
A Natural Bio-Stimulant Consisting of a Mixture of Fish Protein Hydrolysates and Kelp Extract Enhances the Physiological, Biochemical and Growth Responses of Spinach under Different Water Levels



Abstract
:1. Introduction
2. Results
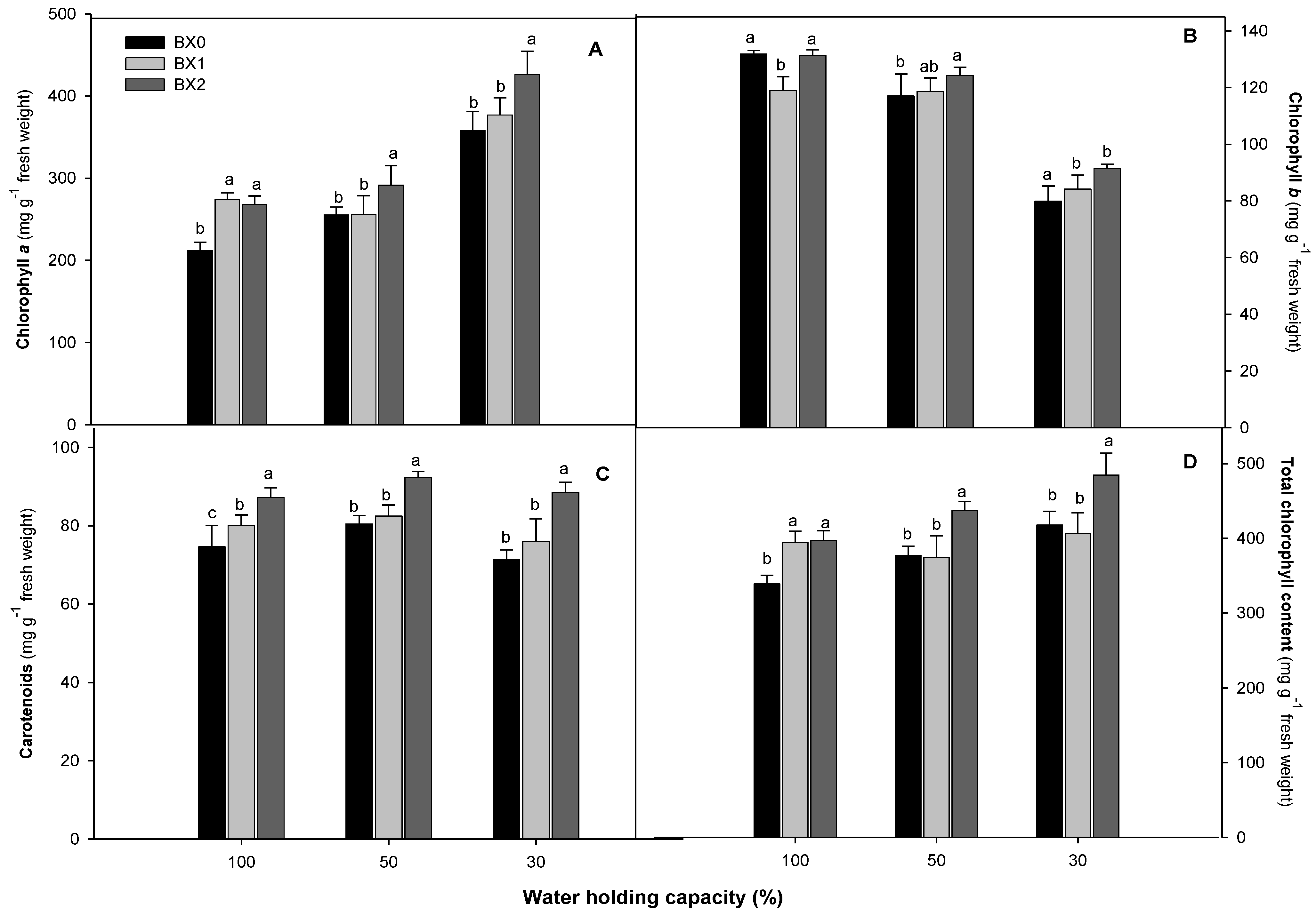
2.1. Photosynthesis Parameters
2.2. Osmolytes and Electrolyte Leakage
2.3. Antioxidant Enzyme Activities
2.4. Vegetative Growth Responses
2.5. The Correlations between the Photosynthetic, Vegetative and Biochemical Responses of Xcell Boost Treated Spinach under Severe Water Deficiency
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Setup
4.2. Chlorophyll a Fluorescence and Normalised Difference Vegetation Index (NDVI)
4.3. Stomatal Conductance
4.4. Chlorophyll and Carotenoid Content
4.5. Determination of Total Soluble Sugars
4.6. Proline Determination
4.7. Determination of Electrolyte Leakage
4.8. Enzyme Extract Preparation, Antioxidative Enzyme and Protein Assays
4.9. Growth Parameters
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roughani, A.; Miri, S. Spinach: An important green leafy vegetable and medicinal herb. In Proceedings of the 2nd International Conference on Medicinal Plants, Organic Farming, Natural and Pharmaceutical Ingredients, Mashhad, Iran, 13 March 2019; pp. 1–6. [Google Scholar]
- Salehi, B.; Tumer, T.B.; Ozleyen, A.; Peron, G.; Dall’Acqua, S.; Rajkovic, J.; Naz, R.; Nosheen, A.; Mudau, F.N.; Labanca, F. Plants of the genus Spinacia: From bioactive molecules to food and phytopharmacological applications. Trends Food Sci. Technol. 2019, 88, 260–273. [Google Scholar] [CrossRef]
- Verma, S. A study on medicinal herb Spinacia oleraceae Linn: Amaranthaceae. J. Drug Deliv. Ther. 2018, 8, 59–61. [Google Scholar] [CrossRef]
- Vasconcelos, A.C.F.d.; Zhang, X.; Ervin, E.H.; Kiehl, J.d.C. Enzymatic antioxidant responses to biostimulants in maize and soybean subjected to drought. Sci. Agric. 2009, 66, 395–402. [Google Scholar] [CrossRef]
- Araujo, J.A.; Abiodun, B.J.; Crespo, O. Impacts of drought on grape yields in Western Cape, South Africa. Theor. Appl. Climatol. 2016, 123, 117–130. [Google Scholar] [CrossRef]
- Maseko, I.; Ncube, B.; Mabhaudhi, T.; Tesfay, S.; Chimonyo, V.; Araya, H.; Fessehazion, M.; Du Plooy, C. Moisture stress on physiology and yield of some indigenous leafy vegetables under field conditions. South Afr. J. Bot. 2019, 126, 85–91. [Google Scholar] [CrossRef]
- Ors, S.; Suarez, D.L. Spinach biomass yield and physiological response to interactive salinity and water stress. Agric. Water Manag. 2017, 190, 31–41. [Google Scholar] [CrossRef]
- Ali, Q.; Ashraf, M. Induction of drought tolerance in maize (Zea mays L.) due to exogenous application of trehalose: Growth, photosynthesis, water relations and oxidative defence mechanism. J. Agron. Crop Sci. 2011, 197, 258–271. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, S.; Chi, Y.; Li, Q.; Huang, K.; Yu, Q. Responses of photosynthetic parameters to drought in subtropical forest ecosystem of China. Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, J.; Wang, Y.; Chi, Y.; Zhou, L.; Chen, J.; Zhou, W.; Song, J.; Zhao, N.; Ding, J. Drought stress strengthens the link between chlorophyll fluorescence parameters and photosynthetic traits. PeerJ 2020, 8, e10046. [Google Scholar] [CrossRef]
- Ceusters, N.; Valcke, R.; Frans, M.; Claes, J.E.; Van den Ende, W.; Ceusters, J. Performance index and PSII connectivity under drought and contrasting light regimes in the CAM orchid Phalaenopsis. Front. Plant Sci. 2019, 10, 1012. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Qiang, S.; Goltsev, V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820-nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochim. Biophys. Acta Bioenerg. 2010, 1797, 1313–1326. [Google Scholar] [CrossRef] [Green Version]
- Živčák, M.; Olšovská, K.; Slamka, P.; Galambošová, J.; Rataj, V.; Shao, H.; Brestič, M. Application of chlorophyll fluorescence performance indices to assess the wheat photosynthetic functions influenced by nitrogen deficiency. Plant Soil Environ. 2015, 60, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Shin, Y.K.; Bhandari, S.R.; Jo, J.S.; Song, J.W.; Lee, J.G. Effect of drought stress on chlorophyll fluorescence parameters, phytochemical contents, and antioxidant activities in lettuce seedlings. Horticulturae 2021, 7, 238. [Google Scholar] [CrossRef]
- Xu, C.; Leskovar, D. Growth, physiology and yield responses of cabbage to deficit irrigation. Hortic. Sci. 2014, 41, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Potopová, V.; Štěpánek, P.; Farda, A.; Türkott, L.; Zahradníček, P.; Soukup, J. Drought stress impact on vegetable crop yields in the Elbe River Lowland between 1961 and 2014. Cuad. De Investig. Geogr. 2016, 42, 127–143. [Google Scholar] [CrossRef] [Green Version]
- Hailemichael, G.; Catalina, A.; González, M.; Martin, P. Relationships between water status, leaf chlorophyll content and photosynthetic performance in Tempranillo vineyards. South Afr. J. Enol. Vitic. 2016, 37, 149–156. [Google Scholar] [CrossRef]
- Ekinci, M.; Ors, S.; Sahin, U.; Yildirim, E.; Dursun, A. Responses to the irrigation water amount of spinach supplemented with organic amendment in greenhouse conditions. Commun. Soil Sci. Plant Anal. 2015, 46, 327–342. [Google Scholar] [CrossRef]
- Reyes, A.; Alvarado, O.; Álvarez-Herrera, J. Effect of irrigation suspension on the growth, water state and production of spinach (Spinacia olerácea L.) plants. Agron. Colomb. 2018, 36, 120–125. [Google Scholar] [CrossRef]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000 Res. 2016, 5, 1554. [Google Scholar] [CrossRef]
- Ache, P.; Bauer, H.; Kollist, H.; Al-Rasheid, K.A.; Lautner, S.; Hartung, W.; Hedrich, R. Stomatal action directly feeds back on leaf turgor: New insights into the regulation of the plant water status from non-invasive pressure probe measurements. Plant J. 2010, 62, 1072–1082. [Google Scholar] [CrossRef]
- Parkash, V.; Singh, S. A review on potential plant-based water stress indicators for vegetable crops. Sustainability 2020, 12, 3945. [Google Scholar] [CrossRef]
- Singh, R.; Parihar, P.; Singh, S.; Mishra, R.K.; Singh, V.P.; Prasad, S.M. Reactive oxygen species signaling and stomatal movement: Current updates and future perspectives. Redox Biol. 2017, 11, 213–218. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Masomi, A.; Kafi, M.; Khazaei, H.R.; Davary, K. Effect of drought stress on water status, elecrolyte leakage and enzymatic antioxidants of kochia (kochia scoparia) under saline condition. Pak. J. Bot. 2010, 42, 3517–3524. [Google Scholar]
- Scotti Campos, P.; Quartin, V.; Cochicho Ramalho, J.; Nunes, M.A. Electrolyte leakage and lipid degradation account for cold sensitivity in leaves of Coffea sp. plants. J. Plant Physiol. 2003, 160, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Torres, M.A. ROS in biotic interactions. Physiol. Plant. 2010, 138, 414–429. [Google Scholar] [CrossRef]
- Akram, N.A.; Waseem, M.; Ameen, R.; Ashraf, M. Trehalose pretreatment induces drought tolerance in radish (Raphanus sativus L.) plants: Some key physio-biochemical traits. Acta Physiol. Plant. 2016, 38, 1–10. [Google Scholar] [CrossRef]
- Shafiq, S.; Akram, N.A.; Ashraf, M. Does exogenously-applied trehalose alter oxidative defense system in the edible part of radish (Raphanus sativus L.) under water-deficit conditions? Sci. Hortic. 2015, 185, 68–75. [Google Scholar] [CrossRef]
- Moloi, M.J.; van der Merwe, R. Drought tolerance responses in vegetable-type soybean involve a network of biochemical mechanisms at flowering and pod-filling stages. Plants 2021, 10, 1502. [Google Scholar] [CrossRef]
- Jabeen, M.; Akram, N.A.; Ashraf, M.; Aziz, A. Assessment of biochemical changes in spinach (Spinacea oleracea L.) subjected to varying water regimes. Sains Malays. 2019, 48, 533–541. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Madende, M.; Hayes, M. Fish by-product use as biostimulants: An overview of the current state of the art, including relevant legislation and regulations within the EU and USA. Molecules 2020, 25, 1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants application in horticultural crops under abiotic stress conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- Fan, D.; Hodges, D.M.; Critchley, A.T.; Prithiviraj, B. A commercial extract of brown macroalga (Ascophyllum nodosum) affects yield and the nutritional quality of spinach in vitro. Commun. Soil Sci. Plant Anal. 2013, 44, 1873–1884. [Google Scholar] [CrossRef]
- Carillo, P.; Colla, G.; Fusco, G.M.; Dell’Aversana, E.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cozzolino, E.; Mori, M.; Reynaud, H. Morphological and physiological responses induced by protein hydrolysate-based biostimulant and nitrogen rates in greenhouse spinach. Agronomy 2019, 9, 450. [Google Scholar] [CrossRef] [Green Version]
- Genc, E.; Atici, Ö. Chicken feather protein hydrolysate as a biostimulant improves the growth of wheat seedlings by affecting biochemical and physiological parameters. Turk. J. Bot. 2019, 43, 67–79. [Google Scholar] [CrossRef] [Green Version]
- Andarwulan, N.; Shetty, K. Improvement of pea (Pisum sativum) seed vigour response by fish protein hydrolysates in combination with acetyl salicylic acid. Process Biochem. 1999, 35, 159–165. [Google Scholar] [CrossRef]
- Xu, C.; Mou, B. Drench application of fish-derived protein hydrolysates affects lettuce growth, chlorophyll content, and gas exchange. Horttechnology 2017, 27, 539–543. [Google Scholar] [CrossRef]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.C.; Bonini, P.; Colla, G. Plant-and seaweed-based extracts increase yield but differentially modulate nutritional quality of greenhouse spinach through biostimulant action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef]
- Pinheiro, C.; Chaves, M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2011, 62, 869–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banks, J.M. Continuous excitation chlorophyll fluorescence parameters: A review for practitioners. Tree Physiol. 2017, 37, 1128–1136. [Google Scholar] [CrossRef] [PubMed]
- Krause, G.H.; Weis, E. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.-W.; Qiang, L.; Rong, J.; Wei, C.; Liu, X.-L.; Kong, F.-L.; Ke, Y.-P.; Shi, H.-C.; Yuan, J.-C. Effect of low-nitrogen stress on photosynthesis and chlorophyll fluorescence characteristics of maize cultivars with different low-nitrogen tolerances. J. Integr. Agric. 2019, 18, 1246–1256. [Google Scholar] [CrossRef]
- Majid, K.; Roza, G. The effect of drought stress on leaf chlorophyll content and stress resistance in maize cultivars (Zea mays). Afr. J. Microbiol. Res. 2012, 6, 2844–2848. [Google Scholar]
- Ping, M.; BAI, T.-h.; WANG, X.-q. Effects of light intensity on photosynthesis and photoprotective mechanisms in apple under progressive drought. J. Integr. Agric. 2015, 14, 1755–1766. [Google Scholar]
- Filimon, R.; Rotaru, L.; Filimon, R. Quantitative investigation of leaf photosynthetic pigments during annual biological cycle of Vitis vinifera L. table grape cultivars. South Afr. J. Enol. Vitic. 2016, 37, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Noroozlo, Y.A.; Souri, M.K.; Delshad, M. Stimulation effects of foliar applied glycine and glutamine amino acids on lettuce growth. Open Agric. 2019, 4, 164–172. [Google Scholar] [CrossRef]
- Gandhi, G.M.; Parthiban, B.; Thummalu, N.; Christy, A. Ndvi: Vegetation change detection using remote sensing and gis–A case study of Vellore District. Procedia Comput. Sci. 2015, 57, 1199–1210. [Google Scholar] [CrossRef] [Green Version]
- Sentera, Inc. NDVI and Your Farm: Understanding NDVI for Plant Health Insights. Available online: https://www.agriculture-xprt.com/news/ndvi-and-your-farm-understanding-ndvi-for-plant-health-insights-702065 (accessed on 18 November 2021).
- Crusiol, L.G.T.; Carvalho, J.d.F.C.; Sibaldelli, R.N.R.; Neiverth, W.; do Rio, A.; Ferreira, L.C.; Procópio, S.d.O.; Mertz-Henning, L.M.; Nepomuceno, A.L.; Neumaier, N. NDVI variation according to the time of measurement, sampling size, positioning of sensor and water regime in different soybean cultivars. Precis. Agric. 2017, 18, 470–490. [Google Scholar] [CrossRef] [Green Version]
- Devnarain, N.; Crampton, B.G.; Chikwamba, R.; Becker, J.V.; O′Kennedy, M.M. Physiological responses of selected African sorghum landraces to progressive water stress and re-watering. South Afr. J. Bot. 2016, 103, 61–69. [Google Scholar] [CrossRef]
- Liu, F.; Song, R.; Zhang, X.; Shahnazari, A.; Andersen, M.N.; Plauborg, F.; Jacobsen, S.-E.; Jensen, C.R. Measurement and modelling of ABA signalling in potato (Solanum tuberosum L.) during partial root-zone drying. Environ. Exp. Bot. 2008, 63, 385–391. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, F.; Jensen, C.R. Comparative effects of deficit irrigation and alternate partial root-zone irrigation on xylem pH, ABA and ionic concentrations in tomatoes. J. Exp. Bot. 2012, 63, 1907–1917. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble sugars: Metabolism, sensing and abiotic stress: A complex network in the life of plants. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [Green Version]
- Suprasanna, P.; Nikalje, G.; Rai, A. Osmolyte accumulation and implications in plant abiotic stress tolerance. In Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Springer: Berlin/Heidelberg, Germany, 2016; pp. 1–12. [Google Scholar]
- Yan, M. Seed priming stimulate germination and early seedling growth of Chinese cabbage under drought stress. South Afr. J. Bot. 2015, 99, 88–92. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Akram, N.A.; Ashraf, M. Osmoprotection in plants under abiotic stresses: New insights into a classical phenomenon. Planta 2020, 251, 1–17. [Google Scholar]
- Patel, K.; Agarwal, P.; Agarwal, P.K. Kappaphycus alvarezii sap mitigates abiotic-induced stress in Triticum durum by modulating metabolic coordination and improves growth and yield. J. Appl. Phycol. 2018, 30, 2659–2673. [Google Scholar] [CrossRef]
- Sahin, U.; Ekinci, M.; Ors, S.; Turan, M.; Yildiz, S.; Yildirim, E. Effects of individual and combined effects of salinity and drought on physiological, nutritional and biochemical properties of cabbage (Brassica oleracea var. capitata). Sci. Hortic. 2018, 240, 196–204. [Google Scholar] [CrossRef]
- Trivedi, K.; Anand, K.V.; Vaghela, P.; Ghosh, A. Differential growth, yield and biochemical responses of maize to the exogenous application of Kappaphycus alvarezii seaweed extract, at grain-filling stage under normal and drought conditions. Algal Res. 2018, 35, 236–244. [Google Scholar] [CrossRef]
- Ihuoma, S.O.; Madramootoo, C.A. Recent advances in crop water stress detection. Comput. Electron. Agric. 2017, 141, 267–275. [Google Scholar] [CrossRef]
- Luoh, J.W.; Begg, C.B.; Symonds, R.C.; Ledesma, D.; Yang, R.-Y. Nutritional yield of African indigenous vegetables in water-deficient and water-sufficient conditions. Food Nutr. Sci. 2014, 2014, 59091. [Google Scholar] [CrossRef]
- Xu, C.; Mou, B. Responses of spinach to salinity and nutrient deficiency in growth, physiology, and nutritional value. J. Am. Soc. Hortic. Sci. 2016, 141, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Introlab. Xcell Boost. Available online: https://www.introlab.co.za/ (accessed on 30 July 2021).
- NFThydro. Available online: https://www.store.nfthydro.co.za/nutrients/23-hygrotech-solu-cal-25kg.html (accessed on 18 February 2022).
- Gadi, V.K.; Hussain, R.; Bordoloi, S.; Hossain, S.; Singh, S.R.; Garg, A.; Sekharan, S.; Karangat, R.; Lingaraj, S. Relating stomatal conductance and surface area with evapotranspiration induced suction in a heterogeneous grass cover. J. Hydrol. 2019, 568, 867–876. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Miehé, J.A. Fluorescence imaging as a diagnostic tool for plant stress. Trends Plant Sci. 1997, 2, 316–320. [Google Scholar] [CrossRef]
- Irigoyen, J.; Einerich, D.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativd) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Carillo, P.; Gibon, Y. Protocol: Extraction and determination of proline. PrometheusWiki 2011, 2011, 1–5. [Google Scholar]
- Rolny, N.; Costa, L.; Carrión, C.; Guiamet, J.J. Is the electrolyte leakage assay an unequivocal test of membrane deterioration during leaf senescence? Plant Physiol. Biochem. 2011, 49, 1220–1227. [Google Scholar] [CrossRef] [PubMed]
- Pukacka, S.; Ratajczak, E. Production and scavenging of reactive oxygen species in Fagus sylvatica seeds during storage at varied temperature and humidity. J. Plant Physiol. 2005, 162, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.P.; Mishra, R.K.; Singhal, G.S. Changes in the activities of anti-oxidant enzymes during exposure of intact wheat leaves to strong visible light at different temperatures in the presence of protein synthesis inhibitors. Plant Physiol. 1993, 102, 903–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zieslin, N.; Ben-Zaken, R. Peroxidases, phenylalanine ammonia-lyase and lignification in peduncles of rose flowers. Plant Physiol. Biochem. 1991, 29, 147–151. [Google Scholar]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta 1976, 133, 21–25. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- González, L.; González-Vilar, M. Determination of relative water content. In Handbook of Plant Ecophysiology Techniques; Springer: Berlin/Heidelberg, Germany, 2001; pp. 207–212. [Google Scholar]
- Ryser, P.; Bernardi, J.; Merla, A. Determination of leaf fresh mass after storage between moist paper towels: Constraints and reliability of the method. J. Exp. Bot. 2008, 59, 2461–2467. [Google Scholar] [CrossRef]
- VSN International. GenStat® for Windows, 19th ed.; VSNI: Hemel Hempstead, UK, 2017. [Google Scholar]
- SAS Institute. SAS® 9.4 Output Delivery System: User′s Guide; SAS Institute: Cary, NC, USA, 2014. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WHC (%) | [Xcell Boost] | Fv/Fm | PIABS | PITotal |
|---|---|---|---|---|
| 100 | BX0 | 0.831 ± 0.0099 a | 7.48 ± 1.13 ab | 6.36 ± 0.58 a |
| BX1 | 0.826 ± 0.0032 a | 6.70 ± 0.65 b | 6.03 ± 0.35 a | |
| BX2 | 0.834 ± 0.0096 a | 8.09 ± 0.98 a | 6.32 ± 0.75 a | |
| 50 | BX0 | 0.836 ± 0.0058 a | 9.71 ± 0.5973 a | 8.09 ± 0.80 a |
| BX1 | 0.832 ± 0.0067 a | 8.77 ± 1.1710 a | 7.09 ± 0.66 b | |
| BX2 | 0.831 ± 0.0035 a | 8.90 ± 0.85 a | 7.40 ± 0.62 b | |
| 30 | BX0 | 0.838 ± 0.0059 a | 8.74 ± 0.53 a | 6.95 ± 0.21 a |
| BX1 | 0.831 ± 0.0045 a | 7.74 ± 0.90 a | 6.20 ± 0.63 a | |
| BX2 | 0.832 ± 0.0073 a | 7.78 ± 0.54 a | 6.28 ± 0.39 a |
| WHC (%) | [Xcell Boost] | PH | RL | LA | LN | LDW | SDW | RDW | RWC | SM | RM |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 100 | BX0 | 23.5 ± 2.1 a | 26.5 ± 3.7 a | 0.8165 ± 87.18 a | 7.5 ± 0.65 a | 4.59 ± 0.6 b | 5.21 ± 0.69 a | 5.3 ± 1.21 a | 76.59± 0.82 b | 90.02 ± 1.04 a | 69.4 ± 1.28 ab |
| BX1 | 24.65 ± 2.2 a | 31.7 ± 2.25 a | 1365 ± 62.31 a | 6.33 ± 0.47 a | 5.48 ± 0.58 ab | 5.43 ± 0.50 a | 5.35 ± 1.31 a | 78.31 ± 1.07 b | 90.96 ± 0.41 a | 71.5 ± 2.79 a | |
| BX2 | 23.7 ± 0.53 a | 29 ± 4.65 a | 1465 ± 79.1 a | 8.25 ± 0.75 a | 5.86 ± 0.45 a | 5.97 ± 0.48 a | 6.92 ± 1.22 a | 80.29 ± 1.97 a | 90.25 ± 1.40 a | 65.8 ± 1.78 b | |
| 50 | BX0 | 23.4 ± 0.54 a | 28. ± 2.16 a | 899 ± 65.35 b | 7.33 ± 0.8 a | 4.39 ± 0.41 a | 3.84 ± 0.42 b | 7 ± 1.1 a | 72.73± 0.89 c | 88.94 ± 1.90 a | 60.6 ± 2.26 a |
| BX1 | 24.8 ± 2.03 a | 32 ± 2.62 a | 1290 ± 56.31 a | 6.75 ± 0.5 a | 4.62 ± 0.47 a | 4.54 ± 0.53 ab | 7.5 ± 1.2 a | 75.4 2± 1.54 b | 89.38 ± 1.32 a | 56.2 ± 4.1 b | |
| BX2 | 22.65 ± 1.65 a | 32.25 ± 4.24 a | 1321 ± 54.2 a | 7.75 ± 0.85 a | 5.38 ± 0.4 a | 5.23 ± 0.63 a | 7.43 ± 1.15 a | 78.93± 1.04 a | 88.83 ± 0.52 a | 62.5 ± 1.02 a | |
| 30 | BX0 | 18.4 ± 0.43 b | 24 ± 3.6 b | 723 ± 17.76 b | 7 ± 0.58 a | 3.5 ± 0.34 a | 3.15 ± 0.76 a | 4.92 ± 1.23 a | 68.35± 1.98 c | 89.75 ± 1.22 a | 56.2 ± 1.8 b |
| BX1 | 21.57 ± 1.2 a | 27 ± 1.41 ab | 799 ± 127.5 ab | 6.5 ± 0.625 a | 3.78 ± 0.15 a | 3.24 ± 0.66 a | 5.65 ± 1.2 a | 72.6± 0.63 b | 89.69 ± 1.26 a | 45.6 ± 4.24 c | |
| BX2 | 19.3 ± 0.65 ab | 32 ± 0.8165 a | 843 ± 41.2 a | 7.87 ± 0.85 a | 4.32 ± 0.39 a | 3.41 ± 0.73 a | 6.17 ± 1.23 a | 76.49 ± 0.69 a | 88.11 ± 0.78 | 61.82 ± 1.93 a |
| LDW | SDW | APX | GR | PIabs | PItotal | SC | NDVI | Chl-b | Chl-total | RWC | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| RL | 0.99 | −0.50 | −0.40 | 0.86 | −0.19 | −0.77 | −0.41 | 0.80 | 0.16 | 0.96 | 0.35 |
| LDW | 1.00 | −0.63 | −0.53 | 0.90 | −0.24 | −0.78 | −0.54 | 0.86 | 0.20 | 0.91 | 0.49 |
| SDW | 0.42 | 1.00 | 0.99 * | −0.65 | 0.27 | 0.56 | 0.99 * | −0.70 | −0.16 | −0.28 | −0.98 * |
| GPX | −0.98 * | −0.31 | 0.78 | 0.43 | −0.81 | 0.35 | −0.62 | 0.53 | 0.76 | −0.35 | 0.56 |
| GR | −0.43 | −0.19 | 0.90 | 1.00 | −0.64 | −0.45 | −0.58 | 0.99 * | 0.59 | 0.70 | 0.50 |
| PIabs | −0.02 | −0.54 | 0.38 | 0.75 | 1.00 | −0.40 | 0.27 | −0.69 | −0.99 * | 0.03 | −0.17 |
| SC | −0.48 | −0.07 | 0.96 * | 0.99 * | 0.63 | 0.78 | 1.00 | −0.65 | −0.17 | −0.18 | −0.99 * |
| Car | −0.19 | 0.19 | −0.43 | −0.77 | −0.92 | −0.97 * | −0.68 | 0.72 | 0.03 | −0.14 | 0.96 |
| Chl-a | 0.04 | −0.02 | −0.74 | −0.92 | −0.78 | −0.98 * | −0.89 | 0.47 | 0.65 | −0.90 | 0.05 |
| Proline | 0.10 | −0.42 | 0.36 | 0.74 | 0.99 * | 0.88 | 0.62 | −0.87 | −0.61 | −0.47 | 0.18 |
| Leaf no | 0.97 * | 0.59 | −0.64 | −0.55 | −0.26 | −0.01 | −0.56 | 0.26 | −0.63 | 0.84 | 0.72 |
| MC leaf | −0.49 | −0.96 * | −0.16 | −0.01 | 0.29 | −0.29 | −0.11 | −0.59 | 0.59 | −0.23 | −0.62 |
| MC root | 0.00 | −0.08 | 0.67 | 0.90 | 0.86 | 0.99 * | 0.85 | −0.58 | −0.66 | −0.63 | −0.64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liatile, P.C.; Potgieter, G.; Moloi, M.J. A Natural Bio-Stimulant Consisting of a Mixture of Fish Protein Hydrolysates and Kelp Extract Enhances the Physiological, Biochemical and Growth Responses of Spinach under Different Water Levels. Plants 2022, 11, 3374. https://doi.org/10.3390/plants11233374
Liatile PC, Potgieter G, Moloi MJ. A Natural Bio-Stimulant Consisting of a Mixture of Fish Protein Hydrolysates and Kelp Extract Enhances the Physiological, Biochemical and Growth Responses of Spinach under Different Water Levels. Plants. 2022; 11(23):3374. https://doi.org/10.3390/plants11233374
Chicago/Turabian StyleLiatile, Pule Clement, Gerhard Potgieter, and Makoena Joyce Moloi. 2022. "A Natural Bio-Stimulant Consisting of a Mixture of Fish Protein Hydrolysates and Kelp Extract Enhances the Physiological, Biochemical and Growth Responses of Spinach under Different Water Levels" Plants 11, no. 23: 3374. https://doi.org/10.3390/plants11233374
APA StyleLiatile, P. C., Potgieter, G., & Moloi, M. J. (2022). A Natural Bio-Stimulant Consisting of a Mixture of Fish Protein Hydrolysates and Kelp Extract Enhances the Physiological, Biochemical and Growth Responses of Spinach under Different Water Levels. Plants, 11(23), 3374. https://doi.org/10.3390/plants11233374