight in crops | Most potent is cyO3 (MICs ranging from 0.8 to 12.5 μM), and the least active was cyO13 (MICs ranging from 3 to 25 μM). | [111,112] |
| Oldenlandia affinis (Roem. and Schult.) DC. | kalata B1 mutant [T20K] | Immunosuppressant effect is currently under clinical trial for the management of multiple sclerosis. | Single administration of T20K to healthy animals did not exhibit toxic effects up to the doses of 15 mg/kg (i.v), 75 mg/kg (i.p) and 250 mg/kg (p.o). | [20,120] |
| Clitoria ternatea L. | Cationic cliotides cT7 and cT19 | Strong immunostimulating effect by enhancing the release of cytokines and chemokines. | [26] | |
| Viola philippica Cav. | Kalata B1, Varv A, Viba 15, Viba 17, VarvE and Viphi G | Antibiofilm activity using S. aureus was reported for Viba 17, VarvE and Viphi G with Viphi G being the most potent. | At a concentration of 20 µg/mL, Viphi G inhibited bacterial growth more than 40%, while Viba 17 and Varv E only caused inhibition of 10–20%. | [121] |
| Tragia benthamii Baker | Knottin-rich biologically active peptides detected but not yet characterized. | Strong anti-inflammatory activity in vivo chick model. | Extract (30–300 mg/kg, i.p) produced a dose dependent reduction in foot edema with maximal inhibition of 84.3% at 300 mg/kg body weight. | [77] |
| Ecballium elaterium Accepted name: Ecballium elaterium subsp. dioicum (Batt.) Costich | Ecballium elaterium trypsin inhibitor II (EETI-II) | When engineered with scaffold proteins, were found to inhibit the binding of fibrinogen to αIIbβ3 in vitro. Engineered peptides bind to integrin expressing tumor cells. Could be employed both in the treatment and diagnosis of cancer. | Microprotein ETRGD1 with IC50 4.50 ± 0.70 nM. IC50 20 times lower than that of the standard drug eptifibatide. Echistatin is the most potent with IC50 of 4.9 ± 1.0 nM in binding assay; 9.2 ± 1.5 nM and 0.90 ± 0.2 nM cell adhesion assay using fibronectin and vitronectin, respectively. | [122,123] |
| Oldenlandia affinis (Roem. and Schult.) DC. | Kalata B1 | In bioengineering, they become excellent scaffolds for radioactive imaging probes to be used in positron emission tomography (PET), single photon emission computed tomography (SPECT). These scaffolds have also been combined with non-radioactive probes for fluorescence and ultrasound imaging. Useful as drug delivery vehicles for bioactive therapeutic peptides. | [23,124] | |
| Momordica cochinchinensis (Lour.) Spreng. | Cystine-knot peptide | This peptide has been engineered to bind to cytotoxic T Lymphocyte-associated antigen-4 (CTLA-4) which happens to be a target in the treatment of metastatic melanoma. | [125] |
3. Limitations to the Pharmaceutical Applications of Knottin-Rich Peptides in Clinical Settings
| Knottins/Phytomedicine | Source | Clinical Trial | IC50/Dose | Reference |
|---|---|---|---|---|
| T20K | Oldenlandia affinis | Multiple sclerosis | 2.7 ± 0.62 µM S.D | [120] |
| Integrin αvβ6 PET tracers (R01-MG) | Engineered peptides | imaging | 0.56 ± 0.46 nM S.D | [122] |
| Herbal Cyclotide Complex syrup | Cyclopeptide fraction with Ziziphus spina-cristi and Pimpinella anisum hydroalcoholic extract and orange peel | Post-exposure prevention of infection with COVID-19. | Dose = 20 mL every 8 h for 14 days | Clinical Trial Protocol Iranian Registry of Clinical Trials (https://www.irct.ir/trial/49925, accessed on 27 April 2022) |
| Concentrated Beet Root Juice | Knottin peptide-rich juice of the root of Beta vulgaris | Aortic and brachial blood pressure over 24 h | 70 mL concentrated beetroot juice drink | [133] |
| Concentrated Beet Root Juice | Knottin peptide-rich juice of the root of Beta vulgaris | Inflammatory markers and oxidative stress in patients with type 2 diabetes | 12 mL concentrated beetroot juice twice daily | [134] |
| Zufa syrup (Cyclotide-rich Polyherbal extract) | Herbal mixture containing three plants including the cyclotide-rich Viola odorata | symptomatic benign prostate hyperplasia (BPH) men | 7.5 mL of syrup every 4 h for 10 days | [135] |
| Knottin peptide-rich Gac extract | Momordica cochinchinensis | antiwrinkle agent | 5% Gac extract in cream applied to the entire face twice daily | [136] |
| Knottin peptide-rich Alkaloid extract | Alstonia scholaris | bronchitis, post-infectious cough and asthma. | 8–480 mg, three times daily for 7 days | [137] |
| Flower extract of Clitoria ternatea | Clitoria ternatea | glycemic response and antioxidant capacity | 1 g–2 g in 400 mL of water | [138] |
| Knottin peptide-rich Moringa oleifera leaf powder divided twice daily in corn porridge Moringa oleifera leaf powder | Moringa oleifera | maternal and infant healthsupplementation on the immune status and anthropometric parameters of adult HIV patients | 20 g daily for 3 months15 g Moringa leaf powder sachets (5 g each for 3 times) | [139,140] |
4. Materials and Methods
5. Conclusions and Prospects
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fernandez, A.R.; Sáez, A.; Quintero, C.; Gleiser, G.; Aizen, M.A. Intentional and unintentional selection during plant domestication: Herbivore damage, plant defensive traits and nutritional quality of fruit and seed crops. New Phytol. 2021, 231, 1586–1598. [Google Scholar] [CrossRef]
- Martínez-Romero, E.; Aguirre-Noyola, J.L.; Taco-Taype, N.; Martínez-Romero, J.; Zuñiga-Dávila, D. Plant microbiota modified by plant domestication. Syst. Appl. Microbiol. 2020, 43, 126106. [Google Scholar] [CrossRef] [PubMed]
- Wright, L. Wilderness into Civilized Shapes: Reading the Postcolonial Environment; University of Georgia Press: Athens, GA, USA, 2010; p. 213. [Google Scholar]
- Attah, A.F.; Akindele, O.O.; Nnamani, P.O.; Jonah, U.J.; Sonibare, M.A.; Moody, J.O. Moringa oleifera seed at the interface of food and medicine: Effect of extracts on some reproductive parameters, hepatic and renal histology. Front. Pharmacol. 2022, 13, 816498. [Google Scholar] [CrossRef] [PubMed]
- Anywar, G. Historical use of toxic plants. In Poisonous Plants and Phytochemicals in Drug Discovery; Wiley: Hoboken, NJ, USA, 2020; pp. 1–17. [Google Scholar]
- Srivastava, S.; Dashora, K.; Ameta, K.L.; Singh, N.P.; El-Enshasy, H.A.; Pagano, M.C.; Hesham, A.E.; Sharma, G.D.; Sharma, M.; Bhargava, A. Cysteine-rich antimicrobial peptides from plants: The future of antimicrobial therapy. Phytother. Res. 2020, 35, 256–277. [Google Scholar] [CrossRef] [PubMed]
- Hussein, R.A.; El-Anssary, A.A. Plants secondary metabolites: The key drivers of the pharmacological actions of medicinal plants. In Herbal Medicine; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Huang, Y.H.; Du, Q.; Craik, D.J. Cyclotides: Disulfide-rich peptide toxins in plants. Toxicon 2019, 172, 33–44. [Google Scholar] [CrossRef]
- Postic, G.; Gracy, J.; Périn, C.; Chiche, L.; Gelly, J.C. Knottin: The database of inhibitor cystine knot scaffold after 10 years, toward a systematic structure modeling. Nucleic Acids Res. 2017, 46, D454–D458. [Google Scholar] [CrossRef]
- Almeida, É.S.; de Oliveira, D.; Hotza, D. Properties and applications of Morinda citrifolia (noni): A review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 883–909. [Google Scholar] [CrossRef]
- Oguis, G.K.; Gilding, E.K.; Jackson, M.A.; Craik, D.J. Butterfly pea (Clitoria ternatea), a cyclotide-bearing plant with applications in agriculture and medicine. Front. Plant Sci. 2019, 10, 645. [Google Scholar] [CrossRef]
- Moyer, T.B.; Brechbill, A.M.; Hicks, L.M. Mass spectrometric identification of antimicrobial peptides from medicinal seeds. Molecules 2021, 26, 7304. [Google Scholar] [CrossRef]
- Retzl, B.; Hellinger, R.; Muratspahić, E.; Pinto, M.E.F.; Bolzani, V.S.; Gruber, C.W. Discovery of a beetroot protease inhibitor to identify and classify plant-derived cystine knot peptides. J. Nat. Prod. 2020, 83, 3305–3314. [Google Scholar] [CrossRef]
- Craik, D.J.; Daly, N.L.; Waine, C. The cystine knot motif in toxins and implications for drug design. Toxicon 2000, 39, 43–60. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.J.; Leung, C.L.; Cochran, J.R. Knottins: Disulfide-bonded therapeutic and diagnostic peptides. Drug Discov. Today Technol. 2012, 9, e3–e11. [Google Scholar] [CrossRef] [PubMed]
- Kumari, G. Discovery and Application of Cysteine-Rich Peptides from Medicinal Plants in Drug Development. Ph.D. Thesis, Nanyang Technological University, Singapore, 2017. [Google Scholar] [CrossRef]
- Molesini, B.; Treggiari, D.; Dalbeni, A.; Minuz, P.; Pandolfini, T. Plant cystine-knot peptides: Pharmacological perspectives. Br. J. Clin. Pharmacol. 2017, 83, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Weidmann, J.; Craik, D.J. Discovery, structure, function, and applications of cyclotides: Circular proteins from plants. J. Exp. Bot. 2016, 67, 4801–4812. [Google Scholar] [CrossRef]
- Huang, J.; Wong, K.H.; Tay, S.V.; Serra, A.; Sze, S.K.; Tam, J.P. Astratides: Insulin-modulating, insecticidal, and antifungal cysteine-rich peptides from Astragalus membranaceus. J. Nat. Prod. 2019, 82, 194–204. [Google Scholar] [CrossRef]
- Gründemann, C.; Stenberg, K.G.; Gruber, C.W. T20K: An immunomodulatory cyclotide on its way to the clinic. Int. J. Pept. Res. Ther. 2018, 25, 9–13. [Google Scholar] [CrossRef]
- Slavokhotova, A.A.; Shelenkov, A.A.; Andreev, Y.A.; Odintsova, T.I. Hevein-like antimicrobial peptides of plants. Biochemistry 2017, 82, 1659–1674. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.; Tan, W.L.; Serra, A.; Xiao, T.; Sze, S.K.; Yang, D.; Tam, J.P. Ginkgotides: Proline-rich hevein-like peptides from gymnosperm Ginkgo biloba. Front. Plant Sci. 2016, 7, 1639. [Google Scholar] [CrossRef] [Green Version]
- Daly, N.L.; Craik, D.J. Bioactive cystine knot proteins. Curr. Opin. Chem. Biol. 2011, 15, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Chai, T.-T.; Tan, Y.-N.; Ee, K.-Y.; Xiao, J.; Wong, F.-C. Seeds, fermented foods, and agricultural by-products as sources of plant-derived antibacterial peptides. Crit. Rev. Food Sci. Nutr. 2019, 59, S162–S177. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.N.T.; Nguyen, G.K.T.; Nguyen, P.Q.T.; Ang, K.H.; Dedon, P.C.; Tam, J.P. Immunostimulating and gram-negative-specific antibacterial cyclotides from the butterfly pea (Clitoria ternatea). FEBS J. 2016, 283, 2067–2090. [Google Scholar] [CrossRef] [PubMed]
- Gressent, F.; de Silva, P.; Eyraud, V.; Karaki, L.; Royer, C. Pea albumin 1 subunit b (PA1b), a promising bioinsecticide of plant origin. Toxins 2011, 3, 1502–1517. [Google Scholar] [CrossRef] [PubMed]
- Kam, A.; Loo, S.; Fan, J.S.; Sze, S.K.; Yang, D.; Tam, J.P. Roseltide rT7 Is a Disulfide-Rich, Anionic, and Cell-Penetrating Peptide That Inhibits Proteasomal Degradation. ASBMB. Available online: https://www.jbc.org/article/S0021-9258(20)30275-1/abstract (accessed on 30 August 2022).
- Kini, S.G.; Wong, K.H.; Tan, W.L.; Xiao, T.; Tam, J.P. Morintides: Cargo-free chitin-binding peptides from Moringa oleifera. BMC Plant Biol. 2017, 17, 68. [Google Scholar] [CrossRef] [PubMed]
- Moyer, T.B.; Schug, W.J.; Hicks, L.M. Amaranthus hypochondriacus seeds as a rich source of cysteine rich bioactive peptides. Food Chem. 2022, 377, 131959. [Google Scholar] [CrossRef] [PubMed]
- Culver, K.D.; Allen, J.L.; Shaw, L.N.; Hicks, L.M. Too hot to handle: Antibacterial peptides identified in ghost pepper. J. Nat. Prod. 2021, 84, 2200–2208. [Google Scholar] [CrossRef]
- Oliveira, A.P.B.F.; Resende, L.M.; Rodrigues, R.; Mello, D.O.; Taveira, G.B.; Carvalho, A.D.O.; Gomes, V.M. Antimicrobial peptides of the genus Capsicum: A mini review. Hortic. Environ. Biotechnol. 2022, 63, 453–466. [Google Scholar] [CrossRef]
- Tan, J.; Ni, D.; Taitz, J.; Pinget, G.V.; Read, M.; Senior, A.; Wali, J.A.; Elnour, R.; Shanahan, E.; Wu, H.; et al. Dietary protein increases T-cell-independent sIgA production through changes in gut microbiota-derived extracellular vesicles. Nat. Commun. 2022, 13, 4336. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, S.; Jian, W.; Xie, C.; Yang, X. Plant antimicrobial peptides: Structures, functions, and applications. Bot. Stud. 2021, 62, 5. [Google Scholar] [CrossRef]
- Ngadni, M.; Akhtar, M.; Ismail, I.; Norazhar, A.; Lee, S.; Maulidiani, M.; Shaari, K. Clitorienolactones and isoflavonoids of clitorea ternatea roots alleviate stress-like symptoms in a reserpine-induced zebrafish model. Molecules 2021, 26, 4137. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Functional amino acids in nutrition and health. Amino Acids 2013, 45, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Ezzudin, R.M.; Rabeta, M.S. A potential of telang tree (Clitoria ternatea) in human health. Food Res. 2018, 2, 415–420. [Google Scholar] [CrossRef]
- Turnos, L.J.N. Blue Ternate (Clitoria Ternatea L.): Nutritive Analysis of Flowers and Seeds. Asian Journal of Fundamental and Applied Sciences. Available online: https://myjms.mohe.gov.my/index.php/ajfas/article/view/14510 (accessed on 31 August 2022).
- Rungruangmaitree, R.; Jiraungkoorskul, W. Pea, Pisum sativum, and its anticancer activity. Pharmacogn. Rev. 2017, 11, 39–42. [Google Scholar] [CrossRef] [PubMed]
- Dahl, W.J.; Foster, L.M.; Tyler, R.T. Review of the health benefits of peas (Pisum sativum L.). Br. J. Nutr. 2012, 108, S3–S10. [Google Scholar] [CrossRef] [PubMed]
- Barashkova, A.S.; Rogozhin, E.A. Isolation of antimicrobial peptides from different plant sources: Does a general extraction method exist? Plant Methods 2020, 16, 143. [Google Scholar] [CrossRef] [PubMed]
- Abou-Arab, A.; Abu-Salem, F.; Abou-Arab, E.A. Physico- chemical properties of natural pigments (anthocyanin) extracted from Roselle calyces (Hibiscus subdariffa). J. Am. Sci. 2011, 7, 445–456. [Google Scholar]
- Riaz, G.; Chopra, R. A review on phytochemistry and therapeutic uses of Hibiscus sabdariffa L. Biomed. Pharmacother. 2018, 102, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Loo, S.; Kam, A.; Xiao, T.; Nguyen, G.K.T.; Liu, C.F.; Tam, J.P. Identification and characterization of roseltide, a knottin-type neutrophil elastase inhibitor derived from Hibiscus sabdariffa. Sci. Rep. 2016, 6, 39401. [Google Scholar] [CrossRef]
- Hamedi, S.; Honarvar, M. Beta vulgaris—A mini review of traditional uses in Iran, phytochemistry and pharmacology. Curr. Drug Discov. Technol. 2019, 16, 74–81. [Google Scholar] [CrossRef]
- Shyamala, B.N.; Jamuna, P. Nutritional content and antioxidant properties of pulp waste from Daucus carota and Beta vulgaris. Mal. J. Nutr. 2010, 16, 397–408. Available online: https://nutriweb.org.my/mjn/publication/16-3/h.pdf (accessed on 30 August 2022).
- Wruss, J.; Waldenberger, G.; Huemer, S.; Uygun, P.; Lanzerstorfer, P.; Müller, U.; Höglinger, O.; Weghuber, J. Compositional characteristics of commercial beetroot products and beetroot juice prepared from seven beetroot varieties grown in Upper Austria. J. Food Compos. Anal. 2015, 42, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Baraniak, J.; Kania-Dobrowolska, M. The dual nature of amaranth—Functional food and potential medicine. Foods 2022, 11, 618. [Google Scholar] [CrossRef] [PubMed]
- Trucco, F.; Tranel, P.J. Amaranthus. In Wild Crop Relatives: Genomic and Breeding Resources; Springer: Berlin/Heidelberg, Germany, 2011; pp. 11–21. [Google Scholar] [CrossRef]
- Gruber, C.W.; Čemažar, M.; Anderson, M.A.; Craik, D.J. Insecticidal Plant Cyclotides and Related Cystine Knot Toxins; Elsevier: Amsterdam, The Netherlands, 2007; Available online: https://www.sciencedirect.com/science/article/pii/S004101010600434X (accessed on 30 August 2022).
- Carugo, O.; Lu, S.; Luo, J.; Gu, X.; Liang, S.; Strobl, S.; Pongor, S. Structural analysis of free and enzyme-bound amaranth α-amylase inhibitor: Classification within the knottin fold superfamily and analysis of its functional flexibility. Protein Eng. Des. Sel. 2001, 14, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, A.; Sher, R.; Kashmala, K.; Anum, S.; Faryal, I.; Syeda, A.; Shah, S.; Mii, M. Antimicrobial peptides as effective tools for enhanced disease resistance in plants. Plant Cell Tissue Organ. Cult. 2019, 139, 1–15. [Google Scholar] [CrossRef]
- Anaya-López, J.L.; López-Meza, J.E.; Baizabal-Aguirre, V.M.; Cano-Camacho, H.; Ochoa-Zarzosa, A. Fungicidal and cytotoxic activity of a Capsicum chinense defensin expressed by endothelial cells. Biotechnol. Lett. 2006, 28, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.-L.; Cai, S.-Y.; Gao, M.; Chu, X.-M.; Pan, X.-Y.; Gong, K.-K.; Xiao, C.-W.; Chen, Y.; Zhao, Y.-Q.; Wang, B.; et al. Purification of Antioxidant Peptides of Moringa oleifera Seeds and Their Protective Effects on H2O2 Oxidative Damaged Chang Liver Cells; Elsevier: Amsterdam, The Netherlands, 2020; Available online: https://www.sciencedirect.com/science/article/pii/S175646461930622X (accessed on 30 August 2022).
- Kolmar, H. Engineered cystine-knot miniproteins for diagnostic applications. Expert Rev. Mol. Diagn. 2010, 10, 361–368. [Google Scholar] [CrossRef] [PubMed]
- García-Moreno, P.J.; Gregersen, S.; Nedamani, E.R.; Olsen, T.H.; Marcatili, P.; Overgaard, M.T.; Andersen, M.L.; Hansen, E.B.; Jacobsen, C. Identification of emulsifier potato peptides by bioinformatics: Application to omega-3 delivery emulsions and release from potato industry side streams. Sci. Rep. 2020, 10, 690. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, D.; Sun, C.; McClements, D.J.; Gao, Y. Utilization of interfacial engineering to improve physicochemical stability of β-carotene emulsions: Multilayer coatings formed using protein and protein-polyphenol conjugates. Food Chem. 2016, 205, 129–139. [Google Scholar] [CrossRef]
- Faustino, M.; Veiga, M.; Sousa, P.; Costa, E.M.; Silva, S.; Pintado, M. Agro-food byproducts as a new source of natural food additives. Molecules 2019, 24, 1056. [Google Scholar] [CrossRef] [PubMed]
- Afrianto, W.F.; Tamnge, F.; Hasanah, L.N. Review: A relation between ethnobotany and bioprospecting of edible flower Butterfly Pea (Clitoria ternatea) in Indonesia. Asian J. Ethnobiol. 2020, 3. [Google Scholar] [CrossRef]
- Xiong, T.; Xiong, W.; Ge, M.; Xia, J.; Li, B.; Chen, Y. Effect of high intensity ultrasound on structure and foaming properties of pea protein isolate. Food Res. Int. 2018, 109, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Hidayat, B.T.; Wea, A.; Andriati, N. Physicochemical, sensory attributes and protein profile by SDS-PAGE of beef sausage substituted with texturized vegetable protein. Food Res. 2017, 2, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Babarykin, D.; Smirnova, G.; Pundinsh, I.; Vasiljeva, S.; Krumina, G.; Agejchenko, V. Red beet (Beta vulgaris) impact on human health. J. Biosci. Med. 2019, 7, 61–79. [Google Scholar] [CrossRef]
- Mondal, S.; Moktan, S. A paradoxically significant medicinal plant carapichea ipecacuanha: A review. Indian J. Pharm. Educ. Res. 2020, 54, s56–s66. [Google Scholar] [CrossRef]
- Karami, Z.; Akbari-adergani, B. Bioactive food derived peptides: A review on correlation between structure of bioactive peptides and their functional properties. J. Food Sci. Technol. 2019, 56, 535–554. [Google Scholar] [CrossRef] [PubMed]
- Mirmiran, P.; Houshialsadat, Z.; Gaeini, Z.; Bahadoran, Z.; Azizi, F. Functional properties of beetroot (Beta vulgaris) in management of cardio-metabolic diseases. Nutr. Metab. 2020, 17, 3. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, G.R.; Milani, T.M.G.; Trinca, N.R.R.; Nagai, L.Y.; de Barretto, A.C. Textured soy protein, collagen and maltodextrin as extenders to improve the physicochemical and sensory properties of beef burger. Food Sci. Technol. 2017, 37, 10–16. [Google Scholar] [CrossRef]
- Zaky, A.A.; Chen, Z.; Qin, M.; Wang, M.; Jia, Y. Assessment of antioxidant activity, amino acids, phenolic acids and functional attributes in defatted rice bran and rice bran protein concentrate. Prog. Nutr. 2020, 22, e2020069. [Google Scholar] [CrossRef]
- Adewole, K.E.; Attah, A.F.; Adebayo, J.O. Morinda lucida Benth (Rubiaceae): A review of its ethnomedicine, phytochemistry and pharmacology. J. Ethnopharmacol. 2021, 276, 114055. [Google Scholar] [CrossRef]
- Patel, S. Hibiscus sabdariffa: An ideal yet under-exploited candidate for nutraceutical applications. Biomed. Prev. Nutr. 2014, 4, 23–27. [Google Scholar] [CrossRef]
- Rasskazova, I.; Kirse-Ozolina, A. Field pea Pisum sativum L. as a perspective ingredient for vegan foods: A review. Res. Rural Dev. 2020, 35, 125–131. [Google Scholar] [CrossRef]
- Hatano, K.-I.; Miyakawa, T.; Sawano, Y.; Tanokura, M. Antifungal and Lipid Transfer Proteins from Ginkgo (Ginkgo biloba) Seeds; Elsevier: Amsterdam, The Netherlands, 2011; Available online: https://www.sciencedirect.com/science/article/pii/B9780123756886100635 (accessed on 30 August 2022).
- Rhee, M.; Kim, Y.; Bae, J.; Nah, D.; Kim, Y.; Lee, M.; Kim, H. Effect of Korean red ginseng on arterial stiffness in subjects with hypertension: PP.35.479. J. Hypertens. 2010, 28, e592. [Google Scholar] [CrossRef]
- Wang, T.; Yu, X.F.; Qu, S.C.; Xu, H.L.; Sui, D.Y. Ginsenoside Rb3 inhibits angiotensin II-induced vascular smooth muscle cells proliferation. Basic Clin. Pharmacol. Toxicol. 2010, 107, 685–689. [Google Scholar] [CrossRef]
- Lee, H.; Choi, J.; Shin, S.S.; Yoon, M. Effects of Korean red ginseng (Panax ginseng) on obesity and adipose inflammation in ovariectomized mice. J. Ethnopharmacol. 2016, 178, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Ratan, Z.A.; Haidere, M.F.; Hong, Y.H.; Park, S.H.; Lee, J.-O.; Lee, J.; Cho, J.Y. Pharmacological potential of ginseng and its major component ginsenosides. J. Ginseng Res. 2020, 45, 199–210. [Google Scholar] [CrossRef]
- Hong, B.; Ji, Y.H.; Hong, J.H.; Nam, K.I.Y.; Ahn, T.Y. A double-blind crossover study evaluating the efficacy of Korean red ginseng in patients with erectile dysfunction: A preliminary report. J. Urol. 2002, 168, 2070–2073. [Google Scholar] [CrossRef]
- Lu, G.; Liu, Z.; Wang, X.; Wang, C. Recent advances in panax ginseng C.A. Meyer as a herb for anti-fatigue: An effects and mechanisms review. Foods 2021, 10, 1030. [Google Scholar] [CrossRef] [PubMed]
- Attah, A.F.; Omobola, A.I.; Moody, J.O.; Sonibare, M.A.; Adebukola, O.M.; Onasanwo, S.A. Detection of cysteine-rich peptides in Tragia benthamii Baker (Euphorbiaceae) and in vivo antiinflammatory effect in a chick model. Phys. Sci. Rev. 2021. [Google Scholar] [CrossRef]
- Tam, J.P.; Lu, Y.A.; Yang, J.L.; Chiu, K.W. An unusual structural motif of antimicrobial peptides containing end-to-end macrocycle and cystine-knot disulfides. Proc. Natl. Acad. Sci. USA 1999, 96, 8913–8918. [Google Scholar] [CrossRef]
- Rowe, S.M.; Spring, D.R. The role of chemical synthesis in developing RiPP antibiotics. Chem. Soc. Rev. 2021, 50, 4245–4258. [Google Scholar] [CrossRef]
- Nguyen, G.K.T.; Zhang, S.; Nguyen, N.T.K.; Nguyen, P.Q.T.; Chiu, M.S.; Hardjojo, A.; Tam, J.P. Discovery and characterization of novel cyclotides originated from chimeric precursors consisting of albumin-1 chain a and cyclotide domains in the fabaceae family. J. Biol. Chem. 2011, 286, 24275–24287. [Google Scholar] [CrossRef]
- Wang, C.K.L.; Colgrave, M.L.; Gustafson, K.R.; Ireland, D.C.; Goransson, U.; Craik, D.J. Anti-HIV cyclotides from the Chinese medicinal herb Viola yedoensis. J. Nat. Prod. 2008, 71, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Strömstedt, A.A.; Park, S.; Burman, R.; Göransson, U. Bactericidal activity of cyclotides where phosphatidylethanolamine-lipid selectivity determines antimicrobial spectra. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1986–2000. [Google Scholar] [CrossRef] [PubMed]
- Fensterseifer, I.C.; Silva, O.; Malik, U.; Ravipati, A.S.; Novaes, N.R.; Miranda, P.R.; Rodrigues, E.A.; Moreno, S.E.; Craik, D.; Franco, O.L. Effects of cyclotides against cutaneous infections caused by Staphylococcus aureus. Peptides 2014, 63, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Aboye, T.L.; Strömstedt, A.A.; Gunasekera, S.; Bruhn, J.G.; El-Seedi, H.; Rosengren, K.J.; Göransson, U. A cactus-derived toxin-like cystine knot peptide with selective antimicrobial activity. ChemBioChem 2015, 16, 1068–1077. [Google Scholar] [CrossRef]
- Álvarez, C.A.; Santana, P.A.; Luna, O.; Cárdenas, C.; Albericio, F.; Romero, M.S.; Guzmán, F. Chemical synthesis and functional analysis of VarvA cyclotide. Molecules 2018, 23, 952. [Google Scholar] [CrossRef]
- Álvarez, C.A.; Barriga, A.; Palomera, F.A.; Romero, M.S.; Guzmán, F. Identification of antimicrobial peptides in flowers of sambucus nigra. Molecules 2018, 23, 1033. [Google Scholar] [CrossRef]
- Weidle, U.H.; Auer, J.; Brinkmann, U.; Georges, G.; Tiefenthaler, G. The Emerging Role of New Protein Scaffold-Based Agents for Treatment of Cancer. cgp.iiarjournals.org. Available online: https://cgp.iiarjournals.org/content/10/4/155.short (accessed on 30 August 2022).
- Sottero, T.; Girard, E.J.; Correnti, C.; Stroud, M.R.; Kier, B.L.; Mhyre, A.J.; Olson, J. Abstract LB-231: An optide (optimized knottin-peptide) that inhibits tumor cell growth In vitro and accumulates in sarcoma flank tumors in vivo. Cancer Res. 2016, 76, LB-231. [Google Scholar] [CrossRef]
- Grover, T.; Mishra, R.; Bushra; Gulati, P.; Mohanty, A. An insight into biological activities of native cyclotides for potential applications in agriculture and pharmaceutics. Peptides 2020, 135, 170430. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, S.L.; Chandra, P.K.; Roy, U.; Gunasekera, S.; Göransson, U.; Wimley, W.C.; Braun, S.E.; Mondal, D. The membrane-active phytopeptide cycloviolacin O2 simultaneously targets HIV-1-infected cells and infectious viral particles to potentiate the efficacy of antiretroviral drugs. Medicines 2019, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Bai, D.; Qian, J. Novel cyclotides from Hedyotis biflora inhibit proliferation and migration of pancreatic cancer cell in vitro and in vivo. Med. Chem. Res. 2013, 23, 1406–1413. [Google Scholar] [CrossRef]
- Sun, G.; Wei, L.; Feng, J.; Lin, J.; Peng, J. Inhibitory effects of Hedyotis diffusa Willd. on colorectal cancer stem cells. Oncol. Lett. 2016, 11, 3875–3881. [Google Scholar] [CrossRef] [PubMed]
- Ravipati, A.S.; Henriques, S.T.; Poth, A.G.; Kaas, Q.; Wang, C.K.; Colgrave, M.L.; Craik, D.J. Lysine-rich cyclotides: A new subclass of circular knotted proteins from violaceae. ACS Chem. Biol. 2015, 10, 2491–2500. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, M.A.; Abagheri-Mahabadi, N.; Hashempour, H.; Farhadpour, M.; Gruber, C.W.; Ghassempour, A. Viola plant cyclotide vigno 5 induces mitochondria-mediated apoptosis via cytochrome C release and caspases activation in cervical cancer cells. Fitoterapia 2016, 109, 162–168. [Google Scholar] [CrossRef]
- Treggiari, D.; Zoccatelli, G.; Molesini, B.; Degan, M.; Rotino, G.L.; Sala, T.; Cavallini, C.; MacRae, C.A.; Minuz, P.; Pandolfini, T. A cystine-knot miniprotein from tomato fruit inhibits endothelial cell migration and angiogenesis by affecting vascular endothelial growth factor receptor (VEGFR) activation and nitric oxide production. Mol. Nutr. Food Res. 2015, 59, 2255–2266. [Google Scholar] [CrossRef]
- Engelhard, Y.N.; Gazer, B.; Paran, E. Natural antioxidants from tomato extract reduce blood pressure in patients with grade-1 hypertension: A double-blind, placebo-controlled pilot study. Am. Heart J. 2006, 151, 100.e1–100.e6. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Huang, Y.H.; Wang, C.K.; Kaas, Q.; Craik, D.J. Mutagenesis of bracelet cyclotide hyen D reveals functionally and structurally critical residues for membrane binding and cytotoxicity. J. Biol. Chem. 2022, 298. [Google Scholar] [CrossRef]
- Vilas Boas, L.C.P.; Campos, M.L.; Berlanda, R.L.A.; de Carvalho Neves, N.; Franco, O.L. Antiviral peptides as promising therapeutic drugs. Cell. Mol. Life Sci. 2019, 76, 3525–3542. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, K.R.; Sowder, R.C., II; Henderson, L.E.; Parsons, I.C.; Kashman, Y.; Cardellina, J.H., II; McMahon, J.B.; Buckheit, R.W., Jr.; Pannell, L.K.; Boyd, M.R. Circulins A and B: Novel HlV-inhibitory macrocyclic peptides from the tropical tree Chassalia parnifolia. J. Am. Chem. Soc. 1994, 116, 9337–9338. [Google Scholar] [CrossRef]
- Gustafson, K.R.; Walton, L.K.; Sowder, I.R.C.; Johnson, D.G.; Pannell, L.K.; Cardellina, I.J.H.; Boyd, M.R. New circulin macrocyclic polypeptides from Chassalia parvifolia. J. Nat. Prod. 2000, 63, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Hallock, Y.F.; Ii, R.C.S.; Pannell, L.K.; Hughes, C.B.; Johnson, D.G.; Gulakowski, R.; Ii, J.H.C.; Boyd, M.R. Cycloviolins A-D, anti-HIV macrocyclic peptides from Leonia cymosa. J. Org. Chem. 1999, 65, 124–128. [Google Scholar] [CrossRef]
- Bokesch, H.R.; Pannell, L.K.; Cochran, P.K.; Sowder, R.C.; McKee, T.C.; Boyd, M.R. A novel anti-HIV macrocyclic peptide from Palicourea condensata. J. Nat. Prod. 2001, 64, 249–250. [Google Scholar] [CrossRef] [PubMed]
- Henriques, S.T.; Huang, Y.-H.; Rosengren, K.J.; Franquelim, H.; Carvalho, F.; Johnson, A.; Sonza, S.; Tachedjian, G.; Castanho, M.; Daly, N.; et al. Decoding the membrane activity of the cyclotide kalata B1: The importance of phosphatidylethanolamine phospholipids and lipid organization on hemolytic and anti-HIV activities. J. Biol. Chem. 2011, 286, 24231–24241. [Google Scholar] [CrossRef] [PubMed]
- Henriques, S.T.; Craik, D.J. Cyclotides as templates in drug design. Drug Discov. Today 2010, 15, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Nawae, W.; Hannongbua, S.; Ruengjitchatchawalya, M. Molecular dynamics exploration of poration and leaking caused by Kalata B1 in HIV-infected cell membrane compared to host and HIV membranes. Sci. Rep. 2017, 7, 3638. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Cui, T.; Lam, Y. Synthesis and disulfide bond connectivity-activity studies of a kalata B1-inspired cyclopeptide against dengue NS2B-NS3 protease. Bioorganic Med. Chem. 2010, 18, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.Q.T.; Ooi, J.S.G.; Nguyen, N.T.K.; Wang, S.; Huang, M.; Liu, D.X.; Tam, J.P. Antiviral cystine knot α-amylase inhibitors from Alstonia scholaris. J. Biol. Chem. 2015, 290, 31138–31150. [Google Scholar] [CrossRef]
- Ireland, D.C.; Wang, C.K.L.; Wilson, J.A.; Gustafson, K.R.; Craik, D.J. Cyclotides as natural anti-HIV agents. Biopolymers 2008, 90, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Slazak, B.; Kapusta, M.; Strömstedt, A.; Słomka, A.; Krychowiak-Masnicka, M.; Shariatgorji, R.; Andrén, P.E.; Bohdanowicz, J.; Kuta, E.; Göransson, U. How does the sweet violet (Viola odorata L.) fight pathogens and pests—Cyclotides as a comprehensive plant host defense system. Front. Plant Sci. 2018, 9, 1296. [Google Scholar] [CrossRef]
- Parsley, N.C.; Kirkpatrick, C.L.; Crittenden, C.; Rad, J.G.; Hoskin, D.W.; Brodbelt, J.S.; Hicks, L.M. PepSAVI-MS reveals anticancer and antifungal cycloviolacins in Viola odorata. Phytochemistry 2018, 152, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Parisi, K.; Shafee, T.M.A.; Quimbar, P.; van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. The evolution, function and mechanisms of action for plant defensins. Semin. Cell Dev. Biol. 2019, 88, 107–118. [Google Scholar] [CrossRef]
- Hsiao, P.Y.; Cheng, C.P.; Koh, K.W.; Chan, M.T. The Arabidopsis defensin gene, AtPDF1.1, mediates defence against Pectobacterium carotovorum subsp. carotovorum via an iron-withholding defence system. Sci. Rep. 2017, 7, 9175. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Ng, T.B. Gymnin, a potent defensin-like antifungal peptide from the Yunnan bean (Gymnocladus chinensis Baill). Peptides 2003, 24, 963–968. [Google Scholar] [CrossRef]
- Wang, S.; Rao, P.; Ye, X. Isolation and biochemical characterization of a novel leguminous defense peptide with antifungal and antiproliferative potency. Appl. Microbiol. Biotechnol. 2009, 82, 79–86. [Google Scholar] [CrossRef]
- Eyraud, V.; Balmand, S.; Karaki, L.; Rahioui, I.; Sivignon, C.; Delmas, A.F.; Royer, C.; Rahbé, Y.; Da Silva, P.; Gressent, F. The interaction of the bioinsecticide PA1b (Pea Albumin 1 subunit b) with the insect V-ATPase triggers apoptosis. Sci. Rep. 2017, 7, 4902. [Google Scholar] [CrossRef] [PubMed]
- Koehbach, J.; Gani, J.; Hilpert, K.; Craik, D.J. Comparison of a short linear antimicrobial peptide with its disulfide-cyclized and cyclotide-grafted variants against clinically relevant pathogens. Microorganisms 2021, 9, 1249. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Majumder, S.; Millard, M.; Borra, R.; Bi, T.; Elnagar, A.Y.; Neamati, N.; Shekhtman, A.; Camarero, J.A. In vivo activation of the p53 tumor suppressor pathway by an engineered cyclotide. J. Am. Chem. Soc. 2013, 135, 11623–11633. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, C.; Henriques, S.T.; Wang, C.K.; Cheneval, O.; Chan, L.Y.; Bokil, N.J.; Sweet, M.J.; Craik, D.J. Using the MCoTI-II cyclotide scaffold to design a stable cyclic peptide antagonist of SET, a protein overexpressed in human cancer. Biochemistry 2016, 55, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Glotzbach, B.; Reinwarth, M.; Weber, N.; Fabritz, S.; Tomaszowski, M.; Fittler, H.; Christmann, A.; Avrutina, O.; Kolmar, H. Combinatorial optimization of cystine-knot peptides towards high-affinity inhibitors of human matriptase-1. PLoS ONE 2013, 8, e76956. [Google Scholar] [CrossRef] [PubMed]
- Hellinger, R.; Muratspahić, E.; Devi, S.; Koehbach, J.; Vasileva, M.; Harvey, P.J.; Craik, D.J.; Gründemann, C.; Gruber, C.W. Importance of the cyclic cystine knot structural motif for immunosuppressive effects of cyclotides. ACS Chem. Biol. 2021, 16, 2373–2386. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.; Yang, Y.; Uche, F.I.; Hart, S.R.; Li, W.-W.; Yuan, C. Coupling plant-derived cyclotides to metal surfaces: An antibacterial and antibiofilm study. Int. J. Mol. Sci. 2018, 19, 793. [Google Scholar] [CrossRef] [PubMed]
- Reiss, S.; Sieber, M.; Oberle, V.; Wentzel, A.; Spangenberg, P.; Claus, R.; Kolmar, H.; Lösche, W. Inhibition of platelet aggregation by grafting RGD and KGD sequences on the structural scaffold of small disulfide-rich proteins. Platelets 2006, 17, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Kimura, R.H.; Levin, A.M.; Cochran, F.V.; Cochran, J.R. Engineered cystine knot peptides that bind alphavbeta3, alphavbeta5, and alpha5beta1 integrins with low-nanomolar affinity. Proteins 2009, 77, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, C.H.; Kimura, R.H.; Withofs, N.; Tran, P.T.; Miao, Z.; Cochran, J.R.; Cheng, Z.; Felsher, D.; Kjær, A.; Willmann, J.K.; et al. PET imaging of tumor neovascularization in a transgenic mouse model using a novel 64Cu-DOTA-knottin peptide. Cancer Res. 2010, 70, 9022–9030. [Google Scholar] [CrossRef] [PubMed]
- Maaß, F.; Wüstehube-Lausch, J.; Dickgießer, S.; Valldorf, B.; Reinwarth, M.; Schmoldt, H.-U.; Daneschdar, M.; Avrutina, O.; Sahin, U.; Kolmar, H. Cystine-knot peptides targeting cancer-relevant human cytotoxic T lymphocyte-associated antigen 4 (CTLA-4). J. Pept. Sci. 2015, 21, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Vriens, K.; Cools, T.L.; Harvey, P.J.; Craik, D.J.; Spincemaille, P.; Cassiman, D.; Braem, A.; Vleugels, J.; Nibbering, P.H.; Drijfhout, J.W.; et al. Synergistic activity of the plant defensin HsAFP1 and caspofungin against candida albicans biofilms and planktonic cultures. PLoS ONE 2015, 10, e0132701. [Google Scholar] [CrossRef] [PubMed]
- Oguis, G.K.; Kan, M.W.; Craik, D.J. Natural Functions and Structure–Activity Relationships of Cyclotides; Elsevier: Amsterdam, The Netherlands, 2015; Available online: https://www.sciencedirect.com/science/article/pii/S0065229615000725 (accessed on 31 August 2022).
- Huang, Y.H.; Colgrave, M.L.; Clark, R.J.; Kotze, A.C.; Craik, D.J. Lysine-scanning mutagenesis reveals an amendable face of the cyclotide kalata B1 for the optimization of nematocidal activity. J. Biol. Chem. 2010, 285, 10797–10805. [Google Scholar] [CrossRef]
- Narayani, M.; Babu, R.; Chadha, A.; Srivastava, S. Production of bioactive cyclotides: A comprehensive overview. Phytochem. Rev. 2020, 19, 787–825. [Google Scholar] [CrossRef]
- Gattringer, J.; Ndogo, O.E.; Retzl, B.; Ebermann, C.; Gruber, C.W.; Hellinger, R. Cyclotides Isolated from violet plants of cameroon are inhibitors of human prolyl oligopeptidase. Front. Pharmacol. 2021, 12, 707596. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalmankar, N.V.; Gehi, B.R.; Sowdhamini, R. Effects of a plant cyclotide on conformational dynamics and destabilization of β-amyloid fibrils through molecular dynamics simulations. Front. Mol. Biosci. 2022, 9. [Google Scholar] [CrossRef]
- Kukadia, S.; Dehbi, H.M.; Tillin, T.; Coady, E.; Chaturvedi, N.; Hughes, A.D. A double-blind placebo-controlled crossover study of the effect of beetroot juice containing dietary nitrate on aortic and brachial blood pressure over 24 h. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef]
- Karimzadeh, L.; Behrouz, V.; Sohrab, G.; Hedayati, M.; Emami, G. A randomized clinical trial of beetroot juice consumption on inflammatory markers and oxidative stress in patients with type 2 diabetes. J. Food Sci. 2022. [Google Scholar] [CrossRef]
- Kaplan, S.A. Re: A two-week, double-blind, placebo-controlled trial of Viola odorata, Echium amoenum and Physalis alkekengi mixture in symptomatic benign prostate hyperplasia (BPH) men. J. Urol. 2018, 200, 6, Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Leevutinun, P.; Krisadaphong, P.; Petsom, A.J. Clinical evaluation of Gac extract (Momordica cochinchinensis) in an antiwrinkle cream formulation. J. Cosmet. Sci. 2015, 66, 175–187. [Google Scholar] [PubMed]
- Gou, Z.-P.; Zhao, Y.-L.; Zou, L.-L.; Wang, Y.; Shu, S.-Q.; Zhu, X.-H.; Zheng, L.; Shen, Q.; Luo, Z.; Miao, J.; et al. The safety and tolerability of alkaloids from Alstonia scholaris leaves in healthy Chinese volunteers: A single-centre, randomized, double-blind, placebo-controlled phase I clinical trial. Pharm. Biol. 2021, 59, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Chusak, C.; Thilavech, T.; Henry, C.J.; Adisakwattana, S. Acute effect of Clitoria ternatea flower beverage on glycemic response and antioxidant capacity in healthy subjects: A randomized crossover trial. BMC Complement. Altern. Med. 2018, 18, 6. [Google Scholar] [CrossRef] [PubMed]
- Mogaka, J.N.; Owuor, P.M.; Odhiambo, S.; Waterman, C.; McGuire, M.K.; Fuchs, G.J.; Attia, S.L. Investigating the impact of moringa oleifera supplemented to kenyan breastfeeding mothers on maternal and infant health: A cluster randomized single-blinded controlled pilot trial protocol. JPGN Rep. 2022, 3, e237. [Google Scholar] [CrossRef]
- Gambo, A.; Moodley, I.; Babashani, M.; Babalola, T.K.; Gqaleni, N. A double-blind, randomized controlled trial to examine the effect of Moringa oleifera leaf powder supplementation on the immune status and anthropometric parameters of adult HIV patients on antiretroviral therapy in a resource-limited setting. PLoS ONE 2021, 16, e0261935. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Attah, F.A.; Lawal, B.A.; Yusuf, A.B.; Adedeji, O.J.; Folahan, J.T.; Akhigbe, K.O.; Roy, T.; Lawal, A.A.; Ogah, N.B.; Olorundare, O.E.; et al. Nutritional and Pharmaceutical Applications of Under-Explored Knottin Peptide-Rich Phytomedicines. Plants 2022, 11, 3271. https://doi.org/10.3390/plants11233271
Attah FA, Lawal BA, Yusuf AB, Adedeji OJ, Folahan JT, Akhigbe KO, Roy T, Lawal AA, Ogah NB, Olorundare OE, et al. Nutritional and Pharmaceutical Applications of Under-Explored Knottin Peptide-Rich Phytomedicines. Plants. 2022; 11(23):3271. https://doi.org/10.3390/plants11233271
Chicago/Turabian StyleAttah, Francis Alfred, Bilqis Abiola Lawal, Abdulmalik Babatunde Yusuf, Oluwakorede Joshua Adedeji, Joy Temiloluwa Folahan, Kelvin Oluwafemi Akhigbe, Tithi Roy, Azeemat Adeola Lawal, Ngozi Blessing Ogah, Olufunke Esan Olorundare, and et al. 2022. "Nutritional and Pharmaceutical Applications of Under-Explored Knottin Peptide-Rich Phytomedicines" Plants 11, no. 23: 3271. https://doi.org/10.3390/plants11233271
APA StyleAttah, F. A., Lawal, B. A., Yusuf, A. B., Adedeji, O. J., Folahan, J. T., Akhigbe, K. O., Roy, T., Lawal, A. A., Ogah, N. B., Olorundare, O. E., & Chamcheu, J. C. (2022). Nutritional and Pharmaceutical Applications of Under-Explored Knottin Peptide-Rich Phytomedicines. Plants, 11(23), 3271. https://doi.org/10.3390/plants11233271