

Guava (Psidium guajava L.) Leaf Extract as Bioactive Substances for Anti-Androgen and Antioxidant Activities

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Extraction Yield and Bioactive Compound Estimation
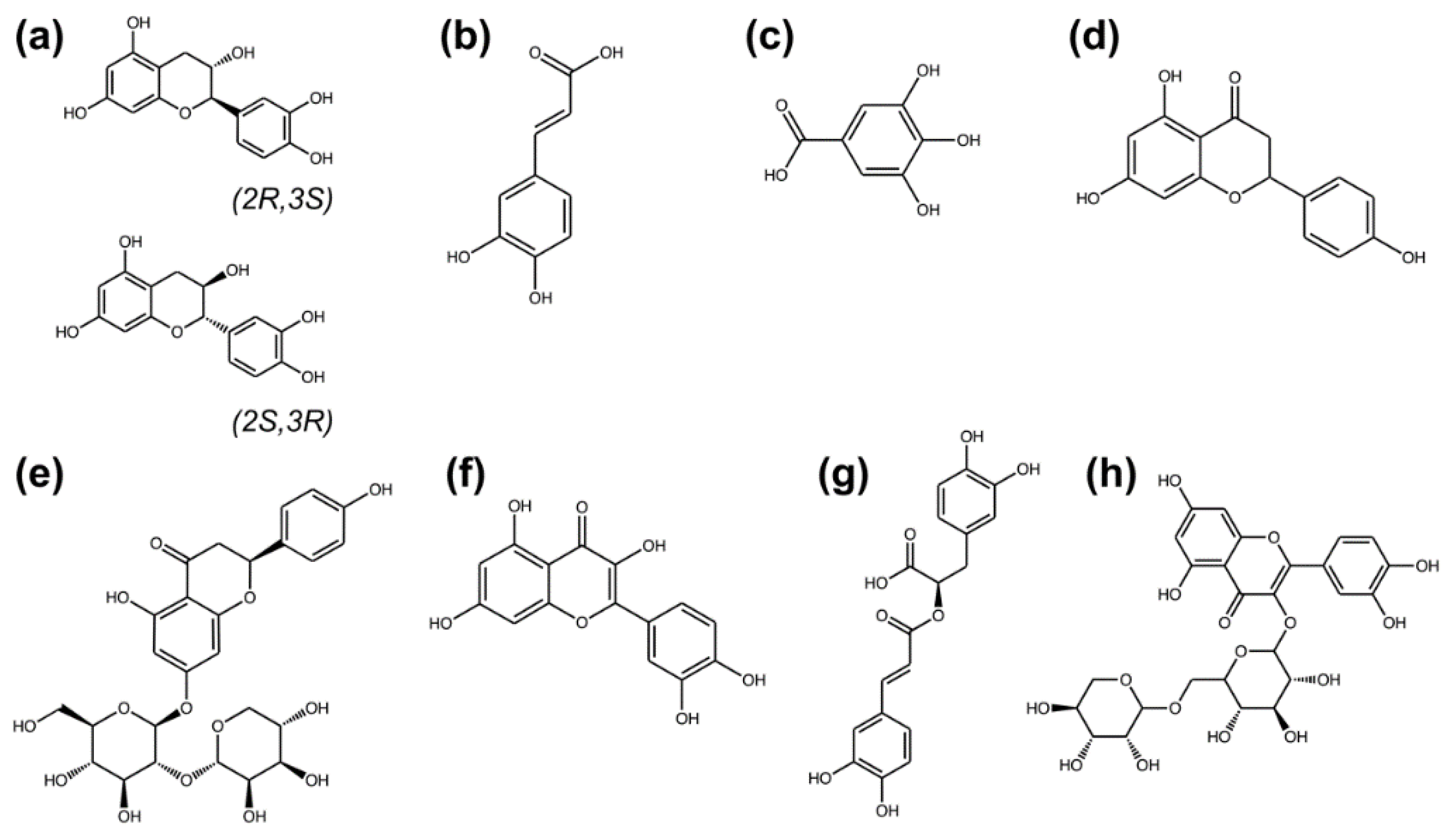
2.2. Characterization of the Phytochemical Profile by Liquid Chromatography–Mass Spectrometry (LC–MS) Analysis
2.3. Antioxidant Activities of Guava Leaf Extract
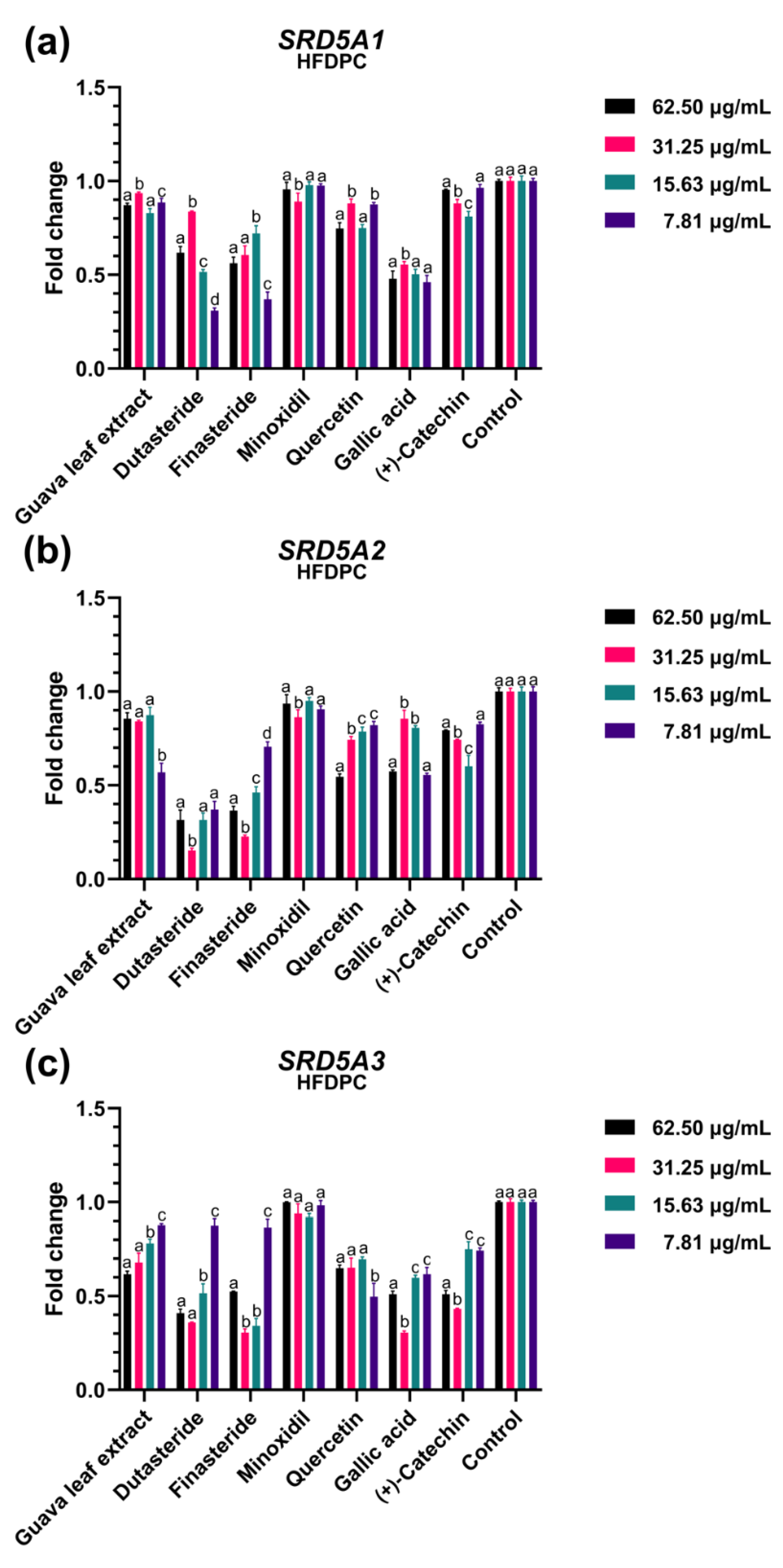
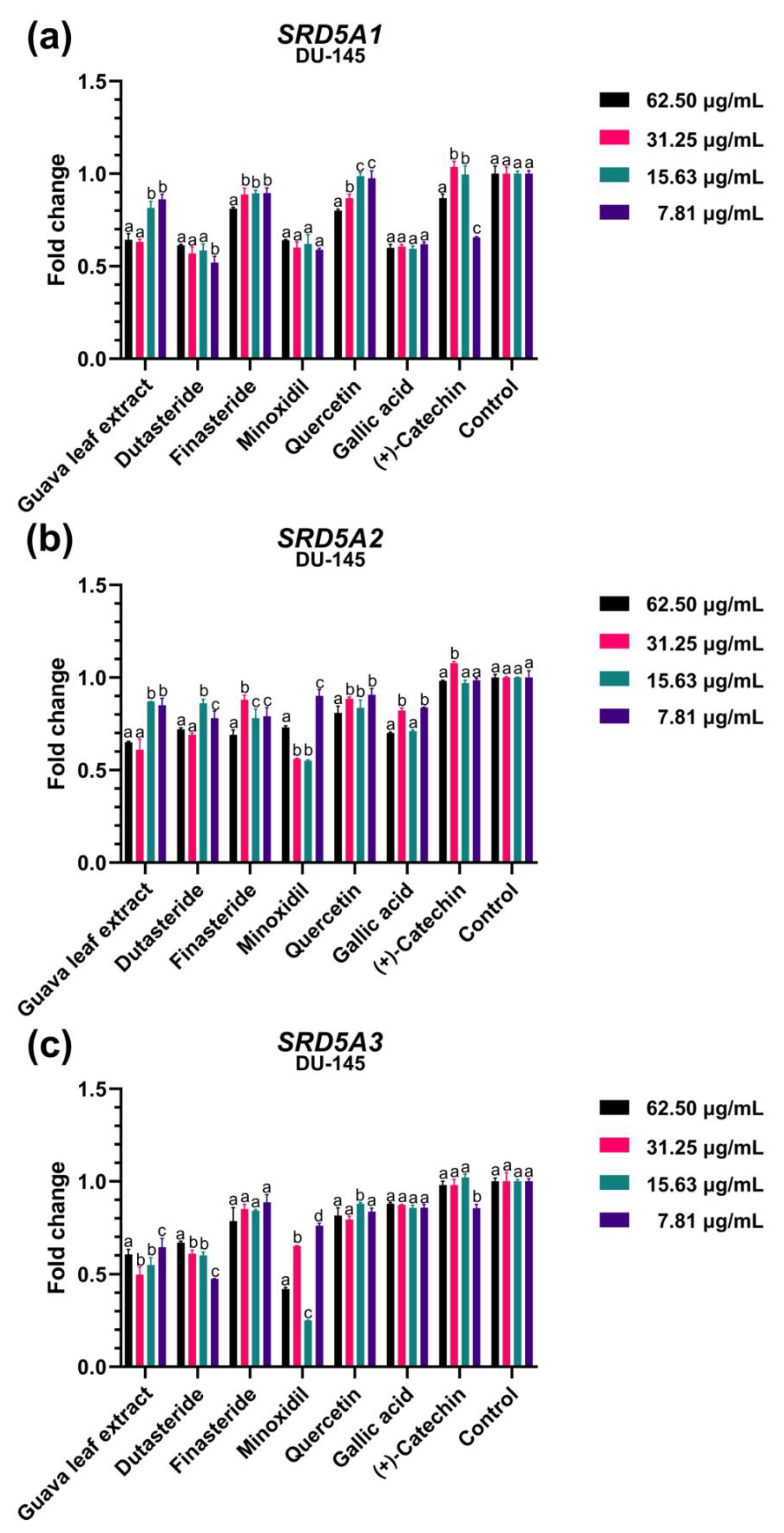
2.4. Effect of Guava Leaf Extract on the Expression of Gene Asscociated with Androgen-Dependent Pathway
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plant Material and Crude Extracts
4.3. Bioactive Compound Estimation
4.3.1. Total Phenolic Content (TPC) Assay
4.3.2. Total Flavonoid Content (TFC) Assay
4.3.3. Total Polysaccharide Content
4.3.4. Total Protein Content
4.4. Characterization of Psidium guajava Leaf Extract by Liquid Chromatography–Mass Spectrometry (LC–MS)
4.5. Antioxidant Activities of Psidium guajava Leaf Extract
4.5.1. DPPH Radical Scavenging Assay
4.5.2. ABTS•+ Radical Scavenging Assay
4.5.3. Metal Chelating Assay
4.6. Cell Lines and Culture
4.7. Cell Viability Assay
4.8. RT-PCR Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flores, G.; Wu, S.B.; Negrin, A.; Kennelly, E.J. Chemical composition and antioxidant activity of seven cultivars of guava (Psidium guajava) fruits. Food Chem. 2015, 170, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Naseer, S.; Hussain, S.; Naeem, N.; Pervaiz, M.; Rahman, M. The phytochemistry and medicinal value of Psidium guajava (guava). Clin. Phytosci. 2018, 4, 32. [Google Scholar] [CrossRef] [Green Version]
- Nantitanon, W.; Yotsawimonwat, S.; Okonogi, S. Factors influencing antioxidant activities and total phenolic content of guava leaf extract. LWT -Food Sci. Technol. 2010, 43, 1095–1103. [Google Scholar] [CrossRef]
- Gutiérrez, R.M.P.; Mitchell, S.; Solis, R.V. Psidium guajava: A review of its traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2008, 117, 1–27. [Google Scholar] [CrossRef]
- Fu, D.; Huang, J.; Li, K.; Chen, Y.; He, Y.; Sun, Y.; Guo, Y.; Du, L.; Qu, Q.; Miao, Y.; et al. Dihydrotestosterone-induced hair regrowth inhibition by activating androgen receptor in C57BL6 mice simulates androgenetic alopecia. Biomed. Pharmacother. 2021, 137, 111247. [Google Scholar] [CrossRef]
- Paik, J.; Yoon, J.; Sim, W.; Kim, B.; Kim, N. The prevalence and types of androgenetic alopecia in Korean men and women. Br. J. Dermatol. 2001, 145, 95–99. [Google Scholar] [CrossRef]
- Ceruti, J.M.; Leirós, G.J.; Balañá, M.E. Androgens and androgen receptor action in skin and hair follicles. Mol. Cell Endocrinol. 2017, 465, 122–133. [Google Scholar] [CrossRef] [Green Version]
- Pathomvanich, D.; Pongratananukul, S.; Thienthaworn, P.; Manoshai, S. A random study of Asian male androgenetic alopecia in Bangkok, Thailand. Dermatol. Surg. 2002, 28, 804–807. [Google Scholar]
- Gupta, M.; Mysore, V. Classifications of patterned hair loss: A review. J. Cutan. Aesthet. Surg. 2016, 9, 3–12. [Google Scholar] [CrossRef]
- Dinh, Q.Q.; Sinclair, R. Female pattern hair loss: Current treatment concepts. Clin. Interv. Aging 2007, 2, 189–199. [Google Scholar]
- English, R.S. A hypothetical pathogenesis model for androgenic alopecia: Clarifying the dihydrotestosterone paradox and rate-limiting recovery factors. Med. Hypotheses 2018, 111, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Manosroi, A.; Ruksiriwanich, W.; Manosroi, W.; Abe, M.; Manosroi, J. In vivo hair growth promotion activity of gel containing niosomes loaded with the Oryza sativa bran fraction (OSF3). Adv. Sci. Lett. 2012, 16, 222–228. [Google Scholar] [CrossRef]
- Lizeth, M.J.; César, D.V.V.; Rocío, O.L.; Jorge, O.C.; Augusto, R.M. Genetic and molecular aspects of androgenetic alopecia. Indian J. Dermatol. Venereol. Leprol. 2018, 84, 263. [Google Scholar]
- Manosroi, A.; Ruksiriwanich, W.; Abe, M.; Sakai, H.; Aburai, K.; Manosroi, W.; Manosroi, J. Physico-chemical properties of cationic niosomes loaded with fraction of rice (Oryza sativa) bran extract. J. Nanosci. Nanotechnol. 2012, 12, 7339–7345. [Google Scholar] [CrossRef] [PubMed]
- Anastassakis, K. The role of the sebaceous gland in AGA/FPHL. In Androgenetic Alopecia from A to Z; Springer: Berlin/Heidelberg, Germany, 2022; pp. 233–240. [Google Scholar]
- Premanand, A.; Reena Rajkumari, B. Androgen modulation of Wnt/β-catenin signaling in androgenetic alopecia. Arch. Dermatol. Res. 2018, 310, 391–399. [Google Scholar] [CrossRef]
- Kwack, M.H.; Sung, Y.K.; Chung, E.J.; Im, S.U.; Ahn, J.S.; Kim, M.K.; Kim, J.C. Dihydrotestosterone-inducible dickkopf 1 from balding dermal papilla cells causes apoptosis in follicular keratinocytes. J. Investig. Dermatol. 2008, 128, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Yamana, K.; Labrie, F.; The, V.L. Human type 3 5α-reductase is expressed in peripheral tissues at higher levels than types 1 and 2 and its activity is potently inhibited by finasteride and dutasteride. Horm. Mol. Biol. Clin. Investig. 2010, 2, 293–299. [Google Scholar] [CrossRef]
- Robitaille, J.; Langlois, V.S. Consequences of steroid-5α-reductase deficiency and inhibition in vertebrates. Gen. Comp. Endocrinol. 2020, 290, 113400. [Google Scholar] [CrossRef]
- Lourith, N.; Kanlayavattanakul, M.; Chaikul, P. Para rubber seed oil: The safe and efficient bio-material for hair loss treatment. J. Cosmet. Dermatol. 2021, 20, 2160–2167. [Google Scholar] [CrossRef]
- Teeranachaideekul, V.; Parichatikanond, W.; Junyaprasert, V.B.; Morakul, B. Pumpkin seed oil-loaded niosomes for topical application: 5α-reductase inhibitory, anti-inflammatory, and in vivo nti-hair loss effects. Pharmaceuticals 2022, 15, 930. [Google Scholar] [CrossRef]
- Madaan, A.; Verma, R.; Singh, A.T.; Jaggi, M. Review of hair follicle dermal papilla cells as in vitro screening model for hair growth. Int. J. Cosmet. Sci. 2018, 40, 429–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semalty, M.; Semalty, A.; Joshi, G.P.; Rawat, M.S.M. Hair growth and rejuvenation: An overview. J. Cosmet. Dermatol. 2011, 22, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Katzer, T.; Leite Junior, A.; Beck, R.; da Silva, C. Physiopathology and current treatments of androgenetic alopecia: Going beyond androgens and anti-androgens. Dermatol.Ther. 2019, 32, e13059. [Google Scholar] [CrossRef] [PubMed]
- Nestor, M.S.; Ablon, G.; Gade, A.; Han, H.; Fischer, D.L. Treatment options for androgenetic alopecia: Efficacy, side effects, compliance, financial considerations, and ethics. J. Cosmet. Dermatol. 2021, 20, 3759–3781. [Google Scholar] [CrossRef]
- Dhariwala, M.Y.; Ravikumar, P. An overview of herbal alternatives in androgenetic alopecia. J. Cosmet. Dermatol. 2019, 18, 966–975. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem in 2021: New data content and improved web interfaces. Nucleic Acids Res. 2021, 49, D1388–D1395. [Google Scholar] [CrossRef]
- Donovan, J.L.; Crespy, V.; Oliveira, M.; Cooper, K.A.; Gibson, B.B.; Williamson, G. (+)-Catechin is more bioavailable than (−)-catechin: Relevance to the bioavailability of catechin from cocoa. Free Radic. Res. 2006, 40, 1029–1034. [Google Scholar] [CrossRef]
- Kumar, M.; Tomar, M.; Amarowicz, R.; Saurabh, V.; Nair, M.S.; Maheshwari, C.; Sasi, M.; Prajapati, U.; Hasan, M.; Singh, S.; et al. Guava (Psidium guajava L.) leaves: Nutritional composition, phytochemical profile, and health-promoting bioactivities. Foods 2021, 10, 752. [Google Scholar] [CrossRef]
- Díaz-de-Cerio, E.; Gómez-Caravaca, A.M.; Verardo, V.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Determination of guava (Psidium guajava L.) leaf phenolic compounds using HPLC-DAD-QTOF-MS. J. Funct. Foods 2016, 22, 376–388. [Google Scholar] [CrossRef]
- Sampath Kumar, N.S.; Sarbon, N.M.; Rana, S.S.; Chintagunta, A.D.; Prathibha, S.; Ingilala, S.K.; Jeevan Kumar, S.P.; Sai Anvesh, B.; Dirisala, V.R. Extraction of bioactive compounds from Psidium guajava leaves and its utilization in preparation of jellies. AMB Express 2021, 11, 36. [Google Scholar] [CrossRef]
- Jung, Y.H.; Chae, C.W.; Choi, G.E.; Shin, H.C.; Lim, J.R.; Chang, H.S.; Park, J.; Cho, J.H.; Park, M.R.; Lee, H.J.; et al. Cyanidin 3-o-arabinoside suppresses DHT-induced dermal papilla cell senescence by modulating p38-dependent ER-mitochondria contacts. J. Biomed. Sci. 2022, 29, 17. [Google Scholar] [CrossRef] [PubMed]
- Iwegbue, C.M.A.; Emakunu, O.S.; Obi, G.; Nwajei, G.E.; Martincigh, B.S. Evaluation of human exposure to metals from some commonly used hair care products in Nigeria. Toxicol. Rep. 2016, 3, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Boonsong, P.; Laohakunjit, N.; Kerdchoechuen, O. Natural pigments from six species of Thai plants extracted by water for hair dyeing product application. J. Clean. Prod. 2012, 37, 93–106. [Google Scholar] [CrossRef]
- Gulcin, İ.; Alwasel, S.H. Metal ions, metal chelators and metal chelating assay as antioxidant method. Processes 2022, 10, 132. [Google Scholar] [CrossRef]
- Seo, J.; Lee, S.; Elam, M.L.; Johnson, S.A.; Kang, J.; Arjmandi, B.H. Study to find the best extraction solvent for use with guava leaves (Psidium guajava L.) for high antioxidant efficacy. Food Sci. Nutr. 2014, 2, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Yen, G.C. Antioxidant activity and free radical-scavenging capacity of extracts from guava (Psidium guajava L.) leaves. Food Chem. 2007, 101, 686–694. [Google Scholar] [CrossRef]
- Tachakittirungrod, S.; Okonogi, S.; Chowwanapoonpohn, S. Study on antioxidant activity of certain plants in Thailand: Mechanism of antioxidant action of guava leaf extract. Food Chem. 2007, 103, 381–388. [Google Scholar] [CrossRef]
- Khokhar, S.; Owusu Apenten, R.K. Iron binding characteristics of phenolic compounds: Some tentative structure–activity relations. Food Chem. 2003, 81, 133–140. [Google Scholar] [CrossRef]
- Inui, S.; Itami, S. Molecular basis of androgenetic alopecia: From androgen to paracrine mediators through dermal papilla. J. Dermatol. Sci. 2011, 61, 1–6. [Google Scholar] [CrossRef]
- Lai, J.J.; Chang, P.; Lai, K.P.; Chen, L.; Chang, C. The role of androgen and androgen receptor in skin-related disorders. Arch. Dermatol. Res. 2012, 304, 499–510. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.S.; Thomas, L.N.; Douglas, R.C.; Lazier, C.B.; Rittmaster, R.S. Relative potency of testosterone and dihydrotestosterone in preventing atrophy and apoptosis in the prostate of the castrated rat. J. Clin. Investig. 1996, 98, 2558–2563. [Google Scholar] [CrossRef] [PubMed]
- Anastassakis, K. Hormonal and genetic etiology of male androgenetic alopecia. In Androgenetic Alopecia from A to Z; Springer: Berlin/Heidelberg, Germany, 2022; pp. 135–180. [Google Scholar]
- Uemura, M.; Tamura, K.; Chung, S.; Honma, S.; Okuyama, A.; Nakamura, Y.; Nakagawa, H. Novel 5α-steroid reductase (SRD5A3, type-3) is overexpressed in hormone-refractory prostate cancer. Cancer Sci. 2008, 99, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, P.; Serrano-Falcón, C.; Torres, J.M.; Serrano, S.; Ortega, E. 5α-Reductase isozymes and aromatase mRNA levels in plucked hair from young women with female pattern hair loss. Arch. Dermatol. Res. 2018, 310, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Azzouni, F.; Zeitouni, N.; Mohler, J. Role of 5α-reductase inhibitors in androgen-stimulated skin disorders. J. Drugs Dermatol. 2013, 12, e30–e35. [Google Scholar] [PubMed]
- Ruksiriwanich, W.; Khantham, C.; Muangsanguan, A.; Chittasupho, C.; Rachtanapun, P.; Jantanasakulwong, K.; Phimolsiripol, Y.; Sommano, S.R.; Sringarm, K.; Ferrer, E. Phytochemical constitution, anti-inflammation, anti-androgen, and hair growth-promoting potential of shallot (Allium ascalonicum L.) extract. Plants 2022, 11, 1499. [Google Scholar] [CrossRef]
- Langlois, V.S.; Duarte-Guterman, P.; Ing, S.; Pauli, B.D.; Cooke, G.M.; Trudeau, V.L. Fadrozole and finasteride exposures modulate sex steroid- and thyroid hormone-related gene expression in Silurana (Xenopus) tropicalis early larval development. Gen. Comp. Endocrinol. 2010, 166, 417–427. [Google Scholar] [CrossRef]
- Khantham, C.; Yooin, W.; Sringarm, K.; Sommano, S.R.; Jiranusornkul, S.; Carmona, F.D.; Nimlamool, W.; Jantrawut, P.; Rachtanapun, P.; Ruksiriwanich, W. Effects on steroid 5-alpha reductase gene expression of Thai rice bran extracts and molecular dynamics study on SRD5A2. Biology 2021, 10, 319. [Google Scholar] [CrossRef]
- Pekmezci, E.; Türkoğlu, M. Minoxidil acts as an antiandrogen: A study of 5α-reductase type 2 gene expression in a human keratinocyte cell line. Acta Dermatovenerol. Croat. 2017, 25, 271–275. [Google Scholar]
- Mellin, T.N.; Busch, R.; Rasmusson, G.H. Azasteroids as inhibitors of testosterone 5α-reductase in mammalian skin. J. Steroid Biochem. Mol. Biol. 1993, 44, 121–131. [Google Scholar] [CrossRef]
- Tao, R.; Miao, L.; Yu, X.; Orgah, J.O.; Barnabas, O.; Chang, Y.; Liu, E.; Fan, G.; Gao, X. Cynomorium songaricum Rupr demonstrates phytoestrogenic or phytoandrogenic like activities that attenuates benign prostatic hyperplasia via regulating steroid 5-α-reductase. J. Ethnopharmacol. 2019, 235, 65–74. [Google Scholar] [CrossRef]
- Hu, X.; Li, X.; Deng, P.; Zhang, Y.; Liu, R.; Cai, D.; Xu, Q.; Jiang, X.; Sun, J.; Bai, W. The consequence and mechanism of dietary flavonoids on androgen profiles and disorders amelioration. Crit. Rev. Food Sci. Nutr. 2022, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Song, L.; Wang, H.; Wang, J.; Xu, Z.; Xing, N. Quercetin in prostate cancer: Chemotherapeutic and chemopreventive effects, mechanisms and clinical application potential. Oncol. Rep. 2015, 33, 2659–2668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiipakka, R.A.; Zhang, H.Z.; Dai, W.; Dai, Q.; Liao, S. Structure–activity relationships for inhibition of human 5α-reductases by polyphenols. Biochem. Pharmacol. 2002, 63, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ding, Z.; Wang, Z.; Lu, J.-F.; Maity, S.N.; Navone, N.M.; Logothetis, C.J.; Mills, G.B.; Kim, J. Androgen regulation of 5α-reductase isoenzymes in prostate cancer: Implications for prostate cancer prevention. PLoS ONE 2011, 6, e28840. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Speiser, P.W. Regulation of HSD17B1 and SRD5A1 in lymphocytes. Mol. Genet. Metab. 1999, 68, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Pihlajamaa, P.; Sahu, B.; Jänne, O.A. Determinants of receptor- and tissue-specific actions in androgen signaling. Endocr. Rev. 2015, 36, 357–384. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Wang, L.; Wang, R.; Luo, X.; Li, Y.; Li, J.; Li, Y.; Chen, Z. Phenolic contents, cellular antioxidant activity and antiproliferative capacity of different varieties of oats. Food Chem. 2018, 239, 260–267. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Luo, D.; Qu, C.; Lin, G.; Zhang, Z.; Xie, J.; Chen, H.; Liang, J.; Li, C.; Wang, H.; Su, Z. Character and laxative activity of polysaccharides isolated from Dendrobium officinale. J. Funct. Foods 2017, 34, 106–117. [Google Scholar] [CrossRef]
- Waterborg, J.H. The Lowry method for protein quantitation. In The Protein Protocols Handbook; Springer: Berlin/Heidelberg, Germany, 2009; pp. 7–10. [Google Scholar]
- Starace, M.; Orlando, G.; Alessandrini, A.; Piraccini, B.M. Female androgenetic alopecia: An update on diagnosis and management. Am. J. Clin. Dermatol. 2020, 21, 69–84. [Google Scholar] [CrossRef]
- Zhou, S.D.; Xu, X.; Lin, Y.F.; Xia, H.Y.; Huang, L.; Dong, M.S. On-line screening and identification of free radical scavenging compounds in Angelica dahurica fermented with Eurotium cristatum using an HPLC-PDA-Triple-TOF-MS/MS-ABTS system. Food Chem. 2019, 272, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, K.; Charles, A.L. In vitro antioxidant activity of Kyoho grape extracts in DPPH and ABTS assays: Estimation methods for EC(50) using advanced statistical programs. Food Chem. 2019, 275, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Grochowski, D.M.; Uysal, S.; Aktumsek, A.; Granica, S.; Zengin, G.; Ceylan, R.; Locatelli, M.; Tomczyk, M. In vitro enzyme inhibitory properties, antioxidant activities, and phytochemical profile of Potentilla thuringiaca. Phytochem. Lett. 2017, 20, 365–372. [Google Scholar] [CrossRef]
- Manosroi, A.; Chankhampan, C.; Kietthanakorn, B.O.; Ruksiriwanich, W.; Chaikul, P.; Boonpisuttinant, K.; Sainakham, M.; Manosroi, W.; Tangjai, T.; Manosroi, J. Pharmaceutical and cosmeceutical biological activities of hemp (Cannabis sativa L. var. sativa) leaf and seed extracts. Chiang Mai J. Sci. 2019, 46, 180–195. [Google Scholar]
- Orellana, E.A.; Kasinski, A.L. Sulforhodamine B (SRB) assay in cell culture to investigate cell proliferation. Bio Protoc. 2016, 6, e1984. [Google Scholar] [CrossRef] [PubMed]
- Wang, F. Semi-quantitative RT-PCR: An effective method to explore the regulation of gene transcription level affected by environmental pollutants. In Environmental Toxicology and Toxicogenomics: Principles, Methods, and Applications; Pan, X., Zhang, B., Eds.; Springer: New York, NY, USA, 2021; pp. 95–103. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Compound Name | Molecular Formula | m/z | Content (mg/g Extract) |
|---|---|---|---|
| Catechin * | C15H14O6 | 290.27 | 2.215 ± 0.031 |
| Caffeic acid | C9H8O4 | 180.16 | 0.074 ± 0.001 |
| Epicatechin ** | C15H14O6 | 290.26 | Nd. |
| Epigallocatechin gallate | C22H18O11 | 458.37 | Nd. |
| Gallic acid | C7H6O5 | 170.20 | 0.751 ± 0.008 |
| Naringenin | C15H12O5 | 272.25 | 0.066 ± 0.001 |
| Naringin | C27H32O14 | 580.50 | 0.045 ± 0.001 |
| p-Coumaric acid | C9H8O3 | 164.15 | Nd. |
| Quercetin | C15H10O7 | 302.23 | 0.520 ± 0.022 |
| Rosmarinic acid | C18H16O8 | 360.30 | 0.039 ± 0.018 |
| Rutin | C27H30O16 | 610.50 | 0.109 ± 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruksiriwanich, W.; Khantham, C.; Muangsanguan, A.; Phimolsiripol, Y.; Barba, F.J.; Sringarm, K.; Rachtanapun, P.; Jantanasakulwong, K.; Jantrawut, P.; Chittasupho, C.; et al. Guava (Psidium guajava L.) Leaf Extract as Bioactive Substances for Anti-Androgen and Antioxidant Activities. Plants 2022, 11, 3514. https://doi.org/10.3390/plants11243514
Ruksiriwanich W, Khantham C, Muangsanguan A, Phimolsiripol Y, Barba FJ, Sringarm K, Rachtanapun P, Jantanasakulwong K, Jantrawut P, Chittasupho C, et al. Guava (Psidium guajava L.) Leaf Extract as Bioactive Substances for Anti-Androgen and Antioxidant Activities. Plants. 2022; 11(24):3514. https://doi.org/10.3390/plants11243514
Chicago/Turabian StyleRuksiriwanich, Warintorn, Chiranan Khantham, Anurak Muangsanguan, Yuthana Phimolsiripol, Francisco J. Barba, Korawan Sringarm, Pornchai Rachtanapun, Kittisak Jantanasakulwong, Pensak Jantrawut, Chuda Chittasupho, and et al. 2022. "Guava (Psidium guajava L.) Leaf Extract as Bioactive Substances for Anti-Androgen and Antioxidant Activities" Plants 11, no. 24: 3514. https://doi.org/10.3390/plants11243514
APA StyleRuksiriwanich, W., Khantham, C., Muangsanguan, A., Phimolsiripol, Y., Barba, F. J., Sringarm, K., Rachtanapun, P., Jantanasakulwong, K., Jantrawut, P., Chittasupho, C., Chutoprapat, R., Boonpisuttinant, K., & Sommano, S. R. (2022). Guava (Psidium guajava L.) Leaf Extract as Bioactive Substances for Anti-Androgen and Antioxidant Activities. Plants, 11(24), 3514. https://doi.org/10.3390/plants11243514