
Microanatomical Changes in the Leaves of Arundo donax (L.) Caused by Potentially Toxic Elements from Municipal Sewage Sediment



Abstract
:1. Introduction
2. Results and Discussion
2.1. Potentially Toxic Elements in Experimental Soil and Arundo Leaves
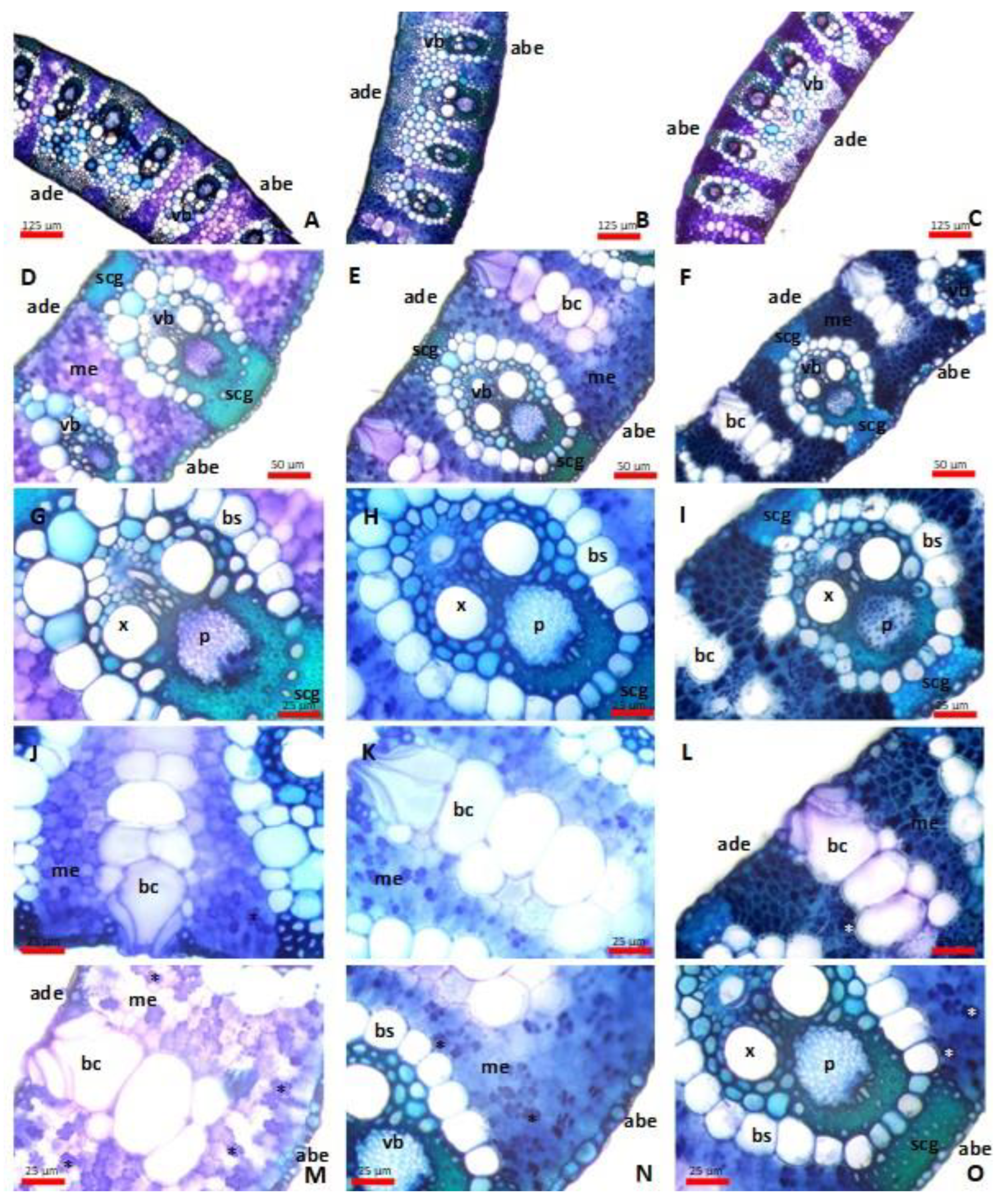
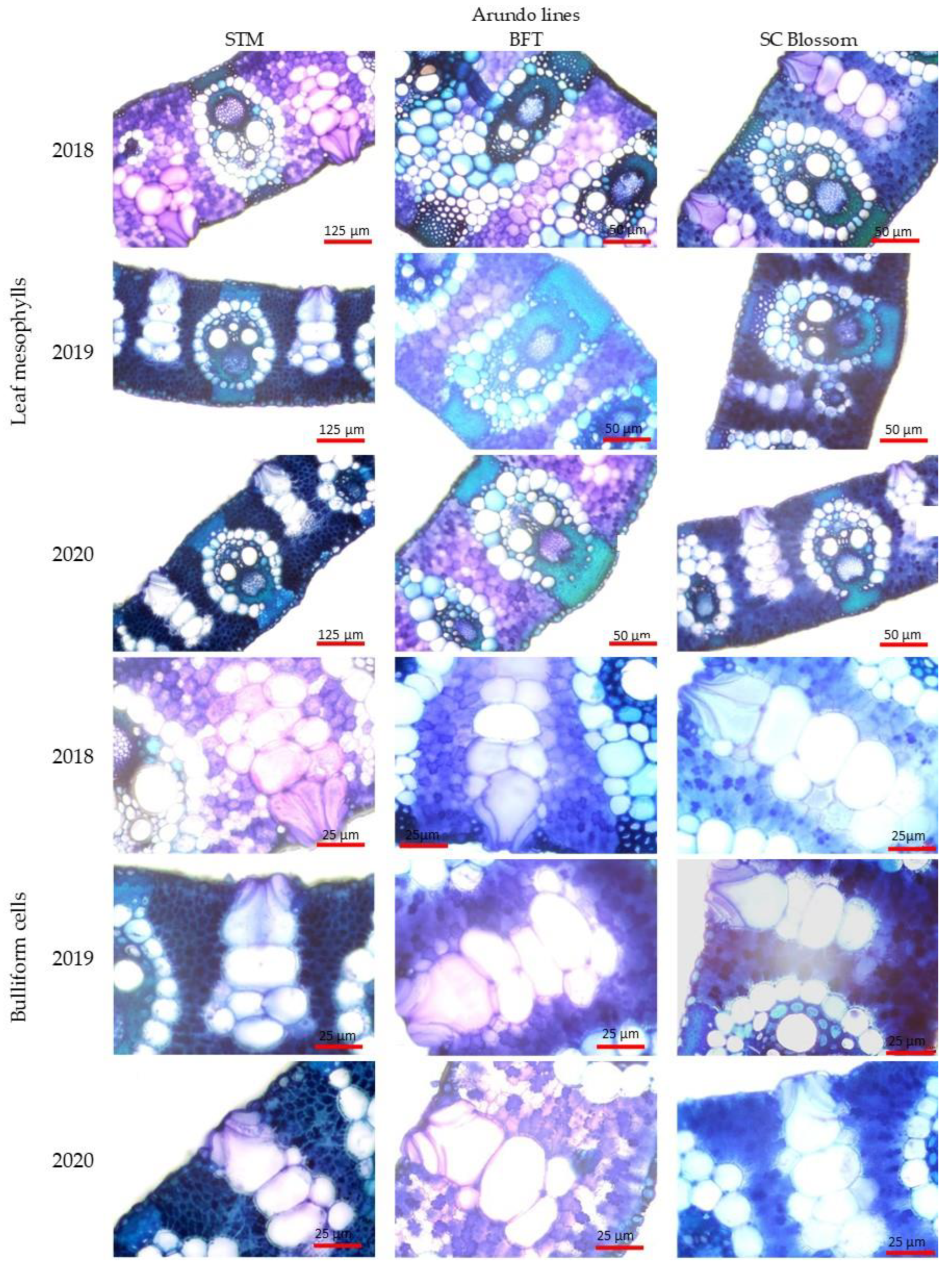
2.2. Anatomical Responses of the Leaf
3. Materials and Methods
3.1. Designation of the Experimental Site, Soil Tests
3.2. Open-Field Experiment with Giant Reed
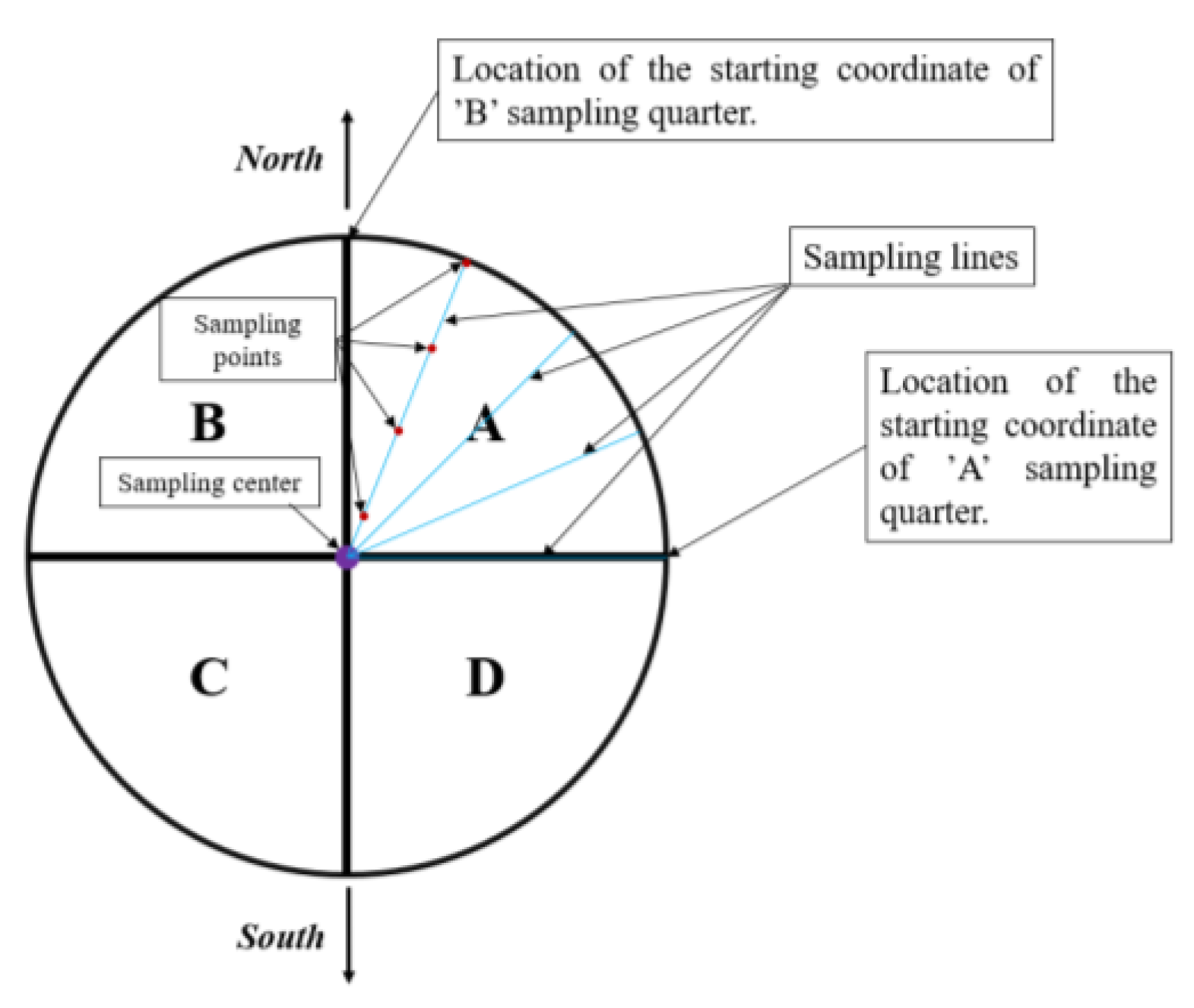
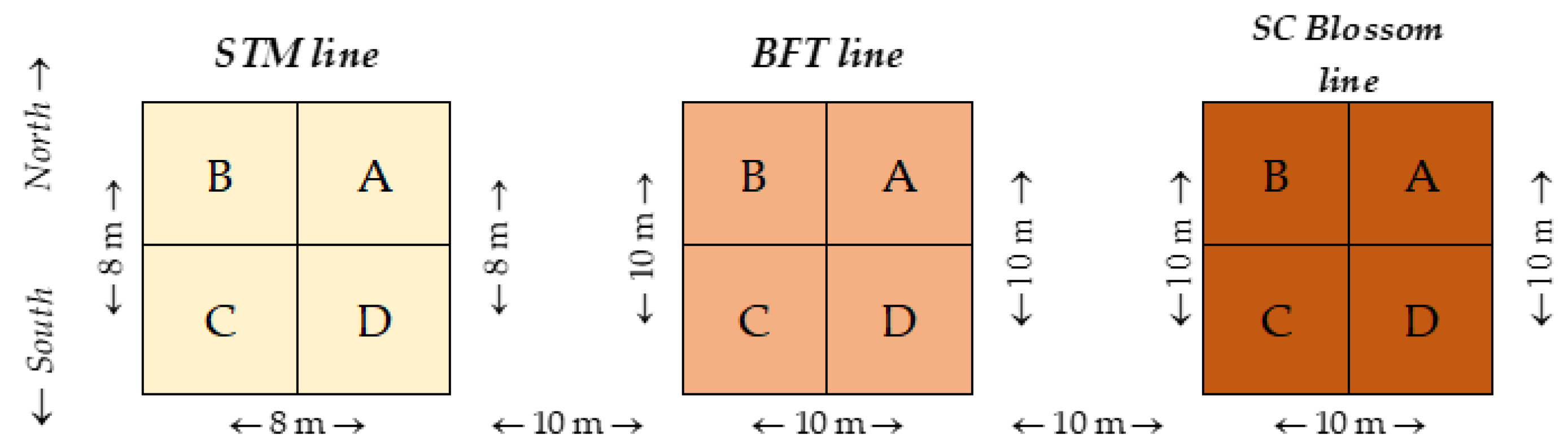
3.3. Plant Sampling during the Experiment
3.4. Soil Samplings at the End of the Experiment
3.5. Element Analysis of Soil and Plant Samples
3.6. Microanatomical Investigations of Leaves
3.7. Bioconcentration Factor (BCF)
3.8. Statistical Analysis of Data
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lino, L.; Espigul, P.; Nogués, S.; Serrat, X. Arundo donax L. growth potential under different abiotic stress. Heliyon 2023, 9, e15521. [Google Scholar] [CrossRef]
- Angelini, L.G.; Ceccarini, L.; Di Nasso, N.N.; Bonari, E. Comparison of Arundo donax and Miscanthus x giganteus in a long-term field experiment in Central Italy: Analysis of productive characteristics and energy balance. Biomass Bioenergy 2009, 33, 635–643. [Google Scholar] [CrossRef]
- Lewandowski, I.; Scurlock, J.M.O.; Lindvall, E.; Christou, M. The development and current status of perennial rhizomatous grasses as energy crops in the US and Europe. Biomass Bioenergy 2003, 25, 335–361. [Google Scholar] [CrossRef]
- Corno, L.; Pilu, R.; Adani, F. Arundo donax L.: A non-food crop for bioenergy and bio-compound production. Biotechnol. Adv. 2014, 32, 1535–1549. [Google Scholar] [CrossRef] [PubMed]
- Nsanganwimana, F.; Marchand, L.; Douay, F.; Mench, M. Arundo donax L., a candidate for phytomanaging water and soils contaminated by trace elements and producing plant-based feedstock. A review. Int. J. Phytoremediation 2014, 16, 982–1017. [Google Scholar] [CrossRef] [PubMed]
- Simon, L. Cultivation and utilization of giant reed (Arundo donax L.) (review). Növényterm (Crop Production) 2017, 66, 89–109. (In Hungarian) [Google Scholar]
- Simon, L.; Szabó, B.; Szabó, M.; Vincze, G.; Varga, C.; Uri, Z.; Koncz, J. Effect of various soil amendments on the mineral nutrition of Salix viminalis and Arundo donax energy crops. Eur. Chem. Bull. 2013, 2, 18–21. [Google Scholar] [CrossRef]
- Papazoglou, E.G.; Karantounias, G.A.; Vemmos, S.N.; Bouranis, D.L. Photosynthesis and growth responses of giant reed (Arundo donax L.) to the heavy metals Cd and Ni. Environ. Int. 2005, 31, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Mirza, N.; Pervez, A.; Mahmood, Q.; Shah, M.M.; Shafqat, M.N. Ecological restoration of arsenic contaminated soil by Arundo donax L. Ecol. Eng. 2011, 37, 1949–1956. [Google Scholar] [CrossRef]
- Yang, M.; Xiao, X.-Y.; Miao, X.-F.; Guo, Z.-H.; Wang, F.-Y. Effect of amendments on growth and metal uptake of giant reed (Arundo donax L.) grown on soil contaminated by arsenic, cadmium and lead. Trans. Nonferrous Met. Soc. China 2012, 22, 1462–1469. [Google Scholar] [CrossRef]
- Bonanno, G. Comparative performance of trace element bioaccumulation and biomonitoring in the plant species Typha domingensis, Phragmites australis and Arundo donax. Ecotoxicol. Environ. Saf. 2013, 97, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Ammari, T.G. Utilization of a natural ecosystem bio-waste; leaves of Arundo donax reed, as a raw material of low-cost eco-biosorbent for cadmium removal from aqueous phase. Ecol. Eng. 2014, 71, 466–473. [Google Scholar] [CrossRef]
- Elhawat, N.; Alshaal, T.; Domokos-Szabolcsy, É.; El-Ramady, H.; Antal, L.; Márton, L.; Czakó, M.; Balogh, P.; Fári, M.G. Copper uptake efficiency and its distribution within bioenergy grass giant reed. Bull. Environ. Contam. Toxicol. 2015, 95, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Bonanno, G.; Cirelli, G.L.; Toscano, A.; Giudice, R.L.; Pavone, P. Heavy metal content in ash of energy crops growing in sewage-contaminated natural wetlands: Potential applications in agriculture and forestry? Sci. Total Environ. 2013, 452–453, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Cano-Ruiz, J.; Plaza, A.; Pinilla, P.; Mauri, P.V.; Lobo, M.C. Valorization of treated sewage sludge for Arundo donax production in a field experiment. Ind. Crops Prod. 2021, 170, 113702. [Google Scholar] [CrossRef]
- Liu, Y.-N.; Guo, Z.-H.; Xiao, X.-Y.; Wang, S.; Jiang, Z.-C.; Zeng, P. Phytostabilisation potential of giant reed for metals contaminated soil modified with complex organic fertiliser and fly ash: A field experiment. Sci. Total Environ. 2017, 576, 292–302. [Google Scholar] [CrossRef]
- Abo-Donia, F.M.; Soliman, N.A.M.; El-Zalaki, U.A. Nutritional and economical feasibility of using reeds (Arundo donax, L) silage compared to corn (Zea mays L) silage as sheep feed. Egypt J. Nutr. Feeds 2009, 12, 243–256. [Google Scholar]
- Tóth, C.; Simon, L. Impact of ammonium-nitrate artificial fertilizer, municipal biocompost and municipal sewage sludge compost on the leaf microanatomy of giant reed (Arundo donax L.). Növényterm 2018, 67, 69–84. (In Hungarian) [Google Scholar]
- Guo, Z.H.; Miao, X.F. Growth changes and tissues anatomical characteristics of giant reed (Arundo donax L.) in soil contaminated with arsenic, cadmium and lead. J. Cent. South Univ. Technol. 2010, 17, 770–777. [Google Scholar] [CrossRef]
- Yadav, V.; Arif, N.; Kovac, J.; Singh, V.P.; Tripathi, D.K.; Chauhan, D.K.; Vaculik, M. Structural modifications of plant organs and tissues by metals and metalloids in the environment: A review. Plant Physiol. Biochem. 2021, 159, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.P.; Lanza de Sá e Melo Marques, T.C.; de Oliveira Gonçalves Nogueira, M.; Mauro de Castro, E.; Soares, A.M. Ecophysiological and anatomical changes due to uptake and accumulation of heavy metal in Brachiaria decumbens. Sci. Agric. 2011, 68, 566–573. [Google Scholar] [CrossRef]
- Guo, Z.; Gao, Y.; Yuan, X.; Yuan, M.; Huang, L.; Wang, S.; Liu, C.; Duan, C. Effects of Heavy Metals on Stomata in Plants. A Review. Int. J. Mol. Sci. 2023, 24, 9302. [Google Scholar] [CrossRef]
- Shi, G.R.; Cai, Q.S. Photosynthetic and anatomic responses of peanut leaves to cadmium stress. Photosynthetica 2008, 46, 627–630. [Google Scholar] [CrossRef]
- Shi, G.R.; Cai, Q.S. Photosynthetic and anatomic responses of peanut leaves to zinc stress. Biol. Plant. 2009, 53, 391–394. [Google Scholar] [CrossRef]
- Gostin, I.N. Air pollution effects on the leaf structure of some Fabaceae species. Not. Bot. Hort. Agrobot. Cluj. 2009, 37, 57–63. [Google Scholar]
- Kastori, R.; Petrović, M.; Petrović, N. Effects of excess lead, cadmium, copper and zinc on water relations in sunflower. J. Plant Nutr. 1992, 15, 2427–2439. [Google Scholar] [CrossRef]
- Chwil, M. The influence of lead on structure of Cucumis sativus L. leaves. Folia Horticulturae. 2005, 17, 11–22. [Google Scholar]
- Shi, G.; Cai, Q. Leaf plasticity in peanut (Arachis hypogaea L.) in response to heavy metal stress. Environ. Exp. Bot. 2009, 67, 112–117. [Google Scholar] [CrossRef]
- Wahid, A.; Ghani, A.; Javed, F. Effect of cadmium on photosynthesis, nutrition and growth of mungbean. Agron. Sustain. Dev. 2008, 28, 273–280. [Google Scholar] [CrossRef]
- Orcen, N. Stomatal Parameters and Growth Responses of Tomatal Nicotiana and Atriplex to Cd, Pb and Cd-Pb-Contaminated Soil. Fresenius Environ. Bull. 2017, 26, 6340–6345. [Google Scholar]
- Greger, M.; Johansson, M. Cadmium effects on leaf transpiration of sugar beet (Beta vulgaris). Physiol. Plant. 1992, 86, 465–473. [Google Scholar] [CrossRef]
- Gálusová, T.; Piršelová, B.; Rybanský, L.; Krasylenko, Y.; Mészáros, P.; Blehová, A.; Bardáčová, M.; Moravčíková, J.; Matušíková, I. Plasticity of Soybean Stomatal Responses to Arsenic and Cadmium at the Whole Plant Level. Pol. J. Environ. Stud. 2020, 29, 3569–3580. [Google Scholar] [CrossRef] [PubMed]
- Kosobrukhov, A.; Knyazeva, I.; Mudrik, V. Plantago major plants responses to increase content of lead in soil: Growth and photosynthesis. Plant Growth Regul. 2004, 42, 145–151. [Google Scholar] [CrossRef]
- Kasim, W.A. Changes induced by copper and cadmium in the anatomy and grain yield of Sorghum bicolor (L.) Moench. Int. J. Agric. Biol. 2006, 8, 123–128. [Google Scholar]
- Baryla, A.; Carrier, P.; Franck, F.; Coulomb, C.; Sahut, C.; Havaux, M. Leaf cloroses in oilseed rape plants (Brassica napus) grown on cadmium polluted soil: Causes and consequences for photosynthesis and growth. Planta 2001, 212, 696–709. [Google Scholar] [CrossRef] [PubMed]
- Makovníková, J. Distribution of Cd and Pb in main soil types of Slovakia. Agriculture 2001, 47, 903–911. [Google Scholar]
- Verma, R.B.; Mahmooduzzafar Siddiqui, T.O.; Iqbal, M. Foliar response of Ipomea pes-tigridis L. to coal-smoke pollution. Turk. J. Bot. 2006, 30, 413–417. [Google Scholar]
- Piršelová, B.; Kuna, R.; Lukáč, P.; Havrlentová, M. Effect of cadmium on growth, photosynthesis pigments, iron and cadmium accumulation of faba bean (Vicia faba cv. Aštar). Agriculture 2016, 62, 72–79. [Google Scholar]
- Pereira, M.P.; Correa, F.F.; Castro, E.M.; Ribeiro, V.E.; Pereira, F.J. Cadmium tolerance in Schinus molle trees is modulated by enhanced leaf anatomy and photosynthesis. Trees 2016, 30, 807–814. [Google Scholar] [CrossRef]
- Azmat, R.; Haider, S.; Nasreen, H.; Aziz, F.; Riaz, M. A viable alternative mechanism in adapting the plants to heavy metal environment. Pak. J. Bot. 2009, 41, 2729–2738. [Google Scholar]
- Piršelová, B.; Kubová, V.; Boleček, P.; Hegedűsová, A. Impact of cadmium toxicity on leaf area and stomatal characteristics in Faba bean. Microbiol. Biotech. Food Sci. 2021, 11, 1–4. [Google Scholar] [CrossRef]
- Wainwright, S.J.; Woolhouse, H.W. Some physiological aspects of copper and zinc tolerance in Agrostis tenuis sibth.: Cell elongation and membrane damage. J. Exp. Bot. 1977, 28, 1029–1036. [Google Scholar] [CrossRef]
- Tran, T.A. Cadmium-induced structural disturbances in pea leaves are alleviated by nitric oxide. Turk. J. Bot. 2013, 37, 698–707. [Google Scholar] [CrossRef]
- Weryszko-Chmielewska, E.; Chwil, M. Lead-Induced Histological and Ultrastructural Changes in the Leaves of Soybean (Glycine max (L.) Merr.). Soil Sci. Plant Nutr. 2005, 51, 203–212. [Google Scholar] [CrossRef]
- Nazir, F.; Hussain, A.; Fariduddin, Q. Interactive role of epibrassinolide and hydrogen peroxide in regulating stomatal physiology, root morphology, photosynthetic and growth traits in Solanum lycopersicum L. under nickel stress. Environ. Exp. Bot. 2019, 162, 479–495. [Google Scholar] [CrossRef]
- Vezza, M.E.; Llanes, A.; Travaglia, C.; Agostini, E.; Talano, M.A. Arsenic stress effects on root water absorption in soybean plants: Physiological and morphological aspects. Plant Physiol. Biochem. 2018, 123, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Stancheva, I.; Geneva, M.; Markovska, Y.; Tzvetkova, N.; Mitova, I.; Todorova, M.; Petrov, P. A comparative study on plant morphology, gas exchange parameters, and antioxidant response of Ocimum basilicum L. and Origanum vulgare L. grown on industrially polluted soil. Turk. J. Biol. 2014, 38, 89–102. [Google Scholar] [CrossRef]
- Barceló, P.D.J.; Vaschez, M.D.; Poschenrieder, C. Cadmium-Induced Structural and Ultrastructural Changes in the Vascular System of Bush Bean Stems. Bot. Acta 1988, 101, 254–261. [Google Scholar] [CrossRef]
- Bini, C.; Wahsha, M.; Fontana, S.; Maleci, L. Effects of heavy metals on morphological characteristics of Taraxacum officinale Web. growing on mine soils in NE Italy. J. Geochem. Explor. 2012, 123, 101–108. [Google Scholar] [CrossRef]
- Sridhar, M.B.B.; Diehl, S.V.; Han, F.X.; Monts, D.L.; Su, Y. Anatomical changes due to uptake and accumulation of Zn an Cu in Indian mustard (Brassica juncea). Environ. Exp. Bot. 2005, 54, 131–141. [Google Scholar] [CrossRef]
- André, O.; Vollenweider, P.; Günthardt-Goerg, M.S. Foliage response to heavy metal contamination in Sycamore Maple (Acer pseudoplatanus L.). For. Snow Landsc. Res. 2006, 80, 275–288. [Google Scholar]
- da Silva de Jesus, D.; Martins, F.M.; de Azevedo Neto, A.D. Structural changes in leaves and roots are anatomical markers of aluminum sensitivity in sunflower. Pesq. Agropec. Trop. Goiânia 2016, 46, 383–390. [Google Scholar] [CrossRef]
- Vollenweider, P.; Cosio, C.; Günthardt-Goerg, M.; Keller, C. Localization and effects of cadmium in leaves of a cadmium-tolerant willow (Salix viminalis L.). II. microlocalization and cellular effects of cadmium. Environ. Exp. Bot. 2006, 58, 25–40. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 39–397. [Google Scholar] [CrossRef] [PubMed]
- Currie, H.A.; Perry, C.C. Silica in plants: Biological: Biochemical and chemical studies. Ann. Bot. 2007, 100, 1383–1389. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Samson, N.P.; Ravichandran, P.; Narasimhan, D.; Dayanandan, P. Phytoliths of Indian grasses and their potential use in identification. Bot. J. Linn. Soc. 2000, 132, 241–252. [Google Scholar] [CrossRef]
- Hodson, M.J.; Evans, D.E. Aluminium/silicon interactions in higher plants. J. Exp. Bot. 1995, 46, 161–171. [Google Scholar] [CrossRef]
- MSZ 21470-50; Environmental Testing of Soils. Determination of Total and Soluble Toxic Element, Heavy Metal and Chromium (VI) Content. Hungarian Standards Association: Budapest, Hungary, 2006. (In Hungarian)
- Shakoor, S.A.; Bhat, M.A.; Soodan, A.S. Taxonomic demarcation of Arundo donax L. and Phragmites karka (Retz.) Trin. ex Steud. (Arundinoideae, Poaceae) from phytolith signatures. Flora 2016, 224, 130–153. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: Boca Raton, FL, USA, 2011; Volume 2. [Google Scholar]
- Simon, L. Potentially harmful elements in agricultural soils. In Environment and Human Health. Potentially Harmful Elements in the Environment and the Impact on Human Health; Bini, C., Bech, J., Eds.; Springer: Dordrecht, The Netherlands; Berlin/Heidelberg, Germany; New York, NY, USA; London, UK, 2014; pp. 85–137, 142–150. ISBN 978-94-017-8964-6. [Google Scholar]
- Buscaroli, A. An overview of indexes to evaluate terrestrial plants for phytoremediation purposes (review). Ecol. Indic. 2017, 82, 367–380. [Google Scholar] [CrossRef]
- Rossa, B.; Tüffers, A.V.; Naidoo, G.; von Willert, D.J. Arundo donax L. (Poaceae)—A C3 species with unusually high photosynthetic capacity. Bot. Acta 1998, 111, 216–221. [Google Scholar] [CrossRef]
- Alshaal, T.; Elhawat, N.; Domokos-Szabolcsy, É.; Kátai, J.; Márton, L.; Czako, M.; El-Ramady, H.; Fári, M. Giant reed (Arundo donax L.): A green technology for clean environment. In Phytoremediation: Management of Environmental Contaminants; Ansari, A.A., Gill, S.S., Gill, R., Lanza, G.R., Newman, L., Eds.; Springer Science Business Media B.V.: Berlin/Heidelberg, Germany, 2015; Volume I, pp. 3–20. [Google Scholar]
- Kering, M.K.; Butler, T.J.; Biermacher, J.T.; Guretzky, J.A. Biomass yield and nutrient removal rates of perennial grasses under nitrogen fertilization. Bioenergy Res. 2012, 5, 61–70. [Google Scholar] [CrossRef]
- Sylvester, A.; Parker-Clark, V.; Murray, G. Leaf shape and anatomy as indicators of phase change in the grasses: Comparison of maize, rice, and bluegrass. Am. J. Bot. 2001, 88, 2157–2167. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.P. Agrostology; an introduction to the systematics of grasses. Bot. Stud. 2015, 10. Available online: https://digitalcommons.humboldt.edu/cgi/viewcontent.cgi?article=1000&context=botany_jps (accessed on 3 March 2024).
- Khan, R.; Abidin, S.Z.U.; Mumtaz, A.S.; Jamsheed, S.; Ullah, H. Comparative leaf and pollen micromorphology on some grasses taxa (Poaceae) distributed in Pakistan. Int. J. Nat. Life Sci. 2017, 1, 72–82. [Google Scholar]
- Chauhan, D.K.; Tripathi, D.K.; Kumar, D.; Kumar, Y. Diversity, Distribution and Frequency Based Attributes of Phytolith in Arundo donax L. Int. J. Innov. Biol. Chem. Sci. 2011, 1, 22–27. [Google Scholar]
- Shakoor, S.; Soodan, A.S.; Kumar, K. Morphological diversity and frequency of phytolith types in gaint reed Arundo donax (L.). World Appl. Sci. J. 2014, 29, 926–932. [Google Scholar]
- Fernandez Honaine, M.; Osterrieth, M.L. Silicification of the adaxial epidermis of leaves of a panicoid grass in relation to leaf position and section and environmental conditions. Plant Biol. 2012, 14, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Pepim, G.F.; Zucola, F.; Arriagam, O. Comparative phytolith analysis of festuca (Pooideae: Poaceae) species native to tierra del fuego, Argentina. Botany 2012, 90, 1113–1124. [Google Scholar]
- de Souza, A.E.; Rios, C.O.; de Araújo, T.O. Is a C4 tropical grass still an option in the revegetation of iron ore tailings in face of climate change? Theor. Exp. Plant Physiol. 2021, 33, 397–409. [Google Scholar] [CrossRef]
- Tóth, C.; Irinyiné-Oláh, K.; Simon, L. The effect of sewage sediment containing toxic elements on the microanatomy of the leaf of Sorghum species. In International Multidisciplinary Conference, 14th ed.; Páy, G.L., Ed.; University of Nyíregyháza: Nyíregyháza, Hungary, 2022; pp. 124–133. [Google Scholar]
- Zhao, F.J.; Lombi, E.; Breedon, T.; McGrath, S.P. Zinc hyperaccumulation and cellular distribution in Arabidopsis halleri. Plant Cell Environ. 2000, 23, 507–514. [Google Scholar] [CrossRef]
- Sridhar, B.B.M.; Han, F.X.; Diehl, V.S.; Monts, D.L.; Su, Y. Effects of Zn and Cd accumulation on structural and physiological characteristics of barley plants. Braz. J. Plant. Physiol. 2007, 19, 15–22. [Google Scholar] [CrossRef]
- Santana, B.V.N.; de Araújo, T.O.; Andrade, G.C.; de Freitas-Silva, L.; Kuki, K.N.; Pereira, E.G.; Azevedo, A.A.; da Silva, L.C. Leaf morphoanatomy of species tolerant to excess iron and evaluation of their phytoextraction potential. Environ. Sci. Pollut. Res. 2014, 21, 2550–2562. [Google Scholar] [CrossRef] [PubMed]
- Koleva, L.; Semerdjieva, I.; Nikolova, A.; Vassilev, A. Comparative morphological and histological study on zinc- and cadmium-treated durum wheat plants with similar growth inhibition. Gen. Appl. Plant Physiol. 2010, 36, 8–11. [Google Scholar]
- Müller, C.; Kuki, K.N.; Pinheiro, D.T.; de Souza, L.R.; Silva, A.I.S.; Loureiro, M.E.; Oliva, M.A.; Almeida, A.M. Differential physiological responses in rice upon exposure to excess distinct iron forms. Plant Soil. 2015, 391, 123–138. [Google Scholar] [CrossRef]
- Brandão, M.C.S.; Martins, F.M.; Accioly, A.M.A. Phytoremediation potential and morphological changes of plants growing in the vicinity of lead smelter plant. Int. J. Environ. Sci. Technol. 2018, 15, 361–372. [Google Scholar] [CrossRef]
- Baas, P.; Werker, E.; Fahn, A. Some ecological trends in vessel characters. Iawa Bull. 1983, 4, 141–159. [Google Scholar] [CrossRef]
- Sandalio, L.M.; Dalurzo, H.C.; Gómes, M.; Romero-Puertas, M.C.; Del Rio, L.A. Cadmium-induced changes in the growth and oxidative metabolism of pea plants. J. Exp. Bot. 2001, 52, 2115–2126. [Google Scholar] [CrossRef]
- Turner, N.C.; Jones, M.M. Turgor maintenance by osmotic adjustment: Are view and evaluation. In Adaptation of Plants to Water and High Temperaturen Stress; Turner, N.C., Kramer, P.J., Eds.; CAB International: Wallingford, UK, 1980; pp. 87–103. [Google Scholar]
- Solìs-Domìnguez, F.A.; Gonzàlez-Chàvez, M.C.; Carrillo-Gonzàlez, R.; Rodrìguez-Vàzquez, R. Accumulation and localization of cadmium in Echinochloa polystachya grown within a hydro ponic system. J. Hazard Mater. 2007, 141, 630–636. [Google Scholar] [CrossRef]
- Tőzsér, D. Investigation the Phytoremediation of Heavy Metal Contaminated Soils. Ph.D. Thesis, University of Debrecen, Debrecen, Hungary, 2018; pp. 1–148. (In Hungarian). [Google Scholar]
- Tőzsér, D.; Harangi, S.; Baranyai, E.; Lakatos, G.; Fülöp, Z.; Tóthmérész, B.; Simon, E. Phytoextraction with Salix viminalis in a moderately to strongly contaminated area. Environ. Sci. Pollut. R. 2018, 25, 3275–3290. [Google Scholar] [CrossRef]
- Simon, L.; Uri, Z.S.; Vigh, S.Z.; Irinyiné Oláh, K.; Makádi, M.; Vincze, G.Y. Phytoextraction of potentially toxic elements from wastewater solids and willow ash—Experiences with energy willow (Salix triandra × S. viminalis ’Inger’). In Bioenergy Crops: A Sustainable Means of Phytoremediation, 1st ed.; Puthur, J.T., Dhankher, O.P., Eds.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2022; Chapter 13; pp. 227–245. ISBN 9781003043522. [Google Scholar] [CrossRef]
- Simon, L.; Makádi, M.; Uri, Z.S.; Vigh, S.Z.; Irinyiné Oláh, K.; Vincze, G.Y.; Tóth, C.S. Phytoextraction of toxic elements and chlorophyll fluorescence in the leaves of energy willow (Salix sp.), treated with wastewater solids and wood ash. Agrokem Talajtan. (Agrochem. Soil Sci.) 2022, 71, 77–99. [Google Scholar] [CrossRef]
- Antal, G. Some Bio-Economic Issues of the Propagation and Cultivation of Giant Reed Using Biotechnological Methods. Ph.D. Thesis, Faculty of Economic Science, University of Debrecen, Debrecen, Hungary, 2018; pp. 1–150. (In Hungarian). [Google Scholar]
- Alshaal, T. Remediation and Restoring Marginal Lands with Biotechnologically Propagated Giant Reed (Arundo donax L.). Ph.D. Thesis, Faculty of Agriculture, Food Science and Environmental Management, University of Debrecen, Debrecen, Hungary, 2013; pp. 1–109. (In Hungarian). [Google Scholar]
- Márton, L.; Czakó, M. Sustained Totipotent Culture of Selected Monocot Genera. U.S. Patent 6,821,782, 23 November 2004. [Google Scholar]
- Barykina, R.P. Guide on Botanical Microtechique; Base and Methods; MSU: Moscow, Russia, 2004; p. 312. [Google Scholar]
- MSZ 20135; Determination of the Soluble Nutrient Element Content of the Soil. Hungarian Standards Board: Budapest, Hungary, 1999. (In Hungarian)
- Hilu, K.W.; Randall, J.L. Convenient method for studying grass leaf epidermis. Taxon 1984, 33, 413–415. [Google Scholar] [CrossRef]
- Gardner, S.D.L.; Taylor, G.; Bosac, C. Leaf growth of hybrid poplar following exposure to elevated CO2. New Phytol. 1995, 131, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Elagöz, V.; Han, S.S.; Manning, W.J. Acquired changes in stomatal characteristics in response to ozone during plant growth and leaf development of bush beans (Phaseolus vulgaris L.) indicate phenotypic plasticity. Environ. Poll. 2006, 140, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Tóth, C.S.; Vincze, G.Y.; Irinyiné-Oláh, K.; Uri, Z.S.; Vigh, S.Z.; Simon, L. The effect of toxic elements on the microanatomy of the leaves of the Salix alba L. Rev. Agricult. Rural Develop. 2022, 11, 139–145. [Google Scholar] [CrossRef]
- Sass, J.E. Botanical Microtechnique, 2nd ed.; Iowa State College Press: Ames, IA, USA, 1951. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PTE µg g−1 | Arundo Lines | ||
|---|---|---|---|
| STM (Location 1) | BFT (Location 2) | SC Blossom (Location 3) | |
| As | 9.85 a | 12.5 b | 13.9 c |
| Ba | 163 a | 434 c | 347 b |
| Cd | 0.301 a | 0.579 b | 0.731 c |
| Cr | 126 a | 310 b | 341 c |
| Cu | 39.3 a | 82.8 b | 88.4 c |
| Mn | 293 a | 411 b | 427 c |
| Ni | 21.9 a | 33.9 b | 37.1 c |
| Pb | 33.9 a | 92.5 b | 111 c |
| Zn | 156 a | 413 b | 435 c |
| PTE µg g−1 | Arundo Lines | ||
|---|---|---|---|
| STM (Location 1) | BFT (Location 2) | SC Blossom (Location 3) | |
| 13 September 2018 | |||
| As | 0.094 b | 0.103 c | 0.065 a |
| Ba | 1.97 c | 1.82 b | 1.72 a |
| Cd | 0.329 b | 0.379 c | 0.270 a |
| Cr | 0.297 b | 0.564 c | 0.146 a |
| Cu | 13.2 b | 13.4 b | 11.4 a |
| Mn | 156 a | 179 b | 159 a |
| Ni | 0.611 b | 1.05 c | 0.468 a |
| Pb | 0.620 b | 1.28 c | 0.425 a |
| Zn | 170 b | 187 c | 147 a |
| 17 October 2019 | |||
| As | 0.209 a | 0.272 b | 0.211 a |
| Ba | 2.25 b | 2.60 c | 1.96 a |
| Cd | 0.191 a | 0.297 b | 0.200 a |
| Cr | 0.070 a | 0.111 b | 0.079 a |
| Cu | 7.65 a | 10.8 b | 8.33 a |
| Mn | 111 a | 194 c | 128 b |
| Ni | 0.507 a | 1.05 c | 0.583 b |
| Pb | 0.369 a | 0.617 b | 0.392 a |
| Zn | 59.8 c | 117 b | 70.2 a |
| 25 June 2020 | |||
| As | 0.214 a | 0.315 b | 0.224 a |
| Ba | 3.60 a | 5.89 b | 3.39 a |
| Cd | 0.501 b | 0.634 c | 0.467 a |
| Cr | 2.46 b | 2.94 c | 2.24 a |
| Cu | 8.39 a | 10.6 c | 9.88 b |
| Mn | 68.7 a | 107 c | 91.3 b |
| Ni | 3.17 a | 3.55 b | 3.05 a |
| Pb | 0.337 a | 0.624 c | 0.376 b |
| Zn | 83.9 a | 103 c | 92.7 b |
| PTE (mg kg−1) | Arundo Lines | ||
|---|---|---|---|
| STM (Location 1) | BFT (Location 2) | SC Blossom (Location 3) | |
| “Pseudo-total” * | Soil depth 0–30 cm | ||
| As | 9.00 a | 10.1 b | 11.5 c |
| Ba | 158 a | 214 a | 293 b |
| Cd | 0.862 a | 1.28 b | 1.72 c |
| Cr | 133 a | 149 a | 267 b |
| Cu | 39.9 a | 44.2 b | 72.1 a |
| Mn | 403 a | 426 a | 425 a |
| Ni | 21.9 a | 26.2 a | 35.4 b |
| Pb | 28.3 a | 41.6 b | 67.1 c |
| Zn | 159 a | 291 a | 393 b |
| Soil depth 30–60 cm | |||
| As | 14.7 a | 15.3 b | 16.3 c |
| Ba | 212 a | 242 a | 348 b |
| Cd | 1.62 a | 1.76 a | 2.85 b |
| Cr | 209 a | 215 a | 412 b |
| Cu | 77.2 a | 80.4 a | 92.8 a |
| Mn | 429 a | 413 a | 456 a |
| Ni | 26.9 a | 29.0 a | 47.5 b |
| Pb | 46.7 a | 55.1 a | 92.6 b |
| Zn | 454 a | 427 a | 482 a |
| “Plant available” ** | Soil depth 0–30 cm | ||
| As | 1.11 a | 1.31 b | 1.51 c |
| Ba | 20.4 a | 19.9 a | 21.2 a |
| Cd | 0.603 a | 0.845 b | 1.14 c |
| Cr | 2.09 a | 2.05 a | 3.68 b |
| Cu | 19.3 a | 25.8 b | 41.3 b |
| Mn | 263 a | 275 a | 267 a |
| Ni | 5.06 a | 6.29 b | 7.98 c |
| Pb | 19.1 a | 29.0 a | 45.1 b |
| Zn | 92.7 a | 178 b | 240 c |
| Soil depth 30–60 cm | |||
| As | 1.50 a | 1.67 b | 1.77 c |
| Ba | 22.2 a | 23.9 b | 25.1 b |
| Cd | 1.11 a | 1.20 a | 1.92 b |
| Cr | 3.04 a | 2.90 a | 5.36 b |
| Cu | 37.8 a | 46.2 ab | 53.1 b |
| Mn | 275 a | 273 a | 287 a |
| Ni | 6.15 a | 7.17 a | 10.6 b |
| Pb | 31.4 a | 36.7 a | 62.1 b |
| Zn | 272 a | 256 b | 290 a |
| 2018 | 2019 | 2020 | |
|---|---|---|---|
| Arundo Line: BFT | |||
| Adaxial epidermis (µm) | 13.88 ± 1.28 a | 13.06 ± 1.41 a | 13.69 ± 1.09 a |
| Abaxial epidermis (µm) | 15.83 ± 1.37 b | 14.50 ± 1.59 a | 14.96 ± 1.42 ab |
| Leaf mesophyll thickness (µm) | 421.29 ± 37.58 c | 335.07 ± 46.71 b | 256.18 ± 110.49 a |
| Vascular bundle width (µm) | 190.46 ± 11.19 c | 166.65 ± 13.63 b | 150.36 ± 61.25 a |
| Vascular bundle height (µm) | 264.53 ± 11.37 c | 232.44 ± 29.19 b | 183.84 ± 77.88 a |
| Vascular bundle area + bundle sheath (µm2) | 72,596.1 ± 6202.48 c | 54,967.57 ± 10,041.21 b | 33,619.79 ± 5461.28 a |
| Bulliform cell number (no.) | 4.2 ± 0 a | 4.35 ± 0.47 ab | 4.6 ± 0.6 b |
| Bulliform cell height (µm) | 109.21 ± 11.52 b | 91.69 ± 11.56 a | 91.29 ± 12.59 ab |
| Bulliform cell width (µm) | 314.22 ± 25.37 c | 268.18 ± 40.30 b | 212.47 ± 15.6 a |
| Adaxial sclerenchyma (µm) | 47.67 ± 4.88 b | 46.30 ± 7.49 b | 38.83 ± 9.0 a |
| Abaxial sclerenchyma (µm) | 42.27 ± 5.64 a | 49.47 ± 9.91 b | 40.29 ± 5.57 a |
| Arundo line: STM | |||
| Adaxial epidermis (µm) | 12.71 ± 1.1 a | 13.64 ± 1.54 a | 13.41 ± 0.87 a |
| Abaxial epidermis (µm) | 16.04 ± 1.96 b | 14.29 ± 0.99 a | 15.14 ± 1.67 ab |
| Leaf mesophyll thickness (µm) | 381.12 ± 24.11 c | 275.95 ± 21.98 b | 255.38 ± 27.5 a |
| Vascular bundle width (µm) | 186.84 ± 9.86 b | 148.04 ± 11.77 a | 146.79 ± 12.8 a |
| Vascular bundle height (µm) | 275.12 ± 18.31 b | 193.12 ± 16.11 a | 183.69 ± 25.03 a |
| Vascular bundle area + bundle sheath (µm2) | 67,349.27 ± 9236.71 b | 39,778.54 ± 5988.5 a | 38,102.77 ± 7324.76 a |
| Bulliform cell number (no.) | 3.8 ± 0.7 a | 4.5 ± 0.5 b | 4.6 ± 0.61 b |
| Bulliform cell height (µm) | 91.01 ± 12.06 a | 84.85 ± 8.63 a | 89.11 ± 10.14 a |
| Bulliform cell width (µm) | 273.71 ± 27.93 c | 231.96 ± 8.63 b | 203.28 ± 28.97 a |
| Adaxial sclerenchyma (µm) | 53.83 ± 13.62 b | 41.47 ± 6.33 a | 36.77 ± 5.57 a |
| Abaxial sclerenchyma (µm) | 42.29 ± 7.82 b | 36.86 ± 4.71 a | 34.41 ± 6.48 a |
| Arundo line: SC Blossom | |||
| Adaxial epidermis (µm) | 14.18 ± 1.04 b | 14.28 ± 1.64 b | 12.12 ± 1.16 a |
| Abaxial epidermis (µm) | 16.15 ± 1.32 a | 15.12 ± 1.75 a | 15.26 ± 1.04 a |
| Leaf mesophyll thickness (µm) | 363.54 ± 22.5 b | 294.59 ± 37.42 b | 299.79 ± 28.83 a |
| Vascular bundle width (µm) | 183.9311.75 ± b | 153.19 ± 12.93 a | 153.38 ± 8.77 a |
| Vascular bundle height (µm) | 246.93 ± 14.76 b | 200.25 ± 20.75 a | 200.38 ± 18.0 a |
| Vascular bundle area + bundle sheath (µm2) | 65,561.40 ± 9536.66 b | 41,548.18 ± 7317.44 a | 43,161.86 ± 4603 a |
| Bulliform cell number (no.) | 4.35 ± 0.57 a | 4.8 ± 0.6 b | 4.5 ± 0.49 ab |
| Bulliform cell height (µm) | 97.01 ± 11.1 b | 82.98 ± 8.31 a | 85.02 ± 12.43 a |
| Bulliform cell width (µm) | 274.18 ± 25.88 b | 222.92 ± 27.73 a | 237.67 ± 14.77 a |
| Adaxial sclerenchyma (µm) | 51.23 ± 7.94 b | 36.71 ± 7.42 a | 37.31 ± 4.34 a |
| Abaxial sclerenchyma (µm) | 39.90 ± 7.7 a | 36.42 ± 5.63 a | 40.97 ± 5.05 a |
| 2018 | 2019 | 2020 | |||
|---|---|---|---|---|---|
| Arundo line: BFT | Adaxial epidermis | stomatal density (no./mm2) | 196.8 ± 20.61 a | 257.6 ± 68.42 b | 310.4 ± 75.53 c |
| width of stomatal complexes (µm) | 7.61 ± 0.76 a | 8.09 ± 2.16 a | 7.7 ± 2.13 a | ||
| length of stomatal complexes (µm) | 31.13 ± 1.89 a | 34.08 ± 8.78 a | 32.84 ± 8.92 a | ||
| number of rows in the intercostal zone (no.) | 4.4 ± 2.42 a | 4.8 ± 2.44 a | 4.8 ± 2.05 a | ||
| number of rows in the costal zone (no.) | 4.6 ± 1.02 a | 5 ± 1.35 a | 5.6 ± 1.55 a | ||
| number of phytoliths (no./mm2) | 206.8 ± 45.49 a | 246.2 ± 67.7 ab | 290 ± 77.87 b | ||
| Abaxial epidermis | stomatal density (no./mm2) | 220.8 ± 20.61 a | 240.8 ± 89.05 a | 338.4 ± 86.51 b | |
| width of stomatal complexes (µm) | 8.51 ± 0.36 a | 8.16 ± 2.09 a | 6.8 ± 2.05 a | ||
| length of stomatal complexes (µm) | 37.89 ± 1.05 a | 36.78 ± 9.24 a | 34.22 ± 9.31 a | ||
| number of rows in the intercostal zone (no.) | 3.2 ± 1.47 a | 5 ± 2.25 a | 5.8 ± 2.64 a | ||
| number of rows in the costal zone (no.) | 5.6 ± 0.8 a | 5.2 ± 1.42 a | 5 ± 1.64 a | ||
| number of phytoliths (no./mm2) | 196.8 ± 35.57 a | 256.4 ± 79.13 a | 256.4 ± 71.78 a | ||
| Arundo line: STM | Adaxial epidermis | stomatal density (no./mm2) | 234.4 ± 19.03 a | 255.2 ± 8.19 a | 227.2 ± 20.77 a |
| width of stomatal complexes (µm) | 7 ± 0.47 a | 7.75 ± 0.5 a | 8 ± 0.54 a | ||
| length of stomatal complexes (µm) | 31.06 ± 1.34 a | 32.47 ± 0.81 a | 33.9 ± 0.99 a | ||
| number of rows in the intercostal zone (no.) | 5.8 ± 2.64 a | 6 ± 2.76 a | 3.4 ± 1.02 a | ||
| number of rows in the costal zone (no.) | 4.8 ± 0.4 a | 5 ± 0.89 a | 6 ± a.63 a | ||
| number of phytoliths (no./mm2) | 186.6 ± 53.05 a | 246.6 ± 24.41 a | 289.6 ± 69.53 a | ||
| Abaxial epidermis | stomatal density (no./mm2) | 267.2 ± 12.49 a | 314.4 ± 13.76 b | 199.2 ± 14.4 c | |
| width of stomatal complexes (µm) | 6.46 ± 0.32 a | 7.24 ± 0.73 a | 8.23 ± 0.74 a | ||
| length of stomatal complexes (µm) | 33.14 ± 1.8 a | 33.48 ± 1.01 a | 34.04 ± 1.71 a | ||
| number of rows in the intercostal zone (no.) | 5 ± 3.29 a | 5.4 ± 2.33 a | 4.8 ± 0.93 a | ||
| number of rows in the costal zone (no.) | 4.4 ± 0.49 a | 4.8 ± 0.98 a | 5.4 ± 0.49 a | ||
| number of phytoliths (no./mm2) | 229.8 ± 48.59 a | 313.4 ± 35.59 a | 219.8 ± 38.37 a | ||
| Arundo line: SC Blossom | Adaxial epidermis | stomatal density (no./mm2) | 178.4 ± 24.99 a | 246.4 ± 69.91 bc | 230.4 ± 59.50 b |
| width of stomatal complexes (µm) | 6.68 ± 0.30 a | 5.52 ± 2.09 a | 8.03 ± 2.14 a | ||
| length of stomatal complexes (µm) | 32.27 ± 1.07 a | 32.12 ± 8.75 a | 33.60 ± 9.11 a | ||
| number of rows in the intercostal zone no.) | 5 ± 1.67 a | 6.8 ± 2.59 a | 5.8 ± 2.06 a | ||
| number of rows in the costal zone (no.) | 4 ± 1.26 a | 4.2 ± 1.33 a | 4.6 ± 1.56 a | ||
| number of phytoliths (no./mm2) | 200.2 ± 60.6 a | 206.6 ± 67.86 a | 266.4 ± 77.09 a | ||
| Abaxial epidermis | stomatal density (no./mm2) | 292 ± 10.43 a | 309.6 ± 84.28 ab | 343.2 ± 99.97 b | |
| width of stomatal complexes (µm) | 7.08 ± 0.98 a | 6.19 ± 1.84 a | 7.17 ± 2.06 a | ||
| length of stomatal complexes (µm) | 31.2 ± 0.92 a | 31.39 ± 8.86 a | 36.92 ± 9.5 a | ||
| number of rows in the intercostal zone (no.) | 3.8 ± 1.46 a | 4.8 ± 2.46 a | 4 ± 2.31 a | ||
| number of rows in the costal zone (no.) | 5.2 ± 0.4 a | 5.2 ± 1.37 a | 6.2 ± 1.61 a | ||
| number of phytoliths (no./mm2) | 226.4 ± 58.83 a | 259.8 ± 81.82 a | 249.8 ± 69.84 a | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tóth, C.; Simon, L.; Tóth, B. Microanatomical Changes in the Leaves of Arundo donax (L.) Caused by Potentially Toxic Elements from Municipal Sewage Sediment. Plants 2024, 13, 740. https://doi.org/10.3390/plants13050740
Tóth C, Simon L, Tóth B. Microanatomical Changes in the Leaves of Arundo donax (L.) Caused by Potentially Toxic Elements from Municipal Sewage Sediment. Plants. 2024; 13(5):740. https://doi.org/10.3390/plants13050740
Chicago/Turabian StyleTóth, Csilla, László Simon, and Brigitta Tóth. 2024. "Microanatomical Changes in the Leaves of Arundo donax (L.) Caused by Potentially Toxic Elements from Municipal Sewage Sediment" Plants 13, no. 5: 740. https://doi.org/10.3390/plants13050740
APA StyleTóth, C., Simon, L., & Tóth, B. (2024). Microanatomical Changes in the Leaves of Arundo donax (L.) Caused by Potentially Toxic Elements from Municipal Sewage Sediment. Plants, 13(5), 740. https://doi.org/10.3390/plants13050740