
Candidate Genes Associated with Abiotic Stress Response in Plants as Tools to Engineer Tolerance to Drought, Salinity and Extreme Temperatures in Wheat: An Overview


Abstract
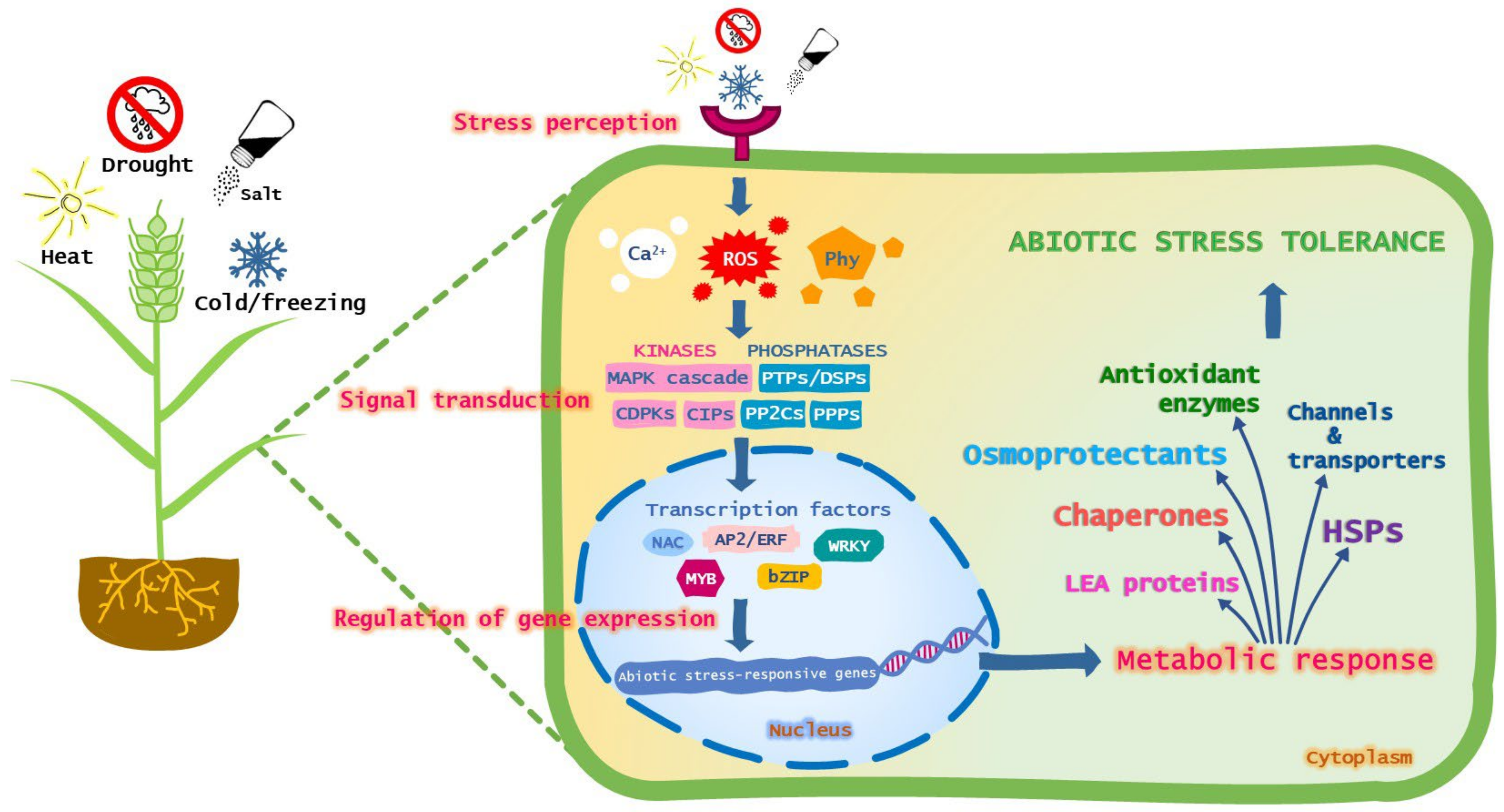
:1. Introduction
2. Candidate Genes
2.1. Transcription Factors
2.2. Osmolytes
2.3. Heat Shock Proteins and Other Chaperones
2.4. Late Embryogenesis Abundant Proteins
2.5. ROS Detoxification
2.6. Water Channels and Ion Transporters
3. Improvement of Abiotic Stress Tolerance in Wheat Plants through Transgenic Approaches
3.1. Drought

{kind=link}
| Gene | Gene Product | Plant Source | Improved Traits | Ref. |
|---|---|---|---|---|
| Transcription factors | ||||
| GmDREB1 | Dehydration-responsive element-binding protein | Soybean |
| [58] |
| [59] | |||
| AtDREBA1 | Dehydration-responsive element-binding protein | Arabidopsis thaliana |
| [60] |
| [61] | |||
| GhDREB | Dehydration-responsive element-binding protein | Cotton |
| [62] |
| TaDREB3 | Dehydration-responsive element-binding protein | Bread wheat |
| [63] |
| TaCBF5L | C-repeat binding factor | Bread wheat |
| [64] |
| TaERF3 | Ethylene response factor | Bread wheat |
| [65] |
| TaNAC69-1 | Protein belonging to the NAM/ATAF1-2/ CUC2 family | Bread wheat |
| [66] |
| SNAC1 | Protein belonging to the NAM/ATAF1-2/ CUC2 family | Rice |
| [67] |
| HaHB4 | Homeodomain-leucine zipper I protein | Sunflower |
| [68] |
| TaWRKY2 | WRKY domain protein | Bread wheat |
| [69] |
| AtWRKY30 | WRKY domain protein | Arabidopsis thaliana |
| [70] |
| TaASR1-D | Abscisic acid stress- ripening protein | Bread wheat |
| [71] |
| TaBZR2 | BRI1-EMS suppressor /brassinazole-resistant family | Bread wheat |
| [72] |
| TaNF-YA7-5B | Nuclear factor Y transcription factors | Bread wheat |
| [73] |
| Osmolytes | ||||
| P5CS | Δ1-pyrroline-5-carboxylate synthase | Vigna aconitifolia |
| [75,76,77] |
| AtOAT | Ornithine aminotransferase | Arabidopsis thaliana |
| [78] |
| mtlD | Mannitol-1-phosphate dehydrogenase | Escherichia coli |
| [80] |
| betA | Choline dehydrogenase | Escherichia coli |
| [81] |
| BADH | Betaine aldehyde dehydrogenase | Atriplex hortensis |
| [82] |
| LEA proteins | ||||
| HVA1 | Group 3 LEA protein | Barley |
| [83] |
| [84] | |||
| [85] | |||
| [86] | |||
| [87] | |||
| ROS detoxification | ||||
| TaNRX | Thioredoxin | Bread wheat |
| [88] |
| MsALR | Aldose reductase | Medicago sativa |
| [89] |
| Other genes | ||||
| PEPC | Phosphoenolpyruvate carboxylase | Maize |
| [91] |
| TaPEPKR2 | Phosphoenolpyruvate carboxylase kinase-related kinase | Bread wheat |
| [92] |
| SeCspA, SeCspB | Cold shock proteins | Escherichia coli |
| [93] |
| IPT | Isopentenyl transferase | Agrobacterium tumefaciens |
| [95] |
| OTS1 | cysteine protease (OVERLY TOLERANT TO SALT-1) | Arabidopsis thaliana |
| [96] |
| TaPYL4 | ABA receptor | Bread wheat |
| [97] |
3.2. Salinity
| Gene | Gene Product | Plant Source | Improved Traits | Ref. |
|---|---|---|---|---|
| Transcription factors | ||||
| GmDREB1 | Dehydration-responsive element-binding protein | Soybean |
| [58] |
| AtDREBA1 | Dehydration-responsive element-binding protein | Arabidopsis thaliana |
| [60] |
| GhDREB | Dehydration-responsive element-binding protein | Cotton |
| [62] |
| TaERF3 | Ethylene response factor | Bread wheat |
| [65] |
| SNAC1 | Protein belonging to the NAM/ATAF1-2/CUC2 family | Rice |
| [67] |
| TaASR1-D | Abscisic acid stress- ripening protein | Bread wheat |
| [71] |
| TabZIP15 | Basic leucine zipper proteins | Bread wheat |
| [101] |
| EsMYB90 | v-myb avian myeloblastosis viral oncogene homolog family | Eutrema salsugineum |
| [102] |
| TaMYB86B | v-myb avian myeloblastosis viral oncogene homolog family | Bread wheat |
| [103] |
| Osmolytes | ||||
| AtOAT | Ornithine aminotransferase | Arabidopsis thaliana |
| [78] |
| mtlD | Mannitol-1-phosphate dehydrogenase | Escherichia coli |
| [80] |
| [104] | |||
| betA | Choline dehydrogenase | Escherichia coli |
| [105] |
| BADH | Betaine aldehyde dehydrogenase | Atriplex hortensis |
| [106] |
| [107] | |||
| HvBADH1 | Betaine aldehyde dehydrogenase | Barley |
| [108] |
| LEA proteins | ||||
| HVA1 | Group 3 LEA protein | Barley |
| [84] |
| ROS detoxification | ||||
| TaPRX-2A | Peroxidase | Bread wheat |
| [109] |
| Aquaporins and ion transporters | ||||
| SbPIP1 | Plasma membrane intrinsic proteins | Salicornia bigelovii |
| [110] |
| TdPIP2;1 | Plasma membrane intrinsic proteins | Durum wheat |
| [111] |
| AtNHX1 | Vacuolar Na+/H+ antiporter | Arabidopsis thaliana |
| [113] |
| HVP1 | Vacuolar pyrophosphatase | Barley |
| [114] |
| Other genes | ||||
| SeCspA, SeCspB | Cold shock proteins | Escherichia coli |
| [93] |
| TaBASS2 | Pyruvate transporter | Bread wheat |
| [116] |
| TaPUB1 | U-box E3 ubiquitin ligase | Bread wheat |
| [117] |
3.3. High Temperatures
| Gene | Gene Product | Plant Source | Improved Traits | Ref. |
|---|---|---|---|---|
| Transcription factors | ||||
| AtWRKY30 | WRKY domain protein | Arabidopsis thaliana |
| [70] |
| TaHsfC2a-B | Heat shock factor | Bread wheat |
| [124] |
| TaHsfA6f | Heat shock factor | Bread wheat |
| [125] |
| Chaperones | ||||
| Zmeftu1 | Elongation Factor thermo-unstable | Maize |
| [126] |
| [127] | |||
| Osmolytes | ||||
| AtOAT | Ornithine aminotransferase | Arabidopsis thaliana |
| [78] |
| BADH | Betaine aldehyde dehydrogenase | Atriplex hortensis |
| [82] |
| LEA proteins | ||||
| HVA1 | Group 3 Late Embryogenesis Abundant protein | Barley |
| [85] |
| ROS detoxification | ||||
| TaFER-5B | Ferritin | Bread wheat |
| [129] |
| Other genes | ||||
| ZmPEPC | Phosphoenolpyruvate carboxylase | Maize |
| [128] |
| TaPEPKR2 | Phosphoenolpyruvate carboxylase kinase-related kinase | Bread wheat |
| [92] |
| SSI | Soluble starch synthase I | Rice |
| [130] |
3.4. Low Temperatures
4. Improvement of Abiotic Stress Tolerance in Wheat Plants through Genome Editing Approaches
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- EU Crops Market Observatory. Available online: https://ec.europa.eu/info/food-farming-fisheries/farming/facts-and-figures/markets/overviews/market-observatories/crops#cereals (accessed on 28 July 2022).
- Garutti, M.; Nevola, G.; Mazzeo, R.; Cucciniello, L.; Totaro, F.; Bertuzzi, C.A.; Caccialanza, R.; Pedrazzoli, P.; Puglisi, F. The impact of cereal grain composition on the health and disease outcomes. Front. Nutr. 2022, 9, 888974. [Google Scholar] [CrossRef]
- Food and Agriculture Projections to 2050. Available online: https://www.fao.org/global-perspectives-studies/food-agriculture-projections-to-2050/en/ (accessed on 28 July 2022).
- Sharma, I.; Tyagi, B.S.; Singh, G.; Venkatesh, K.; Gupta, O.P. Enhancing wheat production-a global perspective. Indian J. Agric. Sci. 2015, 85, 3–13. [Google Scholar]
- Hussain, B. Modernization in plant breeding approaches for improving biotic stress resistance in crop plants. Turk. J. Agric. For. 2015, 39, 515–530. [Google Scholar] [CrossRef]
- Chauhan, H.; Khurana, P. Wheat genetic transformation using mature embryos as explants. Methods Mol. Biol. 2017, 1679, 153–167. [Google Scholar]
- Shrawat, A.K.; Armstrong, C.L. Development and application of genetic engineering for wheat improvement. Crit. Rev. Plant Sci. 2018, 37, 335–421. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Humanes, J.; Wang, Y.; Liang, Z.; Shan, Q.; Ozuna, C.V.; Sánchez-León, S.; Baltes, N.J.; Starker, C.; Barro, F.; Gao, C.; et al. High-efficiency gene targeting in hexaploid wheat using DNA replicons and CRISPR/Cas9. Plant J. 2017, 89, 1251–1262. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Hua, L.; Gupta, A.; Tricoli, D.; Edwards, K.J.; Yang, B.; Li, W. Development of an Agrobacterium-delivered CRISPR/Cas9 system for wheat genome editing. Plant Biotechnol. J. 2019, 17, 1623–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, J.; Minkenberg, B.; Poddar, S.; Staskawicz, B.; Cho, M.J. Improvement of gene delivery and mutation efficiency in the CRISPR-Cas9 wheat (Triticum aestivum L.) genomics system via biolistics. Genes 2022, 13, 1180. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, H.; Shao, H.; Tang, X. Recent advances in utilizing transcription factors to improve plant abiotic stress tolerance by transgenic technology. Front. Plant Sci. 2016, 7, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Nolan, T.M.; Jiang, H.; Yin, Y. AP2/ERF transcription factor regulatory networks in hormone and abiotic stress responses in Arabidopsis. Front. Plant Sci. 2019, 10, 228. [Google Scholar] [CrossRef] [Green Version]
- Nuruzzaman, M.; Sharoni, A.M.; Kikuchi, S. Roles of NAC transcription factors in the regulation of biotic and abiotic stress responses in plants. Front. Microbiol. 2013, 4, 248. [Google Scholar] [CrossRef]
- Yu, Y.; Qian, Y.; Jiang, M.; Xu, J.; Yang, J.; Zhang, T.; Gou, L.; Pi, E. Regulation mechanisms of plant basic leucine zippers to various abiotic stresses. Front. Plant Sci. 2020, 11, 1258. [Google Scholar] [CrossRef]
- Chen, L.; Song, Y.; Li, S.; Zhang, L.; Zou, C.; Yu, D. The role of WRKY transcription factors in plant abiotic stresses. Biochim. Biophys. Acta 2012, 1819, 120–128. [Google Scholar] [CrossRef]
- Wu, H.L.; Ni, Z.F.; Yao, Y.Y.; Guo, G.G.; Sun, Q.X. Cloning and expression profiles of 15 genes encoding WRKY transcription factor in wheat (Triticum aestivum L.). Prog. Nat. Sci. 2008, 18, 697–705. [Google Scholar] [CrossRef]
- Li, J.; Han, G.; Sun, C.; Sui, N. Research advances of MYB transcription factors in plant stress resistance and breeding. Plant Signal. Behav. 2019, 14, 1613131. [Google Scholar] [CrossRef] [PubMed]
- Jogawat, A. Osmolytes and their role in abiotic stress tolerance in plants. In Molecular Plant Abiotic Stress; Roychoudhury, A., Tripathi, D.K., Eds.; John Wiley & Sons, Ltd.: West Sussex, UK, 2019; pp. 91–97. [Google Scholar]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meena, M.; Divyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Yadav, M.; Upadhyay, R.S. Regulation of L-proline biosynthesis, signal transduction, transport, accumulation and its vital role in plants during variable environmental conditions. Heliyon 2019, 5, e02952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’Aversana, E.; Carillo, P. Spatial and temporal profile of glycine betaine accumulation in plants under abiotic stresses. Front. Plant Sci. 2019, 10, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giri, J. Glycinebetaine and abiotic stress tolerance in plants. Plant Signal. Behav. 2011, 6, 1746–1751. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Kundu, A. Sugars and sugar polyols in overcoming environmental stresses. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress; Aryadeep Roychoudhury, A., Tripathi, D.K., Eds.; John Wiley & Sons, Ltd.: West Sussex, UK, 2020; pp. 71–101. [Google Scholar]
- De Carvalho, K.; Petkowicz, C.L.; Nagashima, G.T.; Bespalhok Filho, J.C.; Vieira, L.G.; Pereira, L.F.; Domingues, D.S. Homeologous genes involved in mannitol synthesis reveal unequal contributions in response to abiotic stress in Coffea arabica. Mol. Genet. Genom. 2014, 289, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Timperio, A.M.; Egidi, M.G.; Zolla, L. Proteomics applied on plant abiotic stresses: Role of heat shock proteins (HSP). J. Proteome 2008, 71, 391–411. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.S.; Kavi Kishor, P.B.; Seiler, C.; Kuhlmann, M.; Eschen-Lippold, L.; Lee, J.; Reddy, M.K.; Sreenivasulu, N. Unraveling regulation of the small heat shock proteins by the heat shock factor HvHsfB2c in barley: Its implications in drought stress response and seed development. PLoS ONE 2014, 9, e89125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, D.; Tuteja, N. Chaperones and foldases in endoplasmic reticulum stress signaling in plants. Plant Signal. Behav. 2011, 6, 232–236. [Google Scholar] [CrossRef] [Green Version]
- Garg, G.; Yadav, S.; Ruchi, Y.G. Key roles of calreticulin and calnexin proteins in plant perception under stress conditions: A review. Int. J. Adv. Life Sci. 2015, 5, 18–26. [Google Scholar]
- Ristic, Z.; Momcilović, I.; Fu, J.; Callegari, E.; DeRidder, B.P. Chloroplast protein synthesis elongation factor, EF-Tu, reduces thermal aggregation of rubisco activase. J. Plant Physiol. 2007, 164, 1564–1571. [Google Scholar] [CrossRef]
- Amara, I.; Zaidi, I.; Masmoudi, K.; Ludevid, M.D.; Pagès, M.; Goday, A.; Brini, F. Insights into late embryogenesis abundant (LEA) proteins in plants: From structure to the functions. Am. J. Plant Sci. 2014, 5, 3440–3455. [Google Scholar] [CrossRef] [Green Version]
- Kovacs, D.; Kalmar, E.; Torok, Z.; Tompa, P. Chaperone activity of ERD10 and ERD14, two disordered stress-related plant proteins. Plant Physiol. 2008, 147, 381–390. [Google Scholar] [CrossRef]
- Marttila, S.; Tenhola, T.; Mikkonen, A. A barley (Hordeum vulgare L.) LEA3 protein, HVA1, is abundant in protein storage. Planta 1996, 199, 602–611. [Google Scholar] [CrossRef]
- Kader, A.; Almeslemani, M.; Baghdady, A.; Alzubi, H.; Alasaad, N.; Ali Basha, N.; Dameriha, A.; Jacobsen, H.J.; Hassan, F. Isolation, characterization of the hva1 gene from Syrian barley varieties and cloning into a binary plasmid vector. Int. J. Bot. 2012, 8, 117–126. [Google Scholar]
- Imai, R.; Chang, L.; Ohta, A.; Bray, E.A.; Takagi, M. A lea-class gene of tomato confers salt and freezing tolerance when expressed in Saccharomyces cerevisiae. Gene 1996, 170, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Calderón, A.; Sevilla, F.; Jiménez, A. Redox protein thioredoxins: Function under salinity, drought and extreme temperature conditions. In Antioxidants and Antioxidant Enzymes in Higher Plants; Gupta, D., Palma, J., Corpas, F., Eds.; Springer: Cham, Switzerland, 2018; pp. 123–162. [Google Scholar]
- Bartels, D. Targeting detoxification pathways: An efficient approach to obtain plants with multiple stress tolerance? Trends Plant Sci. 2001, 6, 284–286. [Google Scholar] [CrossRef] [PubMed]
- Roldán-Arjona, T.; Ariza, R.R. Repair and tolerance of oxidative DNA damage in plants. Mutat. Res. 2009, 681, 169–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent developments in enzymatic antioxidant defence mechanism in plants with special reference to abiotic stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Ballesta, M.C.; Carvajal, M. New challenges in plant aquaporin biotechnology. Plant Sci. 2014, 217–218, 71–77. [Google Scholar] [CrossRef]
- Uehlein, N.; Sperling, H.; Heckwolf, M.; Kaldenhoff, R. The Arabidopsis aquaporin PIP1;2 rules cellular CO2 uptake. Plant Cell Environ. 2012, 35, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Zwiazek, J.J.; Xu, H.; Tan, X.; Navarro-Ródenas, A.; Morte, A. Significance of oxygen transport through aquaporins. Sci. Rep. 2017, 7, 40411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kader, M.A.; Lindberg, S.; Seidel, T.; Golldack, D.; Yemelyanov, V. Sodium sensing induces different changes in free cytosolic calcium concentration and pH in salt-tolerant and -sensitive rice (Oryza sativa) cultivars. Physiol. Plant. 2007, 130, 99–111. [Google Scholar] [CrossRef]
- Martínez-Ballesta, M.C.; Cabañero, F.J.; Maurel, C.; Olmos, E.; Carvajal, M. Nutritional Calcium as the regulator of the aquaporin activity in plants grown under salinity. Planta 2008, 228, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Kuwagata, T.; Ishikawa-Sakurai, J.; Hayashi, H.; Nagasuga, K.; Fukushi, K.; Ahamed, A.; Takasugi, K.; Katsuhara, M.; Murai-Hatano, M. Influence of low air humidity and low root temperature on water uptake, growth and aquaporin expression in rice plants. Plant Cell Physiol. 2012, 53, 1418–1431. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.Y.; Kim, D.G.; Kim, Y.O.; Kim, J.S.; Kang, H. An expression analysis of a gene family encoding plasma membrane aquaporins in response to abiotic stresses in Arabidopsis thaliana. Plant Mol. Biol. 2004, 54, 713–725. [Google Scholar] [CrossRef]
- Ahamed, A.; Murai-Hatano, M.; Ishikawa-Sakurai, J.; Hayashi, H.; Kawamura, Y.; Uemura, M. Cold stress-induced acclimation in rice is mediated by root-specific aquaporins. Plant Cell Physiol. 2012, 53, 1445–1456. [Google Scholar] [CrossRef]
- Seth, R.; Maritim, T.K.; Parmar, R.; Sharma, R.K. Underpinning the molecular programming attributing heat stress associated thermotolerance in tea (Camellia sinensis (L.) O. Kuntze). Hortic. Res. 2021, 8, 99. [Google Scholar] [CrossRef]
- Christou, A.; Filippou, P.; Manganaris, G.A.; Fotopoulos, V. Sodium hydrosulfide induces systemic thermotolerance to strawberry plants through transcriptional regulation of heat shock proteins and aquaporin. BMC Plant Biol. 2014, 14, 42. [Google Scholar] [CrossRef] [Green Version]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Wu, H. Plant salt tolerance and Na+ sensing and transport. Crop J. 2018, 6, 215–225. [Google Scholar] [CrossRef]
- Dietz, K.-J.; Zörb, C.; Geilfus, C.-M. Drought and crop yield. Plant Biol. 2021, 23, 881–893. [Google Scholar] [CrossRef]
- Shiqing, G.; Huijun, X.; Xianguo, C.; Ming, C.; Zhaoshi, X.; Liancheng, L.; Xingguo, Y.; Lipu, D.; Xiaoyan, H.; Youzhi, M. Improvement of wheat drought and salt tolerance by expression of a stress-inducible transcription factor GmDREB of soybean (Glycine max). Chin. Sci. Bull. 2005, 50, 2714–2723. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, M.; Guo, J.; Wang, Y.; Min, D.; Jiang, Q.; Ji, H.; Huang, C.; Wei, W.; Xu, H.; et al. Overexpression of soybean DREB1 enhances drought stress tolerance of transgenic wheat in the field. J. Exp. Bot. 2020, 71, 1842–1857. [Google Scholar] [CrossRef] [PubMed]
- Noor, S.; Ali, S.; Hafeez-ur-Rahman; Farhatullah; Ali, G.M. Comparative study of transgenic (DREB1A) and non-transgenic wheat lines on relative water content, sugar, proline and chlorophyll under drought and salt stresses. Sarhad J. Agric. 2018, 34, 986–993. [Google Scholar] [CrossRef]
- Saint Pierre, C.; Crossa, J.L.; Bonnett, D.; Yamaguchi-Shinozaki, K.; Reynolds, M.P. Phenotyping transgenic wheat for drought resistance. J. Exp. Bot. 2012, 63, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.Q.; Chen, M.; Xia, L.Q.; Xiu, H.J.; Xu, Z.S.; Li, L.C.; Zhao, C.P.; Cheng, X.G.; Ma, Y.Z. A cotton (Gossypium hirsutum) DRE-binding transcription factor gene, GhDREB, confers enhanced tolerance to drought, high salt, and freezing stresses in transgenic wheat. Plant Cell Rep. 2009, 28, 301–311. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Baho, M.; Lopato, S.; Langridge, P. The TaDREB3 transgene transferred by conventional crossings to different genetic backgrounds of bread wheat improves drought tolerance. Plant Biotechnol. J. 2016, 14, 313–322. [Google Scholar] [CrossRef]
- Yang, Y.; Al-Baidhani, H.H.J.; Harris, J.; Riboni, M.; Li, Y.; Mazonka, I.; Bazanova, N.; Chirkova, L.; Sarfraz Hussain, S.; Hrmova, M.; et al. DREB/CBF expression in wheat and barley using the stress-inducible promoters of HD-Zip I genes: Impact on plant development, stress tolerance and yield. Plant Biotechnol. J. 2020, 18, 829–844. [Google Scholar] [CrossRef] [Green Version]
- Rong, W.; Qi, L.; Wang, A.; Ye, X.; Du, L.; Liang, H.; Xin, Z.; Zhang, Z. The ERF transcription factor TaERF3 promotes tolerance to salt and drought stresses in wheat. Plant Biotechnol. J. 2014, 12, 468–479. [Google Scholar] [CrossRef]
- Xue, G.P.; Way, H.M.; Richardson, T.; Drenth, J.; Joyce, P.A.; McIntyre, C.L. Overexpression of TaNAC69 leads to enhanced transcript levels of stress up-regulated genes and dehydration tolerance in bread wheat. Mol. Plant. 2011, 4, 697–712. [Google Scholar] [CrossRef]
- Saad, A.S.; Li, X.; Li, H.P.; Huang, T.; Gao, C.S.; Guo, M.W.; Cheng, W.; Zhao, G.Y.; Liao, Y.C. A rice stress-responsive NAC gene enhances tolerance of transgenic wheat to drought and salt stresses. Plant Sci. 2013, 203–204, 33–40. [Google Scholar] [CrossRef]
- González, F.G.; Capella, M.; Ribichich, K.F.; Curín, F.; Giacomelli, J.I.; Ayala, F.; Watson, G.; Otegui, M.E.; Chan, R.L. Field-grown transgenic wheat expressing the sunflower gene HaHB4 significantly outyields the wild-type. J. Exp. Bot. 2019, 70, 1669–1681. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Wang, Y.; Xu, P.; Zhang, Z. Overexpression of a WRKY transcription factor TaWRKY2 enhances drought stress tolerance in transgenic wheat. Front. Plant Sci. 2018, 9, 997. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Ahmad, M. Overexpression of AtWRKY30 transcription factor enhances heat and drought stress tolerance in wheat (Triticum aestivum L.). Genes 2019, 10, 163. [Google Scholar] [CrossRef] [Green Version]
- Qiu, D.; Hu, W.; Zhou, Y.; Xiao, J.; Hu, R.; Wei, Q.; Zhang, Y.; Feng, J.; Sun, F.; Sun, J.; et al. TaASR1-D confers abiotic stress resistance by affecting ROS accumulation and ABA signalling in transgenic wheat. Plant Biotechnol. J. 2021, 19, 1588–1601. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.Y.; Gao, Y.; Guo, J.; Yu, T.F.; Zheng, W.J.; Liu, Y.W.; Chen, J.; Xu, Z.S.; Ma, Y.Z. BES/BZR Transcription factor TaBZR2 positively regulates drought responses by activation of TaGST1. Plant Physiol. 2019, 180, 605–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Zhang, Y.; Li, T.; Ni, C.; Bai, X.; Lin, R.; Xiao, K. TaNF-YA7-5B, a gene encoding nuclear factor Y (NF-Y) subunit A in Triticum aestivum, confers plant tolerance to PEG-inducing dehydration simulating drought through modulating osmotic stress-associated physiological processes. Plant Physiol. Biochem. 2022, 188, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Debnath, B.; Islam, W.; Li, M.; Sun, Y.; Lu, X.; Mitra, S.; Hussain, M.; Liu, S.; Qiu, D. Melatonin mediates enhancement of stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 1040. [Google Scholar] [CrossRef] [Green Version]
- Vendruscolo, E.C.; Schuster, I.; Pileggi, M.; Scapim, C.A.; Molinari, H.B.; Marur, C.J.; Vieira, L.G. Stress-induced synthesis of proline confers tolerance to water deficit in transgenic wheat. J. Plant Physiol. 2007, 164, 1367–1376. [Google Scholar] [CrossRef]
- Pavei, D.; Gonçalves-Vidigal, M.C.; Schuelter, A.R.; Schuster, I.; Vieira, E.S.N.; Vendruscolo, E.C.G.; Poletine, J.P. Response to water stress in transgenic (p5cs gene) wheat plants (Triticum aestivum L.). Aust. J. Crop Sci. 2016, 10, 776–783. [Google Scholar] [CrossRef]
- De Lima, L.A.D.C.; Schuster, I.; da Costa, A.C.T.; Vendruscolo, E.C.G. Evaluation of wheat events transformed with the p5cs gene under conditions of water stress. Rev. Ciências Agrárias 2019, 42, 448–455. [Google Scholar]
- Anwar, A.; Wang, K.; Wang, J.; Shi, L.; Du, L.; Ye, X. Expression of Arabidopsis Ornithine Aminotransferase (AtOAT) encoded gene enhances multiple abiotic stress tolerances in wheat. Plant Cell Rep. 2021, 40, 1155–1170. [Google Scholar] [CrossRef] [PubMed]
- Tarczynski, M.C.; Jensen, R.G.; Bohnert, H.J. Expression of a bacterial mtlD gene in transgenic tobacco leads to production and accumulation of mannitol. Proc. Natl. Acad. Sci. USA 1992, 89, 2600–2604. [Google Scholar] [CrossRef] [Green Version]
- Abebe, T.; Guenzi, A.C.; Martin, B.; Cushman, J.C. Tolerance of mannitol-accumulating transgenic wheat to water stress and salinity. Plant Physiol. 2003, 131, 1748–1755. [Google Scholar] [CrossRef]
- He, C.; Zhang, W.; Gao, Q.; Yang, A.; Hu, X.; Zhang, J. Enhancement of drought resistance and biomass by increasing the amount of glycine betaine in wheat seedlings. Euphytica 2011, 177, 151–167. [Google Scholar] [CrossRef]
- Wang, G.-P.; Hui, Z.; Li, F.; Zhao, M.-R.; Zhang, J.; Wang, W. Improvement of heat and drought photosynthetic tolerance in wheat by overaccumulation of glycine betaine. Plant Biotechnol. Rep. 2010, 4, 213–222. [Google Scholar] [CrossRef]
- Sivamani, E.; Bahieldin, A.; Wraith, J.M.; Al-Niemi, T.; Dyer, W.E.; Ho, T.D.; Qu, R. Improved biomass productivity and water use efficiency under water deficit conditions in transgenic wheat constitutively expressing the barley HVA1 gene. Plant Sci. 2000, 155, 1–9. [Google Scholar] [CrossRef]
- Habib, I.; Shahzad, K.; Rauf, M.; Ahmad, M.; Alsamadany, H.; Fahad, S.; Saeed, N.A. Dehydrin responsive HVA1 driven inducible gene expression enhanced salt and drought tolerance in wheat. Plant Physiol. Biochem. 2022, 180, 124–133. [Google Scholar] [CrossRef]
- Samtani, H.; Sharma, A.; Khurana, P. Overexpression of HVA1 enhances drought and heat stress tolerance in Triticum aestivum doubled haploid plants. Cells 2022, 11, 912. [Google Scholar] [CrossRef]
- Chauhan, H.; Khurana, P. Use of doubled haploid technology for development of stable drought tolerant bread wheat (Triticum aestivum L.) transgenics. Plant Biotechnol. J. 2011, 9, 408–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahieldin, A.; Mahfouz, H.T.; Eissa, H.F.; Saleh, O.M.; Ramadan, A.M.; Ahmed, I.A.; Dyer, W.A.; El-Itribya, H.A.; Madkour, M.A. Field evaluation of transgenic wheat plants stably expressing the HVA1 gene for drought tolerance. Physiol. Plant. 2005, 123, 421–427. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, J.; Wei, F.; Song, T.; Yu, Y.; Yu, M.; Fan, Q.; Yang, Y.; Xue, G.; Zhang, X. Nucleoredoxin gene TaNRX1 positively regulates drought tolerance in transgenic wheat (Triticum aestivum L.). Front. Plant Sci. 2021, 12, 756338. [Google Scholar] [CrossRef] [PubMed]
- Fehér-Juhász, E.; Majer, P.; Sass, L.; Lantos, C.; Csiszár, J.; Turóczy, Z.; Mihály, R.; Mai, A.; Horváth, G.V.; Vass, I.; et al. Phenotyping shows improved physiological traits and seed yield of transgenic wheat plants expressing the alfalfa aldose reductase under permanent drought stress. Acta Physiol. Plant. 2014, 36, 663–673. [Google Scholar] [CrossRef] [Green Version]
- Miyao, M. Molecular evolution and genetic engineering of C4 photosynthetic enzymes. J. Exp. Bot. 2003, 54, 179–189. [Google Scholar] [CrossRef]
- Qin, N.; Xu, W.; Hu, L.; Li, Y.; Wang, H.; Qi, X.; Fang, Y.; Hua, X. Drought tolerance and proteomics studies of transgenic wheat containing the maize C4 phosphoenolpyruvate carboxylase (PEPC) gene. Protoplasma 2016, 253, 1503–1512. [Google Scholar] [CrossRef]
- Zang, X.; Geng, X.; He, K.; Wang, F.; Tian, X.; Xin, M.; Yao, Y.; Hu, Z.; Ni, Z.; Sun, Q.; et al. Overexpression of the wheat (Triticum aestivum L.) TaPEPKR2 gene enhances heat and dehydration tolerance in both wheat and Arabidopsis. Front. Plant Sci. 2018, 9, 1710. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.F.; Xu, Z.S.; Guo, J.K.; Wang, Y.X.; Abernathy, B.; Fu, J.D.; Chen, X.; Zhou, Y.B.; Chen, M.; Ye, X.G.; et al. Improved drought tolerance in wheat plants overexpressing a synthetic bacterial cold shock protein gene SeCspA. Sci. Rep. 2017, 7, 44050. [Google Scholar] [CrossRef] [Green Version]
- Nakaminami, K.; Karlson, D.T.; Imai, R. Functional conservation of cold shock domains in bacteria and higher plants. Proc. Natl. Acad. Sci. USA 2006, 103, 10122–10127. [Google Scholar] [CrossRef] [Green Version]
- Beznec, A.; Faccio, P.; Miralles, D.J.; Abeledo, L.G.; Oneto, C.D.; Garibotto, M.B.; Gonzalez, G.; Moreyra, F.; Elizondo, M.; Ruíz, M.; et al. Stress-induced expression of IPT gene in transgenic wheat reduces grain yield penalty under drought. J. Genet. Eng. Biotechnol. 2021, 19, 67. [Google Scholar] [CrossRef]
- Le Roux, M.L.; Kunert, K.J.; van der Vyver, C.; Cullis, C.A.; Botha, A.M. Expression of a small ubiquitin-like modifier protease increases drought tolerance in wheat (Triticum aestivum L.). Front. Plant Sci. 2019, 10, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mega, R.; Abe, F.; Kim, J.S.; Tsuboi, Y.; Tanaka, K.; Kobayashi, H.; Sakata, Y.; Hanada, K.; Tsujimoto, H.; Kikuchi, J.; et al. Tuning water-use efficiency and drought tolerance in wheat using abscisic acid receptors. Nat. Plants 2019, 5, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Arora, N.K. Impact of climate change on agriculture production and its sustainable solutions. Environ. Sustain. 2019, 2, 95–96. [Google Scholar] [CrossRef] [Green Version]
- EL Sabagh, A.; Islam, M.S.; Skalicky, M.; Ali Raza, M.; Singh, K.; Anwar Hossain, M.; Hossain, A.; Mahboob, W.; Iqbal, M.A.; Ratnasekera, D.; et al. Salinity stress in wheat (Triticum aestivum L.) in the changing climate: Adaptation and management srategies. Front. Agron. 2021, 3, 661932. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of plant responses to salt stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- Bi, C.; Yu, Y.; Dong, C.; Yang, Y.; Zhai, Y.; Du, F.; Xia, C.; Ni, Z.; Kong, X.; Zhang, L. The bZIP transcription factor TabZIP15 improves salt stress tolerance in wheat. Plant Biotechnol. J. 2021, 19, 209–211. [Google Scholar] [CrossRef]
- Li, C.; Zhao, Y.; Qi, Y.; Duan, C.; Zhang, H.; Zhang, Q. Eutrema EsMYB90 gene improves growth and antioxidant capacity of transgenic wheat under salinity stress. Front. Plant Sci. 2022, 13, 856163. [Google Scholar] [CrossRef]
- Song, Y.; Yang, W.; Fan, H.; Zhang, X.; Sui, N. TaMYB86B encodes a R2R3-type MYB transcription factor and enhances salt tolerance in wheat. Plant Sci. 2020, 300, 110624. [Google Scholar] [CrossRef]
- El-Yazal, M.A.S.; Eissa, H.F.; Ahmed, S.M.A.E.; Howladar, S.M.; Zaki, S.S.; Rady, M.M. The mtlD gene-overexpressed transgenic wheat tolerates salt stress through accumulation of mannitol and sugars. Plant 2016, 4, 78–90. [Google Scholar] [CrossRef]
- He, C.; Yang, A.; Zhang, W.; Gao, Q.; Zhang, J. Improved salt tolerance of transgenic wheat by introducing betA gene for glycine betaine synthesis. Plant Cell Tiss. Organ Cult. 2010, 101, 65–78. [Google Scholar] [CrossRef]
- Liang, C.; Zhang, X.Y.; Luo, Y.; Wang, G.P.; Zou, Q.; Wang, W. Overaccumulation of glycine betaine alleviates the negative effects of salt stress in wheat. Russ. J. Plant Physiol. 2009, 56, 370–376. [Google Scholar] [CrossRef]
- Tian, F.; Wang, W.; Liang, C.; Wang, X.; Wang, G.; Wang, W. Overaccumulation of glycine betaine makes the function of the thylakoid membrane better in wheat under salt stress. Crop J. 2017, 5, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Cai, J.; Luo, X.; Chang, T.; Li, J.; Zhao, Y.; Xu, Y. Transformation of wheat Triticum aestivum with the HvBADH1 transgene from hulless barley improves salinity-stress tolerance. Acta Physiol. Plant. 2019, 41, 155. [Google Scholar] [CrossRef]
- Su, P.; Yan, J.; Li, W.; Wang, L.; Zhao, J.; Ma, X.; Li, A.; Wang, H.; Kong, L. A member of wheat class III peroxidase gene family, TaPRX-2A, enhanced the tolerance of salt stress. BMC Plant Biol. 2020, 20, 392. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.H.; Zhang, X.; Ma, H.X. Changes in the physiological parameters of SbPIP1-transformed wheat plants under salt stress. Int. J. Genom. 2015, 2015, 384356. [Google Scholar]
- Ayadi, M.; Brini, F.; Masmoudi, K. Overexpression of a wheat aquaporin gene, TdPIP2;1, enhances salt and drought tolerance in transgenic durum wheat cv. Maali. Int. J. Mol. Sci. 2019, 20, 2389. [Google Scholar] [CrossRef]
- Afzal, Z.; Howton, T.C.; Sun, Y.; Mukhtar, M.S. The roles of aquaporins in plant stress responses. J. Dev. Biol. 2016, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Xue, Z.-Y.; Zhi, D.-Y.; Xue, G.-P.; Zhang, H.; Zhao, Y.-X.; Xia, G.-M. Enhanced salt tolerance of transgenic wheat (Tritivum aestivum L.) expressing a vacuolar Na+/H+ antiporter gene with improved grain yields in saline soils in the field and a reduced level of leaf Na+. Plant Sci. 2004, 167, 849–859. [Google Scholar] [CrossRef]
- Haq, R.F.U.; Saeed, N.A.; Ahmed, M.; Arshad, Z.; Mansoor, S.; Habib, I.; Tester, M. Barley vacuolar pyrophosphatase (HVP1) gene confers salinity tolerance in locally adapted wheat (Triticum aestivum). Int. J. Agric. Biol. 2019, 22, 1338–1346. [Google Scholar]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Ai, X.; Wang, M.; Xiao, L.; Xia, G. A putative pyruvate transporter TaBASS2 positively regulates salinity tolerance in wheat via modulation of ABI4 expression. BMC Plant Biol. 2016, 16, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Wang, W.; Wu, Y.; Li, Q.; Zhang, G.; Shi, R.; Yang, J.; Wang, Y.; Wang, W. The involvement of wheat U-box E3 ubiquitin ligase TaPUB1 in salt stress tolerance. J. Integr. Plant Biol. 2020, 62, 631–651. [Google Scholar] [CrossRef] [PubMed]
- Wahid, A.; Gelania, S.; Ashrafa, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Streck, N.A. Climate change and agroecosystems: The effect of elevated CO2 and temperature on crop growth, development, and yield. Ciência Rural 2005, 35, 730–740. [Google Scholar] [CrossRef] [Green Version]
- Akter, N.; Rafiqul Islam, M. Heat stress effects and management in wheat. A review. Agron. Sustain. Dev. 2017, 37, 37. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Mittler, R.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Plant adaptations to the combination of drought and high temperatures. Physiol. Plant. 2018, 162, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Al-Whaibi, M.H. Plant heat-shock proteins: A mini review. J. King Saud Univ. Sci. 2011, 23, 139–150. [Google Scholar] [CrossRef]
- Xue, G.P.; Sadat, S.; Drenth, J.; McIntyre, C.L. The heat shock factor family from Triticum aestivum in response to heat and other major abiotic stresses and their role in regulation of heat shock protein genes. J. Exp. Bot. 2014, 65, 539–557. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.J.; Chen, D.; Lynne Mclntyre, C.; Fernanda Dreccer, M.; Zhang, Z.B.; Drenth, J.; Kalaipandian, S.; Chang, H.; Xue, G.P. Heat shock factor C2a serves as a proactive mechanism for heat protection in developing grains in wheat via an ABA-mediated regulatory pathway. Plant Cell Environ. 2018, 41, 79–98. [Google Scholar] [CrossRef]
- Xue, G.P.; Drenth, J.; McIntyre, C.L. TaHsfA6f is a transcriptional activator that regulates a suite of heat stress protection genes in wheat (Triticum aestivum L.) including previously unknown Hsf targets. J. Exp. Bot. 2015, 66, 1025–1039. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Momcilović, I.; Clemente, T.E.; Nersesian, N.; Trick, H.N.; Ristic, Z. Heterologous expression of a plastid EF-Tu reduces protein thermal aggregation and enhances CO2 fixation in wheat (Triticum aestivum) following heat stress. Plant Mol. Biol. 2008, 68, 277–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.; Ristic, Z. Analysis of transgenic wheat (Triticum aestivum L.) harboring a maize (Zea mays L.) gene for plastid EF-Tu: Segregation pattern, expression and effects of the transgene. Plant Mol. Biol. 2010, 73, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Xu, W.; Zhang, J.; Guo, R.; Zhao, M.; Hu, L.; Wang, H.; Dong, H.; Li, Y. Physiological characteristics and metabolomics of transgenic wheat containing the maize C4 phosphoenolpyruvate carboxylase (PEPC) gene under high temperature stress. Protoplasma 2017, 254, 1017–1030. [Google Scholar] [CrossRef] [PubMed]
- Zang, X.; Geng, X.; Wang, F.; Liu, Z.; Zhang, L.; Zhao, Y.; Tian, X.; Ni, Z.; Yao, Y.; Xin, M.; et al. Overexpression of wheat ferritin gene TaFER-5B enhances tolerance to heat stress and other abiotic stresses associated with the ROS scavenging. BMC Plant Biol. 2017, 17, 14. [Google Scholar] [CrossRef] [Green Version]
- Keeling, P.L.; Bacon, P.J.; Holt, D.C. Elevated temperature reduces starch deposition in wheat endosperm by reducing the activity of soluble starch synthase. Planta 1993, 191, 342–348. [Google Scholar] [CrossRef]
- Kumari, A.; Kumar, R.R.; Singh, J.P.; Verma, P.; Singh, G.P.; Chinnusamy, V.; Praveen, S.; Goswami, S. Characterization of the starch synthase under terminal heat stress and its effect on grain quality of wheat. 3 Biotech 2020, 10, 531. [Google Scholar] [CrossRef]
- Tian, B.; Talukder, S.K.; Fu, J.; Fritz, A.K.; Trick, H.N. Expression of a rice soluble starch synthase gene in transgenic wheat improves the grain yield under heat stress conditions. In Vitro Cell. Dev. Biol. Plant. 2018, 54, 216–227. [Google Scholar] [CrossRef]
- Smidansky, E.D.; Clancy, M.; Meyer, F.D.; Lanning, S.P.; Blake, N.K.; Talbert, L.E.; Giroux, M.J. Enhanced ADP-glucose pyrophosphorylase activity in wheat endosperm increases seed yield. Proc. Natl. Acad. Sci. USA 2002, 99, 1724–1729. [Google Scholar] [CrossRef] [Green Version]
- Frederiks, T.M.; Christopher, J.T.; Sutherland, M.W.; Borrell, A.K. Post-head-emergence frost in wheat and barley: Defining the problem, assessing the damage, and identifying resistance. J. Exp. Bot. 2015, 66, 3487–3498. [Google Scholar] [CrossRef] [Green Version]
- Hassan, M.A.; Xiang, C.; Farooq, M.; Muhammad, N.; Yan, Z.; Hui, X.; Yuanyuan, K.; Bruno, A.K.; Lele, Z.; Jincai, L. Cold stress in wheat: Plant acclimation responses and management strategies. Front. Plant Sci. 2021, 12, 676884. [Google Scholar] [CrossRef]
- Zhang, X.-Y.; Liang, C.; Wang, G.-P.; Luo, Y.; Wang, W. The protection of wheat plasma membrane under cold stress by glycine betaine overproduction. Biol. Plant. 2010, 54, 83–88. [Google Scholar] [CrossRef]
- Choi, C.; Hwang, C.H. The barley lipid transfer protein, BLT101, enhances cold tolerance in wheat under cold stress. Plant Biotechnol. Rep. 2015, 9, 197–207. [Google Scholar] [CrossRef]
- Levine, T.P. A lipid transfer protein that transfers lipid. J. Cell Biol. 2007, 179, 11–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goddard, N.J.; Dunn, M.A.; Zhang, L.; White, A.H.; Jack, P.L.; Hughes, M.A. Molecular analysis and spatial expression of a low-temperature specific barley gene, blt101. Plant Mol. Biol. 1993, 23, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Armario Najera, V.; Twyman, R.M.; Christou, P.; Zhu, C. Applications of multiplex genome editing in higher plants. Curr. Opin. Biotechnol. 2019, 59, 93–102. [Google Scholar] [CrossRef]
- Awan, M.J.A.; Pervaiz, K.; Rasheed, A.; Amin, I.; Saeed, N.A.; Dhugga, K.S.; Mansoor, S. Genome edited wheat- current advances for the second green revolution. Biotechnol. Adv. 2022, 60, 108006. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Alptekin, B.; Budak, H. CRISPR/Cas9 genome editing in wheat. Funct. Integr. Genom. 2018, 18, 31–41. [Google Scholar] [CrossRef]
- Zheng, M.; Lin, J.; Liu, X.; Chu, W.; Li, J.; Gao, Y.; An, K.; Song, W.; Xin, M.; Yao, Y.; et al. Histone acetyltransferase TaHAG1 acts as a crucial regulator to strengthen salt tolerance of hexaploid wheat. Plant Physiol. 2021, 186, 1951–1969. [Google Scholar] [CrossRef]
- Tian, X.; Qin, Z.; Zhao, Y.; Wen, J.; Lan, T.; Zhang, L.; Wang, F.; Qin, D.; Yu, K.; Zhao, A.; et al. Stress granule-associated TaMBF1c confers thermotolerance through regulating specific mRNA translation in wheat (Triticum aestivum). New Phytol. 2022, 233, 1719–1731. [Google Scholar] [CrossRef]
- Suzuki, N.; Bajad, S.; Shuman, J.; Shulaev, V.; Mittler, R. The transcriptional co-activator MBF1c is a key regulator of thermotolerance in Arabidopsis thaliana. J. Biol. Chem. 2008, 283, 9269–9275. [Google Scholar] [CrossRef]

| Gene | Gene Product | Plant Source | Improved Traits | Ref. |
|---|---|---|---|---|
| Transcription factors | ||||
| GhDREB | Dehydration-responsive element-binding protein | Cotton |
| [62] |
| Osmolytes | ||||
| BADH | Betaine aldehyde dehydrogenase | Atriplex hortensis |
| [136] |
| Other genes | ||||
| BLT101 | Lipid transfer protein | Barley |
| [137] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trono, D.; Pecchioni, N. Candidate Genes Associated with Abiotic Stress Response in Plants as Tools to Engineer Tolerance to Drought, Salinity and Extreme Temperatures in Wheat: An Overview. Plants 2022, 11, 3358. https://doi.org/10.3390/plants11233358
Trono D, Pecchioni N. Candidate Genes Associated with Abiotic Stress Response in Plants as Tools to Engineer Tolerance to Drought, Salinity and Extreme Temperatures in Wheat: An Overview. Plants. 2022; 11(23):3358. https://doi.org/10.3390/plants11233358
Chicago/Turabian StyleTrono, Daniela, and Nicola Pecchioni. 2022. "Candidate Genes Associated with Abiotic Stress Response in Plants as Tools to Engineer Tolerance to Drought, Salinity and Extreme Temperatures in Wheat: An Overview" Plants 11, no. 23: 3358. https://doi.org/10.3390/plants11233358
APA StyleTrono, D., & Pecchioni, N. (2022). Candidate Genes Associated with Abiotic Stress Response in Plants as Tools to Engineer Tolerance to Drought, Salinity and Extreme Temperatures in Wheat: An Overview. Plants, 11(23), 3358. https://doi.org/10.3390/plants11233358