
Paeonia × suffruticosa (Moutan Peony)—A Review of the Chemical Composition, Traditional and Professional Use in Medicine, Position in Cosmetics Industries, and Biotechnological Studies



Abstract
:1. Paeonia Genus and Paeonia lactiflora and Paeonia veitchii as Known Medicinal Plants—General Characteristic
2. Paeonia × suffruticosa as a New Pharmacopoeial Plant Species
3. Paeonia × suffruticosa—General Characteristics
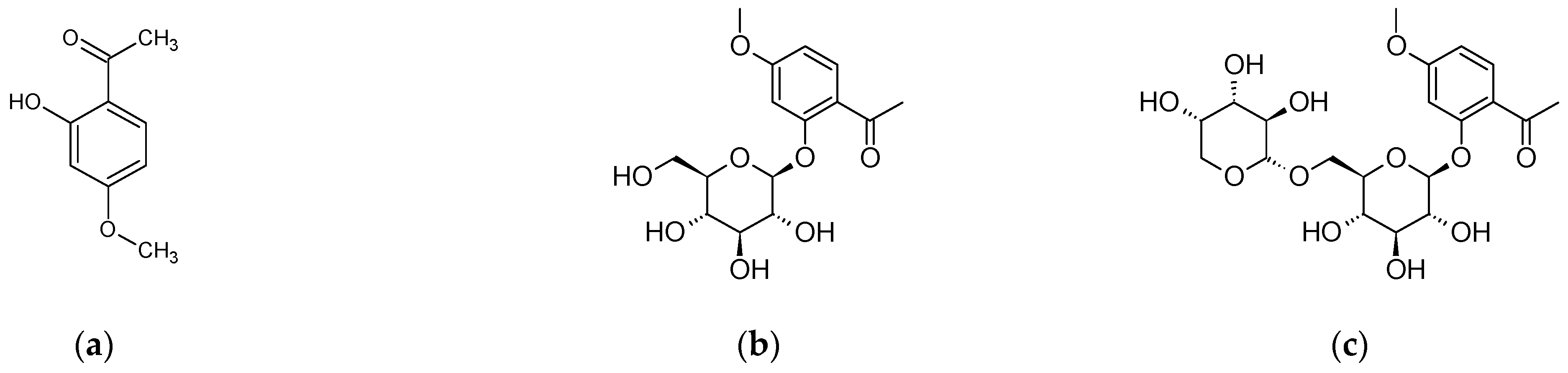
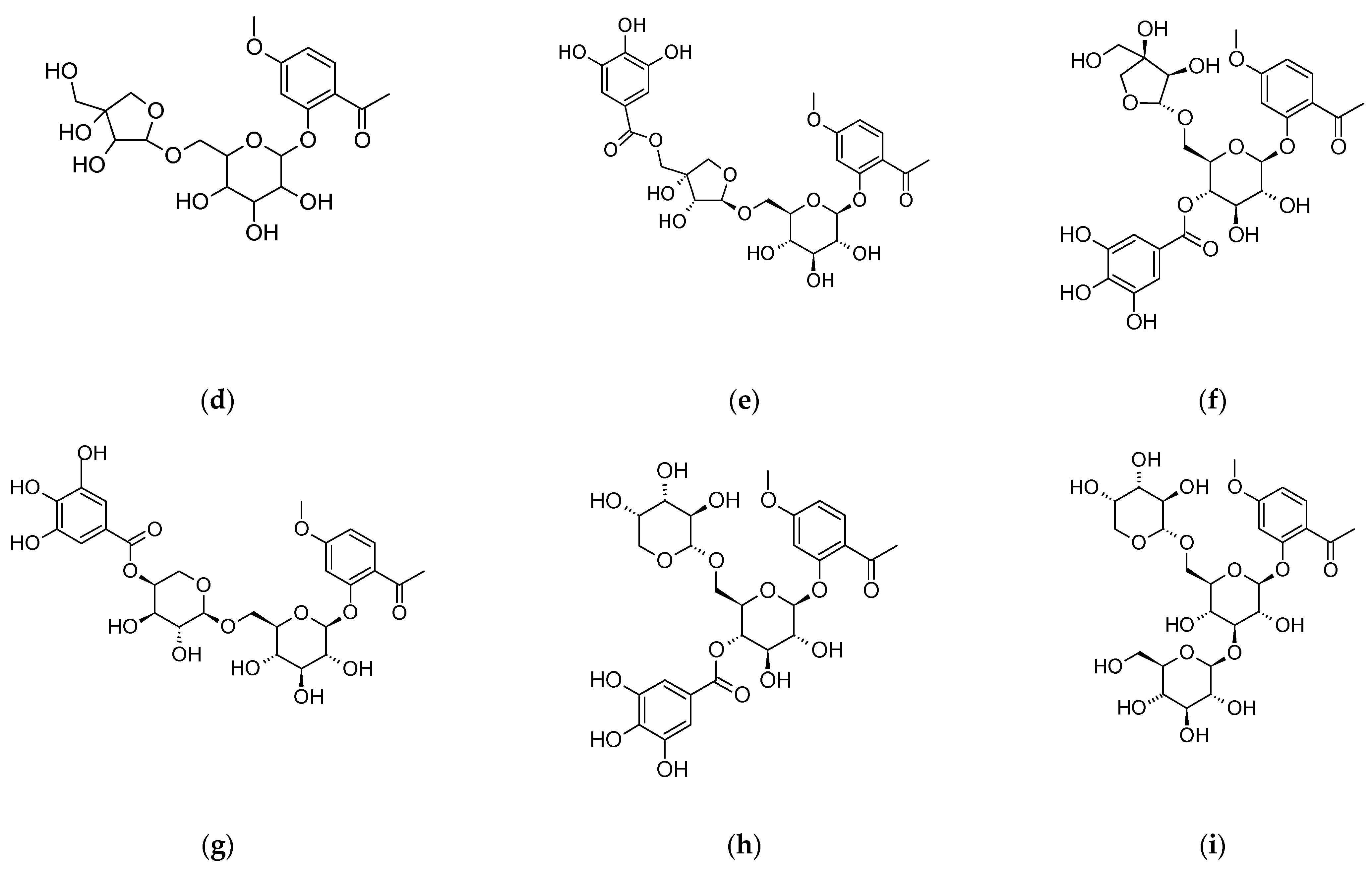
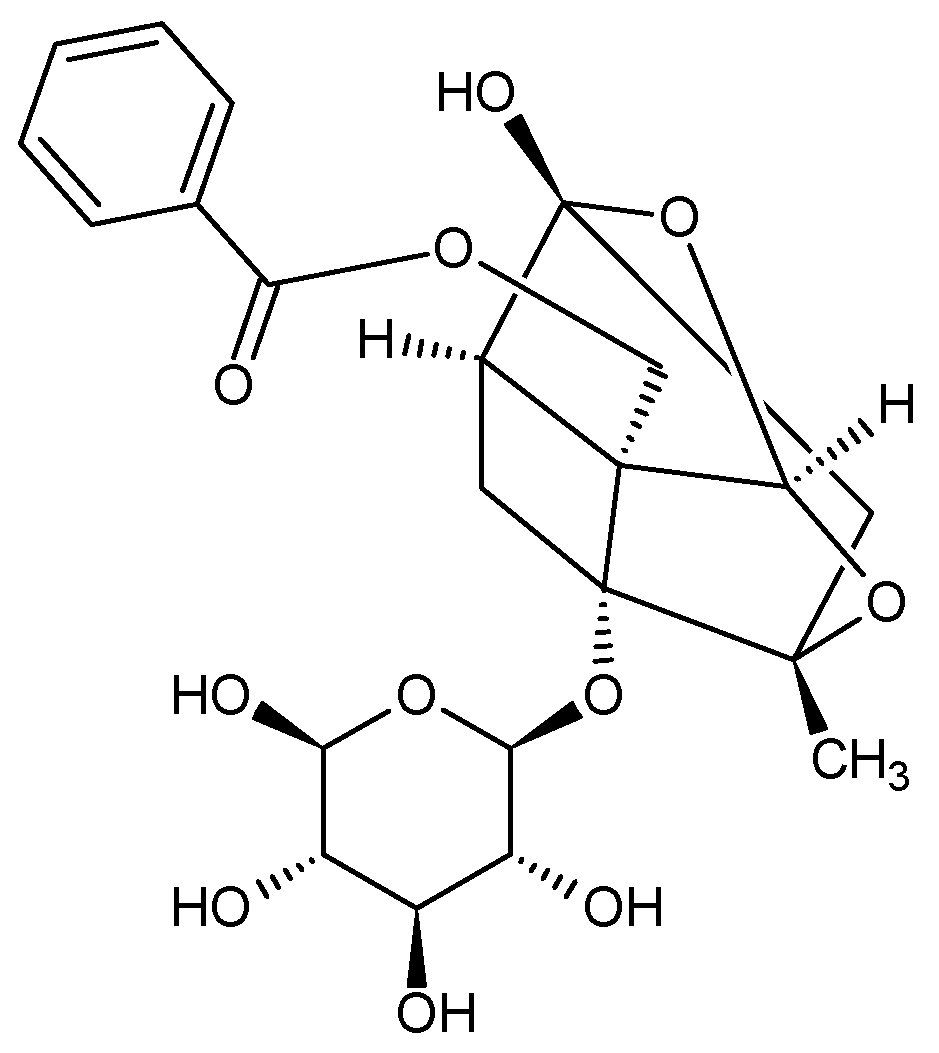
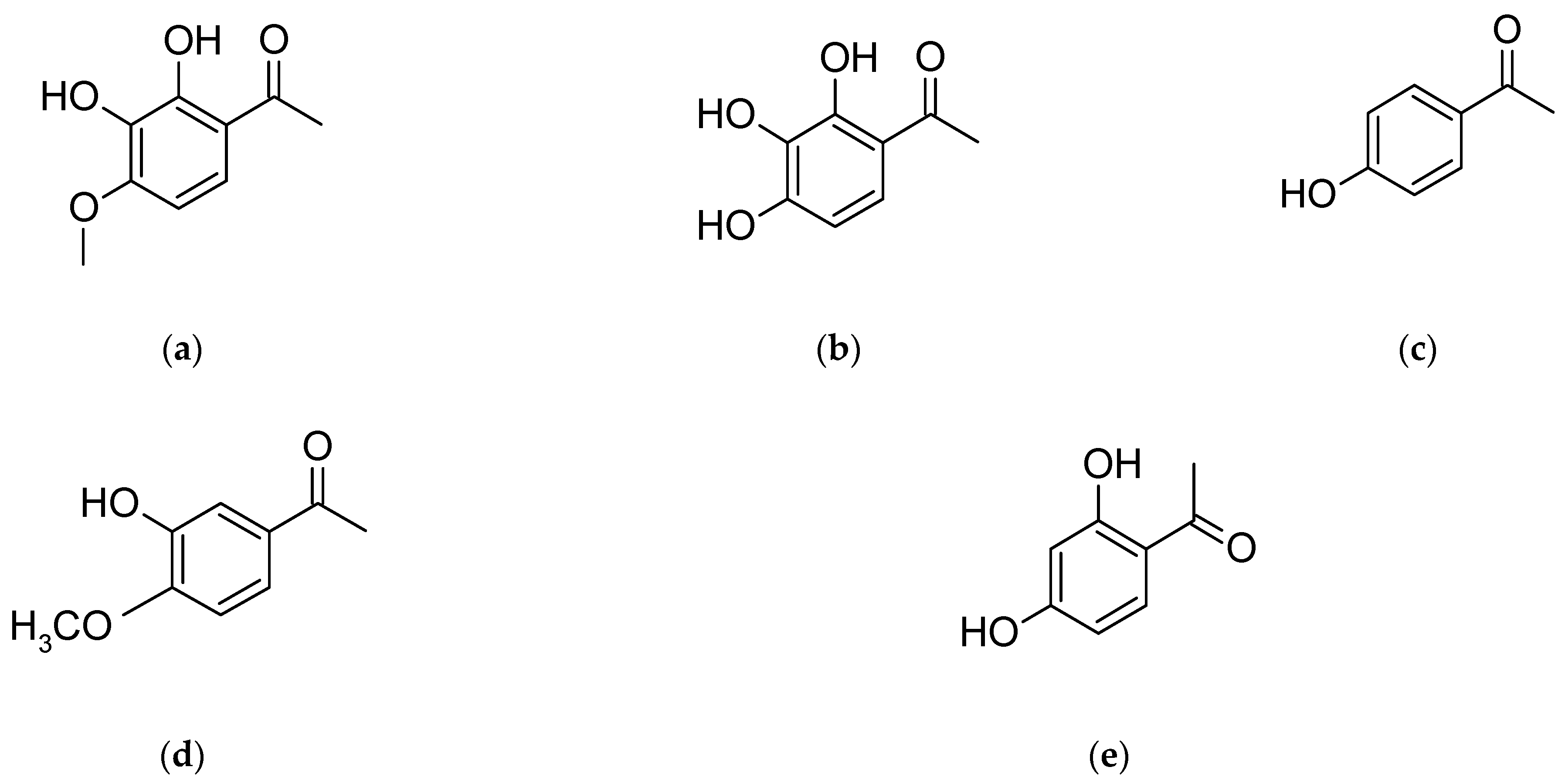
4. Paeonia × suffruticosa—Phytochemical Characteristics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group of Compounds | Compounds | References |
|---|---|---|
| Terpenoid compounds | Monoterpenenoids deoxypaeonisuffrone, isopaeonisuffral, 6-methoxy-paeoniflorigenone, 3-O–methylpaeonisuffral, paeonifloringenone, paeoniflorin A, paeonisothujone, paeonisuffral, paeonisuffrone, (-)-paeonisuffrone Monoterpenenoid glycosides: benzoylpaeoniflorin, α-benzoyloxypaeoniflorin, β-benzoyloxypaeoniflorin, 8-O-benzoylpaeonidanin, 9-O-butyloxypaeonidanin, 4-O-butyloxypaeoniflorin, 9-O-butylpaeonidanin, 4-O-butylpaeoniflorin, 8-O-debenzoylpaeoniflorin, galloyloxypaeoniflorin, galloylpaeoniflorin, p-hydroxylbenzoylpaeonidanin, 4-methylbenzoyloxypaeoniflorin, 4-methylgalloyloxypaeoniflorin, 4-methylmoudanpioside C, 4-methyloxypaeoniflorin, 4-methylsuffrupaeoniflorin B, 4-O-methylpaeoniflorin, 4-O-methylbenzoylpaeoniflorin, mudanpioside A-J, oxypaeonidanin, 9-epi- oxypaeonidanin, oxypaeoniflorin sulfonate, paeonidanin A and C, paeoniflorin, paeoniflorin B, paeoniflorin-4-ethyl ether, paeoniside A and B, suffrupaeonidanin A-F, suffrupaeoniflorin A and B, suffruyabiosides A and B, 6-O-vanillyoxypaeoniflorin Triterpenoids: betulinic acid, mudanpinoic acid A, oleanolic acid, palbinone, ursolic acid | [6,27,28,29,30,31,32] |
| Triterpenoid saponins | hederagenin, 30-norhederagenin | [6] |
| Acetophenons | 2,3-dihydroxy-4-methoxyacetophenone, gallacetophenone, p-hydroxyacetophenone, 3-hydroxy-4-methoxyacetophenone, reacetophenone | [33] |
| Phenols and their derivatives | apiopaeonoside, galusan metylu, iriflophenone 2-O-β-D-glucopyranoside, methyl 3-hydroxy-4-methoxybenzoate, mudanoside C, paeonol, paeonolide, paeoniside, phenol, suffruticosides A–E | [6,33] |
| Flavonoids | apigenin 7-O-rhamnoside, isorhamnetin 3,7-di-O-glucoside, quercetin, kaempferol, kaempferol 3,7-di-O-glucoside, quercetin 3-O-galloylglucoside | [6,32] |
| Phenolic and cyclohexanecarboxylic acids | gallic acid, galloylquinic acid, trans-caffeic acid stearyl ester, quinic acid, p-hydroxybenzoic acid, 3-hydroxy-4-methoxybenzoic acid | [32] |
| Resveratrol oligomers | suffruticosol A, suffruticosol B, suffruticosol C, trans-gnetin H, trans-suffruticosol D, trans-ε-viniferin | [63] |
| Catechins | catechin, (+)-catechin-7-O-glucopyranoside, dimeric proanthocyanidin epicatechin-(4β-8)-catechin, epicatechin-3-O-gallate, flavan-3-ols catechin, epigallocatechin gallate | [104] |
| Sterols | campesterol, daucosterol, β-sitosterol | [6,31] |
| Polysaccharides | polisacharyd-2b | [34] |
| Others | adenosine, ainsliaside E, 1,2,3,4,6-penta-O-galloyl-β-D-glucopyranose, paesuffrioside, trigalloilo-glucose, thymidine, 1-tryptophan, uridine | [30] |
5. Paeonia × suffruticosa-Studies on Biological Activity
5.1. Antioxidant Effect
5.2. Cytoprotective Effect
5.3. Anti-Inflammatory Effect
5.4. Anticancer Effect
5.5. Cardioprotective and Anti-Atherosclerotic Effects
5.6. Antidiabetic Effect
5.7. Neuroprotective Effect
5.8. Effects in Neurodegenerative Diseases
5.9. Hepatoprotective Effect
5.10. Anti-Allergic Effect
5.11. Immunomodulatory Activity
5.12. Antibacterial and Antifungal Effects
| Activity | Mechanism of Action | References |
|---|---|---|
| Antioxidant activity | Reduction in the production of reactive oxygen species (ROS) (ethanolic extract of Moutan cortex) | [45] |
| Activation of the signaling of the transcription factor Nrf2, that controls the expression of genes encoding enzymes and cytoprotective proteins (paeonol) | [56,64] | |
| Cytoprotective activity | Protection of thymocytes (“pre-T lymphocytes”) against oxidative damage caused by 60Co radiation (paeoniflorin) | [38] |
| Protection of human cell lines (EA.hy926) against gamma-induced oxidative damage via the transcription factor pathway protection Nrf2/HO (paeoniflorin) | [39] | |
| Protection of the retinal pigment epithelium cells against oxidative stress, reduction in the production of ROS and inhibition of the activation of the caspase-3 pathway (paeoniflorin) | [40] | |
| Protection against damage and cell death caused by hydrogen peroxide in human HaCaT keratinocytes (galloilopaeoniflorin) | [41] | |
| Anti-inflammatory activity | Inhibition of the expression of nitric oxide synthase (iNOS) and induced cyclooxygenase (COX-2) by suppressing the phosphorylation of the inhibitory protein (I-κBα), the transcription factor NF-κB (extract of Moutan cortex) | [20] |
| Inhibition of the HMGB1-RAGE/TLR-2/TLR-4-NF-κB pathway (paeoniflorin) | [66] | |
| Inhibition of the synthesis of tumor necrosis factor (TNF-α) and the production of interleukin-6 (IL-6) in synoviocytes (cells of the synovial membrane of the joint capsule) (paeonol, paeoniflorin, 1,2,3,4,6-penta-O-galloyl- β-D-glucopyranose) | [67] | |
| Inhibition of the secretion of interleukin-8 (IL-8) and monocyte chemotactic protein in human monocytic cells (U937) (methanol extract from shoulder) regulating the production of TNF-α and interleukins-IL-1β, IL-6 and IL-10 by inactivating I-κBα (NF-κB inhibitory protein), extracellular signal regulated kinase (ERK1/2), N-kinases c-JUN terminal (JNK) and mitogen-activated protein kinases (p38 MAPK) (paeonol) | [69] | |
| Inhibition of the total expression of HMGB1, NF-κB P65 and TNF-α in the lung tissue of acute lung injury rats (paeonol) | [35] | |
| Suppressing the activity of M1 macrophage cells (pro-inflammatory) and increasing the function of M2 macrophage cells (anti-inflammatory) (paeoniflorin) | [36] | |
| Inhibition expressions of Il-6, TNF-α, NADPH oxidase 2 (NOX2), prostaglandin-endoperoxide synthase 2 (PTGS2), nucleobindin-2 (NUCB2)/nesfatin-1, intercellular adhesion molecule 1 (ICAM -1), vascular cell adhesion molecule 1 (VCAM-1), matrix metalloproteinase-3/13 (MMP-3/13) and degradation of type II collagen (paeonol) | [37] | |
| Anticancer activity | Reduction resistance to paclitaxel in human breast cancer cells by regulating the expression of transgelin 2 (paeonol) | [42] |
| Induction of an anti-tumor effect on colon cancer cells by suppressing prostaglandin synthesis (PGE-2) and expression of COX-2 (paeonol) | [43] | |
| Inhibition of the metastasis of melanoma and chondrosarcoma (paeonol) | [44] | |
| Induction of apoptosis of EMT6 breast cancer cells, HepG2 human liver cancer cells and HepA cells in a mouse model (paeonol) | [46,48] | |
| Inhibition of cell proliferation and induction of apoptosis on mouse gastric cancer cell lines (MFC) and on human gastric tumor cells SGC-7901 (paeonol) | [49] | |
| Decreases the expression of proteins regulating the release of cytochrome c from mitochondria (Bcl-2) and increased the expression of the apoptosis accelerating protein (Bax) in MFC and SGC-7901 (paeonol) | [49] | |
| Cells’ suppression of the signaling pathway for the gene encoding the single-pass Notch-1 transmembrane receptor in breast cancer cells (paeoniflorin) | [50] | |
| Inhibition of macrophage-dependent metastasis of lung cancer (paeoniflorin) | [50] | |
| Inhibition of proliferation and induction of apoptosis of human glioblastoma cells by up-regulation of microRNA-16 and down-regulation of metalloproteinase-9 (paeoniflorin) | [51] | |
| Reduction in proliferation on human SK-HEP-1 hepatocellular carcinoma cells (1,2,3,4,6-penta-O-galloyl-β-D-glucopyranose) | [52] | |
| Inhibition of the growth of bladder cancer cells (extract of Moutan cortex) | [53] | |
| Induction stress on endoplasmic reticulum (ER)-related proteostasis and affected mitochondrial membrane potential to increase autophagosome numbers and block their degradation (aqueous extracts of Moutan cortex) | [54] | |
| Inhibition of the proliferation of human liver cancer cell line (HepG2) and human hepatocarcinoma cell line (SMMC-7721) and induction apoptosis, concomitant with the down-regulation of survivin (paeonol) | [55] | |
| Cardioprotective and anti-atherosclerotic activity | Inhibition of thrombocyte aggregation and blood coagulation (paeonol, paeoniflorin, benzoylpaeoniflorin and benzoyloxypaeoniflorin) | [21] |
| Protective effect in the rat model of ischemia and reperfusion (extract of Moutan cortex) | [78] | |
| Inhibition of inflammatory processes and signaling pathways of iNOS (paeoniflorin) | [79] | |
| Reduction in vascular damage and the expression of E-selectin and intercellular adhesion molecule (ICAM-1) in a mouse model of the skin Arthus reaction (paeoniflorin) | [80] | |
| Increases thrombus recanalization by inducing endothelial growth factor-165 and up-regulating urokinase plasminogen activator (paeonol and paeoniflorin) | [81,82] | |
| Inhibition of the adhesion of monocytes, induced by the oxidized form of LDL, to the vascular endothelium by inhibiting the mitogen-activated kinase (MAPK) signaling pathway (paeonol) | [83] | |
| Prevention arterial thrombosis (paeoniflorin) | [84] | |
| Antidiabetic activity | Inhibition of glucose uptake by intestinal brush border membrane vesicles (BBMV) and increased glucose uptake in human skin fibroblasts (Hs68) and Mouse adipocytes (3T3-L1) (extract of Moutan cortex) | [85] |
| improved glucose tolerance (paeonol) | [85] | |
| Decrease in receptor expression for glycation products and NF-κB in the hippocampus and cortical neurons of the brain (paeonol) | [70] | |
| Increases the content of glutathione and noticeably reduces the activity of iNOS in the tissue of the hippocampus (paeonol) | [71] | |
| Delays the onset and alleviates the degree of lens opacities in diabetic cataracts (polysaccharide-2b present Moutan cortex) | [34] | |
| Stimulation of human insulin-resistant HepG2 cells glucose uptake and glycogen synthesis via the AMPK pathway (palbinon and some triterpenoids isolated from Moutan cortex) | [72] | |
| Neuroprotective activity | Relieving neuropathic pain (Moutan cortex extract) | [19] |
| Protective effect on neurons in a mouse model of Parkinson’s disease (Moutan cortex extract) | [57] | |
| Reduction in cell damage and extending cell viability by inhibiting the ability to bind the N-methyl-D-aspartate (NMDA) receptor and reducing the intracellular concentration of calcium ions (paeonol) | [59] | |
| Suppression of neuroinflammatory reactions by suppressing the NF-κB pathway and mitogen-activated kinases (MAPK) (paeonol) | [59] | |
| Inhibition of the release of NO and the expression of iNOS and COX-2 in microglia cells (paeonol) | [60] | |
| Regulation of AMPK-α kinase and glycogen synthase 3α/β kinase (GSK 3α/β) (paeonol) | [60] | |
| Protection of nerve cells against oxidative stress by inducing the expression of the hemoxygenase-1 gene (1,2,3,4,6-penta-O-galloyl-β-D-glucopyranose) | [62] | |
| Improves the cognitive performance of scopolamine-treated mice in behavioral tests (seed coat extracts of P. × suffruticosa) | [63] | |
| Increases acetylcholinesterase, choline acetyltransferase, superoxide dismutase (SOD) and catalase (CAT) activities and acetylcholine, glutathione (GSH) and IL-4 levels, and decreases IL-1β, IL-6 and TNF-α levels in a cell line that was derived from a transplantable rat pheochromocytoma (seed coat extracts of P. × suffruticosa) | [63] | |
| Activity in neurodegenerative diseases | Increasing the level of acetylcholine and GSH (paeonol) | [22] |
| Restoration of superoxide dismutase activity and concentration of Na +/K+-ATPase (paeonol) | [110] | |
| Reduction in MDA levels and cholinesterase activity (paeonol) | [110] | |
| Alleviated neuronal damage, both in the hippocampus and in the temporal cortex (paeonol) | [110] | |
| Inhibition of formation and destabilization of initial formation of amyloid-β fibrils in in vitro and in vivo models (1,2,3,4,6-penta-O-galloyl-β-D-glucopyranose) | [111] | |
| Hepatoprotective activity | Reduction in GSH deficit, cytochrome P450 2E1 activity and protection against damage to hepatic DNA (extract of Moutan cortex) | [73,74,75,76,77] |
| Decrease in serum transaminase levels, damage to liver cells, steatosis and infiltration of inflammatory cells (paeonol) | [74] | |
| Reduction in hepatic mRNA expression of lipogenic genes (paeonol) | [74] | |
| Lowering the level of inflammatory cytokines in serum and tissues, peroxidation of tissue lipids, neutrophil infiltration and inhibition of hepatocyte apoptosis (paeonol) | [75] | |
| Alleviation of liver fibrosis by inhibiting the hypoxia-1α-induced factor, partly by the m-TOR dependent pathway (paeoniflorin) | [76,77] | |
| Anti-allergic effect | Inhibition of systemic anaphylactic shock (ethanol extract of Moutan cortex) | [112] |
| Inhibition of the scratch reflex (ethanol extract of Moutan cortex) | [112] | |
| Inhibition of increased vascular permeability (ethanol extract of Moutan cortex) | [112] | |
| Reduction in histamine release from mast cells (ethanol extract of Moutan cortex) | [113] | |
| Immunomodulatory activity | Stimulation of Raw264.7 (macrophage cell line) cell proliferation (polysaccharides obtained from P. × suffruticosa flowers) | [114] |
| Activation of Raw264.7 cells by releasing immunoactive molecules such as NO, TNF-α and IL-6 (polysaccharides obtained from P. × suffruticosa flowers) | [114] | |
| Antibacterial and antifungal activity | Inhibition of growth of Gram-positive bacteria: Staphylococcus aureus and Gram-negative bacteria: Escherichia coli (methanol extract of Moutan cortex) | [32,116] |
| Inhibition of growth of Candida glabrata (1,2,3,4,6-penta-O-galloyl-β-D-glucopyranose) | [116] |
6. Applications in Cosmetology
7. Paeonia × suffruticosa—Toxicity
8. Paeonia × suffruticosa—Pant Biotechnological Studies
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Hong, D. Peonies of the World. Taxonomy and Phytogeography; Hong, D., Ed.; Royal Botanic Gardens: Richmond, UK, 2010. [Google Scholar]
- World Flora Online. Available online: http://www.worldfloraonline.org/ (accessed on 9 August 2022).
- Zhao, M.; Wu, S.P. A review of the ethnobotany, phytochemistry and pharmacology of tree peony (Sect. Moutan). S. Afr. J. Bot. 2019, 124, 556–563. [Google Scholar] [CrossRef]
- Connor, P. Paeon. In Gods, Goddesses, and Mythology; Littleton, C., Ed.; Marshall Cavendish: Tarrytown, NY, USA, 2005; p. 1069. [Google Scholar]
- Sang, T.; Crawford, D.J.; Stuessy, T.F. Documentation of reticulate evolution in peonies (Paeonia) using internal transcripted spacer sequences of nuclear ribosomal DNA: Implications for biogeography and concerted evolution. Proc. Natl. Acad. Sci. USA 1995, 92, 6813–6817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.; Peng, B.; Dan, Y.; Peng, Y.; Xiao, P. Chemical taxonomy of tree peony species from China based on root cortex metabolic fingerprinting. Phytochemistry 2014, 107, 69–79. [Google Scholar] [CrossRef] [PubMed]
- European Directorate for the Quality of Medicine & HealthCare. European Pharmacopoeia 10.0; Council of Europe: Strasbourg, France, 2021. [Google Scholar]
- Lan, B.; Li, J.; Duan, Q. An Encyclopedia of the Tree Peonies in China; China Science and Technology Press: Beijing, China, 2002. [Google Scholar]
- HMPC European Medicines Agency; The Committee on Herbal Medicinal Products (HMPC). Public Statement on the Use of Herbal Medicinal Products Containing Estragole. Available online: https://www.ema.europa.eu/en/documents/other/second-draft-revision-1-public-statement-use-herbal-medicinal-products-containing-estragole_en.pdf (accessed on 9 August 2022).
- Institute for Traditional Medicine. Available online: http://www.itmonline.org/arts/peony.html (accessed on 9 August 2022).
- European Directorate for the Quality of Medicines & HealthCare. European Pharmacopoeia 9.0; Council of Europe: Strasbourg, France, 2017. [Google Scholar]
- Zhang, X.; Wang, Y.; Liang, Q.; Ma, Z.; Xiao, C.; Tan, H.; Gao, Y. The correlation between chemical composition, as determined by UPLC-TOF-MS, and acute toxicity of Veratrum nigrum L. and Radix Paeoniae alba. Evid.-Based Complement. Altern. Med. 2014, 2014, 892797. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.L.; Wang, J.X.; Hu, X.X.; Chen, L.; Qiu, Z.K.; Zhao, N.; Yu, Z.D.; Sun, S.Z.; Xu, Y.Y.; Guo, Y.; et al. Antidepressant-like effects of albiflorin extracted from Radix Paeoniae alba. J. Ethnopharmacol. 2016, 179, 9–15. [Google Scholar] [CrossRef]
- Tan, Y.Q.; Chen, H.W.; Li, J.; Wu, Q.J. Efficacy, Chemical Constituents, and Pharmacological Actions of Radix Paeoniae rubra and Radix Paeoniae alba. Front. Pharmacol. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Kataloh Sortiv Roslyn, Prydatnykh dlia Poshyrennia v Ukraini na 2018 rik (Vytiah); Ukraine, 2018.
- Markovskyi, Y. Dekoratyvnue mnoholetnyky. SPb. Myr Y Semia 2002. [Google Scholar]
- Pantsyreva, H. Research of sortal resources of grape species of Paeonia L. in Ukraine. Sci. Bull. UNFU 2018, 28, 74–78. [Google Scholar] [CrossRef]
- Qiu, Z.K.; He, J.L.; Liu, X.; Zeng, J.; Chen, J.S.; Nie, H. Anti-PTSD-like effects of albiflorin extracted from Radix Paeoniae alba. J. Ethnopharmacol. 2017, 198, 324–330. [Google Scholar] [CrossRef]
- Tatsumi, S.; Mabuchi, T.; Abe, T.; Xu, L.; Minami, T.; Ito, S. Analgesic effect of extracts of Chinese medicinal herbs Moutan cortex and Coicis semen on neuropathic pain in mice. Neurosci. Lett. 2004, 370, 130–134. [Google Scholar] [CrossRef]
- Chun, S.C.; Jee, S.Y.; Lee, S.G.; Park, S.J.; Lee, J.R.; Kim, S.C. Anti-inflammatory activity of the methanol extract of Moutan cortex in LPS-activated Raw264.7 cells. Evid.-Based Complement. Altern. Med. 2007, 4, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Koo, Y.K.; Kim, J.M.; Koo, J.Y.; Kang, S.S.; Bae, K.H.; Kim, Y.S.; Chung, J.H.; Yun-Choi, H.S. Platelet anti-aggregatory and blood anti-coagulant effects of compounds isolated from Paeonia lactiflora and Paeonia suffruticosa. Pharmazie 2010, 65, 624–628. [Google Scholar] [PubMed]
- Zhou, J.; Zhou, L.; Hou, D.; Tang, J.; Sun, J.; Bondy, S.C. Paeonol increases levels of cortical cytochrome oxidase and vascular actin and improves behavior in a rat model of Alzheimer’s disease. Brain Res. 2011, 1388, 141–147. [Google Scholar] [CrossRef] [Green Version]
- European Directorate for the Quality of Medicines & HelthCare. Supplement 9.4. In European Pharmacopoeia 9.0; Council of Europe: Strasbourg, France, 2018; p. 2474. [Google Scholar]
- The Ministry of Health Labour and Welfare. The Japanese Pharmacopoeia, 18th ed.; Pharmaceutical s and Medical Devices Agency: Tokyo, Japan, 2021. [Google Scholar]
- Ministry of Food and Drug Safety. Korean Pharmacopoeia, 10th ed.; Ministry of Food and Drug Safety: Cheongju-si, Republic of Korea, 2012. [Google Scholar]
- Wagner, H. Cortex Moutan—Mudanpi. In Chromatographic Fingerprint Analysis of Herbal Medicines Thin-Layer and High Performance Liquid Chromatography of Chinese Drugs; Wagner, H., Bauer, R., Melchart, D., Xiao, P.-G., Staudinger, A., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 91–103. [Google Scholar]
- Yoshikawa, M.; Harada, E.; Minematsu, T.; Muraoka, O.; Yamahara, J.; Murakami, N.; Kitagawa, I. Absolute stereostructures of paeonisothujone, a novel skeletal monoterpene ketone, and deoxypaeonisuffrone, and isopaeonisuffral, two new monoterpenes, from Moutan cortex. Chem. Pharm. Bull. 1994, 42, 736–738. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Ohta, T.; Kawaguchi, A.; Matsuda, H. Bioactive constituents of chinese natural medicines. V. Radical scavenging effect of Moutan cortex. (1): Absolute stereostructures of two monoterpenes, paeonisuffrone and paeonisuffral. Chem. Pharm. Bull. 2000, 48, 1327–1331. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Zhao, F.; Chen, L.; Jiang, Z.; Liu, Y.; Li, Z.; Qiu, F.; Yao, X. New monoterpene glycosides from Paeonia suffruticosa Andrews and their inhibition on NO production in LPS-induced RAW 264.7 cells. Bioorgan. Med. Chem. Lett. 2012, 22, 7243–7247. [Google Scholar] [CrossRef] [PubMed]
- Xiao, K.; Song, Q.H.; Zhang, S.W.; Xuan, L.J. A pyrrole derivative from Paeonia suffruticosa. Nat. Prod. Res. 2008, 22, 1614–1619. [Google Scholar] [CrossRef]
- He, C.N.; Peng, Y.; Zhang, Y.C.; Xu, L.J.; Gu, J.; Xiao, P.G. Phytochemical and biological studies of Paeoniaceae. Chem. Biodivers. 2010, 91, 52–80. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, Y.; Zong, H.; Lu, X.; Shen, W.; Zhuge, B. Chemical constituents, antibacterial activity and mechanism of Paeonia suffruticosa Andr. buds extract against Staphylococcus aureus and Escherichia coli O157:H7. Nat. Prod. Res. 2021, 35, 1005–1009. [Google Scholar] [CrossRef]
- Ding, L.; Zuo, Q.; Li, D.; Feng, X.; Gao, X.; Zhao, F.; Qiu, F. A new phenone from the roots of Paeonia suffruticosa Andrews. Nat. Prod. Res. 2017, 31, 253–260. [Google Scholar] [CrossRef]
- Zhao, G.H.; Shen, Y.S.; Ma, J.B.; Li, F.; Shi, X.Q. Protection of polysaccharides-2b from mudan cortex of Paeonia suffruticosa Andr on diabetic cataract in rats. Zhongguo Zhong Yao Za Zhi 2007, 32, 2036–2039. [Google Scholar] [PubMed]
- Liu, X.; Xu, Q.; Mei, L.; Lei, H.; Wen, Q.; Miao, J.; Huang, H.; Chen, D.; Du, S.; Zhang, S.; et al. Paeonol attenuates acute lung injury by inhibiting HMGB1 in lipopolysaccharide-induced shock rats. Int. Immunopharmacol. 2018, 61, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Zhai, T.; Sun, Y.; Li, H.; Zhang, J.; Huo, R.; Li, H.; Shen, B.; Li, N. Unique immunomodulatory effect of paeoniflorin on type I and II macrophages activities. J. Pharmacol. Sci. 2016, 130, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Xu, X.; Kang, Z.; Zhang, Z.; Li, Y. Paeonol prevents IL-1β-induced inflammatory response and degradation of type II collagen in human primary chondrocytes. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2139–2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.R.; Zhou, Z.; Zhu, D.; Sun, Y.N.; Dai, J.M.; Wang, S.Q. Protective effect of paeoniflorin on irradiation-induced cell damage involved in modulation of reactive oxygen species and the mitogen-activated protein kinases. Int. J. Biochem. Cell Biol. 2007, 39, 426–438. [Google Scholar] [CrossRef]
- Yu, J.; Zhu, X.; Qi, X.; Che, J.; Cao, B. Paeoniflorin protects human EA.hy926 endothelial cells against gamma-radiation induced oxidative injury by activating the NF-E2-related factor 2/heme oxygenase-1 pathway. Toxicol. Lett. 2013, 218, 224–234. [Google Scholar] [CrossRef]
- Wankun, X.; Wenzhen, Y.; Min, Z.; Weiyan, Z.; Huan, C.; Wei, D.; Lvzhen, H.; Xu, Y.; Xiaoxin, L. Protective effect of paeoniflorin against oxidative stress in human retinal pigment epithelium in vitro. Mol. Vis. 2011, 17, 3512–3522. [Google Scholar]
- Weng, Y.C.; Jing, P.M.; Cheon, K.K.; Won, C.J.; Won, H.J. 6’-O-galloylpaeoniflorin protects human keratinocytes against oxidative stress-induced cell damage. Biomol. Ther. 2013, 21, 349–357. [Google Scholar]
- Cai, J.; Chen, S.; Zhang, W.; Hu, S.; Lu, J.; Xing, J.; Dong, Y. Paeonol reverses paclitaxel resistance in human breast cancer cells by regulating the expression of transgelin 2. Phytomedicine 2014, 21, 984–991. [Google Scholar] [CrossRef]
- Zhang, L.; Tao, L.; Shi, T.; Zhang, F.; Sheng, X.; Cao, Y.; Zheng, S.; Wang, A.; Qian, W.; Jiang, L.; et al. Paeonol inhibits B16F10 melanoma metastasis in vitro and in vivo via disrupting proinflammatory cytokines-mediated NF-κB and STAT3 pathways. IUBMB Life 2015, 67, 778–788. [Google Scholar] [CrossRef] [Green Version]
- Horng, C.T.; Shieh, P.C.; Tan, T.W.; Yang, W.H.; Tang, C.H. Paeonol suppresses chondrosarcoma metastasis through up-regulation of miR-141 by modulating PKCδ and c-Src signaling pathway. Int. J. Mol. Sci. 2014, 15, 11760–11772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rho, S.; Chung, H.S.; Kang, M.; Lee, E.; Cho, C.; Kim, H.; Park, S.; Kim, H.Y.; Hong, M.; Shin, M.; et al. Inhibition of production of reactive oxygen species and gene expression profile by treatment of ethanol extract of Moutan cortex radicis in oxidative stressed PC12 cells. Biol. Pharm. Bull. 2005, 28, 661–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou, Y.; Li, Q.; Wang, J.; Li, K.; Zhou, S. Antitumor and apoptosis induction effects of paeonol on mice bearing EMT6 breast carcinoma. Biomol. Ther. 2014, 22, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Shu-Ping, X.; Guo-Ping, S.; Yu-Xian, S.; Wei, W.; Wan-Ren, P.; Hua, W. Antiproliferation and apoptosis induction of paenol in HepG2 cells. World J. Gastroenterol. 2007, 13, 250–256. [Google Scholar]
- Sun, G.-P.; Wang, H.; Xu, S.-P.; Shen, Y.-X.; Wu, Q.; Chen, Z.-D.; Wei, W. Anti-tumor effects of paeonol in a HepA-hepatoma bearing mouse model via induction of tumor cell apoptosis and stimulation of IL-2 and TNF-α production. Eur. J. Pharmacol. 2008, 584, 246–252. [Google Scholar] [CrossRef]
- Li, N.; Fan, L.L.; Sun, G.P.; Wan, X.A.; Wang, Z.G.; Wu, Q.; Wang, H. Paeonol inhibits tumor growth in gastric cancer in vitro and in vivo. World J. Gastroenterol. 2010, 16, 4483–4490. [Google Scholar] [CrossRef]
- Wu, Q.; Chen, G.L.; Li, Y.J.; Chen, Y.; Lin, F.Z. Paeoniflorin inhibits macrophage-mediated lung cancer metastasis. Chin. J. Nat. Med. 2015, 13, 925–932. [Google Scholar] [CrossRef]
- Li, W.; Qi, Z.; Wei, Z.; Liu, S.; Wang, P.; Chen, Y.; Zhao, Y. Paeoniflorin inhibits proliferation and induces apoptosis of human glioma cells via microRNA-16 upregulation and matrix metalloproteinase-9 downregulation. Mol. Med. Rep. 2015, 12, 2735–2740. [Google Scholar] [CrossRef] [Green Version]
- Oh, G.S.; Pae, H.O.; Oh, H.; Hong, S.G.; Kim, I.K.; Chai, K.Y.; Yun, Y.G.; Kwon, T.O.; Chung, H.T. In vitro anti-proliferative effect of 1,2,3,4,6-penta-O-galloyl-beta-D-glucose on human hepatocellular carcinoma cell line, SK-HEP-1 cells. Cancer Lett. 2001, 174, 17–24. [Google Scholar] [CrossRef]
- Lin, M.-Y.; Shen, C.-H.; Chiang, S.-Y.; Chen, S.-Y.; Lin, Y.-S.; Hsu, C.-D. Cortex Moutan inhibits bladder cancer cell proliferation and expression of angiogenic factors. Pharmacol. Pharm. 2014, 05, 846–858. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.H.; Weng, Y.P.; Tsai, H.Y.; Chen, C.J.; Lee, D.Y.; Hsieh, C.L.; Wu, Y.C.; Lin, J.Y. Aqueous extracts of Paeonia suffruticosa modulates mitochondrial proteostasis by reactive oxygen species-induced endoplasmic reticulum stress in pancreatic cancer cells. Phytomedicine 2018, 46, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, C. Paeonol induces antitumor effects in hepatocellular carcinoma cells through survivin via the cyclooxygenase-2/prostaglandin E2 signaling pathway. Transl. Cancer Res. 2020, 9, 7183–7195. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.H.; Lin, A.H.; Lee, H.F.; Ko, H.K.; Lee, T.S.; Kou, Y.R. Paeonol attenuates cigarette smoke-induced lung inflammation by inhibiting ros-sensitive inflammatory signaling. Mediators Inflamm. 2014, 2014, 651890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.G.; Park, G.; Piao, Y.; Kang, M.S.; Pak, Y.K.; Hong, S.-P.; Oh, M.S. Effects of the root bark of Paeonia suffruticosa on mitochondria-mediated neuroprotection in an MPTP-induced model of Parkinson’s disease. Food Chem. Toxicol. 2014, 65, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-b.; Song, N.-n.; Wei, X.-b.; Guan, H.-s.; Zhang, X. Protective effects of paeonol on cultured rat hippocampal neurons against oxygen-glucose deprivation-induced injury. J. Neurol. Sci. 2008, 264, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Himaya, S.W.A.; Ryu, B.; Qian, Z.J.; Kim, S.K. Paeonol from Hippocampus kuda Bleeler suppressed the neuro-inflammatory responses in vitro via NF-κB and MAPK signaling pathways. Toxicol. In Vitro 2012, 26, 878–887. [Google Scholar] [CrossRef]
- Lin, C.; Lin, H.; Chen, J.; Tseng, W.; Ko, P.; Liu, Y.; Yeh, W.; Lu, D. Effects of paeonol on anti-neuroinflammatory responses in microglial cells. Int. J. Mol. Sci. 2015, 16, 8844–8860. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Wang, G.; Zhao, A.; Gu, J.; Sun, X.; Hu, G. Paeoniflorin protects against ischemia-induced brain damages in rats via inhibiting MAPKs/NF-kB-mediated inflammatory responses. PLoS ONE 2012, 7, e49701. [Google Scholar] [CrossRef] [Green Version]
- Choi, B.-M.; Kim, H.-J.; Oh, G.-S.; Pae, H.-O.; Oh, H.; Jeong, S.; Kwon, T.-O.; Kim, Y.-M.; Chung, H.-T. 1,2,3,4,6-Penta-O-galloyl-beta-D-glucose protects rat neuronal cells (Neuro 2A) from hydrogen peroxide-mediated cell death via the induction of heme oxygenase-1. Neurosci. Lett. 2002, 328, 185–189. [Google Scholar] [CrossRef]
- Liu, S.; Li, Y.; Yi, F.; Liu, Q.; Chen, N.; He, X.; He, C.; Xiao, P. Resveratrol Oligomers from Paeonia suffruticosa Protect Mice against Cognitive Dysfunction by Regulating Cholinergic, Antioxidant and Anti-Inflammatory Pathways; Elsevier B.V.: Amsterdam, The Netherlands, 2020; Volume 260, ISBN 1282644149. [Google Scholar]
- Li, H.; Xie, Y.H.; Yang, Q.; Wang, S.W.; Zhang, B.L.; Wang, J.B.; Cao, W.; Bi, L.L.; Sun, J.Y.; Miao, S.; et al. Cardioprotective effect of paeonol and danshensu combination on isoproterenol-induced myocardial injury in rats. PLoS ONE 2012, 7, e48872. [Google Scholar] [CrossRef] [Green Version]
- Yun, C.S.; Choi, Y.G.; Jeong, M.Y.; Lee, J.H.; Lim, S. Moutan cortex radicis inhibits inflammatory changes of gene expression in lipopolysaccharide-stimulated gingival fibroblasts. J. Nat. Med. 2013, 67, 576–589. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Z.; Wu, J.H.; Yu, S.Y.; Shao, Q.R.; Dong, X.M. Inhibitory effects of paeoniflorin on lysophosphatidylcholine-induced inflammatory factor production in human umbilical vein endothelial cells. Int. J. Mol. Med. 2013, 31, 493–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Gu, Z. Screening of bioactive compounds from Moutan cortex and their anti-inflammatory activities in rat synoviocytes. Evid.-Based Complement. Altern. Med. 2009, 6, 57–63. [Google Scholar]
- Oh, G.S.; Pae, H.O.; Choi, B.M.; Jeong, S.; Oh, H.; Rho, Y.D.; Kim, D.H.; Shin, M.K.; Chung, H.T. Inhibitory effects of the root cortex of Paeonia suffruticosa on interleukin-8 and macrophage chemoattractant protein-1 secretions in U937 cells. J. Ethnopharmacol. 2003, 84, 85–89. [Google Scholar] [CrossRef]
- Chen, N.; Liu, D.; Soromou, L.W.; Sun, J.; Zhong, W.; Guo, W.; Huo, M.; Li, H.; Guan, S.; Chen, Z.; et al. Paeonol suppresses lipopolysaccharide- induced inflammatory cytokines in macrophage cells and protects mice from lethal endotoxin shock. Fundam. Clin. Pharmacol. 2013, 28, 268–276. [Google Scholar] [CrossRef]
- Liu, J.; Wang, S.; Feng, L.; Ma, D.; Fu, Q.; Song, Y.; Jia, X.; Ma, S. Hypoglycemic and antioxidant activities of paeonol and its beneficial effect on diabetic encephalopathy in streptozotocin-induced diabetic rats. J. Med. Food 2013, 16, 577–586. [Google Scholar] [CrossRef]
- Fu, J.; Li, Y.; Wang, L.; Gao, B.; Zhang, N.; Ji, Q. Paeoniflorin prevents diabetic nephropathy in rats. Comp. Med. 2009, 59, 557–566. [Google Scholar]
- Ha, D.T.; Tuan, D.T.; Thu, N.B.; Nhiem, N.X.; Ngoc, T.M.; Yim, N.; Bae, K. Palbinone and triterpenes from Moutan cortex (Paeonia suffruticosa, Paeoniaceae) stimulate glucose uptake and glycogen synthesis via activation of AMPK in insulin-resistant human HepG2 Cells. Bioorganic Med. Chem. Lett. 2009, 19, 5556–5559. [Google Scholar] [CrossRef]
- Shon, Y.-H.; Nam, K.-S. Protective effect of Moutan cortex extract on acetaminophen-induced hepatotoxicity in mice. J. Ethnopharmacol. 2004, 90, 415–419. [Google Scholar] [CrossRef]
- Hu, S.; Shen, G.; Zhao, W.; Wang, F.; Jiang, X.; Huang, D. Paeonol, the main active principles of Paeonia moutan, ameliorates alcoholic steatohepatitis in mice. J. Ethnopharmacol. 2010, 128, 100–106. [Google Scholar] [CrossRef]
- Wu, J.; Xue, X.; Zhang, B.; Jiang, W.; Cao, H.; Wang, R.; Sun, D.; Guo, R. The protective effects of paeonol against epirubicin-induced hepatotoxicity in 4T1-tumor bearing mice via inhibition of the PI3K/Akt/NF-kB pathway. Chem. Biol. Interact. 2016, 244, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cao, L.; Luo, Y.; Feng, X.; Sun, L.; Wen, M.; Peng, S. Paeoniflorin protects against concanavalin A-induced hepatitis in mice. Int. Immunopharmacol. 2015, 24, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ma, X.; Wang, J.; Zhu, Y.; Li, R.; Wang, J.; He, X.; Shan, L.; Wang, R.; Wang, L.; et al. Paeoniflorin alleviates liver fibrosis by inhibiting HIF-1α through mTOR-dependent pathway. Fitoterapia 2014, 99, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.; Liping, Z.; Xiaolin, Q.; Xiaohui, P.; Mingyan, W.; Xuan, T.; Shanggong, Y.; Nianbai, F. Moutan cortex extract exerts protective effects in a rat model of cardiac ischemia/reperfusion. Can. J. Physiol. Pharmacol. 2015, 94, 245–250. [Google Scholar]
- Chen, C.; Du, P.; Wang, J. Paeoniflorin ameliorates acute myocardial infarction of rats by inhibiting inflammation and inducible nitric oxide synthase signaling pathways. Mol. Med. Rep. 2015, 12, 3937–3943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Guo, Z.-P.; Wang, L.; Qin, S.; Cao, N.; Li, M.-M.; Jia, R.-Z.; Wang, T.-T. Paeoniflorin suppresses vascular damage and the expression of E-selectin and ICAM-1 in a mouse model of cutaneous Arthus reaction. Exp. Dermatol. 2013, 22, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Liu, X.; Mao, B.; Yang, L.; Liu, N. Paeonol enhances thrombus recanalization by inducing vascular endothelial growth factor 165 via ERK1/2 MAPK signaling pathway. Mol. Med. Rep. 2016, 13, 4853–4858. [Google Scholar] [CrossRef]
- Ye, S.; Mao, B.; Yang, L.; Fu, W.; Hou, J. Thrombosis recanalization by paeoniflorin through the upregulation of urokinase-type plasminogen activator via the MAPK signaling pathway. Mol. Med. Rep. 2016, 13, 4593–4598. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Dai, M.; Zhong, J.; Yin, D. Paeonol inhibits oxidized low density lipoprotein-induced monocyte adhesion to vascular endothelial cells by inhibiting the mitogen activated protein kinase pathway. Biol. Pharm. Bull. 2012, 35, 767–772. [Google Scholar] [CrossRef] [Green Version]
- Ngo, T.; Kim, K.; Bian, Y.; Noh, H.; Lim, K.M.; Chung, J.H.; Bae, O.N. Antithrombotic effects of paeoniflorin from Paeonia suffruticosa by selective inhibition on shear stress-induced platelet aggregation. Int. J. Mol. Sci. 2019, 20, 5040. [Google Scholar] [CrossRef] [Green Version]
- Lau, C.H.; Chan, C.M.; Chan, Y.W.; Lau, K.M.; Lau, T.W.; Lam, F.C.; Law, W.T.; Che, C.T.; Leung, P.C.; Fung, K.P.; et al. Pharmacological investigations of the anti-diabetic effect of Cortex Moutan and its active component paeonol. Phytomedicine 2007, 14, 778–784. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.Y.; Chou, T.H.; Lin, R.J.; Chan, L.P.; Wang, G.H.; Liang, C.H. Antioxidant and antimelanogenic behaviors of Paeonia suffruticosa. Plant Foods Hum. Nutr. 2011, 66, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Wang, S.H.; Song, T.Y.; Hsieh, C.W.; Tsai, M.S. Safety and efficacy of tyrosinase inhibition of Paeonia suffruticosa Andrews extracts on human melanoma cells. J. Cosmet. Dermatol. 2019, 18, 1921–1929. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, Q.; Zhao, Z.; Liu, Y.; Wang, Y.; Xiong, H.; Mei, Z. Paeonol ameliorates chronic itch and spinal astrocytic activation via CXCR3 in an experimental dry skin model in mice. Front. Pharmacol. 2022, 12, 1–15. [Google Scholar] [CrossRef]
- Hong, D.Y.; Pan, K.Y. Notes on taxonomy of Paeonia sect. Moutan DC. (Paeoniaceae). Acta Phytotaxon. Sin. 2005, 43, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Baoqing, L. Chinese Peony Book I. Science and Technology of China Press; China Press: Beijing, China, 2002. [Google Scholar]
- Wang, Z.; He, C.; Peng, Y.; Chen, F.; Xiao, P. Origins, phytochemistry, pharmacology, analytical methods and safety of Cortex Moutan (Paeonia suffruticosa Andrew): A systematic review. Molecules 2017, 22, 946. [Google Scholar] [CrossRef] [Green Version]
- Fu, P.K.; Yang, C.Y.; Tsai, T.H.; Hsieh, C.L. Moutan cortex radicis improves lipopolysaccharide-induced acute lung injury in rats through anti-inflammation. Phytomedicine 2012, 19, 1206–1215. [Google Scholar] [CrossRef]
- Junan, F.; Yan, Z.; Yongpeng, X.; Zongyin, Q.; Mengquan, Z. Analysis on production history and production status of mudanpi in Chongqing Dianjiang. J. Chinese Med. Mater. 2006, 29, 41–43. [Google Scholar]
- Zhou, S.L.; Zou, X.H.; Zhou, Z.Q.; Liu, J.; Xu, C.; Yu, J.; Wang, Q.; Zhang, D.M.; Wang, X.Q.; Ge, S.; et al. Multiple species of wild tree peonies gave rise to the “king of flowers”, Paeonia suffruticosa Andrews. Proceedings. Biol. Sci. 2014, 281, 20141687. [Google Scholar] [CrossRef] [Green Version]
- Ha, D.T.; Ngoc, T.M.; Lee, I.; Le, Y.M.; Kim, J.S.; Jung, H.; Lee, S.; Na, M.; Bae, K. Inhibitors of aldose reductase and formation of advanced glycation end-products in Moutan cortex (Paeonia suffruticosa). J. Nat. Prod. 2009, 72, 1465–1470. [Google Scholar] [CrossRef]
- Wu, S.H.; Wu, D.G.; Chen, Y.W. Chemical constituents and bioactivities of plants from the genus Paeonia. Chem. Biodivers. 2010, 7, 90–104. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Jiang, Z.; Liu, Y.; Chen, L.; Zhao, Q.; Yao, X.; Zhao, F.; Qiu, F. Monoterpenoid inhibitors of NO production from Paeonia suffruticosa. Fitoterapia 2012, 83, 1598–1603. [Google Scholar] [CrossRef] [PubMed]
- Song, W.H.; Cheng, Z.H.; Chen, D.F. Anticomplement monoterpenoid glucosides from the root bark of Paeonia suffruticosa. J. Nat. Prod. 2014, 77, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Seo, C.S.; Lee, K.S.; Kim, H.J.; Chang, H.W.; Jung, J.S.; Song, D.K.; Son, J.K. Protective constituents against sepsis in mice from the root cortex of Paeonia suffruticosa. Arch. Pharm. Res. 2004, 27, 1123–1126. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.C.; Ding, H.Y.; Wu, Y.C. Two novel compounds from Paeonia suffructicosa. J. Nat. Prod. 1998, 61, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H.; Yoo, H.J.; Noh, C.; Lee, J.M.; Park, J.W.; Choi, W.S.; Choi, J.H. Bioassay-guided isolation of novel compound from Paeonia suffruticosa Andrews roots as an IL-1β inhibitor. Arch. Pharm. Res. 2012, 35, 801–805. [Google Scholar] [CrossRef]
- Lin, H.C.; Ding, H.Y.; Wu, T.S.; Wu, P.L. Monoterpene glycosides from Paeonia suffruticosa. Phytochemistry 1996, 41, 237–242. [Google Scholar]
- Lai, X.; Wei, J.; Ding, X. Paeoniflorin antagonizes TNF-α-Induced L929 fibroblastoma cells apoptosis by inhibiting NF-κBp65 activation. Dose-Response 2018, 16, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Satoh, K.; Nagai, F.; Ushiyama, K.; Yasuda, I.; Seto, T.; Kano, I. Inhibition of Na+, K+-ATPase by 1,2,3,4,6-Penta-O-galloyl-β-D-glucose, a major constituent of both Moutan cortex and Paeoniae radix. Biochem. Pharmacol. 1997, 53, 611–614. [Google Scholar] [CrossRef]
- Yin, C.; Zhang, X.; Li, K.; Bai, Y.; Yang, P.; Li, C.; Song, X. Evaluation on the fresh eating quality of tree peony flowers. Food Biosci. 2022, 47, 101611. [Google Scholar] [CrossRef]
- Lei, G.; Song, C.; Wen, X.; Gao, G.; Qi, Y. Chemical diversity and potential target network of woody peony flower essential oil from eleven representative cultivars (Paeonia × suffruticosa Andr.). Molecules 2022, 27, 2829. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Dong, Y.; Liu, X.; Wan, Y.; Gu, T.; Zhou, X.; Liu, M. Comparison of chemical compositions, antioxidant, and anti-photoaging activities of Paeonia suffruticosa flowers at different flowering stages. Antioxidants 2019, 8, 345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, W.; Wang, Y.; Shehzad, Q.; Liu, Z.; Zeng, R. Effect of different solvents on the extraction of oil from peony seeds (Paeonia suffruticosa Andr.): Oil yield, fatty acids composition, minor components, and antioxidant capacity. J. Oleo Sci. 2022, 71, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yuan, Y.; Cui, J.; Xiao, T.; Jiang, D. Paeoniflorin inhibits proliferation and invasion of breast cancer cells through suppressing Notch-signaling pathway. Biomed. Pharmacother. 2016, 78, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.-Z.; Ge, Q.-H.; Rong, Q.; Li, Q. Paeonol attenuates neurotoxicity and ameliorates cognitive impairment induced by d-galactose in ICR mice. J. Neurol. Sci. 2009, 277, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, H.; Tabuchi, M.; Yamaguchi, T.; Iwasaki, K.; Furukawa, K.; Sekiguchi, K.; Ikarashi, Y.; Kudo, Y.; Higuchi, M.; Saido, T.C.; et al. A traditional medicinal herb Paeonia suffruticosa and its active constituent 1,2,3,4,6-penta-O-galloyl-β-D-glucopyranose have potent anti-aggregation effects on Alzheimer’s amyloid β proteins in vitro and in vivo. J. Neurochem. 2009, 109, 1648–1657. [Google Scholar] [CrossRef]
- Jiang, S.; Nakano, Y.; Yatsuzuka, R.; Ono, R.; Kamei, C. Inhibitory effects of Moutan cortex on immediate allergic reactions. Biol. Pharm. Bull. 2007, 30, 1707–1710. [Google Scholar] [CrossRef] [Green Version]
- Hong, M.H.; Kim, J.H.; Na, S.H.; Bae, H.; Shin, Y.C.; Kim, S.H.; Ko, S.G. Inhibitory effects of Paeonia suffruticosa on allergic reactions by inhibiting the NF-kappaB/IkappaB-alpha signaling pathway and phosphorylation of ERK in an animal model and human mast cells. Biosci. Biotechnol. Biochem. 2010, 74, 1152–1156. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Jiao, K.; Luo, L.; Xiang, J.; Fan, J.; Zhang, X.; Yi, J.; Zhu, W. Characterization and macrophage immunomodulatory activity of two polysaccharides from the flowers of Paeonia suffruticosa Andr. Int. J. Biol. Macromol. 2019, 124, 955–962. [Google Scholar] [CrossRef]
- Espiritu, A.G.; Doma, B.T.; Wang, Y.F.; Masim, F.C. Efficacy of methanol extracts of Mentha haplocalyx Briq. and Paeonia suffruticosa Andr. for potential antibacterial activity. Sustain. Environ. Res. 2014, 24, 319–324. [Google Scholar]
- Zhao, Y.; Wang, B.; Zhang, S.; Yang, S.; Wang, H.; Ren, A.; Yi, E. Isolation of antifungal compound from Paeonia suffruticosa and its antifungal mechanism. Chin. J. Integr. Med. 2015, 21, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.W.; Du, J.; Hwang, E.; Yi, T.-H. Paeonol extracted from Paeonia suffruticosa Andr. ameliorated UVB-induced skin photoaging via DLD/Nrf2/ARE and MAPK/ AP-1 pathway. Phyther. Res. 2018, 32, 1741–1749. [Google Scholar] [CrossRef] [PubMed]
- European Commission. CosIng—Cosmetic Database. Available online: https://ec.europa.eu/growth/tools-databases/cosing/index.cfm?fuseaction=search.simple (accessed on 9 August 2022).
- Yue, L.F.; Huang, X.W.; Gao, J.F.; Zhanghong, L.; Wang, D.M.; Sun, J.K.; Zhang, Z.G.; Wang, J.H.; Liu, J.; Wang, Y.M.; et al. Determination of components in Radix Paeoniae rubra based on QAMS. Pak. J. Pharm. Sci. 2022, 35, 1037–1041. [Google Scholar]
- Baye, H.; Hymete, A. Lead and cadmium accumulation in medicinal plants collected from environmentally different sites. Bull. Environ. Contam. Toxicol. 2010, 84, 197–201. [Google Scholar] [CrossRef]
- Liu, Y.; Xia, Y.; Guo, P.; Wang, G.; Shen, Z.; Xu, Y.; Chen, Y. Copper and bacterial diversity in soil enhance paeonol accumulation in Cortex Moutan of Paeonia suffruticosa “Fengdan”. Hortic. Environ. Biotechnol. 2013, 54, 331–337. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.; Yang, H.; Yu, P.; Tang, Y. Five heavy metals accumulation and health risk in a traditional Chinese medicine Cortex Moutan collected from different sites in China. Hum. Ecol. Risk Assess. Int. J. 2018, 24, 2288–2298. [Google Scholar] [CrossRef]
- Liu, X.H. Analysis of heavy metal elements lead and cadmium in Cortex Moutan and its planting soil. Chinese J. Spectrosc. Lab. 2013, 30, 821–824. [Google Scholar]
- Gildow, F.E.; Mitchell, J.P. Initiation, growth and nuclear characteristics of tissue cultures of Paeonia suffruticosa. Physiol. Plant. 1977, 39, 295–298. [Google Scholar] [CrossRef]
- Zhu, X.; Li, X.; Ding, W.; Jin, S.; Wang, Y. Callus induction and plant regeneration from leaves of peony. Hortic. Environ. Biotechnol. 2018, 59, 575–582. [Google Scholar] [CrossRef]
- Chen, X.; Ye, C.; Yang, H.; Ji, W.; Xu, Z.; Ye, S.; Wang, H.; Jin, S.; Yu, C.; Zhu, X. Callogenesis and plant regeneration in peony (Paeonia × suffruticosa) using flower petal explants. Horticulturae 2022, 8, 1–10. [Google Scholar] [CrossRef]





| Group of Compounds | Compounds | References |
|---|---|---|
| Monoterpenoids | 3-carene, citronellal, citronellol, citronellyl acetate, nerol, allo-ocimene, β-ocimene, (E)-β-ocimene, (Z)-β-ocimene, O-cymene, β-cymene, (E)- geraniol, (Z)- geraniol, cis-linalool oxide, linalool, trans-linalool oxide (furanoid) β-myrcene, α-phellandrene, α-pinene, (1s)-(-)-β-pinene, α-terpinolene, α-terpineol | [105,106] |
| Monoterpenoid glycosides | benzoylpaeoniflorin, benzoyloxypaeoniflorin, debenzoylgalloylpaeoniflorin, 8-debenzoylpaeoniflorin, galloyloxypaeoniflorin, galloylpaeoniflorin, oxypaeoniflorin, paeoniflorin, paeonolide | [107] |
| Phenols and their derivatives | mudanoside A, mudanpioside C, mudanpioside E, mudanpioside H, mudanpioside J, paeonol | [107] |
| Sesquiterpenoids | β-cadinene, cis-calamenene, caryophyllene, β-copaene, α-cubebene, α-farnesene, germacrene D, γ-muurolene | [105,106] |
| Alcohols | 1-decanol, 2-ethyl hexanol, cis-3-hexen-1-ol, 1-nonanol, cis-3-nonen-1-ol | [105,106] |
| Alkanes | 1-chloro-octadecane, cyclotetradecane, dodecane, eicosane, heptadecane, hexadecane, 8-hexylpentadecane, nonane, octane, tetradecane, undecane | [105,106] |
| Alkenes | 1-pentadecene | [105,106] |
| Alkatrienes | 3,4-dimethyl-2,4,6-octatriene, (E)-4,8-dimethylnona-1,3,7-triene, 1,3,8-triene-p-menthatriene, 1,5,8-triene-p-menthatriene | [105,106] |
| Adehydes | benzeneacetaldehyde, decanal, geranial, heptanal, myrtanal, neral, octanal | [106] |
| Phenolic acids | gallic acid, gallic acid-di-O-glucoside, gentisic acid-5-O-glucoside, p-hydroxybenzoic acid | [107] |
| Tannins | glucogallin | [107] |
| Flavonoids | apigenin, apigenin-7-O-glucoside, chrysoeriol, 6,3′-dimethoxyquercetin-di-O-glucoside, diosmin, eriodictyol-7-O-glucoside, hederagenin, isorhamnetin-3-O-glucoside, isorhamnetin-7-O-glucoside, isorhamnetin-3,7-di-O-glucoside, kaempferol-3-O-(2′’-O-galloyl)-glucoside, kaempferol-3-O-glucoside, kaempferol-3-O-rutinoside, kaempferol-3,7-di-O-glucoside, luteolin, monoxerutin, patuletin-3,5-di-O-glucoside, rhoifolin, quercetin-3-O-glucoside, quercetin-O-di-glucoside | [107] |
| Iridoid glycosides | geniposide, loganin, morroniside | [107] |
| Furans | 2-pentylfuran | [106] |
| Aminoacids | leucine, tryptophan | [107] |
| Sugars | sucrose | [107] |
| INCI Name | Description | Functions |
|---|---|---|
| Paeonia × suffruticosa bark extract | The extract of the bark of the Chinese Peony, Paeonia × suffruticosa, Paeoniaceae. | Skin conditioning |
| Paeonia × suffruticosa branch/flower/leaf extract | The extract of the branches, flowers and leaves of the Chinese Peony, Paeonia × suffruticosa, Paeoniaceae. | Skin conditioning |
| Paeonia × suffruticosa callus extract | The extract of the callus of Paeonia × suffruticosa, Paeoniaceae. | Antimicrobial Antioxidant Hair conditioning Skin protecting |
| Paeonia × suffruticosa extract | The extract of the whole plant Paeonia × suffruticosa, Paeoniaceae. | Skin conditioning |
| Paeonia × suffruticosa flower extract | The extract of the flowers of Paeonia × suffruticosa, Paeoniaceae. | Skin conditioning |
| Paeonia × suffruticosa flower water | The aqueous solution of the steam distillate obtained from the flowers of the Chinese Peony, Paeonia × suffruticosa, Paeoniaceae. | Fragrance |
| Paeonia × suffruticosa phytoplacenta extract | The extract of the phytoplacenta cells directly isolated from Paeonia × suffruticosa or grown in culture, Paeoniaceae. | Antimicrobial Antioxidant Hair conditioning Skin conditioning |
| Paeonia × suffruticosa root extract | The extract of the roots of the Chinese Peony, Paeonia × suffruticosa, Paeoniaceae. | Skin protecting |
| Paeonia × suffruticosa root water | The aqueous solution of the steam distillates obtained from the roots of the Chinese Peony, Paeonia × suffruticosa, Paeoniaceae. | Flavoring Fragrance Perfuming Skin conditioning |
| Paeonia × suffruticosa seed oil | The oil expressed from the seeds of Paeonia × suffruticosa, Paeoniaceae. | Skin conditioning Skin conditioning-emollient |
| Aspergillus/Paeonia × suffruticosa bark/Honeysuckle flower/Forsythia fruit (Astragalus membranaceus/Gentiana scabra/Licorice/Rehmannia glutinosa/Rheum palmatum/Scrophularia buergeriana) root/(castor/rice) seed ferment extract | The extract of the product obtained by the fermentation of the bark of Paeonia × suffruticosa; the flower of Lonicera japonica (honeysuckle); the fruit of Forsythia suspensa (forsythia); the roots of Astragalus membranaceus, Gentiana scabra, Glycyrrhiza glabra (licorice), Rehmannia glutinosa (rehmannia), Rheum palmatum and Scrophularia buergeriana; the seeds of Oryza sativa (rice), and Ricinus communis (castor), by the microorganism Aspergillus. | Skin conditioning |
| Lactobacillus/(Achyranthes bidentata/Angelica gigas/Angelica pubescens/Angelica tenuissima/Asarum sieboldi/Cnidium officinale/Ledebouriella divaricata/Paeonia × suffruticosa) Root/Eucommia ulmoides Bark/Magnolia liliflora Bud Ferment Filtrate | The filtrate of the product obtained by the fermentation of the roots of Achyranthes bidentata, Angelica gigas, Angelica pubescens, Angelica tenuissima, Asarum sieboldi, Cnidium officinale, Ledebouriella divaricata, Paeonia × suffruticosa, the bark of Eucommia ulmoides and the buds of Magnolia liliflora by the microorganism, Lactobacillus. | Skin conditioning-miscellaneous |
| Lactobacillus/(Cudrania tricuspidata/Paeonia × suffruticosa) Bark/Lycium chinense Fruit/Apricot Kernel/Artemisia capillaris Leaf/(Angelica dahurica/Scutellaria baicalensis) Root/Soybean Seed/Houttuynia cordata/Mistletoe/Poria Cocos ferment filtrate | The filtrate of the product obtained by the fermentation of the bark of Cudrania tricuspidata, and Paeonia × suffruticosa; the fruits of Lycium chinense; the kernels of Prunus armeniaca (apricot); the leaves of Artemisia capillaris; the roots of Angelica dahurica, and Scutellaria baicalensis; the seeds of Glycine soja (soybean); the whole plants, Houttuynia cordata, and Viscum album (mistletoe); and the fungus, Poria cocos, by the microorganism Lactobacillus. | Humectant Skin conditioning |
| Lactobacillus/Ledebouriella divaricate root/Angelica dahurica root/Angelica tenuissima root/Magnolia liliflora bud/Asarum sieboldin root/Paeonia × suffruticosa root/Cnidium officinale root/Angelica gigas root/Aukcklandia lappa root/Achyranthes japonica root/Eucommia ulmoides bark/Angelica pubescens root/Aconitum koreanum root ferment filtrate | The a filtrate of the product obtained by the fermentation of the roots of Ledebouriella divaricata, Angelica dahurica, Angelica tenuissima, Asarum sieboldi, Paeonia × suffruticosa, Cnidium officinale, Angelica gigas, Auklandia lappa, Achyranthes japonica, Angelica pubescens, and Aconitum koreanum, the buds of Magnolia liliflora and the bark of Eucommia ulmoides by the microorganism, Lactobacillus. | Skin conditioning |
| Leuconostoc/Ledebouriella divaricata root/Angelica dahurica root/Angelica tenuissima root/Magnolia liliflora bud/Asarum sieboldi root/Paeonia × suffruticosa root/Cnidium officinale root/Angelica gigas root/Aucklandia lappa root/Achyranthes japonoica root/Eucommia ulmoides bark/Angelica pubescens root/Aconitum koreanum root ferment | The product obtained by the fermentation of the roots of Ledebouriella divaricata, Angelica dahurica, Angelica tenuissima, Asarum sieboldi, Paeonia × suffruticosa, Cnidium officinale, Angelica gigas, Aucklandia lappa, Achyranthes japonoica, Angelica pubescens, Aconitum koreanum, the buds of Magnolia liliflora, and the bark of Eucommia ulmoides, by the microorganism, Leuconostoc. | Skin Conditioning |
| Monascus/Paeonia × suffruticosa flower/rice bran ferment filtrate | The filtrate of the product obtained by the fermentation of the flowers of Paeonia × suffruticosa and the bran of Oryza sativa (rice) by the microorganism, Monascus. | Anti-sebum Skin conditioning |
| Acetobacter/Ledebouriella divaricata root/Angelica dahurica root/Angelica tenuissama root/Magnolia liliflora bud/Asarum sieboldi root/Paeonia × suffruticosa root/Cnidium officinale root/Angelica gigas root/Aucklandia lappa root/Achyranthes japonica root/Eucommia ulmoides bark/Angelica pubescens root/Aconitum koreanum root ferment filtrate | The filtrate of the product obtained by the fermentation of Ledebouriella divaricata roots, Angelica dahurica roots, Angelica tenuissama roots, Magnolia liliflora buds, Asarum sieboldi roots, Paeonia × suffruticosa roots, Cnidium officinale roots, Angelica gigas roots, Auklandia lappa roots, Achyranthes japonica roots, Eucommia ulmoides bark, Angelica pubescens roots, Aconitum koreanum roots by the microorganism Acetobacter. | Skin conditioning |
| Saccharomyces/(Achyranthes bidentata/Angelica gigas/Angelica pubescens/Angelica tenuissima/Asarum sieboldi/Cnidium officinale/Ledebouriella divaricata/Paeonia × suffruticosa) root/Eucommia ulmoides bark/Magnolia liliflora bud ferment filtrate | The filtrate of the product obtained by the fermentation of the roots of Achyranthes bidentata, Angelica gigas, Angelica pubescens, Angelica tenuissima, Asarum sieboldi, Cnidium officinale, Ledebouriella divaricata, Paeonia × suffruticosa, the bark of Eucommia ulmoides and the buds of Magnolia liliflora by the microorganism, Saccharomyces. | Skin conditioning |
| Saccharomyces/Achyranthes bidentat root/Angelica gigas root/Angelica pubescens root/Angelica tenuissima root/Asarum sieboldi root/Cnidium officinale root/Eucommia ulmoides bark/Ledebouriella divaricata root/Magnolia liliflora bud/Paeonia × suffruticosa root ferment filtrate | The filtrate of the product obtained by the fermentation of the roots of Achyranthes bidentata, Angelica gigas, Angelica pubescens, Angelica tenuissima, Asarum sieboldi, Cnidium officinale, Ledebouriella divaricata, Paeonia × suffruticosa, the buds of Magnolia liliflora, and the bark of Eucommia ulmoides by the microorganism, Saccharomyces. | Skin conditioning |
| Saccharomyces/Achyranthes bidentata root/Angelica gigas root/Angelica pubescens root/Angelica tenuissima root/Cnidium officinale root/Eucommia ulmoides bark/Ledebouriella divaricata root/Paeonia × suffruticosa root ferment filtrate | The filtrate of the product obtained by the fermentation of the roots of Achyranthes bidentata, Angelica gigas, Angelica pubescens, Angelica tenuissima, Cnidium officinale, Ledebouriella divaricata, Paeonia × suffruticosa, and the bark of Eucommia ulmoides by the microorganism, Saccharomyces. | Skin conditioning |
| Saccharomyces/Camellia japonica flower/Castanea crenata shell/Diospyros kaki leaf/Paeonia × suffruticosa root/Rhus javanica/Sanguisorba officinalis root extract ferment filtrate | The filtrate of the product obtained by the fermentation of Camellia japonica flower extract, Castanea crenata shell extract, Diospyros kaki leaf extract, Paeonia × suffruticosa root extract, Rhus javanica extract and Sanguisorba officinalis root extract by the microorganism, Saccharomyces. | Skin conditioning |
| Saccharomyces/Cyperus rotundus root/Magnolia obovata bark/Paeonia × suffruticosa root/Peach kernel ferment extract filtrate | The filtrate of the extract of the product obtained by the fermentation of the roots of Cyperus rotundus and Paeonia × suffruticosa, the bark of Magnolia obovata, and the kernel of Prunus persica (peach) by the microorganism, Saccharomyces. | Skin conditioning |
| Saccharomyces/Ledebouriella divaricata root/Achyranthes bidentata root/Cnidium officinale root/Eucommia ulmoides bark/Angelica gigas root/Paeonia × suffruticosa root/Angelica tenuissim root/Asarum sieboldi root/Angelica pubescens root/Magnolia liliflora bud ferment filtrate | The filtrate of the product obtained by the fermentation of the roots of Ledebouriella divaricata, Achyranthes bidentata, Cnidium officinale, Angelica gigas, Paeonia × suffruticosa, Angelica tenuissima, Asarum sieboldi, Angelica pubescens, the bark of Eucommia ulmoides and the buds of Magnolia liliflora by the microorganism, Saccharomyces. | Skin conditioning |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ekiert, H.; Klimek-Szczykutowicz, M.; Szopa, A. Paeonia × suffruticosa (Moutan Peony)—A Review of the Chemical Composition, Traditional and Professional Use in Medicine, Position in Cosmetics Industries, and Biotechnological Studies. Plants 2022, 11, 3379. https://doi.org/10.3390/plants11233379
Ekiert H, Klimek-Szczykutowicz M, Szopa A. Paeonia × suffruticosa (Moutan Peony)—A Review of the Chemical Composition, Traditional and Professional Use in Medicine, Position in Cosmetics Industries, and Biotechnological Studies. Plants. 2022; 11(23):3379. https://doi.org/10.3390/plants11233379
Chicago/Turabian StyleEkiert, Halina, Marta Klimek-Szczykutowicz, and Agnieszka Szopa. 2022. "Paeonia × suffruticosa (Moutan Peony)—A Review of the Chemical Composition, Traditional and Professional Use in Medicine, Position in Cosmetics Industries, and Biotechnological Studies" Plants 11, no. 23: 3379. https://doi.org/10.3390/plants11233379
APA StyleEkiert, H., Klimek-Szczykutowicz, M., & Szopa, A. (2022). Paeonia × suffruticosa (Moutan Peony)—A Review of the Chemical Composition, Traditional and Professional Use in Medicine, Position in Cosmetics Industries, and Biotechnological Studies. Plants, 11(23), 3379. https://doi.org/10.3390/plants11233379