
Strawberry Biostimulation: From Mechanisms of Action to Plant Growth and Fruit Quality

 , ,
, ,  , ,
, ,  ,
,  ,
, 
Abstract
:1. Introduction
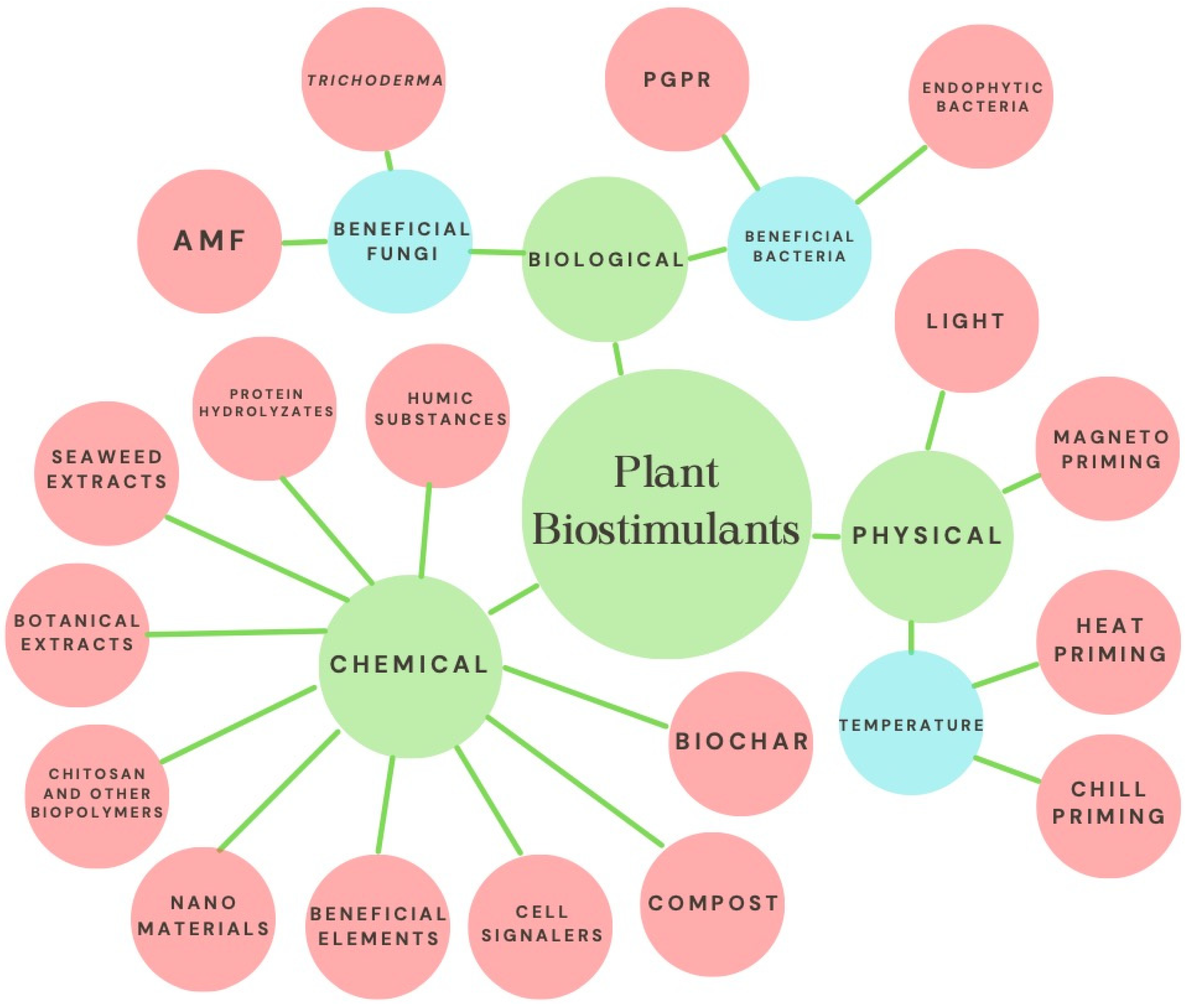
2. General Mechanism of Plant Biostimulation
2.1. Plant Cell Receptors
2.2. From Perception to Transduction and Signaling
3. Use of Chemical and Biochemical Biostimulants in Strawberry Cropping
3.1. Humic Substances (HS)
3.2. Protein Hydrolysates (PHs)
3.3. Seaweed and Algal and Microalgal Extracts
3.4. Botanical Extracts
3.5. Chitosan and Other Biopolymers
3.6. Beneficial Elements
3.7. Metal, Carbon, Zeolite, and Chitosan Nanomaterials
3.8. Compost
3.9. Biochar
3.10. H2O2, NO, H2S, H2, CH4, and CO
4. Use of Biological Biostimulants in Strawberry Cropping
4.1. Beneficial Bacteria
4.1.1. PGPR
4.1.2. Endophytic Bacteria
4.2. Beneficial Fungi
4.2.1. Arbuscular Mycorrhizal Fungi (AMF)
4.2.2. Trichoderma
5. Use of Physical Biostimulants in Strawberry Cropping
5.1. Biostimulation and Priming Using UV and Visible Light
5.2. Biostimulation and Priming Using Heat Shock and Chill Priming
5.3. Magnetopriming
6. Comments and Future Perspectives
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Dijk, M.; Morley, T.; Rau, M.L.; Saghai, Y. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat. Food 2021, 2, 494–501. [Google Scholar] [CrossRef]
- Clapp, J.; Newell, P.; Brent, Z.W. The global political economy of climate change, agriculture and food systems. J. Peasant Stud. 2018, 45, 80–88. [Google Scholar] [CrossRef]
- FAO Food and Agriculture Data. Available online: https://www.fao.org/faostat/en/#data (accessed on 20 October 2022).
- du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Juárez-Maldonado, A.; Ortega-Ortíz, H.; Morales-Díaz, A.B.; González-Morales, S.; Morelos-Moreno, Á.; Cabrera-De la Fuente, M.; Sandoval-Rangel, A.; Cadenas-Pliego, G.; Benavides-Mendoza, A. Nanoparticles and nanomaterials as plant biostimulants. Int. J. Mol. Sci. 2019, 20, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; He, C. A review of redox signaling and the control of MAP kinase pathway in plants. Redox Biol. 2017, 11, 192–204. [Google Scholar] [CrossRef] [PubMed]
- González-Morales, S.; Solís-Gaona, S.; Valdés-Caballero, M.V.; Juárez-Maldonado, A.; Loredo-Treviño, A.; Benavides-Mendoza, A. Transcriptomics of Biostimulation of Plants Under Abiotic Stress. Front. Genet. 2021, 12, 583888. [Google Scholar] [CrossRef] [PubMed]
- Righini, H.; Roberti, R.; Baraldi, E. Use of algae in strawberry management. J. Appl. Phycol. 2018, 30, 3551–3564. [Google Scholar] [CrossRef]
- Zipfel, C. Plant pattern-recognition receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef]
- Liang, X.; Zhou, J.M. Receptor-Like Cytoplasmic Kinases: Central Players in Plant Receptor Kinase-Mediated Signaling. Annu. Rev. Plant Biol. 2018, 69, 267–299. [Google Scholar] [CrossRef] [Green Version]
- Kanneganti, V.; Gupta, A.K. Wall associated kinases from plants—An overview. Physiol. Mol. Biol. Plants 2008, 14, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, N.; Mahajan, S. Calcium signaling network in plants: An overview. Plant Signal. Behav. 2007, 2, 79–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nephali, L.; Piater, L.A.; Dubery, I.A.; Patterson, V.; Huyser, J.; Burgess, K.; Tugizimana, F. Biostimulants for plant growth and mitigation of abiotic stresses: A metabolomics perspective. Metabolites 2020, 10, 505. [Google Scholar] [CrossRef]
- Taj, G.; Agarwal, P.; Grant, M.; Kumar, A. MAPK machinery in plants. Plant Signal. Behav. 2010, 5, 1370–1378. [Google Scholar] [CrossRef] [Green Version]
- Heinemann, B.; Künzler, P.; Eubel, H.; Braun, H.P.; Hildebrandt, T.M. Estimating the number of protein molecules in a plant cell: Protein and amino acid homeostasis during drought. Plant Physiol. 2021, 185, 385–404. [Google Scholar] [CrossRef]
- Thor, K. Calcium—Nutrient and messenger. Front. Plant Sci. 2019, 10, 440. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Tanaka, K.; Nguyen, C.T.; Stacey, G. Extracellular ATP is a central signaling molecule in plant stress responses. Curr. Opin. Plant Biol. 2014, 20, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic acid and abiotic stress tolerance in crop plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayakannan, M.; Bose, J.; Babourina, O.; Rengel, Z.; Shabala, S. Salicylic acid in plant salinity stress signalling and tolerance. Plant Growth Regul. 2015, 76, 25–40. [Google Scholar] [CrossRef]
- Kapoor, D.; Sharma, R.; Handa, N.; Kaur, H.; Rattan, A.; Yadav, P.; Gautam, V.; Kaur, R.; Bhardwaj, R. Redox homeostasis in plants under abiotic stress: Role of electron carriers, energy metabolism mediators and proteinaceous thiols. Front. Environ. Sci. 2015, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive Oxygen Species in Plant Signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [PubMed]
- Astier, J.; Lindermayr, C. Nitric oxide-dependent posttranslational modification in plants: An update. Int. J. Mol. Sci. 2012, 13, 15193–15208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Morales, S.; López-Sánchez, R.C.; Juárez-Maldonado, A.; Robledo-Olivo, A.; Benavides-Mendoza, A. A Transcriptomic and Proteomic View of Hydrogen Sulfide Signaling in Plant Abiotic Stress. In Plant in Challenging Environments; Gupta, D., Palma, J.M., Corpas, F.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 161–186. ISBN 9783030736774. [Google Scholar]
- Joshi, R.; Wani, S.H.; Singh, B.; Bohra, A.; Dar, Z.A.; Lone, A.A.; Pareek, A.; Singla-Pareek, S.L. Transcription factors and plants response to drought stress: Current understanding and future directions. Front. Plant Sci. 2016, 7, 1029. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Gu, Y. Structural and functional insight into the nuclear pore complex and nuclear transport receptors in plant stress signaling. Curr. Opin. Plant Biol. 2020, 58, 60–68. [Google Scholar] [CrossRef]
- Rawat, N.; Singla-Pareek, S.L.; Pareek, A. Membrane dynamics during individual and combined abiotic stresses in plants and tools to study the same. Physiol. Plant. 2021, 171, 653–676. [Google Scholar] [CrossRef] [PubMed]
- Hayes, S.; Schachtschabel, J.; Mishkind, M.; Munnik, T.; Arisz, S.A. Hot topic: Thermosensing in plants. Plant Cell Environ. 2021, 44, 2018–2033. [Google Scholar] [CrossRef] [PubMed]
- Muscolo, A.; Sidari, M.; Nardi, S. Humic substance: Relationship between structure and activity. deeper information suggests univocal findings. J. Geochem. Explor. 2013, 129, 57–63. [Google Scholar] [CrossRef]
- Weber, J.; Chen, Y.; Jamroz, E.; Miano, T. Preface: Humic substances in the environment. J. Soils Sediments 2018, 18, 2665–2667. [Google Scholar] [CrossRef] [Green Version]
- Trevisan, S.; Francioso, O.; Quaggiotti, S.; Nardi, S. Humic substances biological activity at the plant-soil interface: From environmental aspects to molecular factors. Plant Signal. Behav. 2010, 5, 635–643. [Google Scholar] [CrossRef] [Green Version]
- Rose, M.T.; Patti, A.F.; Little, K.R.; Brown, A.L.; Jackson, W.R.; Timothy, R. A meta-analysis and review of plant-growth response to humic substances: Practical implications for agriculture. Adv. Agron. 2014, 124, 37–89. [Google Scholar]
- García, A.C.; van Tol de Castro, T.A.; Santos, L.A.; Tavares, O.C.H.; Castro, R.N.; Berbara, R.L.L.; García-Mina, J.M. Structure-Property-Function Relationship of Humic Substances in Modulating the Root Growth of Plants: A Review. J. Environ. Qual. 2019, 48, 1622–1632. [Google Scholar] [CrossRef]
- Aeschbacher, M.; Graf, C.; Schwarzenbach, R.; Sander, M. Antioxidant Properties of Humic Substances. Environ. Sci. Technol. 2012, 46, 4916–4925. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 871, 1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and fulvic acids as biostimulants in horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Aghaeifard, F.; Babalar, M.; Fallahi, E.; Ahmadi, A. Influence of humic acid and salicylic acid on yield, fruit quality, and leaf mineral elements of strawberry (Fragaria × Ananassa duch.) cv. Camarosa. J. Plant Nutr. 2016, 39, 1821–1829. [Google Scholar] [CrossRef]
- Arancon, N.Q.; Lee, S.; Edwards, C.A.; Atiyeh, R. Effects of humic acids derived from cattle, food and paper-waste vermicomposts on growth of greenhouse plants. Pedobiologia 2003, 47, 741–744. [Google Scholar] [CrossRef] [Green Version]
- Arancon, N.Q.; Edwards, C.A.; Lee, S.; Byrne, R. Effects of humic acids from vermicomposts on plant growth. Eur. J. Soil Biol. 2006, 42, 65–69. [Google Scholar] [CrossRef]
- Belyaev, A.A.; Pospelova, N.P.; Lelyak, A.A.; Shternshis, M.V.; Shpatova, T.V. The use of Bacillus spp. Strains for biocontrol of Ramularia leaf spot on strawberry and improving plant health in Western Siberia. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 1594–1607. [Google Scholar]
- de Santiago, A.; Carmona, E.; Quintero, J.M.; Delgado, A. Effectiveness of mixtures of vivianite and organic materials in preventing iron chlorosis in strawberry. Spanish J. Agric. Res. 2013, 11, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Kazemi, M. The impact of foliar humic acid sprays on reproductive biology and fruit quality of strawberry. Thai J. Agric. Sci. 2014, 47, 221–225. [Google Scholar]
- Mufty, R.K.; Taha, S.M. Response Two Strawberry Cultivars (Fragaria × Ananassa Duch.) for Foliar Application of Two Organic Fertilizers. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Babil, Iraq, 4–5 October 2021; Volume 910. [Google Scholar] [CrossRef]
- Narouei, Z.; Sedaghathoor, S.; Kaviani, B.; Ansari, M.H. Biochemical and physiological traits of strawberry as influenced by organic acids and deficit irrigation under colored netting. Agric. Nat. Resour. 2021, 55, 1023–1038. [Google Scholar] [CrossRef]
- Neri, J.C.; Meléndez-Mori, J.B.; Tejada-Alvarado, J.J.; Vilca-Valqui, N.C.; Huaman-Huaman, E.; Oliva, M.; Goñas, M. An Optimized Protocol for Micropropagation and Acclimatization of Strawberry (Fragaria × ananassa Duch.) Variety ‘Aroma’. Agronomy 2022, 12, 968. [Google Scholar] [CrossRef]
- Neri, D.; Lodolini, E.M.; Savini, G.; Sabbatini, P.; Bonanomi, G.; Zucconi, F. Foliar application of humic acids on strawberry (cv Onda). Acta Hortic. 2002, 594, 297–302. [Google Scholar] [CrossRef]
- Rafeii, S.; Pakkish, Z. Improvement of vegetative and reproductive growth of ‘camarosa’ strawberry: Role of humic acid, Zn, and B. Agric. Conspec. Sci. 2014, 79, 239–244. [Google Scholar]
- Rätsep, R.; Vool, E.; Karp, K. Influence of humic fertilizer on the quality of strawberry cultivar “Darselect”. Acta Hortic. 2014, 1049, 911–916. [Google Scholar] [CrossRef]
- Rostami, M.; Shokouhian, A.; Mohebodini, M. Effect of Humic Acid, Nitrogen Concentrations and Application Method on the Morphological, Yield and Biochemical Characteristics of Strawberry ‘Paros’. Int. J. Fruit Sci. 2022, 22, 203–214. [Google Scholar] [CrossRef]
- Rzepka-Plevnes, D.; Kulpa, D.; Golebiowska, D.; Porwolik, D. Effects of auxins and humic acids on in vitro rooting of strawberry (Fragaria × ananassa Duch.). J. Food Agric. Environ. 2011, 9, 592–595. [Google Scholar]
- Saidimoradi, D.; Ghaderi, N.; Javadi, T. Salinity stress mitigation by humic acid application in strawberry (Fragaria × ananassa Duch.). Sci. Hortic. 2019, 256, 108594. [Google Scholar] [CrossRef]
- Soppelsa, S.; Kelderer, M.; Casera, C.; Bassi, M.; Robatscher, P.; Matteazzi, A.; Andreotti, C. Foliar applications of biostimulants promote growth, yield and fruit quality of strawberry plants grown under nutrient limitation. Agronomy 2019, 9, 483. [Google Scholar] [CrossRef] [Green Version]
- Tehranifar, A.; Ameri, A. Effect of humic acid on nutrient uptake and physiological characteristics of Fragaria × ananassa “Camarosa”. Acta Hortic. 2014, 1049, 391–394. [Google Scholar] [CrossRef]
- Wasi Amiri, A.; Nache Gowda, V.; Shymmlama, S.; Vinaya Kumar Reddy, P. Influence of bio-inoculants on nursery establishment of strawberry “Sujatha”. Acta Hortic. 2011, 890, 155–160. [Google Scholar] [CrossRef]
- Yaman, M.; Yilmaz, K.U. The Effects of Different Chemicals on Runner Yield and Quality of ‘Kabarla’ Strawberry Young Plants Grown in Cappadocia Region. Erwerbs-Obstbau 2022, 64, 85–90. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Di Mattia, E.; El-Nakhel, C.; Cardarelli, M. Co-inoculation of Glomus intraradices and Trichoderma atroviride acts as a biostimulant to promote growth, yield and nutrient uptake of vegetable crops. J. Sci. Food Agric. 2015, 95, 1706–1715. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Hernández, J.M.; Benítez-García, I.; Mazorra-Manzano, M.A.; Ramírez-Suárez, J.C.; Sánchez, E. Strategies for production, characterization and application of protein-based biostimulants in agriculture: A review. Chil. J. Agric. Res. 2020, 80, 274–289. [Google Scholar] [CrossRef]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant action of protein hydrolysates: Unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef]
- Nardi, S.; Pizzeghello, D.; Schiavon, M.; Ertani, A. Plant biostimulants: Physiological responses induced by protein hydrolyzed-based products and humic substances in plant metabolism. Sci. Agric. 2016, 73, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Bogunovic, I.; Duralija, B.; Gadze, J.; Kisic, I. Biostimulant usage for preserving strawberries to climate damages. Hortic. Sci. 2015, 42, 132–140. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Wang, G.; Du, M.; Niu, C.; Zhang, P.; Zhang, X.; Ma, D.; Ma, F.; Bao, Z. Biostimulants promote plant vigor of tomato and strawberry after transplanting. Sci. Hortic. 2020, 267, 9355. [Google Scholar] [CrossRef]
- Gerdakaneh, M.; Mozafari, A.A.; sioseh-mardah, A.; Sarabi, B. Effects of different amino acids on somatic embryogenesis of strawberry (Fragaria × ananassa Duch.). Acta Physiol. Plant. 2011, 33, 1847–1852. [Google Scholar] [CrossRef]
- Marfà, O.; Cáceres, R.; Polo, J.; Ródenas, J. Animal protein hydrolysate as a biostimulant for transplanted strawberry plants subjected to cold stress. Acta Hortic. 2009, 842, 315–318. [Google Scholar] [CrossRef]
- Mohseni, F.; Pakkish, Z.; Panahi, B. Arginine impact on yield and fruit qualitative characteristics of strawberry. Agric. Conspec. Sci. 2017, 82, 19–26. [Google Scholar]
- Talukder, M.R.; Asaduzzaman, M.; Tanaka, H.; Asao, T. Light-emitting diodes and exogenous amino acids application improve growth and yield of strawberry plants cultivated in recycled hydroponics. Sci. Hortic. 2018, 239, 93–103. [Google Scholar] [CrossRef]
- Wang, B.; Lai, T.; Huang, Q.W.; Yang, X.M.; Shen, Q.R. Effect of N Fertilizers on Root Growth and Endogenous Hormones in Strawberry Project supported by the National High Technology Research and Development Program (863 Program) of China (No. 2004AA246080) and the Program for the Development of High-Tech Indu. Pedosphere 2009, 19, 86–95. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Stirk, W.A.; Rengasamy, K.R.R.; Kulkarni, M.G.; van Staden, J. Plant biostimulants from seaweed: An overview. In The Chemical Biology of Plant Biostimulants; Geelen, D., Xu, L., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2020; pp. 31–55. ISBN 9781119357193. [Google Scholar]
- Al-Juthery, H.W.A.; Abbas Drebee, H.; Al-Khafaji, B.M.K.; Hadi, R.F. Plant Biostimulants, Seaweeds Extract as a Model (Article Review). In Proceedings of the IOP Conference Series: Earth and Environmental Science, Al-Qadisiyah, Iraq, 31 May–1 June 2020; Volume 553. [Google Scholar] [CrossRef]
- EL Boukhari, M.E.M.; Barakate, M.; Bouhia, Y.; Lyamlouli, K. Trends in Seaweed Extract Based Biostimulants: Manufacturing Process and beneficial effect on soil-plant systems. Plants 2020, 9, 359. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.Z.; Braun, G.; Norrie, J.; Hodges, D.M. Effect of Ascophyllum extract application on plant growth, fruit yield and soil microbial communities of strawberry. Can. J. Plant Sci. 2013, 93, 23–36. [Google Scholar] [CrossRef]
- Al-Shatri, A.H.N.; Pakyürek, M.; Yaviç, A. Effect of seaweed application on nutrient uptake of strawberry cv. Albion grown under the environmental conditions of northern iraq. Appl. Ecol. Environ. Res. 2020, 18, 1267–1279. [Google Scholar] [CrossRef]
- Al-Shatri, A.H.N.; Pakyürek, M.; Yaviç, A. Effect of seaweed application on the vegetative growth of strawberry cv. Albion grown under iraq ecological conditions. Appl. Ecol. Environ. Res. 2020, 18, 1211–1225. [Google Scholar] [CrossRef]
- Bajpai, S.; Shukla, P.S.; Asiedu, S.; Pruski, K.; Prithiviraj, B. A biostimulant preparation of brown seaweed Ascophyllum nodosum suppresses powdery mildew of strawberry. Plant Pathol. J. 2019, 35, 406–416. [Google Scholar] [CrossRef]
- Celiktopuz, E.; Kapur, B.; Sarıdas, M.A.; Kargı, S.P. Response of strawberry fruit and leaf nutrient concentrations to the application of irrigation levels and a biostimulant. J. Plant Nutr. 2021, 44, 153–165. [Google Scholar] [CrossRef]
- El-Miniawy, S.M.; Ragab, M.E.; Youssef, S.M.; Metwally, A.A. Influence of Foliar Spraying of Seaweed Extract on Growth, Yield and Quality of Strawberry Plants. J. Appl. Sci. Res. 2016, 10, 88–94. [Google Scholar]
- Holden, D.; Ross, R. Six years of strawberry trials in commercial fields demonstrate that an extract of the brown seaweed Ascophyllum nodosum improves yield of strawberries. Acta Hortic. 2017, 1156, 249–254. [Google Scholar] [CrossRef]
- Kapur, B.; Çeliktopuz, E.; Sarıdaş, M.A.; Kargi, S.P. Irrigation regimes and bio-stimulant application effects on yield and morpho-physiological responses of strawberry. Hortic. Sci. Technol. 2018, 36, 313–325. [Google Scholar] [CrossRef] [Green Version]
- Kapur, B.; Sarıdaş, M.A.; Çeliktopuz, E.; Kafkas, E.; Paydaş Kargı, S. Health and taste related compounds in strawberries under various irrigation regimes and bio-stimulant application. Food Chem. 2018, 263, 67–73. [Google Scholar] [CrossRef]
- Mattner, S.W.; Milinkovic, M.; Arioli, T. Increased growth response of strawberry roots to a commercial extract from Durvillaea potatorum and Ascophyllum nodosum. J. Appl. Phycol. 2018, 30, 2943–2951. [Google Scholar] [CrossRef]
- Spinelli, F.; Fiori, G.; Noferini, M.; Sprocatti, M.; Costa, G. A novel type of seaweed extract as a natural alternative to the use of iron chelates in strawberry production. Sci. Hortic. 2010, 125, 263–269. [Google Scholar] [CrossRef]
- Kocira, S.; Szparaga, A.; Krawczuk, A.; Bartoš, P.; Zaguła, G.; Plawgo, M.; Černý, P. Plant material as a novel tool in designing and formulating modern biostimulants—Analysis of botanical extract from Linum usitatissimum L. Materials 2021, 14, 6661. [Google Scholar] [CrossRef]
- Szparaga, A.; Kocira, S.; Kapusta, I. Identification of a biostimulating potential of an organic biomaterial based on the botanical extract from arctium lappa l. Roots. Materials 2021, 14, 4920. [Google Scholar] [CrossRef] [PubMed]
- Arif, Y.; Bajguz, A.; Hayat, S. Moringa oleifera Extract as a Natural Plant Biostimulant. J. Plant Growth Regul. 2022, 1–16. [Google Scholar] [CrossRef]
- Godlewska, K.; Ronga, D.; Michalak, I. Plant extracts-importance in sustainable agriculture. Ital. J. Agron. 2021, 16, 1851. [Google Scholar] [CrossRef]
- Hayat, S.; Ahmad, H.; Ali, M.; Hayat, K.; Khan, M.A.; Cheng, Z. Aqueous garlic extract as a plant biostimulant enhances physiology, improves crop quality and metabolite abundance, and primes the defense responses of receiver plants. Appl. Sci. 2018, 8, 1505. [Google Scholar] [CrossRef] [Green Version]
- Ismail, S.A.A.; Ganzour, S.K. Efficiency of foliar spraying with moringa leaves extract and potassium nitrate on yield and quality of strawberry in sandy soil. Int. J. Agric. Stat. Sci. 2021, 17, 383–398. [Google Scholar]
- Pestana, M.; Domingos, I.; Gama, F.; Dandlen, S.; Miguel, M.; Castro Pinto, J.; de Varennes, A.; Correia, P.J. Strawberry recovers from iron chlorosis after foliar application of a grass-clipping extract. J. Plant Nutr. Soil Sci. 2011, 174, 473–479. [Google Scholar] [CrossRef]
- Saavedra, T.; Gama, F.; Correia, P.J.; Da Silva, J.P.; Miguel, M.G.; de Varennes, A.; Pestana, M. A novel plant extract as a biostimulant to recover strawberry plants from iron chlorosis. J. Plant Nutr. 2020, 43, 2054–2066. [Google Scholar] [CrossRef]
- Palacio-Márquez, A.; Ramírez-Estrada, C.A.; Sánchez, E.; Ojeda-Barrios, D.L.; Chávez-Mendoza, C.; Sida-Arreola, J.P.; Preciado-Rangel, P. Use of biostimulant compounds in agriculture: Chitosan as a sustainable option for plant development. Not. Sci. Biol. 2022, 14, 11124. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Chaski, C.; Polyzos, N.; Tzortzakis, N.; Petropoulos, S.A. Sustainable agriculture systems in vegetable production using chitin and chitosan as plant biostimulants. Biomolecules 2021, 11, 819. [Google Scholar] [CrossRef]
- Stasinska, M.; Hawrylak-Nowak, B. Protective, Biostimulating, and Eliciting Effects of Chitosan and Its Derivatives on Crop Plants. Molecule 2022, 27, 2801. [Google Scholar] [CrossRef]
- Pichyangkura, R.; Chadchawan, S. Biostimulant activity of chitosan in horticulture. Sci. Hortic. 2015, 196, 49–65. [Google Scholar] [CrossRef]
- Li, K.; Xing, R.; Liu, S.; Li, P. Chitin and Chitosan Fragments Responsible for Plant Elicitor and Growth Stimulator. J. Agric. Food Chem. 2020, 68, 12203–12211. [Google Scholar] [CrossRef]
- González-García, Y.; Cadenas-Pliego, G.; Benavides-Mendoza, A.; Juárez-Maldonado, A. Impact of chitosan and chitosan-based nanoparticles on plants growth and development. In Role of Chitosan and Chitosan-Bases Nanomaterials in Plant Sciences; Kumar, S., Madihally, S., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 255–272. ISBN 9780323853910. [Google Scholar]
- Abdel-Mawgoud, A.M.R.; Tantawy, A.S.; El-Nemr, M.A.; Sassine, Y.N. Growth and yield responses of strawberry plants to chitosan application. Eur. J. Sci. Res. 2010, 39, 161–168. [Google Scholar]
- Akter Mukta, J.; Rahman, M.; As Sabir, A.; Gupta, D.R.; Surovy, M.Z.; Rahman, M.; Islam, M.T. Chitosan and plant probiotics application enhance growth and yield of strawberry. Biocatal. Agric. Biotechnol. 2017, 11, 9–18. [Google Scholar] [CrossRef]
- Rahman, M.; Rahman, M.; Sabir, A.A.; Mukta, J.A.; Khan, M.M.A.; Mohi-Ud-Din, M.; Miah, M.G.; Islam, M.T. Plant probiotic bacteria Bacillus and Paraburkholderia improve growth, yield and content of antioxidants in strawberry fruit. Sci. Rep. 2018, 8, 2504. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Bose, S.K.; Wang, W.; Jia, X.; Lu, H.; Yin, H. Pre-harvest treatment of chitosan oligosaccharides improved strawberry fruit quality. Int. J. Mol. Sci. 2018, 19, 2194. [Google Scholar] [CrossRef] [Green Version]
- Bhaskara Reddy, M.V.; Belkacemi, K.; Corcuff, R.; Castaigne, F.; Arul, J. Effect of pre-harvest chitosan sprays on post-harvest infection by Botrytis cinerea quality of strawberry fruit. Postharvest Biol. Technol. 2000, 20, 39–51. [Google Scholar] [CrossRef]
- Nithin, K.M.; Madaiah, D.; Shivakumar, B.S.; Kumar, M.D.; Dhananjaya, B.C. Influence of Chitosan Foliar Application on Quality and Biochemical Traits of Strawberry (Fragaria × ananassa Duch.) under Naturally Ventilated Polyhouse. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 243–250. [Google Scholar] [CrossRef]
- El-Miniawy, S.M.; Ragab, M.E.; Youssef, S.M.; Metwally, A.A. Response of Strawberry Plants to Foliar Spraying of Chitosan Response of Strawberry Plants to Foliar Spraying of Chitosan. Res. J. Agric. Biol. Sci. 2013, 9, 366–372. [Google Scholar]
- Saavedra, G.M.; Figueroa, N.E.; Poblete, L.A.; Cherian, S.; Figueroa, C.R. Effects of preharvest applications of methyl jasmonate and chitosan on postharvest decay, quality and chemical attributes of Fragaria chiloensis fruit. Food Chem. 2016, 190, 448–453. [Google Scholar] [CrossRef]
- Pilon-Smits, E.A.; Quinn, C.F.; Tapken, W.; Malagoli, M.; Schiavon, M. Physiological functions of beneficial elements. Curr. Opin. Plant Biol. 2009, 12, 267–274. [Google Scholar] [CrossRef]
- Vatansever, R.; Ozyigit, I.I.; Filiz, E. Essential and Beneficial Trace Elements in Plants, and Their Transport in Roots: A Review. Appl. Biochem. Biotechnol. 2017, 181, 464–482. [Google Scholar] [CrossRef]
- Savvas, D.; Ntatsi, G. Biostimulant activity of silicon in horticulture. Sci. Hortic. 2015, 196, 66–81. [Google Scholar] [CrossRef]
- Luyckx, M.; Hausman, J.F.; Lutts, S.; Guerriero, G. Silicon and plants: Current knowledge and technological perspectives. Front. Plant Sci. 2017, 8, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.H.; Khan, A.L.; Waqas, M.; Lee, I.J. Silicon regulates antioxidant activities of crop plants under abiotic-induced oxidative stress: A review. Front. Plant Sci. 2017, 8, 510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, R.; Awasthi, S.; Srivastava, S.; Dwivedi, S.; Pilon-Smits, E.A.H.; Dhankher, O.P.; Tripathi, R.D. Understanding selenium metabolism in plants and its role as a beneficial element. Crit. Rev. Environ. Sci. Technol. 2019, 49, 1937–1958. [Google Scholar] [CrossRef]
- Gonzali, S.; Kiferle, C.; Perata, P. Iodine biofortification of crops: Agronomic biofortification, metabolic engineering and iodine bioavailability. Curr. Opin. Biotechnol. 2017, 44, 16–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyu, S.; Wei, X.; Chen, J.; Wang, C.; Wang, X.; Pan, D. Titanium as a beneficial element for crop production. Front. Plant Sci. 2017, 8, 597. [Google Scholar] [CrossRef] [Green Version]
- Yaghubi, K.; Ghaderi, N.; Vafaee, Y.; Javadi, T. Potassium silicate alleviates deleterious effects of salinity on two strawberry cultivars grown under soilless pot culture. Sci. Hortic. 2016, 213, 87–95. [Google Scholar] [CrossRef]
- Yaghubi, K.; Vafaee, Y.; Ghaderi, N.; Javadi, T. Potassium Silicate Improves Salinity Resistant and Affects Fruit Quality in Two Strawberry Cultivars Grown Under Salt Stress. Commun. Soil Sci. Plant Anal. 2019, 50, 1439–1451. [Google Scholar] [CrossRef]
- Xiao, J.; Li, Y.; Jeong, B.R. Foliar Silicon Spray to Strawberry Plants during Summer Cutting Propagation Enhances Resistance of Transplants to High Temperature Stresses. Front. Sustain. Food Syst. 2022, 6, 938128. [Google Scholar] [CrossRef]
- Tabatabaei, S.J. Interactive effects of Si and NaCl on growth, yield, photosynthesis, and ions content in strawberry (Fragaria × ananassa var Camarosa). J. Plant Nutr. 2016, 39, 1524–1535. [Google Scholar] [CrossRef]
- Ambros, E.; Karpova, E.; Kotsupiy, O.; Zaytseva, Y.; Trofimova, E.; Novikova, T. Silicon chelates from plant waste promote in vitro shoot production and physiological changes in strawberry plantlets. Plant Cell Tissue Organ Cult. 2021, 145, 209–221. [Google Scholar] [CrossRef]
- Dehghanipoodeh, S.; Ghobadi, C.; Baninasab, B.; Gheysari, M.; Bidabadi, S.S. Effects of potassium silicate and nanosilica on quantitative and qualitative characteristics of a commercial strawberry (Fragaria × ananassa cv. ‘camarosa’). J. Plant Nutr. 2016, 39, 502–507. [Google Scholar] [CrossRef]
- Tabrizian, S.T.; Hajilou, J.; Bolandnazar, S.; Dehghan, G. Silicon Improves Strawberry Ability to Cope with Water Deficit Stress. Int. J. Hortic. Sci. Technol. 2022, 9, 213–226. [Google Scholar] [CrossRef]
- Munaretto, L.M.; Botelho, R.V.; Resende, J.T.V.; Schwarz, K.; Sato, A.J. Productivity and quality of organic strawberries pre-harvest treated with silicon. Hortic. Bras. 2018, 36, 40–46. [Google Scholar] [CrossRef]
- Valentinuzzi, F.; Cologna, K.; Pii, Y.; Mimmo, T.; Cesco, S. Assessment of silicon biofortification and its effect on the content of bioactive compounds in strawberry (Fragaria × ananassa ‘Elsanta’) fruits. Acta Hortic. 2018, 1217, 307–312. [Google Scholar] [CrossRef]
- Peris-Felipo, F.J.; Benavent-Gil, Y.; Hernández-Apaolaza, L. Silicon beneficial effects on yield, fruit quality and shelf-life of strawberries grown in different culture substrates under different iron status. Plant Physiol. Biochem. 2020, 152, 23–31. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, J.; Hu, J.; Jeong, B.R. Method of silicon application affects quality of strawberry daughter plants during cutting propagation in hydroponic substrate system. Agronomy 2020, 10, 1753. [Google Scholar] [CrossRef]
- Hajiboland, R.; Moradtalab, N.; Eshaghi, Z.; Feizy, J. Effect of silicon supplementation on growth and metabolism of strawberry plants at three developmental stages. N. Z. J. Crop Hortic. Sci. 2018, 46, 144–161. [Google Scholar] [CrossRef]
- Park, Y.G.; Muneer, S.; Kim, S.; Hwang, S.J.; Jeong, B.R. Foliar or subirrigational silicon supply modulates salt stress in strawberry during vegetative propagation. Hortic. Environ. Biotechnol. 2018, 59, 11–18. [Google Scholar] [CrossRef]
- Hajiboland, R.; Moradtalab, N.; Aliasgharzad, N.; Eshaghi, Z.; Feizy, J. Silicon influences growth and mycorrhizal responsiveness in strawberry plants. Physiol. Mol. Biol. Plants 2018, 24, 1103–1115. [Google Scholar] [CrossRef] [PubMed]
- Moradtalab, N.; Hajiboland, R.; Aliasgharzad, N.; Hartmann, T.E.; Neumann, G. Silicon and the Association with an Arbuscular-Mycorrhizal Fungus (Rhizophagus clarus) Mitigate the Adverse Effects of Drought Stress on Strawberry. Agronomy 2019, 9, 41. [Google Scholar] [CrossRef] [Green Version]
- Dehghanipoodeh, S.; Ghobadi, C.; Baninasab, B.; Gheysari, M.; Shiranibidabadi, S. Effect of Silicon on Growth and Development of Strawberry under Water Deficit Conditions. Hortic. Plant J. 2018, 4, 226–232. [Google Scholar] [CrossRef]
- Mimmo, T.; Tiziani, R.; Valentinuzzi, F.; Lucini, L.; Nicoletto, C.; Sambo, P.; Scampicchio, M.; Pii, Y.; Cesco, S. Selenium biofortification in Fragaria × ananassa: Implications on strawberry fruits quality, content of bioactive health beneficial compounds and metabolomic profile. Front. Plant Sci. 2017, 8, 1887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Qin, N.; Sun, L.; Yu, M.; Hu, W.; Qi, Z. Selenium improves physiological parameters and alleviates oxidative stress in strawberry seedlings under low-temperature stress. Int. J. Mol. Sci. 2018, 19, 1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, K.M.; Gallardo-Williams, M.T.; Benson, R.F.; Martin, D.F. Effects of selenium supplementation on four agricultural crops. J. Agric. Food Chem. 2003, 51, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.; Wu, L.; Zhang, X.; Zhang, Y.; Shan, C. Selenium improves the content of vitamin C in the fruit of strawberry by regulating the enzymes responsible for vitamin C metabolism. Plant Soil Environ. 2022, 68, 205–211. [Google Scholar] [CrossRef]
- Narváez-Ortiz, W.A.; Martínez-Hernández, M.; Fuentes-Lara, L.O.; Benavides-Mendoza, A.; Valenzuela-García, J.R.; González-Fuentes, J.A. Effect of selenium application on mineral macro-and micronutrients and antioxidant status in strawberries. J. Appl. Bot. Food Qual. 2018, 91, 321–331. [Google Scholar] [CrossRef]
- Antoniou, O.; Chrysargyris, A.; Xylia, P.; Tzortzakis, N. Effects of selenium and/or arbuscular mycorrhizal fungal inoculation on strawberry grown in hydroponic trial. Agronomy 2021, 11, 721. [Google Scholar] [CrossRef]
- Budke, C.; thor Straten, S.; Mühling, K.H.; Broll, G.; Daum, D. Iodine biofortification of field-grown strawberries—Approaches and their limitations. Sci. Hortic. 2020, 269, 109317. [Google Scholar] [CrossRef]
- Medrano-Macías, J.; López-Caltzontzit, M.G.; Rivas-Martínez, E.N.; Narváez-Ortiz, W.A.; Benavides-Mendoza, A.; Martínez-Lagunes, P. Enhancement to salt stress tolerance in strawberry plants by iodine products application. Agronomy 2021, 11, 602. [Google Scholar] [CrossRef]
- Li, R.; Liu, H.P.; Hong, C.L.; Dai, Z.X.; Liu, J.W.; Zhou, J.; Hu, C.Q.; Weng, H.X. Iodide and iodate effects on the growth and fruit quality of strawberry. J. Sci. Food Agric. 2017, 97, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Paszt, L.S.; Sumorok, B.; Malusá, E.; Gluszek, S.; Derkowska, E. the Influence of Bioproducts on Root Growthand Mycorrhizaloccurrence in the Rhizosphere of Strawberryplants ‘Elsanta’. J. Fruit Ornam. Plant Res. 2011, 19, 13–34. [Google Scholar]
- Choi, H.G.; Moon, B.Y.; Bekhzod, K.; Park, K.S.; Kwon, J.K.; Lee, J.H.; Cho, M.W.; Kang, N.J. Effects of foliar fertilization containing titanium dioxide on growth, yield and quality of strawberries during cultivation. Hortic. Environ. Biotechnol. 2015, 56, 575–581. [Google Scholar] [CrossRef]
- Skupień, K.; Oszmiański, J. Influence of Titanium Treatment on Antioxidants Content and Antioxidant Activity of Strawberries. Acta Sci. Pol. Technol. Aliment. 2007, 6, 83–94. [Google Scholar]
- Usman, M.; Farooq, M.; Wakeel, A.; Nawaz, A.; Cheema, S.A.; ur Rehman, H.; Ashraf, I.; Sanaullah, M. Nanotechnology in agriculture: Current status, challenges and future opportunities. Sci. Total Environ. 2020, 721, 137778. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Skalicky, M.; Brestic, M.; Mahari, S.; Kerry, R.G.; Maitra, S.; Sarkar, S.; Saha, S.; Bhadra, P.; Popov, M.; et al. Application of Nanomaterials to Ensure Quality and Nutritional Safety of Food. J. Nanomater. 2021, 2021, 9336082. [Google Scholar] [CrossRef]
- Benavides-Mendoza, A.; Gonzalez-Moscoso, M.; Ojeda-Barrios, D.L.; Fuentes-Lara, L.O. Biostimulation and Toxicity: Two Levels of Action of Nanomaterials in Plants. In Nanotechnology in Plant Growth Promotion and Protection: Recent Advances and Impacts; Ingle, A., Ed.; John Wiley & Sons Ltd.: Chichester, UK, 2021; pp. 283–303. ISBN 9781119745853. [Google Scholar]
- González-Morales, S.; Cárdenas-Atayde, P.A.; Garza-Alonso, C.A.; Robledo-Olivo, A.; Benavides-Mendoza, A. Plant Biostimulation with Nanomaterials: A Physiological and Molecular Standpoint Susana. In Inorganic Nanopesticides and Nanofertilizers; Fraceto, L.F., Pereira de Carvalho, H.W., De Lima, R., Ghoshal, S., Santaella, C., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2022; pp. 153–185. ISBN 9783030941543. [Google Scholar]
- Juárez-Maldonado, A.; Tortella, G.; Rubilar, O.; Fincheira, P.; Benavides-Mendoza, A. Biostimulation and toxicity: The magnitude of the impact of nanomaterials in microorganisms and plants. J. Adv. Res. 2021, 31, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, S.M.; Abdelrahman, M.; Hosseini, M.S.; Hoveizeh, N.F.; Tran, L.S.P. Alleviation of the effect of salinity on growth and yield of strawberry by foliar spray of selenium-nanoparticles. Environ. Pollut. 2019, 253, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Luksiene, Z.; Rasiukeviciute, N.; Zudyte, B.; Uselis, N. Innovative approach to sunlight activated biofungicides for strawberry crop protection: ZnO nanoparticles. J. Photochem. Photobiol. B Biol. 2020, 203, 111656. [Google Scholar] [CrossRef]
- Saini, S.; Kumar, P.; Sharma, N.C.; Sharma, N.; Balachandar, D. Nano-enabled Zn fertilization against conventional Zn analogues in strawberry (Fragaria × ananassa Duch.). Sci. Hortic. 2021, 282, 110016. [Google Scholar] [CrossRef]
- Kumar, U.J.; Bahadur, V.; Prasad, V.M.; Mishra, S.; Shukla, P.K. Effect of Different Concentrations of Iron Oxide and Zinc Oxide Nanoparticles on Growth and Yield of Strawberry (Fragaria × ananassa Duch) cv. Chandler. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 2440–2445. [Google Scholar] [CrossRef]
- Mahmood, M.M.; Al-Dulaimy, A.F. Response of Strawberry CV. Festival to Culture Media and Foliar Application of Nano and Normal Micronutrients. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Anbar, Iraq, 17–18 November 2021; Volume 904. [Google Scholar] [CrossRef]
- Dai, Y.; Chen, F.; Yue, L.; Li, T.; Jiang, Z.; Xu, Z.; Wang, Z.; Xing, B. Uptake, Transport, and Transformation of CeO2 Nanoparticles by Strawberry and Their Impact on the Rhizosphere Bacterial Community. ACS Sustain. Chem. Eng. 2020, 8, 4792–4800. [Google Scholar] [CrossRef]
- Dai, Y.; Li, T.; Wang, Z.; Xing, B. Physiological and proteomic analyses reveal the effect of CeO2 nanoparticles on strawberry reproductive system and fruit quality. Sci. Total Environ. 2022, 814, 152494. [Google Scholar] [CrossRef] [PubMed]
- akbar Mozafari, A.; Havas, F.; Ghaderi, N. Application of iron nanoparticles and salicylic acid in in vitro culture of strawberries (Fragaria × ananassa Duch.) to cope with drought stress. Plant Cell Tissue Organ Cult. 2018, 132, 511–523. [Google Scholar] [CrossRef]
- Mozafari, A.; Dedejani, S.; Ghaderi, N. Positive responses of strawberry (Fragaria × ananassa Duch.) explants to salicylic and iron nanoparticle application under salinity conditions. Plant Cell Tissue Organ Cult. 2018, 134, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Tung, H.T.; Thuong, T.T.; Cuong, D.M.; Luan, V.Q.; Hien, V.T.; Hieu, T.; Nam, N.B.; Phuong, H.T.N.; Van The Vinh, B.; Khai, H.D.; et al. Silver nanoparticles improved explant disinfection, in vitro growth, runner formation and limited ethylene accumulation during micropropagation of strawberry (Fragaria × ananassa). Plant Cell Tissue Organ Cult. 2021, 145, 393–403. [Google Scholar] [CrossRef]
- Soleymanzadeh, R.; Iranbakhsh, A.; Habibi, G.; Ardebili, Z.O. Selenium nanoparticle protected strawberry against salt stress through modifications in salicylic acid, ion homeostasis, antioxidant machinery, and photosynthesis performance. Acta Biol. Cracoviensia Ser. Bot. 2020, 62, 33–42. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and silica nanostructure-based recovery of strawberry plants subjected to drought stress. Sci. Rep. 2020, 10, 17672. [Google Scholar] [CrossRef]
- Zakaria, S.; Ragab, M.E.; EL-Yazied, A.; Rageh, M.; Farroh, K.; Salaheldin, T. Improving quality and storability of strawberries using preharvest calcium nanoparticles application. Middle East J. Agric. Res. 2018, 07, 1023–1040. [Google Scholar]
- Soleymanzadeh, R.; Iranbakhsh, A.; Habibi, G.; Ardebili, Z.O. Soil supplementation with silicon nanoparticles to alleviate toxicity signs of salinity in strawberry. Iran. J. Plant Physiol. 2022, 12, 4099–4109. [Google Scholar] [CrossRef]
- Avestan, S.; Ghasemnezhad, M.; Esfahani, M.; Byrt, C.S. Application of nano-silicon dioxide improves salt stress tolerance in strawberry plants. Agronomy 2019, 9, 246. [Google Scholar] [CrossRef]
- Moradi, P.; Vafaee, Y.; Mozafari, A.A.; Tahir, N.A.-R. Silicon Nanoparticles and Methyl Jasmonate Improve Physiological Response and Increase Expression of Stress-related Genes in Strawberry cv. Paros under Salinity Stress. Silicon 2022, 14, 10559–10569. [Google Scholar] [CrossRef]
- Rahman, M.H.; Hasan, M.N.; Khan, M.Z.H. Study on different nano fertilizers influencing the growth, proximate composition and antioxidant properties of strawberry fruits. J. Agric. Food Res. 2021, 6, 100246. [Google Scholar] [CrossRef]
- Farouk, F.; Kabil, F. Zinc oxide chitosan nano-composite membrane for enhancing transplants production in strawberry nurseries via targeting chitin elicitor receptor kinase. Int. Nano Lett. 2022, 12, 301–312. [Google Scholar] [CrossRef]
- Azim, K.; Soudi, B.; Boukhari, S.; Perissol, C.; Roussos, S.; Thami Alami, I. Composting parameters and compost quality: A literature review. Org. Agric. 2018, 8, 141–158. [Google Scholar] [CrossRef]
- Ali, U.; Sajid, N.; Khalid, A.; Riaz, L.; Rabbani, M.; Syed, J.; Malik, R. A Review on Vermicomposting of Organic Wastes Usman. Environ. Prog. Sustain. Energy 2015, 34, 1050–1062. [Google Scholar] [CrossRef]
- Martínez-Blanco, J.; Lazcano, C.; Christensen, T.H.; Muñoz, P.; Rieradevall, J.; Møller, J.; Antón, A.; Boldrin, A. Compost benefits for agriculture evaluated by life cycle assessment. A review. Agron. Sustain. Dev. 2013, 33, 721–732. [Google Scholar] [CrossRef] [Green Version]
- Agegnehu, G.; Srivastava, A.K.; Bird, M.I. The role of biochar and biochar-compost in improving soil quality and crop performance: A review. Appl. Soil Ecol. 2017, 119, 156–170. [Google Scholar] [CrossRef]
- Islam, M.K.; Yaseen, T.; Traversa, A.; Ben Kheder, M.; Brunetti, G.; Cocozza, C. Effects of the main extraction parameters on chemical and microbial characteristics of compost tea. Waste Manag. 2016, 52, 62–68. [Google Scholar] [CrossRef]
- Wang, S.Y.; Lin, H.S. Compost as a Soil Supplement Increases the Level of Antioxidant Compounds and Oxygen Radical Absorbance Capacity in Strawberries. J. Agric. Food Chem. 2003, 51, 6844–6850. [Google Scholar] [CrossRef] [PubMed]
- Pokhrel, B.; Laursen, K.H.; Petersen, K.K. Yield, Quality, and Nutrient Concentrations of Strawberry (Fragaria × ananassa Duch. cv. Sonata) Grown with Different Organic Fertilizer Strategies. J. Agric. Food Chem. 2015, 63, 5578–5586. [Google Scholar] [CrossRef] [PubMed]
- Sayğı, H. Effects of Organic Fertilizer Application on Strawberry (Fragaria vesca L.) Cultivation. Agronomy 2022, 12, 1233. [Google Scholar] [CrossRef]
- Hargreaves, J.; Adl, S.; Warman, P.; Rupasinghe, V. The effects of organic and conventional nutrient amendments on strawberry cultivation: Fruit yield and quality. J. Sci. Food Agric. 2008, 88, 2669–2675. [Google Scholar] [CrossRef]
- Welke, S. The Effect of Compost Extract on the Yield of Strawberries and the Severity of Botrytis cinerea. J. Sustain. Agric. 2005, 25, 57–68. [Google Scholar] [CrossRef]
- Khalid, S.; Qureshi, K.M.; Hafiz, I.A.; Khan, K.S.; Qureshi, U.S. Effect of organic amendments on vegetative growth, fruit and yield quality of strawberry. Pak. J. Agric. Resour. 2013, 26, 104–112. [Google Scholar]
- Hammad, S.; Elzehery, T.; Ramadan, A. Influence of compost, effective microorganisms (EM) and potassium on strawberry production in sandy soils. Acta Hortic. 2014, 1049, 407–414. [Google Scholar] [CrossRef]
- Song, Z.; Massart, S.; Yan, D.; Cheng, H.; Eck, M.; Berhal, C.; Ouyang, C.; Li, Y.; Wang, Q.; Cao, A. Composted chicken manure for anaerobic soil disinfestation increased the strawberry yield and shifted the soil microbial communities. Sustainability 2020, 12, 6313. [Google Scholar] [CrossRef]
- Kilic, N.; Burgut, A.; Gündesli, M.A.; Nogay, G.; Ercisli, S.; Kafkas, N.E.; Ekiert, H.; Elansary, H.O.; Szopa, A. The effect of organic, inorganic fertilizers and their combinations on fruit quality parameters in strawberry. Horticulturae 2021, 7, 354. [Google Scholar] [CrossRef]
- Alluqmani, S.M.; Alabdallah, N.M. Dry waste of red tea leaves and rose petals confer salinity stress tolerance in strawberry plants via modulation of growth and physiology. J. Saudi Soc. Agric. Sci. 2022; in press. [Google Scholar] [CrossRef]
- Arancon, N.Q.; Edwards, C.A.; Bierman, P.; Welch, C.; Metzger, J.D. Influences of vermicomposts on field strawberries: Effects on growth and yields. Bioresour. Technol. 2004, 93, 145–153. [Google Scholar] [CrossRef]
- Zuo, Y.; Zhang, J.; Zhao, R.; Dai, H.; Zhang, Z. Application of vermicompost improves strawberry growth and quality through increased photosynthesis rate, free radical scavenging and soil enzymatic activity. Sci. Hortic. 2018, 233, 132–140. [Google Scholar] [CrossRef]
- Singh, S.R.; Zargar, M.Y.; Singh, U.; Ishaq, M. Influence of bio-inoculants and inorganic fertilizers on yield, nutrient balance, microbial dynamics and quality of strawberry (Fragaria × ananassa) under rainfed conditions of Kashmir valley. Indian J. Agric. Sci. 2010, 80, 275–281. [Google Scholar]
- Cabilovski, R.; Manojlovic, M.; Bogdanovic, D.; Magazin, N.; Keserovic, Z.; Sitaula, B.K. Mulch type and application of manure and composts in strawberry (Fragaria × ananassa Duch.) production: Impact on soil fertility and yield. Zemdirbyste 2014, 101, 67–74. [Google Scholar] [CrossRef]
- Negi, Y.K.; Sajwan, P.; Uniyal, S.; Mishra, A.C. Enhancement in yield and nutritive qualities of strawberry fruits by the application of organic manures and biofertilizers. Sci. Hortic. 2021, 283, 110038. [Google Scholar] [CrossRef]
- Yuan, P.; Wang, J.; Pan, Y.; Shen, B.; Wu, C. Review of biochar for the management of contaminated soil Preparation, application and prospect. Sci. Total Environ. 2019, 659, 473–490. [Google Scholar] [CrossRef] [PubMed]
- Tomczyk, A.; Sokołowska, Z.; Boguta, P. Biochar physicochemical properties: Pyrolysis temperature and feedstock kind effects. Rev. Environ. Sci. Biotechnol. 2020, 19, 191–215. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, C.J.; Fitzgerald, J.D.; Hipps, N.A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: A review. Plant Soil 2010, 337, 1–18. [Google Scholar] [CrossRef]
- Palansooriya, K.N.; Ok, Y.S.; Awad, Y.M.; Lee, S.S.; Sung, J.K.; Koutsospyros, A.; Moon, D.H. Impacts of biochar application on upland agriculture: A review. J. Environ. Manag. 2019, 234, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Jiang, P.; Zhang, H.; Yuan, W. Biochar production and applications in agro and forestry systems: A review. Sci. Total Environ. 2020, 723, 137775. [Google Scholar] [CrossRef] [PubMed]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 871, 1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harel, Y.M.; Kolton, M.; Elad, Y.; Dalia, R.-D.; Cytryn, E.; Ezra, D.; Borenstein, M.; Shulchani, R.; Graber, E.R. Induced systemic resistance in strawberry (Fragaria × ananassa) to powdery mildew using various control agents. IOBC/wprs Bull 2011, 71, 47–51. [Google Scholar]
- De Tender, C.; Vandecasteele, B.; Verstraeten, B.; Ommeslag, S.; Kyndt, T.; Debode, J. Biochar-Enhanced Resistance to Botrytis cinerea in Strawberry Fruits (But Not Leaves) Is Associated with Changes in the Rhizosphere Microbiome. Front. Plant Sci. 2021, 12, 479. [Google Scholar] [CrossRef]
- De Tender, C.A.; Debode, J.; Vandecasteele, B.; D’Hose, T.; Cremelie, P.; Haegeman, A.; Ruttink, T.; Dawyndt, P.; Maes, M. Biological, physicochemical and plant health responses in lettuce and strawberry in soil or peat amended with biochar. Appl. Soil Ecol. 2016, 107, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Koron, D.; Lavrič, L.; Someus, E. Comparison of animal bone biochar and plant-based biochar in strawberry production. Acta Hortic. 2018, 1217, 313–315. [Google Scholar] [CrossRef]
- Fang, L.; Ju, W.; Yang, C.; Jin, X.; Liu, D.; Li, M.; Yu, J.; Zhao, W.; Zhang, C. Exogenous application of signaling molecules to enhance the resistance of legume-rhizobium symbiosis in Pb/Cd-contaminated soils. Environ. Pollut. 2020, 265, 114744. [Google Scholar] [CrossRef] [PubMed]
- Beavers, A.; Koether, M.; McElroy, T.; Greipsson, S. Effects of exogenous application of plant growth regulators (SNP and GA3) on phytoextraction by switchgrass (Panicum virgatum L.) grown in lead (Pb) contaminated soil. Sustainability 2021, 13, 10866. [Google Scholar] [CrossRef]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef]
- El-Banna, M.; Abdelaal, K. Response of Strawberry Plants Grown in the Hydroponic System to Pretreatment with H2O2 before Exposure to Salinity Stress. J. Plant Prod. 2018, 9, 989–1001. [Google Scholar] [CrossRef]
- Jamali, B.; Eshghi, S.; Kholdebarin, B. Response of strawberry “Selva” plants on foliar application of sodium nitroprusside (nitric oxide donor) under saline conditions. J. Hortic. Res. 2014, 22, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Jamali, B.; Eshghi, S.; Tafazoli, E. Mineral composition of ‘Selva’ strawberry as affected by time of application of nitric oxide under saline conditions. Hortic. Environ. Biotechnol. 2015, 56, 273–279. [Google Scholar] [CrossRef]
- Kaya, C.; Akram, N.A.; Ashraf, M. Influence of exogenously applied nitric oxide on strawberry (Fragaria × ananassa) plants grown under iron deficiency and/or saline stress. Physiol. Plant. 2019, 165, 247–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eshghi, S.; Jamali, B.; Rowshan, V. Headspace Analysis of Aroma Composition and Quality Changes of Selva Strawberry (Fragaria × ananassa Duch.), Fruits as Influenced by Salinity Stress and Application Timing of Nitric Oxide. Anal. Chem. Lett. 2014, 4, 178–189. [Google Scholar] [CrossRef]
- Manafi, H.; Baninasab, B.; Gholami, M.; Talebi, M. Nitric oxide induced thermotolerance in strawberry plants by activation of antioxidant systems and transcriptional regulation of heat shock proteins. J. Hortic. Sci. Biotechnol. 2021, 96, 783–796. [Google Scholar] [CrossRef]
- Jamali, B.; Eshghi, S. Application timing of nitric oxide ameliorates on deleterious effects of salinity on growth and fruit quality of strawberry cv. “Selva”. J. Berry Res. 2014, 4, 137–145. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M. The mechanism of hydrogen sulfide mitigation of iron deficiency-induced chlorosis in strawberry (Fragaria × ananassa) plants. Protoplasma 2019, 256, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Christou, A.; Filippou, P.; Manganaris, G.A.; Fotopoulos, V. Sodium hydrosulfide induces systemic thermotolerance to strawberry plants through transcriptional regulation of heat shock proteins and aquaporin. BMC Plant Biol. 2014, 14, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Li, Y.; Liu, Y.; Kang, D.I.; Wei, H.; Jeong, B.R. Hydrogen Sulfide Affects the Root Development of Strawberry during Plug Transplant Production. Agriculture 2020, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Bahmanbiglo, F.A.; Eshghi, S. The effect of hydrogen sulfide on growth, yield and biochemical responses of strawberry (Fragaria × ananassa cv. Paros) leaves under alkalinity stress. Sci. Hortic. 2021, 282, 110013. [Google Scholar] [CrossRef]
- Christou, A.; Manganaris, G.A.; Papadopoulos, I.; Fotopoulos, V. Hydrogen sulfide induces systemic tolerance to salinity and non-ionic osmotic stress in strawberry plants through modification of reactive species biosynthesis and transcriptional regulation of multiple defence pathways. J. Exp. Bot. 2013, 64, 1953–1966. [Google Scholar] [CrossRef] [PubMed]
- Aamir, M.; Rai, K.K.; Zehra, A.; Dubey, M.K.; Kumar, S.; Shukla, V.; Upadhyay, R.S. Microbial Bioformulation-Based Plant Biostimulants: A Plausible Approach toward Next Generation of Sustainable Agriculture. In Microbial Endophytes; Kumar, A., Radhakrishnan, E.K., Eds.; Elsevier Inc.: Duxford, UK, 2020; pp. 195–225. ISBN 9780128196540. [Google Scholar]
- De Pascale, S.; Rouphael, Y.; Colla, G. Plant biostimulants: Innovative tool for enhancing plant nutrition in organic farming. Eur. J. Hortic. Sci. 2017, 82, 277–285. [Google Scholar] [CrossRef]
- Hamid, B.; Zaman, M.; Farooq, S.; Fatima, S.; Sayyed, R.Z.; Baba, Z.A.; Sheikh, T.A.; Reddy, M.S.; El Enshasy, H.; Gafur, A.; et al. Bacterial plant biostimulants: A sustainable way towards improving growth, productivity, and health of crops. Sustainability 2021, 13, 2856. [Google Scholar] [CrossRef]
- Ruzzi, M.; Aroca, R. Plant growth-promoting rhizobacteria act as biostimulants in horticulture. Sci. Hortic. 2015, 196, 124–134. [Google Scholar] [CrossRef]
- Kumari, B.; Mallick, M.A.; Solanki, M.K.; Solanki, A.C. Plant Growth-Promoting Rhizobacteria (PGPR): Modern Prospects for Sustainable Agriculture. In Plant Health Under Biotic Stress; Ansario, R., Mahmood, I., Eds.; Springer: Singapore, 2019; pp. 107–127. ISBN 9789811360398. [Google Scholar]
- Ipek, M.; Pirlak, L.; Esitken, A.; Figen Dönmez, M.; Turan, M.; Sahin, F. Plant Growth-Promoting Rhizobacteria (Pgpr) Increase Yield, Growth and Nutrition of Strawberry under High-Calcareous Soil Conditions. J. Plant Nutr. 2014, 37, 990–1001. [Google Scholar] [CrossRef]
- Kurokura, T.; Hiraide, S.; Shimamura, Y.; Yamane, K. PGPR improves yield of strawberry species under less-fertilized conditions. Environ. Control Biol. 2017, 55, 121–128. [Google Scholar] [CrossRef]
- Kumar, P.; Sharma, N.; Sharma, S.; Gupta, R. Rhizosphere stochiometry, fruit yield, quality attributes and growth response to PGPR transplant amendments in strawberry (Fragaria × ananassa Duch.) growing on solarized soils. Sci. Hortic. 2020, 265, 109215. [Google Scholar] [CrossRef]
- Thakur, S. Studies on the effect of plant growth promoting rhizobacteria (PGPR) on growth, physiological parameters, yield and fruit quality of strawberry cv. chandler. J. Pharmacogn. Phytochem. 2018, 7, 383–387. [Google Scholar]
- Karlidag, H.; Yildirim, E.; Turan, M.; Pehluva, M.; Donmez, F. Plant Growth-promoting Rhizobacteria Mitigate Deleterious Effects of Salt Stress on Strawberry Plants (Fragaria × ananassa). HortScience 2013, 48, 563–567. [Google Scholar] [CrossRef] [Green Version]
- Pirlak, L.; Köse, M. Effects of plant growth promoting rhizobacteria on yield and some fruit properties of strawberry. J. Plant Nutr. 2009, 32, 1173–1184. [Google Scholar] [CrossRef]
- Erturk, Y.; Ercisli, S.; Cakmakci, R. Yield and growth response of strawberry to plant growth-promoting Rhizobacteria inoculation. J. Plant Nutr. 2012, 35, 817–826. [Google Scholar] [CrossRef]
- Anuradh; Goyal, R.K.; Sindhu, S.S. Response of strawberry (Fragaria × ananassa Duch.) to pgpr inoculation. Bangladesh J. Bot. 2020, 49, 1071–1076. [Google Scholar] [CrossRef]
- PeŠaković, M.; Karaklajić-Stajić, Ž.; Milenković, S.; Mitrović, O. Biofertilizer affecting yield related characteristics of strawberry (Fragaria × ananassa Duch.) and soil micro-organisms. Sci. Hortic. 2013, 150, 238–243. [Google Scholar] [CrossRef]
- Arikan, Ş.; İpek, M.; Eşitken, A.; Pırlak, L.; Dönmez, M.F.; Turan, M. Plant growth promoting rhizobacteria mitigate deleterious combined effects of salinity and lime in soil in strawberry plants. J. Plant Nutr. 2020, 43, 2028–2039. [Google Scholar] [CrossRef]
- Kitir, N.; Gunes, A.; Turan, M.; Yildirim, E.; Topcuoglu, B.; Turker, M.; Ozlu, E.; Karaman, M.R.; Fırıldak, G. Bio-Boron Fertilizer Applications Affect Amino Acid and Organic Acid Content and Physiological Properties of Strawberry Plant. Erwerbs-Obstbau 2019, 61, 129–137. [Google Scholar] [CrossRef]
- Esitken, A.; Yildiz, H.E.; Ercisli, S.; Figen Donmez, M.; Turan, M.; Gunes, A. Effects of plant growth promoting bacteria (PGPB) on yield, growth and nutrient contents of organically grown strawberry. Sci. Hortic. 2010, 124, 62–66. [Google Scholar] [CrossRef]
- Vicente-Hernández, A.; Salgado-Garciglia, R.; Valencia-Cantero, E.; Ramírez-Ordorica, A.; Hernández-García, A.; García-Juárez, P.; Macías-Rodríguez, L. Bacillus methylotrophicus M4-96 Stimulates the Growth of Strawberry (Fragaria × ananassa ‘Aromas’) Plants In Vitro and Slows Botrytis cinerea Infection by Two Different Methods of Interaction. J. Plant Growth Regul. 2019, 38, 765–777. [Google Scholar] [CrossRef]
- Gunes, A.; Turan, M.; Kitir, N.; Tufenkci, M.S.; Cimrin, K.M.; Yildirim, E.; Ercisli, S. Auswirkungen von Bio-Bor Düngeanwendungen auf Fruchtertrag, antioxidative Enzym-Aktivität und Frostschäden bei Erdbeeren. Erwerbs-Obstbau 2016, 58, 177–184. [Google Scholar] [CrossRef]
- Mei, C.; Amaradasa, B.S.; Chretien, R.L.; Liu, D.; Snead, G.; Samtani, J.B.; Lowman, S. A potential application of endophytic bacteria in strawberry production. Horticulturae 2021, 7, 504. [Google Scholar] [CrossRef]
- Hernández-Soberano, C.; Ruíz-Herrera, L.F.; Valencia-Cantero, E. Endophytic bacteria Arthrobacter agilis UMCV2 and Bacillus methylotrophicus M4-96 stimulate achene germination, in vitro growth, and greenhouse yield of strawberry (Fragaria × ananassa). Sci. Hortic. 2020, 261, 109005. [Google Scholar] [CrossRef]
- de Andrade, F.M.; de Assis Pereira, T.; Souza, T.P.; Guimarães, P.H.S.; Martins, A.D.; Schwan, R.F.; Pasqual, M.; Dória, J. Beneficial effects of inoculation of growth-promoting bacteria in strawberry. Microbiol. Res. 2019, 223–225, 120–128. [Google Scholar] [CrossRef]
- Pedraza, R.O.; Motok, J.; Salazar, S.M.; Ragout, A.L.; Mentel, M.I.; Tortora, M.L.; Guerrero-Molina, M.F.; Winik, B.C.; Díaz-Ricci, J.C. Growth-promotion of strawberry plants inoculated with Azospirillum brasilense. World J. Microbiol. Biotechnol. 2010, 26, 265–272. [Google Scholar] [CrossRef]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted interactions between endophytes and plant: Developments and Prospects. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Giovannini, L.; Palla, M.; Agnolucci, M.; Avio, L.; Sbrana, C.; Turrini, A.; Giovannetti, M. Arbuscular mycorrhizal fungi and associated microbiota as plant biostimulants: Research strategies for the selection of the best performing inocula. Agronomy 2020, 10, 108. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; De Pascale, S.; Bonini, P.; Colla, G. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Mikiciuk, G.; Sas-Paszt, L.; Mikiciuk, M.; Derkowska, E.; Trzciński, P.; Głuszek, S.; Lisek, A.; Wera-Bryl, S.; Rudnicka, J. Mycorrhizal frequency, physiological parameters, and yield of strawberry plants inoculated with endomycorrhizal fungi and rhizosphere bacteria. Mycorrhiza 2019, 29, 489–501. [Google Scholar] [CrossRef] [PubMed]
- ichi Matsubara, Y.; Ishigaki, T.; Koshikawa, K. Changes in free amino acid concentrations in mycorrhizal strawberry plants. Sci. Hortic. 2009, 119, 392–396. [Google Scholar] [CrossRef]
- Li, Y.; Yanagi, A.; Miyawaki, Y.; Okada, T.; Matsubara, Y.I. Disease tolerance and changes in antioxidative abilities in mycorrhizal strawberry plants. J. Jpn. Soc. Hortic. Sci. 2010, 79, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Boyer, L.R.; Feng, W.; Gulbis, N.; Hajdu, K.; Harrison, R.J.; Jeffries, P.; Xu, X. The use of arbuscular mycorrhizal fungi to improve strawberry production in coir substrate. Front. Plant Sci. 2016, 7, 1237. [Google Scholar] [CrossRef] [Green Version]
- Castellanos-Morales, V.; Villegas, J.; Wendelin, S.; Vierheilig, H.; Eder, R.; Cárdenas-Navarro, R. Root colonisation by the arbuscular mycorrhizal fungus Glomus intraradices alters the quality of strawberry fruits (Fragaria × ananassa Duch.) at different nitrogen levels. J. Sci. Food Agric. 2010, 90, 1774–1782. [Google Scholar] [CrossRef]
- Stewart, L.I.; Hamel, C.; Hogue, R.; Moutoglis, P. Response of strawberry to inoculation with arbuscular mycorrhizal fungi under very high soil phosphorus conditions. Mycorrhiza 2005, 15, 612–619. [Google Scholar] [CrossRef]
- Boyer, L.R.; Brain, P.; Xu, X.M.; Jeffries, P. Inoculation of drought-stressed strawberry with a mixed inoculum of two arbuscular mycorrhizal fungi: Effects on population dynamics of fungal species in roots and consequential plant tolerance to water deficiency. Mycorrhiza 2015, 25, 215–227. [Google Scholar] [CrossRef]
- Lingua, G.; Bona, E.; Manassero, P.; Marsano, F.; Todeschini, V.; Cantamessa, S.; Copetta, A.; D’Agostino, G.; Gamalero, E.; Berta, G. Arbuscular mycorrhizal fungi and plant growth-promoting pseudomonads increases anthocyanin concentration in strawberry fruits (Fragaria × ananassa var. Selva) in conditions of reduced fertilization. Int. J. Mol. Sci. 2013, 14, 16207–16225. [Google Scholar] [CrossRef]
- Gryndler, M.; Vosátka, M.; Hrŝelová, H.; Catská, V.; Chvátalová, I.; Jansa, J. Effect of dual inoculation with arbuscular mycorrhizal fungi and bacteria on growth and mineral nutrition of strawberry. J. Plant Nutr. 2002, 25, 1341–1358. [Google Scholar] [CrossRef]
- Fan, L.; Dalpé, Y.; Fang, C.; Dubé, C.; Khanizadeh, S. Influence of arbuscular mycorrhizae on biomass and root morphology of selected strawberry cultivars under salt stress. Botany 2011, 89, 397–403. [Google Scholar] [CrossRef]
- Chiomento, J.L.T.; De Nardi, F.S.; Filippi, D.; Trentin, T.d.S.; Dornelles, A.G.; Fornari, M.; Nienow, A.A.; Calvete, E.O. Morpho-horticultural performance of strawberry cultivated on substrate with arbuscular mycorrhizal fungi and biochar. Sci. Hortic. 2021, 282, 110053. [Google Scholar] [CrossRef]
- Taylor, J.; Harrier, L.A. A comparison of development and mineral nutrition of micropropagated Fragaria × ananassa cv. Elvira (strawberry) when colonised by nine species of arbuscular mycorrhizal fungi. Appl. Soil Ecol. 2001, 18, 205–215. [Google Scholar] [CrossRef]
- Martinez, F.; Weiland, C.; Palencia, P. Effect of arbuscular mycorrhizal fungi on quality of strawberry fruit in soilless growing system. Acta Hortic. 2013, 1013, 493–498. [Google Scholar] [CrossRef]
- Chauhan, S.; Kumar, A.; Mangla, C.; Aggarwal, A. Response of Strawberry plant (Fragaria ananassa Duch.) to inoculation with arbuscular mycorrhizal fungi and Trichoderma viride. J. Appl. Nat. Sci. 2010, 2, 213–218. [Google Scholar] [CrossRef] [Green Version]
- Lombardi, N.; Caira, S.; Troise, A.D.; Scaloni, A.; Vitaglione, P.; Vinale, F.; Marra, R.; Salzano, A.M.; Lorito, M.; Woo, S.L. Trichoderma Applications on Strawberry Plants Modulate the Physiological Processes Positively Affecting Fruit Production and Quality. Front. Microbiol. 2020, 11, 1364. [Google Scholar] [CrossRef] [PubMed]
- Porras, M.; Barrau, C.; Romero, F. Effects of soil solarization and Trichoderma on strawberry production. Crop Prot. 2007, 26, 782–787. [Google Scholar] [CrossRef]
- Sekmen Cetinel, A.H.; Gokce, A.; Erdik, E.; Cetinel, B.; Cetinkaya, N. The effect of trichoderma citrinoviride treatment under salinity combined to rhizoctonia solani infection in strawberry (Fragaria × ananassa Duch.). Agronomy 2021, 11, 1589. [Google Scholar] [CrossRef]
- Khan, F.; Kim, N.E.; Bhujel, A.; Jaihuni, M.; Lee, D.H.; Basak, J.K.; Kim, H.T. Assessment of Combined Trichoderma-Enriched Biofertilizer and Nutrients Solutions on the Growth and Yield of Strawberry Plants. J. Biosyst. Eng. 2021, 46, 225–235. [Google Scholar] [CrossRef]
- López-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrella, A. Trichoderma as biostimulant: Exploiting the multilevel properties of a plant beneficial fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Visconti, D.; Fiorentino, N.; Cozzolino, E.; Woo, S.L.; Fagnano, M.; Rouphael, Y. Can Trichoderma-based biostimulants optimize N use efficiency and stimulate growth of leafy vegetables in greenhouse intensive cropping systems? Agronomy 2020, 10, 121. [Google Scholar] [CrossRef] [Green Version]
- Formisano, L.; Miras-Moreno, B.; Ciriello, M.; El-Nakhel, C.; Corrado, G.; Lucini, L.; Colla, G.; Rouphael, Y. Trichoderma and phosphite elicited distinctive secondary metabolite signatures in zucchini squash plants. Agronomy 2021, 11, 1205. [Google Scholar] [CrossRef]
- Fiorentino, N.; Ventorino, V.; Woo, S.L.; Pepe, O.; De Rosa, A.; Gioia, L.; Romano, I.; Lombardi, N.; Napolitano, M.; Colla, G.; et al. Trichoderma-based biostimulants modulate rhizosphere microbial populations and improve N uptake efficiency, yield, and nutritional quality of leafy vegetables. Front. Plant Sci. 2018, 9, 743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, B.J.; Rezazadeh, A.; Stafne, E.T.; Sakhanokho, H.F. Effect of Light-emitting Diodes, Ultraviolet-B, and Fluorescent Supplemental Greenhouse Lights on Strawberry Plant Growth and Response to Infection by the Anthracnose Pathogen Colletotrichum gloeosporioides. HortScience 2022, 57, 856–863. [Google Scholar] [CrossRef]
- Darko, E.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakurin, A.O.; Shchennikova, A.V.; Kamionskaya, A.M. Artificial-Light Culture in Protected Ground Plant Growing: Photosynthesis, Photomorphogenesis, and Prospects of LED Application. Russ. J. Plant Physiol. 2020, 67, 413–424. [Google Scholar] [CrossRef]
- Paik, I.; Huq, E. Plant photoreceptors: Multi-functional sensory proteins and their signaling networks. Semin. Cell Dev. Biol. 2019, 92, 114–121. [Google Scholar] [CrossRef]
- Devireddy, A.R.; Liscum, E.; Mittler, R. Phytochrome B is required for systemic stomatal responses and reactive oxygen species signaling during light stress. Plant Physiol. 2020, 184, 1563–1572. [Google Scholar] [CrossRef]
- Küpers, J.J.; Oskam, L.; Pierik, R. Photoreceptors regulate plant developmental plasticity through auxin. Plants 2020, 9, 940. [Google Scholar] [CrossRef]
- Luo, Y.; Shi, H. Direct Regulation of Phytohormone Actions by Photoreceptors. Trends Plant Sci. 2019, 24, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, K.; Dan, K.; Imamura, H.; Miyoshi, Y.; Takayama, T.; Sameshima, K.; Kitano, M.; Okimura, M. Effect of supplemental lighting from different light sources on growth and yield of strawberry. Environ. Control Biol. 2013, 51, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.G.; Moon, B.Y.; Kang, N.J. Effects of LED light on the production of strawberry during cultivation in a plastic greenhouse and in a growth chamber. Sci. Hortic. 2015, 189, 22–31. [Google Scholar] [CrossRef]
- Nhut, D.T.; Takamura, T.; Watanabe, H.; Okamoto, K.; Tanaka, M. Responses of strawberry plantlets cultured in vitro under superbright red and blue light-emitting diodes (LEDs). Plant Cell Tissue Organ Cult. 2003, 73, 43–52. [Google Scholar] [CrossRef]
- Yoshida, H.; Hikosaka, S.; Goto, E.; Takasuna, H.; Kudou, T. Effects of light quality and light period on flowering of everbearing strawberry in a closed plant production system. Acta Hortic. 2012, 956, 107–112. [Google Scholar] [CrossRef]
- Wu, C.C.; Hsu, S.T.; Chang, M.Y.; Fang, W. Effect of light environment on runner plant propagation of strawberry. Acta Hortic. 2011, 907, 297–302. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, W.; Peng, X.; Sun, B.; Wang, X.; Tang, H. Characterization of anthocyanin and proanthocyanidin biosynthesis in two strawberry genotypes during fruit development in response to different light qualities. J. Photochem. Photobiol. B Biol. 2018, 186, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, K.; Okamoto, A.; Araki, T.; Miyoshi, Y.; Dan, K.; Imamura, H.; Kitano, M.; Sameshima, K.; Okimura, M. Effect of photoperiod of supplemental lighting with light-emitting diodes on growth and yield of strawberry. Environ. Control Biol. 2014, 52, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Malekzadeh Shamsabad, M.R.; Esmaeilizadeh, M.; Roosta, H.R.; Dąbrowski, P.; Telesiński, A.; Kalaji, H.M. Supplemental light application can improve the growth and development of strawberry plants under salinity and alkalinity stress conditions. Sci. Rep. 2022, 12, 9272. [Google Scholar] [CrossRef]
- Shamsabad, M.R.M.; Esmaeilizadeh, M.; Roosta, H.R.; Dehghani, M.R.; Dąbrowski, P.; Kalaji, H.M. The effect of supplementary light on the photosynthetic apparatus of strawberry plants under salinity and alkalinity stress. Sci. Rep. 2022, 12, 13257. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Wu, W.; Hu, B. Light and temperature receptors and their convergence in plants. Biol. Plant. 2020, 64, 159–166. [Google Scholar] [CrossRef]
- Casal, J.J.; Qüesta, J.I. Light and temperature cues: Multitasking receptors and transcriptional integrators. New Phytol. 2018, 217, 1029–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saidi, Y.; Finka, A.; Goloubinoff, P. Heat perception and signalling in plants: A tortuous path to thermotolerance. New Phytol. 2011, 190, 556–565. [Google Scholar] [CrossRef]
- von Koskull-Döring, P.; Scharf, K.D.; Nover, L. The diversity of plant heat stress transcription factors. Trends Plant Sci. 2007, 12, 452–457. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Zheng, C.; Huang, Y.P.; Wang, X.L.; Luo, Z.S.; Zheng, Y. Hua Hot air treatment activates defense responses and induces resistance against Botrytis cinerea in strawberry fruit. J. Integr. Agric. 2016, 15, 2658–2665. [Google Scholar] [CrossRef] [Green Version]
- Widiastuti, A.; Yoshino, M.; Saito, H.; Maejima, K.; Zhou, S.; Odani, H.; Narisawa, K.; Hasegawa, M.; Nitta, Y.; Sato, T. Heat shock-induced resistance in strawberry against crown rot fungus Colletotrichum gloeosporioides. Physiol. Mol. Plant Pathol. 2013, 84, 86–91. [Google Scholar] [CrossRef]
- Brown, R.; Wang, H.; Dennis, M.; Slovin, J.; Turechek, W.W. The Effects of Heat Treatment on the Gene Expression of Several Heat Shock Protein Genes in Two Cultivars of Strawberry. Int. J. Fruit Sci. 2016, 16, 239–248. [Google Scholar] [CrossRef]
- Kesici, M.; Ipek, A.; Ersoy, F.; Ergin, S.; Gülen, H. Genotype-Dependent Gene Expression in Strawberry (Fragaria × ananassa) Plants Under High Temperature Stress. Biochem. Genet. 2020, 58, 848–866. [Google Scholar] [CrossRef]
- Araújo, S.d.S.; Paparella, S.; Dondi, D.; Bentivoglio, A.; Carbonera, D.; Balestrazzi, A. Physical methods for seed invigoration: Advantages and challenges in seed technology. Front. Plant Sci. 2016, 7, 646. [Google Scholar] [CrossRef] [Green Version]
- Bukhari, S.A.; Farah, N.; Mustafa, G.; Mahmood, S.; Naqvi, S.A.R. Magneto-Priming Improved Nutraceutical Potential and Antimicrobial Activity of Momordica charantia L. without Affecting Nutritive Value. Appl. Biochem. Biotechnol. 2019, 188, 878–892. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.K.; Anand, A.; Paul, V.; Dahuja, A.; Singh, A.K. Reactive oxygen species mediated improvement in vigour of static and pulsed magneto-primed cherry tomato seeds. Indian J. Plant Physiol. 2015, 20, 197–204. [Google Scholar] [CrossRef]
- Rathod, G.R.; Anand, A. Effect of seed magneto-priming on growth, yield and Na/K ratio in wheat (Triticum aestivum L.) under salt stress. Indian J. Plant Physiol. 2016, 21, 15–22. [Google Scholar] [CrossRef]
- Maffei, M.E. Magnetic field effects on plant growth, development, and evolution. Front. Plant Sci. 2014, 5, 445. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
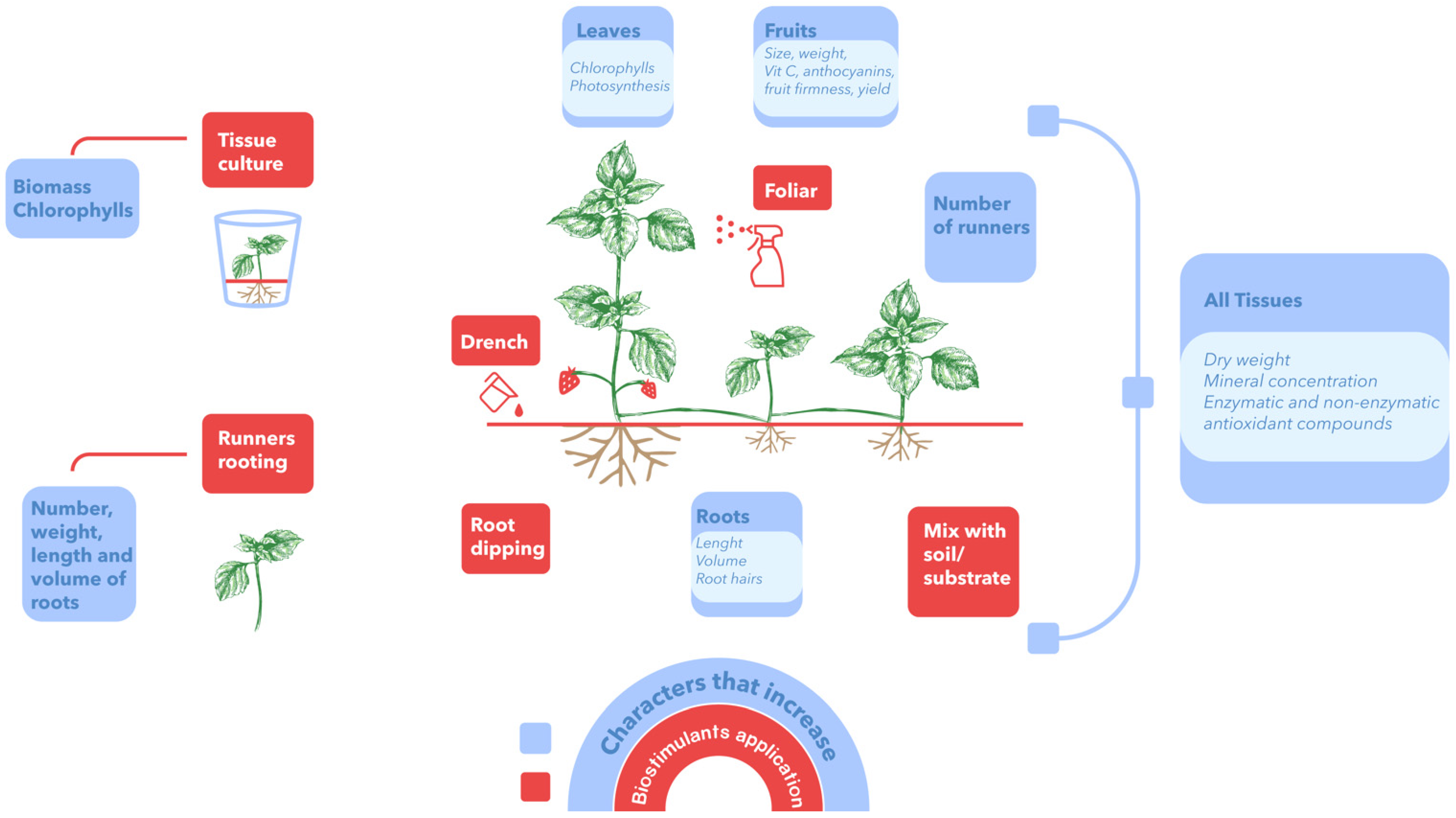
| Product | Experimental Conditions | Forms and Levels of Application | Variables That Increase | Reference |
|---|---|---|---|---|
| HA NS * | Greenhouse, pots with substrate | Foliar 0, 25, 50, and 100 mg L−1 | Fruit yield, TSS, TA, Vit. C, K, P, Ca, Mg. | [36] |
| HA from cow manure, food waste, paper waste | Greenhouse, pots with substrate | Substrate mix 0, 250, and 500 mg kg−1 of substrate | Root dry weight. | [37] |
| HA from cow manure, food waste, paper waste | Greenhouse, pots with substrate | Substrate mix 0, 250, and 500 mg kg−1 of substrate | Number of fruits. | [38] |
| HA Commercial formulation | Open field, pots with soil | Root immersion by 2 h, 0.05% | Number and length of runners, length of roots, and total biomass. | [39] |
| HA + FA Commercial formulation | Greenhouse, pots with substrate | Substrate mix, 0.06 g kg−1 | P in roots, Mn and P in leaves. | [40] |
| HA NS | Greenhouse, pots with soil | Foliar 15 and 25 mL L−1 | Biomass, length of roots, leaf area, number of runners and flowers, fruit weight, TSS, TA, and Vit. C. | [41] |
| HA NS | Open field, soil | Foliar 0, 2, and, 4 mL L−1 | N concentration in leaves, number of flowers, and fruit yield. | [42] |
| HA NS | Greenhouse, pots with soil | Foliar 100 mg L−1 | Proline concentration, phenolics, and antioxidant capacity. | [43] |
| HA Commercial formulation | Greenhouse, pots with substrate | Substrate mix 4 g HA pot−1 | Plant height, number of leaves, crowns, and roots, fresh and dry weight of leaves and roots, stomatal conductance. | [44] |
| HA + FA Extracted from vermicompost | Open field, soil conditions | Foliar 180 mg L−1 | Chlorophyll concentration and net photosynthesis. | [45] |
| HA NS | Greenhouse, soil conditions | Foliar 20 and 40 mg L−1 | Number and weight of fruits, yield per plant, leaf area, length and dry weight of shoot and root. | [46] |
| HA + FA NS | Open field, soil conditions | Drench 5 mL L−1 | TSS, TA, anthocyanins, Vit. C, phenolics. | [47] |
| HA + FA Commercial formulation | Open field, soil conditions | Drench and Foliar 2, 4, and, 6 ton ha−1 | Leaf area, biomass, chlorophyll, carotenoids, TSS, and Vit. C. | [48] |
| HA Extracted from soil | In vitro | Growing medium 1 and 5 mg dm−3 | Number and length of roots, plant weight, number and size of leaves. | [49] |
| HA Commercial formulation | Greenhouse, pots with substrate | Drench 150 and 300 mg L−1 | K concentration, chlorophyll, carbohydrates, shoot and root dry weight, leaf area, SOD, fruit number and yield. | [50] |
| HA Commercial formulation | Greenhouse, pots with substrate | Foliar 1 g L−1 | Root dry weight, Si, fruit chromaticity. | [51] |
| HA NS | Greenhouse, soil conditions | Drench and foliar 10, 20, 30, and 40 mg L−1 | Chlorophyll, N, P, K. | [52] |
| HA NS | Greenhouse, pots with soil | 2 g kg−1 soil | Plant height, leaf area, fresh weight, N, P, K. | [53] |
| HA + FA Commercial formulation | Open field, soil conditions | Drench 10 mL L−1 | Number and length of runners; number, length, and weight of roots. | [54] |
| Product | Experimental Conditions | Forms and Levels of Application | Variables That Increase | Reference |
|---|---|---|---|---|
| Porcine blood | Open field, soil conditions | Drench 0.5, 1, and 1.5 g plant−1 | Resistance to cold stress, fruit weight. | [60] |
| Fish protein concentrates | Greenhouse, pots with soil | Drench NS | Fresh and dry biomass, chlorophyll fluorescence. | [61] |
| Amino acids (Proline, Alanine, Glutamine | In vitro | Growing medium 50, 100, 150, and 200 mg L−1 | Somatic embryogenesis. | [62] |
| Porcine blood | High-tunnel, soil conditions | Drench 2.5 g L−1 | Dry weight of roots, % of flowering, fruit weight. | [63] |
| Arginine NS | Greenhouse, soil conditions | Foliar 0, 250 and 500 μM | Number of fruits, TSS, anthocyanins, phenolics, Vit. C. | [64] |
| Alfalfa protein hydrolizated | Greenhouse, pots with substrate | Foliar 3 g L−1 | Root dry weight, leaf area, Si concentration, SPAD, fruit weight, phenolics. | [51] |
| Microalga protein hydrolizated | Greenhouse, pots with substrate | Foliar 4 g L−1 | Root dry weight, Fe and Si concentration in roots, TA in fruits. | |
| Mix of amino acids | Greenhouse, pots with substrate | Foliar 3 g L−1 | TSS in fruits. | |
| Amino acids (hydroxyproline and glutamic acid), commercial formulation | Controlled environment room, pots with substrate | Foliar 228 and 319 mg L−1 | Number of flowers, number, and weight of fruits, Vit. C. | [65] |
| Hydrolyzed feather meal | Greenhouse, pots with soil | 0.10 g kg−1 soil | Indole Acetic Acid (IAA), Abscisic acid (ABA), Isopentenyl adenosine (iPA). | [66] |
| Amino acids (Glycine) | Open field, soil conditions | Drench 0.5 g L−1 | Number and length of runners, roots length. | [54] |
| Product | Experimental Conditions | Forms and Levels of Application | Variables That Increase | Reference |
|---|---|---|---|---|
| Ascophyllum nodosum, commercial extract | Greenhouse, pots with substrate | Drench 0.2, 0.4, 1.0, or 2.0 g L−1 | Number, surface area, volume, and length of roots. | [71] |
| Open field, soil conditions | Drench 2 and 4 g L−1 | Leaf area, shoot dry weight, number of fruits and yield. | ||
| Sargassum spp., commercial extract | Open field, pots with substrate | Drench 0, 2, 4, and 8 g L−1 | Mn concentration. | [72] |
| Sargassum spp., commercial extract | Open field, pots with substrate | Drench 0, 2, 4, and 8 g L−1 | Number of crowns, number and volume of fruits, yield. | [73] |
| Ascophyllum nodosum, commercial extract | Greenhouse, pots with substrate | Foliar 0.1, 0.2, and 0.3% | Phenolics and flavonoids concentration; activity of PAL and POD. More resistance to Podosphaera aphanis. | [74] |
| Seaweed extract, NS | High tunnel, soil conditions | Drench 20 g ha−1 | Concentration of N, P, K, Ca, Mg, and Mn. | [75] |
| Mix of Sargassum sp., Ascophyllum nodosum, Laminaria sp. | Open field, soil conditions | Foliar 1 and 2 mL L−1 | Plant height, number of leaves, leaf area, root dry weight, fruit weight, TSS. | [76] |
| Ascophyllum nodosum, commercial extract | Open field, soil conditions | 4.68 L ha−1 | Number of crowns, root dry weight, fruit yield. | [77] |
| Seaweed extract, NS | High tunnel, soil conditions | Foliar 1.3 g L−1 | Leaf area, fruit N concentration, fruit yield. | [78] |
| Seaweed extract, NS | High tunnel, soil conditions | Foliar 1.3 g L−1 | TSS, fructose, sucrose, and quercetin. | [79] |
| Mix of Duvillaea potatorum and Ascophyllum nodosum | Open field, soil conditions | 10 L ha−1 | Number of runners, fruit yield, roots length. | [80] |
| Seaweed extract, NS | Open field, soil | Foliar 2 and 4 mL L−1 | Leaf and root dry weight, N concentration, number of flowers, yield. | [42] |
| Ascophyllum nodosum, commercial formulation | Greenhouse, pots with substrate | Foliar 3 g L−1 | Root dry weight, leaf area, Si in roots, phenolics. | [51] |
| Spirulina spp., commercial formulation | Greenhouse, pots with substrate | Foliar 3 g L−1 | Root dry weight, Fe and Si in roots, fruit firmness and TA. | |
| Ascophyllum nodosum, commercial formulation | Greenhouse, pots with substrate | Drench 0.5 mL L−1 | Vegetative growth, chlorophyll concentration, photosynthetic rate, number, and weight of fruits. | [81] |
| Product | Experimental Conditions | Forms and Levels of Application | Variables that Increase | Reference |
|---|---|---|---|---|
| Chitosan, commercial product | Open field, soil conditions | Foliar 1, 2, 3, and 4 mL L−1 | Plant height, number of leaves, biomass, number and weight of fruits. | [96] |
| Chitosan, commercial product | Open field, soil conditions | Foliar 125, 250, 500, and 1000 mg L−1 | Leaf size, fresh and dry weight of shoot and roots, fruit weight and yield. | [97] |
| Chitosan, commercial product | Open field, soil conditions | Foliar 125, 250, 500, and 1000 mg L−1 | Anthocyanins, phenolics, flavonoids, carotenoids, antioxidant capacity. | [98] |
| Chitosan oligosaccharide, commercial formulation | Open field, soil conditions | Foliar 50 mg L−1 | Fruit firmness, TSS, Vit. C, phenolics, flavonoids, antioxidant capacity. | [99] |
| Chitosan, commercial product | Greenhouse, pots with substrate | Foliar 10 mL L−1 | Root dry weight, B and Si concentration in roots, weight, firmness, and fruit yield. | [51] |
| Chitosan, commercial product | Greenhouse, pots with substrate | Foliar 2, 4, and 6 g L−1 | Reduction of % postharvest decay, fruit firmness, citric acid. | [100] |
| Chitosan, commercial product | Greenhouse, pots with substrate | Foliar 1, 2, and 3 g L−1 | Plant height, number of leaves, leaf aera, dry biomass, fruit size, weight, and yield. | [101] |
| Chitosan, commercial product | Open field, soil conditions | 2.5 and 5 mL L−1 | Plant height, number of leaves, leaf area, root dry weight, N, P, K, fruit weight, yield. | [102] |
| Chitosan, commercial product | Open field, soil conditions | Foliar 15 g L−1 | Fruit firmness, anthocyanin concentration, phenolics and antioxidant capacity. | [103] |
| Product | Experimental Conditions | Forms and Levels of Application | Variables That Increase | Reference |
|---|---|---|---|---|
| Silicon | ||||
| K2SiO3 | Greenhouse, pots with substrate | Drench 1000 and 1500 mg L−1 | Shoot dry weight, leaf area, root volume, relative water content. | [112] |
| K2SiO3 | Greenhouse, pots with substrate | Drench 1000 and 1500 mg L−1 | Plant biomass, fruit number, TSS, TA, antioxidant activity. | [113] |
| K2SiO3 | Greenhouse, pots with substrate | Drench and Foliar 75 mg L−1 | General vegetative growth, chlorophyll, stomatal conductance, soluble sugars, CAT, APX, POD, SOD, anthocyanins. | [114] |
| Si(OH)4 | Greenhouse, pots with substrate | Drench 1 and 2 mM | Leaf number, leaf area, dry weight, photosynthetic rate, stomatal conductance. | [115] |
| Si chelate | In vitro | Growing media 2.5, 5, and 10 mg L−1 | Number and length of shoots, CAT, SOD. | [116] |
| K2SiO3 | Shade house, pots with substrate | Drench and Foliar 5, 10, and 15 mM | Shoot and root dry weight, chlorophyll, number of flowers and fruits, yield, fruit firmness. | [117] |
| Si, commercial formulation | Greenhouse, pots with substrate | Foliar 0.3 mL L−1 | Zn and Si concentration, weight of fruit, yield. | [51] |
| Na2SiO3 | Greenhouse, soil conditions | Foliar 3 and 6 mM | SOD, phenolics, flavonoids, anthocyanins. | [118] |
| SiO2 | Open field, soil conditions | Foliar 5, 10, and 15 mg L−1 | Fruit firmness and anthocyanins. | [119] |
| Na2SiO3 | Greenhouse, soilless system | Drench 50 and 100 mg L−1 | Flavonoids and Si concentration. | [120] |
| SiO4H4 | Open field, pots with substrate | Drench and Foliar 1.5 mM | Leaf area, SPAD, fruit size and weight, fructose concentration. | [121] |
| K2SiO3 | Greenhouse, pots with substrate | Drench and Foliar 75 mg L−1 | Leaf size, fresh and dry weight of shoot, Si concentration, chlorophyll fluorescence. | [122] |
| Na2SiO3 | Greenhouse, pots with substrate | Drench 3 mM | Shoot and root dry weight, net photosynthesis, relative water content, protein, phenolics. | [123] |
| K2SiO3 Na2SiO3 CaSiO3 | Greenhouse, pots with substrate | Drench and Foliar 35 and 70 mg L−1 | CAT, SOD and POD activity. | [124] |
| Na2SiO3 | Greenhouse, pots with substrate | Drench 3 mM | Shoot and root dry weight, Si, Zn, soluble sugars, soluble proteins, PAL, phenolics. | [125] |
| Na2SiO3 | Greenhouse, pots with substrate | Drench 3 mM | Shoot and root biomass, net photosynthesis, stomatal conductance, water efficiency use, CAT, SOD, POD. | [126] |
| K2SiO3 | Shade house, pots with substrate | 5, 10, and 15 mM | Root dry weight, chlorophyll fluorescence, net photosynthesis, water efficiency use. | [127] |
| Selenium | ||||
| Na2SeO4 | Greenhouse, soilless system | Nutrient solution 10 and 100 µM | Shoot fresh weight, leaf area, K, Ca, Mg in roots, TSS, fructose, sucrose. | [128] |
| Na2SeO3 | Greenhouse, pots with soil | Foliar 2.5, 5, and 10 mg L−1 | Net photosynthesis, stomatal conductance, chlorophyll, SOD, CAT, POD. | [129] |
| Se NS | Growth chamber, pots with soil | Mix with soil 40 mg kg−1 soil | Fruit weight, Se concentration. | [130] |
| Na2SeO3 | Growth chamber, pots with soil | Foliar 10, 30, and 60 mg L−1 | Number of fruits, yield, Vit. C, APX. | [131] |
| Na2SeO3 | Greenhouse, pots with substrate | Drench 2 and 4 mg L−1 | Fresh and dry weight of crown, K, Ca, Mg, Zn, Se. | [132] |
| Na2SeO4 | Greenhouse, pots with substrate | Drench 1, 5, and 10 mg L−1 | Plant biomass, phenolics, flavonoids, antioxidant capacity. | [133] |
| Iodine | ||||
| KIO3 | Greenhouse, pots with substrate | Drench 1, 2.5, and 7.5 mg L−1 | Fruit I concentration. | [134] |
| KI | Foliar 0.25, 0.75, and 1.5 mg L−1 | |||
| I-based commercial product | Greenhouse, pots with soil | Foliar 0.5 mL L−1 | Phenolics, APX, CAT, K, I concentration. | [135] |
| KIO3 | Foliar 100 µM | Fruit firmness, Vit. C, I concentration. | ||
| KI | Greenhouse, soilless system | Nutrient solution 0.25, 0.5, 1, 2.5, 5 mg L−1 | Vit. C, soluble sugars, I concentration. | [136] |
| KIO3 | Nutrient solution 0.25, 0.5, 1, 2.5, 5 mg L−1 | |||
| Titanium | ||||
| Ti, commercial product | Greenhouse, soil conditions | Soil mix 0.05% | Number of root tips, root dry weight. | [137] |
| TiO2 | Greenhouse, soil conditions | Foliar 50, 100, and 150 mg L−1 | Chlorophylls, yield, glucose, oxalic, malic, and citric acid. | [138] |
| Ti, commercial product | Open field, soil conditions | Foliar 0.02% | Phenolics, Vit. C, antioxidant capacity, anthocyanins. | [139] |
| Material/Form/Size | Experimental Conditions | Forms and Levels of Application | Variables That Increase | Reference |
|---|---|---|---|---|
| Se-NPs/spherical/10–45 nm | Greenhouse, pots with substrate | Foliar 10 and 20 mg L−1 | Root and shot dry weight, number and weight of fruits, yield, chlorophyll concentrations, POD, SOD. | [145] |
| ZnO NPs 25–50 nm | Open field, soil conditions | Foliar 7.5 × 10−3 M | Number of flowers. | [146] |
| ZnO NPs <100 nm | Open field, soil conditions | Foliar 200, 400, and 600 μg g−1 | Plant height, number of leaves, leaf area, number of runners, fruit size and yield. | [147] |
| ZnO NPs NS | Open field, soil conditions | Foliar 50, 100, and 150 mg L−1 | Plant height, number of leaves, number of fruits and yield. | [148] |
| Zn NPs NS | Greenhouse, soil conditions | Foliar 10 and 20 mg L−1 | Number, weight, and fruit yield. | [149] |
| CeO2 NPs 2–50 nm | Greenhouse, soil conditions | Drench 300, 600, 1000, and 2000 mg L−1 | Shoot and root biomass, root surface area, SPAD. | [150] |
| CeO2 NPs 2–50 nm | Greenhouse, soil conditions | 6, 20, 41, 70, and 115 mg L−1 | Phenolics, Vit. C, soluble protein, IAA, number of fruits. | [151] |
| Fe NPs NS | In vitro | Growing medium 0.8 mg L−1 | Shoot length, root dry weight, relative water content. | [152] |
| Fe NPs NS | In vitro | Growing medium 0.8 mg L−1 | Branch number, root length, plant weight. | [153] |
| FeO NPs NS | Open field, soil conditions | Foliar 50, 100, and 150 mg L−1 | Plant height, number of leaves, number of fruits and yield. | [148] |
| Fe NPs NS | Greenhouse, soil conditions | Foliar 20 and 40 mg L−1 | Number, weight, and fruit yield. | [149] |
| Ag NPs <20 nm | In vitro | Growing medium 0.2, 0.4, 0.6, 0.8, and 1 mg L−1 | Number and height of shoots, fresh and dry weight, chlorophyll concentration, number and length of roots. | [154] |
| Se-NPs/10–45 nm | Greenhouse, soil conditions | Foliar 10 and 100 μM | CAT, catechin, caffeic acid, coumaric acid, salicylic acid. | [155] |
| Se NPs 10–45 nm | Greenhouse, pots with soil | Foliar 25 mg L−1 | Root fresh weight, chlorophyll, GPX, number of leaves, water efficiency use. | [156] |
| Ca5(PO4)3(OH) NPs 20–40 nm | Open field, soil conditions | Foliar 15, 30, 60, and 120 mg L−1 | Fruit postharvest life, firmness, Vit. C. | [157] |
| SiO2 NPs 20–30 nm | Greenhouse, soil conditions | Mix with soil 0.75 and 1.5 g kg−1 | Root fresh weight, Vit. C, quercetin, proline, PAL, Ca concentration. | [158] |
| SiO2 NPs 20–30 nm | Greenhouse, pots with soil | Foliar 125 mg L−1 | Number of flowers, anthocyanins, phenolics. | [156] |
| SiO2 NPs NS | Greenhouse, pots with substrate | Drench 50 and 100 mg L−1 | Shoot and root biomass, chlorophylls, fruit yield. | [159] |
| SiO2 NPs NS | Greenhouse, pots with substrate | Drench 2 mM | Resistance to salt stress through improve membrane stability and decrease H2O2. | [160] |
| SiO2 NPs 30–35 nm | Shade house, pots with substrate | Drench and Foliar 5, 10, and 15 mM | Shoot and root dry weight, chlorophyll, number of flowers and fruits, yield, fruit firmness. | [117] |
| Nanozeolite NS | Open field, soil conditions | Mix with soil 5 g bed−1 | Length of plant, number of leaves, number and weight of fruit and yield. | [161] |
| Se/SiO2 NPs 50–80 nm | Greenhouse, pots with soil | Foliar 50 and 100 mg L−1 | Shot and root biomass, chlorophyll, CAT, APX, GPX, SOD, fruit size and yield. | [156] |
| Zn/Fe/Cu NPs NS | Open field, soil conditions | Mix with soil + Foliar 5 mg plant−1 + 100 mg L−1 | Length of plant, number of leaves, Chlorophyll, Vit A, number and weight of fruits, yield. | [161] |
| ZnO-chitosan 50 nm | Greenhouse, soil conditions | Foliar 400, 800, and 1200 mg L−1 | Number of leaves, number of fruits, chlorophylls, N, Mg, Mn. | [162] |
| Origin of Compost | Experimental Conditions | Forms and Levels of Application | Variables That Increase | Reference |
|---|---|---|---|---|
| Agricultural waste | Greenhouse Soil conditions | Mix with soil 50% soil–50% compost | Plant dry weight, chlorophyll, fruit weight, TSS, fructose, glucose, sucrose, malic acid, citric acid, yield. | [168] |
| Chicken manure | High tunnel Soil conditions | Mix with soil 66 g plant−1 | Plant dry matter, fruit firmness, TSS. | [169] |
| Vermicompost Chicken manure Cattle manure | Open field Soil conditions | Mix with soil 250 kg ha−1 | Fruit weight, firmness, yield, TSS, total sugars, Vit. C, N, P, K, Ca, Fe, Zn, Mn, Cu. | [170] |
| Poultry manure | Greenhouse, pots with soil | 0.10 g kg−1 soil | Indole Acetic Acid (IAA), Isopentenyl adenosine (iPA). | [66] |
| Ruminant manure | Open field Soil conditions | 150 kg ha−1 | Fruit yield. | [171] |
| Cattle manure (compost tea) | Open field Soil conditions | Foliar 8:1 compost:water 1.3 L m−2 | Fruit yield, resistance to Botrytis cinerea. | [172] |
| Vermicompost | Greenhouse Pots with soil | Mix with soil 200 g kg−1 soil | Leaf fresh weight, leaf area, root length. | [173] |
| Farmyard manure | Open field Soil conditions | Mix with soil 12.5 kg m−2 | Fruit dry weight, firmness, and yield. | [174] |
| Chicken manure | Greenhouse Soil conditions | Mix with soil 6 and 12 ton ha−1 | Plant height, stem thick, fruit yield. | [175] |
| Mixture of rose oil processing wastes, separated dairy manure, poultry manure, and wheat straws | Greenhouse Pots with substrate | Mix with substrate 12.5, 25, and 50% of total substrate | Number of leaves, number of roots, root length, stem thickness, K, Zn. | [176] |
| Compost NS | Greenhouse Pots with soil | 50% soil and 50% compost 100% compost | Vit. C, GSH, phenolics, anthocyanins. | [168] |
| Wastes of taif rose petals and red tea leaves | Greenhouse, pots with soil | Mix with soil 1.5 g kg−1 soil | Root fresh and dry weight, leaf area. | [177] |
| Vermicompost from food and paper wastes | High tunnel, soil conditions | Mix with soil 5 and 10 ton ha−1 | Number of runners and flowers, fruit yield. | [178] |
| Vermicompost | Greenhouse, pots with soil | 50% soil and 50% vermicompost | Plant height, leaf area, number of leaves, plant biomass, fruit weight and yield. | [179] |
| Vermicompost from cow dung and vegetable waste | Open field, soil conditions | Foliar 2 mL L−1 | Leaf area, plant biomass, fruit weight, firmness, TSS, yield. | [180] |
| Vermicompost Mushroom compost Farmyard manure | Open field, soil conditions | 170 kg ha−1 | Number of flowers, yield. | [181] |
| Farmyard manure Vermicompost | Open field, soil conditions | 30 and 80 ton ha−1 | Plant height, number of leaves, leaf area, number of runners, number, size, and yield of fruits, TSS, Vit. C, phenolics. | [182] |
| Product | Experimental Conditions | Forms and Levels of Application | Variables That Increase | Reference |
|---|---|---|---|---|
| H2O2 | ||||
| H2O2 | Greenhouse Hydroponic system (NFT) | Root dipping 1 M | Plant height, root length, leaf number, leaf area, number of adventious roots, plant biomass. | [196] |
| NO | ||||
| Sodium nitroprusside (SNP) as NO source | Greenhouse, pots with substrate | Foliar 50 and 75 μM | Phenolics, SOD, CAT, APX, POD. | [197] |
| Sodium nitroprusside (SNP) as NO source | Greenhouse, pots with substrate | Foliar 50 and 75 μM | Plant biomass, N, P, K, Ca, Mg, Fe, Zn, Mn, Cu. | [198] |
| Sodium nitroprusside (SNP) as NO source | Greenhouse, pots with substrate | Foliar 0.1 mM | Shoot biomass, chlorophyll, Fe, CAT, POD. | [199] |
| Sodium nitroprusside (SNP) as NO source | Greenhouse, pots with substrate | Foliar 75 μM | Vit. C, anthocyanins, phenolics. | [200] |
| Sodium nitroprusside (SNP) as NO source | Greenhouse, pots with substrate | Foliar 50 and 100 μM | SOD, CAT, APX, GPX, Vit. C, GSH. | [201] |
| Sodium nitroprusside (SNP) as NO source | Greenhouse, pots with substrate | Foliar 50 and 75 μM | Shoot and root dry weight, leaf area, chlorophyll, number of flowers, fruit size and weight, Vit. C, anthocyanins, phenolics. | [202] |
| H2S | ||||
| NaHS as H2S source | Greenhouse, pots with substrate | Foliar 0.2 mM | Plant biomass, chlorophyll, SOD, CAT, POD, Zn, Ca, Mg. | [203] |
| NaHS as H2S source | Greenhouse, pots with substrate | Root dipping 100 μΜ | Vit. C, GSH, DHA, heat shock proteins and overexpression of aquaporin-related genes. | [204] |
| NaHS as H2S source | Greenhouse, pots with substrate | Root dipping 0.125, 0.250, 1.250, 2.500, 12.500, 25.000, and 37.500 mM | Length and dry weight of roots, soluble sugars, SOD. | [205] |
| NaHS as H2S source | Greenhouse, pots with substrate | 0.2 and 0.5 mM | SPAD, chlorophyll fluorescence, fruit yield, SOD, APX, GR. | [206] |
| NaHS as H2S source | Greenhouse, pots with substrate | Root dipping 100 μΜ | Overexpression of genes such as cAPX, CAT, MnSOD, or GR, related with ascorbate-glutathione biosynthesis, transcription factor, and salt overly sensitive pathways. | [207] |
| PGPR Species | Experimental Conditions | Forms and Levels of Application | Variables That Increase | Reference |
|---|---|---|---|---|
| Plant Growth-Promoting Rhizobacteria (PGPR) | ||||
| Alcaligenes faecalis, Staphylococcus arlettae, S. simulans, Agrobacterium rubi, Pantoea agglomerans | Greenhouse, soil conditions | Root dipping 108 CFU mL−1 | Leaf area, number and weight of fruits, total yield. | [213] |
| Bacillus cereus | Growth chamber, pots with substrate | Mix with substrate 106 CFU g−1 substrate | Leaf area, number, weight and yield of fruits, sucrose concentration. | [214] |
| Pseudomonas florescence, Bacillus subtilis, Azotobacter chroococcum | Open field, soil conditions | Root dipping 109 CFU mL−1 | Plant height, number of leaves, leaf area, number of runners, chlorophylls, root fresh weight, fruit number, size and yield. | [215] |
| Bacillus licheniformis, B. subtilis, B. sp. RG1, B. sp. S1, B. sp. S2 | Open field, soil conditions | Root dipping + foliar 109 CFU ml−1 | Plant height, leaf area, number of runners, number of fruits, yield, chlorophyll, photosynthetic rate. | [216] |
| Bacillus subtilis, B. atrophaeus, B. spharicus, Staphylococcus kloosii Kocuria erythromyxa | Open field, soil conditions | Root dipping 108 CFU mL−1 | Shoot and root dry weight, chlorophyll, relative water content, yield, N, P, K, Ca, Mg, Fe, Mn, Zn, Cu. | [217] |
| Pseudomonas BA-8, Bacillus OSU-142, Bacillus M-3 | Open field, soil conditions | Root dipping + foliar 109 CFU ml−1 | Fruit yield, total sugars. | [218] |
| Bacillus megaterium, Bacillus spp., Paenibacillus polymyxa, Bacillus simplex | Open field, soil conditions | Root dipping 109 CFU mL−1 | Number and weight of fruits, TSS, Vit. C, yield. | [219] |
| Pseudomonas sp. | Greenhouse, soil conditions | NS | Plant height, fresh-dry weight, number of runners, number of fruits, yield. | [220] |
| Azotobacter chroococcum, A. vinelandi, Derxia sp., Bacillus megatherium, B. lichenformis, B. subtilis | Open field, soil conditions | Drench 20–40 × 106 CFU mL−1 | TSS, total sugars, TA, yield. | [221] |
| Kocuria E43, Alcaligenes 637Ca Pseudomonas 53/6 | Greenhouse, pots with soil | Root dipping 109 CFU mL−1 | Fruit number, weight, and yield, SPAD, stomatal conductance, CAT, SOD, APX. | [222] |
| Azospirillum brasilense | Open field, soil conditions | Root dipping 109 CFU mL−1 | SPAD, photosynthesis, yield, amino acids and organic acids. | [223] |
| Pseudomonas BA-8, Bacillus OSU-142, Bacillus M-3 | Open field, soil conditions | Root dipping 109 CFU mL−1 | Fruit yield, P, Fe, Zn. | [224] |
| B. methylotrophicus | In vitro | Growing medium 104 CFU | Shoot and root fresh weight, petiole length. | [225] |
| Commercial formulation of several PGPR | Open field, soil conditions | Root dipping 109 CFU mL−1 | CAT, POD, SOD, fruit yield. | [226] |
| Azotobacter chroococcum, Pseudomonas fluorescens | Open field, soil conditions | Root dipping 3 × 107 CFU mL−1 | Plant height, number of leaves, leaf area, number of runners, number, size, and yield of fruits, TSS, Vit. C, phenolics | [182] |
| Endophytic bacteria | ||||
| B. velezensis | Greenhouse, pots with substrate | Drench 5 × 105 spores plant−1 | Shoot and root fresh weight, fruit yield. | [227] |
| Arthrobacter agilis, B. methylotrophicus | In vitro | Growing medium 100 μL of bacterial suspension | % Seed germination, shoot fresh weight. | [228] |
| Greenhouse | Root dipping 100 μL of bacterial suspension | Fruit yield. | ||
| Azospirillum brasilense, Burkholderia cepacian, Enterobacter cloacae | Greenhouse, pots with soil | Root dipping 109 CFU mL−1 | Root length and dry weight, aerial dry weight. | [229] |
| Azospirillum brasilense | Growth chamber, pots with substrate | Root dipping 106 CFU ml−1 | Root length and dry weight, shoot dry weight, total sugars of root exudates. | [230] |
| B. amyloliquefaciens, Paraburkholderia fungorum | Open field, soil conditions | Root dipping 109 CFU mL−1 | Root length, fresh and dry weight, shoot dry weight, fruit weight, anthocyanins, carotenoids, flavonoids, phenolics, antioxidant capacity. | [98] |
| Fungi Species | Experimental Conditions | Forms and Levels of Application | Variables That Increase | Reference |
|---|---|---|---|---|
| Arbuscular Mycorrhizal Fungi (AMF) | ||||
| R. intraradices | Greenhouse, pots with substrate | 0.5 g plant−1 | CO2 assimilation, stomatal conductance, relative water content. | [235] |
| G. mosseae, G. aggregatum | Greenhouse, pots with substrate | NS | P concentration, free amino acids concentration. | [236] |
| G. mosseae | Greenhouse, pots with soil | 1 g plant−1 | Dry weight of shoots, phenolics, antioxidant activity, SOD. | [237] |
| F. mosseae, F. geosporus, C. claroideum, G. microagregatum, R. irregularis | Greenhouse, pots with substrate | 20 g plant−1 | Fruit yield, root length. | [238] |
| G. intraradices | Greenhouse, pots with substrate | 2 mL plant−1 from solution of 50 g L−1 | K, Cu, phenolics, anthocyanins, flavonoids. | [239] |
| G. intraradices | Open field, soil conditions | 1 g plant−1 | Root biomass, daughter plants per mother plant. | [240] |
| R. clarus | Greenhouse, pots with substrate | 60 g plant−1 | Shoot and root biomass, relative water content, net photosynthesis. | [126] |
| F. mosseae, F. geosporus | Greenhouse, pots with substrate | 1:10 inoculated substrate: growing substrate mix | Shoot and root length and fresh weight, SPAD, fruit weight. | [241] |
| Mix of various Glomus species | Greenhouse, pots with substrate | 100 mL of mycorrhizal preparation plant−1 | Anthocyanins concentration. | [242] |
| G. fasciculatum, G. etunicatum | Greenhouse, pots with substrate | 2.5 g plant−1 | Shoot dry weight, P and K concentration. | [243] |
| G. irregularis | Greenhouse, pots with substrate | 80–100 spores plant−1 | Length, volume, and dry weight of roots. | [244] |
| Cetraspora pellucida, Claroideoglomus etunicatum and mycorrhizal community | Greenhouse, pots with substrate | 10 g plant−1 | Aerial biomass, root length and biomass, anthocyanins, flavonoids, phenolics. | [245] |
| Gigaspora margarita | Greenhouse, pots with soil | 30 spores plant−1 | Root biomass, Mg, Mn. | [246] |
| G. clarum | P, Mg, Ca, S, Fe, Cu, Zn. | |||
| Gigaspora rosea | N, P, Mg, Ca, S, Fe, Cu, Mn Zn. | |||
| G. mosseae, G. intraradices | Greenhouse, pots with substrate | 20 spores g−1 of substrate | SPAD, number of leaves and flowers, number of fruits. | [247] |
| G. mosseae | NS | 10% of inoculated substrate | Plant height, leaf area, fresh and dry weight of shoot and roots, chlorophyll. | [248] |
| AMF NS | Open field, soil conditions | 20 g plant−1 | Plant height, biomass, fruit size, yield. | [180] |
| Trichoderma | ||||
| T. harzianum T. virens | Greenhouse, pots with soil | 25 mL plant−1 (107 spores mL−1) | Root length and dry weight, number of fruits, yield, Vit. C, anthocyanins. | [249] |
| T. harzianum T. viride | Open field, soil conditions | Root dipping in fungi preparation (106 spores mL−1) | Root biomass, fruit yield. | [250] |
| T. citrinoviride | Greenhouse, pots with substrate | Root dipping in fungi preparation (2 × 106 CFU mL−1) | Plant dry weight, PSII efficiency. | [251] |
| T. harzianum | Greenhouse, pots with soil | 50 mL plant−1 (9.90 × 106 CFU 100 mL−1) | Vegetative growth, number of flowers, number, weight, and yield of fruits, TSS, TA, Vit. C. | [252] |
| T. viride | NS | 10% of inoculated substrate | Plant height, leaf area, fresh and dry weight of shoot and roots, chlorophyll. | [248] |
| Light Source | Experimental Conditions | Wavelength (nm)/Photosynthetic Photon Flux Density (PPFD) (µmol m−2 s−1) | Variables That Increase | Reference |
|---|---|---|---|---|
| LED | Greenhouse, pots with substrate | 450–550/400 | Photosynthetic rate, leaf area, leaf dry weight, fruit number, weight, yield, TSS and firmness. | [264] |
| Fluorescent lamp (FL) | 405–610/NS | Photosynthetic rate, leaf area, leaf dry weight. | ||
| Blue LED | Greenhouse, pots with substrate | 447/335 | Leaf area, number of leaves, number of flowers, N, K, Ca, Fe, Mn, and Zn concentration. | [257] |
| Red LED | 666/375 | |||
| White LED | 494/330 | |||
| FL | 479/275 | |||
| FL+UV | 480/314 | |||
| Red:Blue LED (8:2) | Greenhouse, pots with substrate | 445–659/106–117 | Number of leaves, crown diameter, plant dry weight, number of flowers, number, and weight of fruits, TSS, Vit. C. | [65] |
| Red:Blue LED (5:5) | 445–659/107–125 | Crown diameter, plant dry weight, TSS of fruits. | ||
| Red:Blue LED (2:8) | 445–659/105–121 | Crown diameter, plant dry weight, K concentration. | ||
| Blue LED | Greenhouse, pots with substrate | 448/75 | Fruit yield, glucose concentration. | [265] |
| Red LED | 661/75 | Sucrose, citric acid, malic acid concentration. | ||
| Blue + Red LED | 634/75 | Fruit yield, fructose, glucose | ||
| Red LED | In vitro | 660/45 | Plant height, number of leaves, root length. | [266] |
| Blue LED | Greenhouse, pots with substrate | 470/190 | Days to anthesis, fruit yield. | [267] |
| Light with various color temperatures (3000, 4000, 5000, and 6500 K) | Growth chamber, pots with substrate | NS | Leaf number and size, crown diameter, dry weight of plant, SPAD. | [268] |
| Red, Blue and Red:Blue LED | Greenhouse, pots with substrate | 450–730/190 | Fruit anthocyanins and proanthocyanins. | [269] |
| LED NS | Greenhouse, pots with substrate | 450–550/400 | Less days to flowering, number of flowers, dry biomass of plant, number, weight, and yield of fruits, TSS, firmness. | [270] |
| Red LED | Greenhouse, pots with substrate | 660/200 | Leaf fresh weight, fruit number and size. | [271] |
| Blue/Red | 460–660/200 | Leaf fresh weight, leaf area, SPAD, fruit number and size, TSS. | ||
| White–Yellow | 400–700/200 | Leaf fresh weight, crown fresh weight, SPAD, fruit number and size. | ||
| Red LED | Greenhouse, pots with substrate | 660/200 | CO2 assimilation rate, water use efficiency, stomatal conductance, transpiration. | [272] |
| Blue/Red | 460–660/200 | |||
| White–Yellow | 400–700/200 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garza-Alonso, C.A.; Olivares-Sáenz, E.; González-Morales, S.; Cabrera-De la Fuente, M.; Juárez-Maldonado, A.; González-Fuentes, J.A.; Tortella, G.; Valdés-Caballero, M.V.; Benavides-Mendoza, A. Strawberry Biostimulation: From Mechanisms of Action to Plant Growth and Fruit Quality. Plants 2022, 11, 3463. https://doi.org/10.3390/plants11243463
Garza-Alonso CA, Olivares-Sáenz E, González-Morales S, Cabrera-De la Fuente M, Juárez-Maldonado A, González-Fuentes JA, Tortella G, Valdés-Caballero MV, Benavides-Mendoza A. Strawberry Biostimulation: From Mechanisms of Action to Plant Growth and Fruit Quality. Plants. 2022; 11(24):3463. https://doi.org/10.3390/plants11243463
Chicago/Turabian StyleGarza-Alonso, Carlos Alberto, Emilio Olivares-Sáenz, Susana González-Morales, Marcelino Cabrera-De la Fuente, Antonio Juárez-Maldonado, José Antonio González-Fuentes, Gonzalo Tortella, Marin Virgilio Valdés-Caballero, and Adalberto Benavides-Mendoza. 2022. "Strawberry Biostimulation: From Mechanisms of Action to Plant Growth and Fruit Quality" Plants 11, no. 24: 3463. https://doi.org/10.3390/plants11243463
APA StyleGarza-Alonso, C. A., Olivares-Sáenz, E., González-Morales, S., Cabrera-De la Fuente, M., Juárez-Maldonado, A., González-Fuentes, J. A., Tortella, G., Valdés-Caballero, M. V., & Benavides-Mendoza, A. (2022). Strawberry Biostimulation: From Mechanisms of Action to Plant Growth and Fruit Quality. Plants, 11(24), 3463. https://doi.org/10.3390/plants11243463